Survival of cyanobacteria and mitigation of Fe(II) toxicity effects in a silica-rich Archean ocean
Carolin L. Dreher, Olaf A. Cirpka, Manuel Schad, Kurt O. Konhauser, Andreas Kappler

TL;DR
This study shows how cyanobacteria survived in ancient oceans by reducing the harmful effects of oxygen radicals using silica and day-night cycles.
Contribution
The study reveals that silica and diel light cycles mitigated ROS toxicity, enabling cyanobacterial growth in early Earth's oceans.
Findings
High SiO2(aq) suppressed ROS formation and promoted cyanobacterial growth and O2 production.
Diel light cycles reduced ROS formation compared to continuous illumination.
Modeling suggests oxygenated surface waters at relevant upwelling rates in Archean oceans.
Abstract
Banded iron formations (BIF) were deposited abundantly between 2.7-2.4 Ga from iron- and silica-rich oceans, with cyanobacterial oxygen (O2) as a possible oxidant for Fe(II)(aq) oxidation and Fe(III) oxyhydroxide precipitation. However, toxic reactive oxygen species (ROS) from Fe(II)/O2 interactions might have inhibited cyanobacterial growth, contributing to the delay between cyanobacterial evolution (>3.0 Ga) and the Great Oxidation Event (2.5 Ga). Here, we explored the impact of Fe(II)(aq) and SiO2(aq) on Synechococcus sp. PCC 7002. High Fe(II)(aq) ( > 500 µM) increased ROS formation, but elevated SiO2(aq) (2200 µM) suppressed ROS formation, promoting growth and O2 production. Diel light cycles further reduced ROS formation compared to continuous illumination. Modelling O2 distribution based on experimental rates revealed oxygenated surface waters at relevant upwelling rates.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
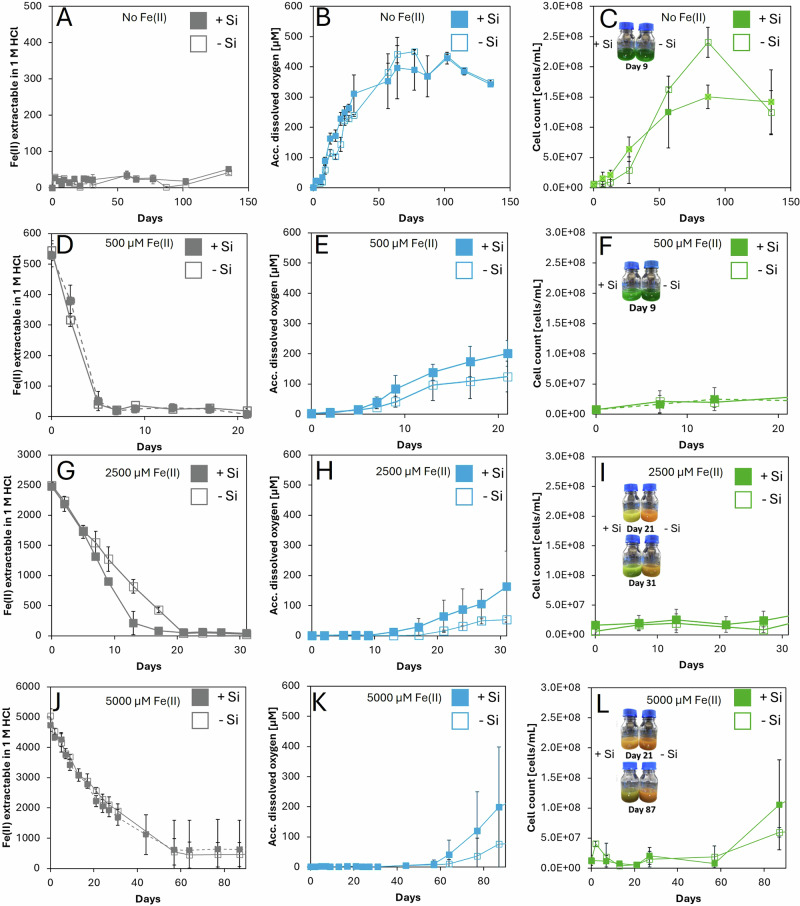 Figure 1
Figure 1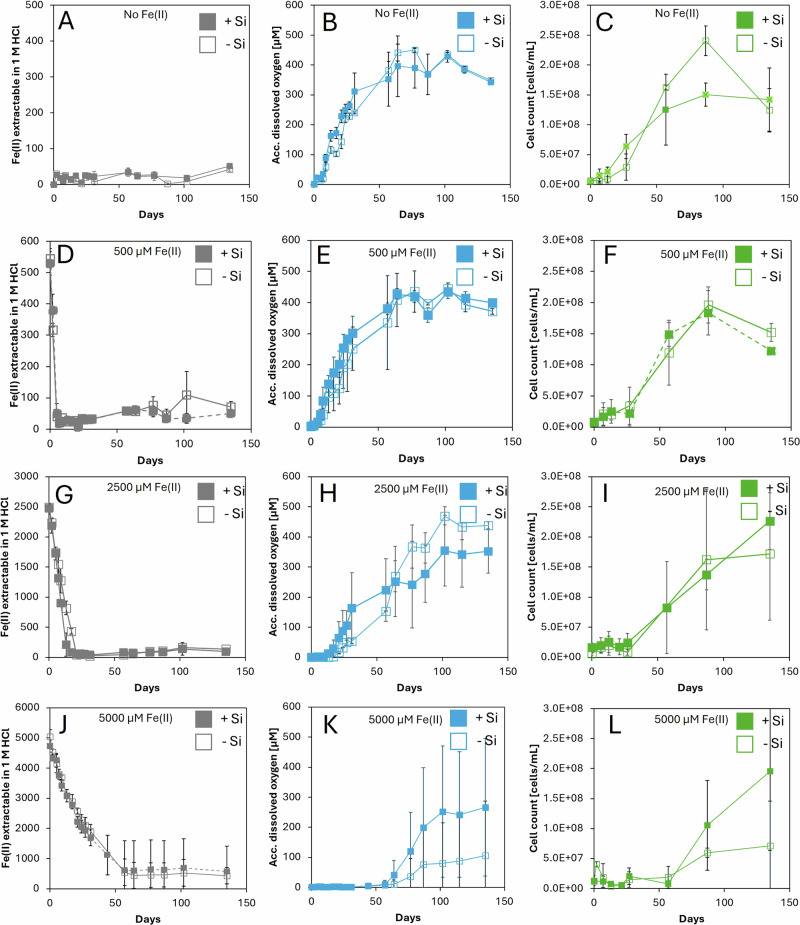 Figure 2
Figure 2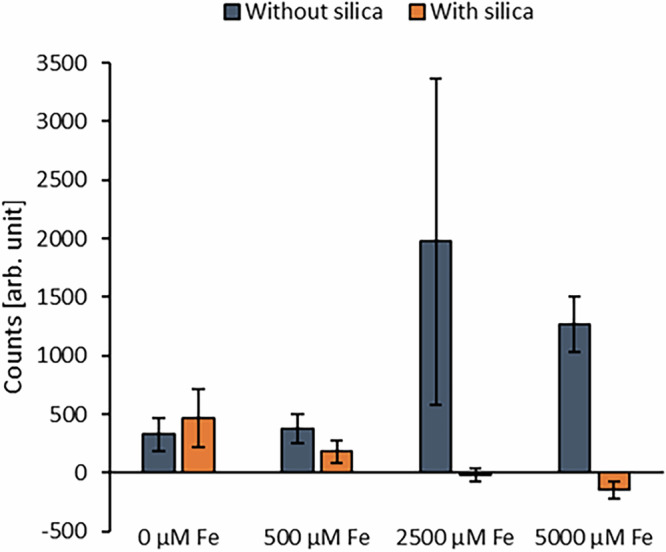 Figure 3
Figure 3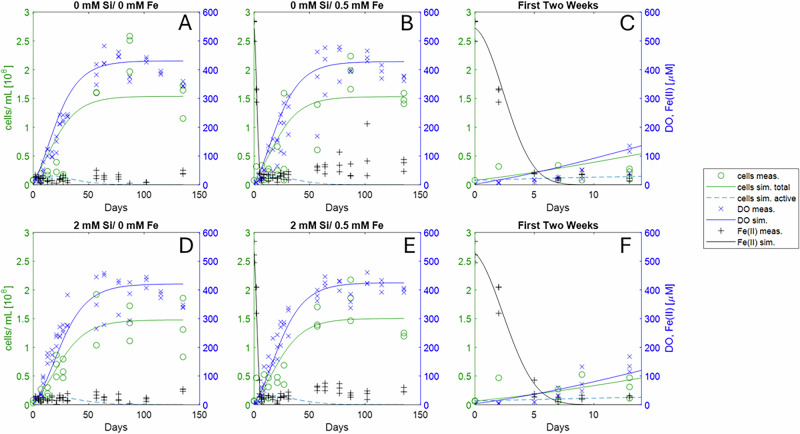 Figure 4
Figure 4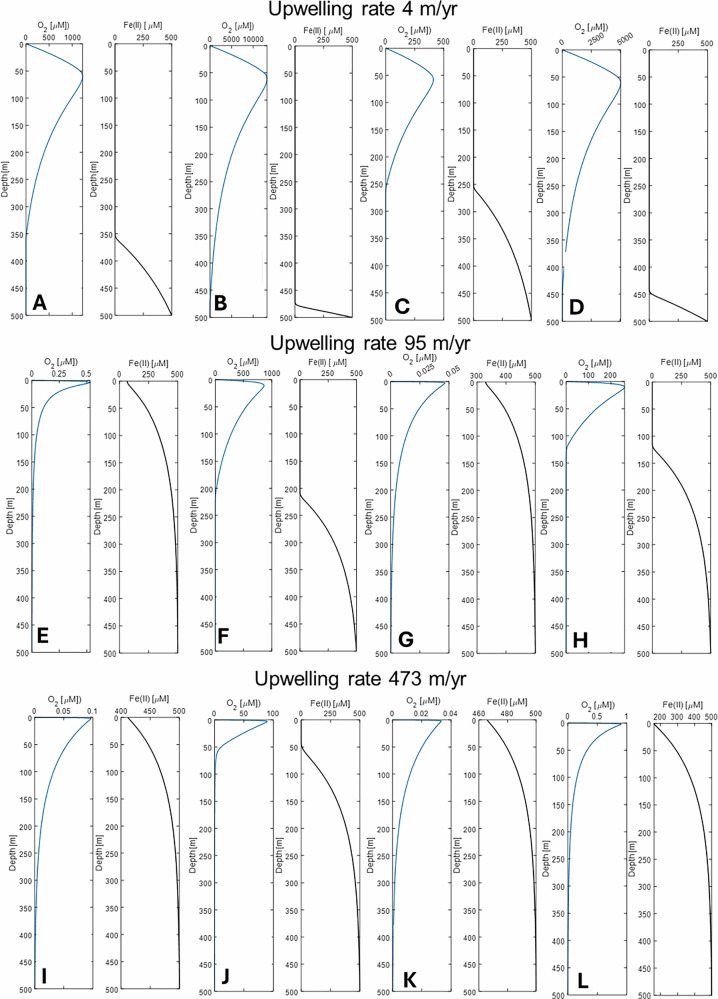 Figure 5
Figure 5- —https://doi.org/10.13039/501100001659Deutsche Forschungsgemeinschaft (German Research Foundation)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMarine and coastal ecosystems · Microbial Fuel Cells and Bioremediation · Geochemistry and Elemental Analysis
Introduction
Sometime between the evolution of oxygen (O_2_)-producing cyanobacteria, possibly as early as 3.4 billion years ago^1,2^ and the permanent rise of atmospheric oxygen during the Great Oxidation Event (GOE) at 2.5–2.3 Ga^3^, free O_2_ began to accumulate in seawater. This accumulation likely began in the form of localized and/or transient oxygen oases, with estimated O_2_ concentrations ranging from 5 to 100 µM^4–7^, before episodically expanding across broader regions of the continental shelf^8–10^. The reasons for the protracted delay between the emergence of cyanobacteria and the GOE remain unresolved but may, at least in part, reflect environmental constraints that inhibited cyanobacterial growth and dispersal^11,12^.
One of the most distinctive sedimentary archives of the Archean, BIFs, may hold clues to these limiting environmental conditions (see^13^ for review). These chemical sediments consist predominantly of iron- (15–40 wt.% Fe) and silica-rich (40–60 wt.% SiO_2) layers that precipitated from seawater following the oxidation of Fe(II)(aq)_, with estimated concentrations ranging from 0.03 to 0.5 mM^14,15^, and possibly exceeding 1.0 mM in some settings^16,17^. Prior to the emergence of silica-precipitating microorganisms (e.g., diatoms), silica inputs from chemical weathering of early continental crust and hydrothermal vents led to elevated dissolved silica concentrations in the seawater. These concentrations were constrained chiefly by saturation with respect to cristobalite or amorphous silica, on the order of 0.67-2.2 mM^18,19^. Recent studies further suggest that under these ‘high silica’ concentrations, the primary precipitate may have been a gel-like composite of ferric oxyhydroxide and silica^20–22^.
While the deposition of the oldest BIF was probably linked to anoxygenic phototrophic Fe(II)-oxidizers^23,24^, photochemical Fe(II) oxidation^25,26^ or abiotic deposition processes such as chemical greenalite deposition^27,28^, the proliferation of cyanobacteria would have accelerated Fe(II) oxidation and thus BIF deposition^13,29^. Cyanobacteria fix and reduce CO_2_ using water as electron donor to form biomass, also yielding O_2_ as by-product (reaction 1). As such, they were key players in the oxygenation of Earth^30,31^.
\documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$6{{\rm{C}}}{{{\rm{O}}}}_{2}+12{{{\rm{H}}}}_{2}{{\rm{O}}}{\to }^{h\nu }{{{\rm{C}}}}_{6}{{{\rm{H}}}}_{12}{{{\rm{O}}}}_{6}+6{{{\rm{O}}}}_{2}+6{\,{{\rm{H}}}}_{2}{{\rm{O}}}$$\end{document}In early Precambrian oceans, the O_2_ may have reacted with Fe(II)(aq) to form Fe(III)(aq), which hydrolyzed into Fe(III) oxyhydroxide minerals. The reaction between dissolved Fe(II)(aq) and O_2_ may also yield reactive oxygen species (ROS), that is, highly reactive oxygen-containing molecules (i.e., the Fenton reaction^32^):
\documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$${{{\rm{O}}}}_{2}+{{{\rm{Fe}}}}^{2+}\to {{{\rm{Fe}}}}^{3+}+{{{\rm{O}}}}_{2}^{-}{{{\bullet }}}$$\end{document} \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$${{{\rm{O}}}}_{2}^{-}{{{\bullet }}}+{{{\rm{Fe}}}}^{2+}+2{{{{\rm{H}}}}^{+}\to {{\rm{Fe}}}}^{3+}+{{{\rm{H}}}}_{2}{{{\rm{O}}}}_{2}$$\end{document} \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$${{{\rm{Fe}}}}^{2+}+\,{{{\rm{H}}}}_{2}{{{\rm{O}}}}_{2}\,\to \,{{{\rm{Fe}}}}^{3+}+{{\rm{O}}}{{{\rm{H}}}}^{-}+{{\rm{OH}}}{{{\bullet }}}\,$$\end{document}Briefly, superoxide ( \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$${{{\rm{O}}}}_{2}^{-}$$\end{document} •), generated by the reaction of O_2_ and Fe(II)(aq) (reaction 2; note we provide actual chemical species here versus the generic Fe(II)), further reacts with Fe(II)aq to produce Fe(III)(aq) and hydrogen peroxide (H_2_O_2_) (reaction 3). The H_2_O_2_ then oxidizes the remaining Fe(II)(aq) to Fe(III)(aq), along with the formation of hydroxide anions ( \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$${{{\rm{OH}}}}^{-}$$\end{document} ) and hydroxyl radicals (OH•) (reaction 4). While the Fe^3+^ drives the formation of Fe(III) minerals, the generated radicals can undergo further reactions, forming additional ROS like ozone (O_3_) and hydroperoxyl radicals (OOH•) (for a review see ref. ^33^).
ROS are harmful to cells; extracellular ROS can damage cell membranes through oxidative stress^34^, while intracellular ROS can oxidize critical biomolecules such as RNA, DNA, proteins, and lipids^35^. Therefore, ROS formation in Archean oceans may have profoundly influenced microbial activity and evolution. Previously, Swanner and colleagues investigated ROS production under early ocean analog conditions using phosphate-buffered saline (PBS) media and varying concentrations of Fe(II)(aq) (0.1 and 1 mM) in the presence of Synechococcus sp. PCC 7002 at a density of 2×10^7^ cells/mL^12^. Under continuous light, they found a 4-fold increase in intracellular ROS levels in the presence of 1 mM Fe(II)(aq) compared to 0.1 mM Fe(II)(aq), suggesting potential toxicity to cyanobacteria at elevated Fe(II)(aq) concentrations. Additionally, in incubation experiments of cyanobacteria under initially anoxic conditions (in the presence of up to 300 µM Fe(II)aq) reduced autofluorescence of the cyanobacteria was reported, accompanied by significantly lower actinic yield, and decreased growth rates at Fe(II)(aq) concentrations >180 µM.
In a follow-up study, Swanner and colleagues conducted Fe(II) oxidation experiments with Synechococcus sp. PCC 7002 under continuous illumination and a range of Fe(II)(aq) concentrations (7.5, 29, 577, and 4805 µM)^11^. At lower Fe(II)aq concentrations, cultures changed from colorless to green, indicating a high density of cyanobacterial cells. By contrast, higher Fe(II)(aq) concentrations produced an orange coloration, consistent with the formation of Fe(III) oxyhydroxide minerals. These observations were supported by direct cell counts, which confirmed reduced growth at higher Fe(II)(aq) concentrations, again suggesting toxic effects. Microscopic analyses further revealed that cells grown under high Fe(II)(aq) concentrations were smaller and exhibited reduced pigment content, as indicated by lower carotenoid and chlorophyll concentrations inferred from optical density measurements. Although Fe(II) oxidation rates increased with rising Fe(II)(aq) concentrations (from 13 µM/day at 29 µM to 475 µM/day at 4805 µM) the rate of O_2_ production in the liquid phase decreased from 120 µM/day to 47 µM/day, indicating diminished cellular activity. Notably, O_2_ was already detectable during Fe(II) oxidation at 29 and 475 µM Fe(II)(aq) but absent at 4805 µM Fe(II)(aq). Taken together, these findings suggest that elevated Fe(II)(aq) concentrations in Archean seawater could have imposed oxidative stress, limiting cyanobacterial productivity and expansion. This could potentially explain the lag between the evolution of cyanobacteria and the GOE^11,12^.
However, the potential mitigating impact of high SiO_2(aq)_ concentrations on Fe(II)(aq) toxicity has not been previously considered. Our working hypothesis is that silica binds Fe(II)aq, forming Fe-Si-aggregates that slow Fe(II) oxidation and consequently inhibit ROS formation. Previous studies have shown that reduced Fe(II) oxidation rates lead to lower ROS formation^36^. Consistent with this, Dreher and colleagues^37^ showed, using SEM/EDS, that Fe-Si aggregates precipitate under similar experimental conditions, further supporting our hypothesis. To test this hypothesis, we experimentally examined the combined effects of Fe(II)(aq) and SiO_2(aq)_ on cyanobacterial O_2_ production, Fe(II) oxidation, and potential ROS-related toxicity. Specifically, we incubated Synechococcus sp. PCC 7002 under alternating day-night-cycles (16 h light; 8 h dark) in artificial seawater medium, in the absence of SiO_2(aq)_ (0 µM, which we refer to as ‘no-silica’) and in the presence of SiO_2(aq)_ (2200 µM, which we refer to as ‘high-silica’) and varying initial Fe(II)(aq) concentrations (0, 500, 2500, 5000 µM). Over the course of the experiments, we monitored O_2_ production, Fe(II)aq and SiO_2aq_ concentrations, total iron (Fe_(tot)), cell numbers, and ROS production to assess the interdependent effects of Fe(II)(aq)_ and SiO_2(aq)_ on cyanobacterial physiology and oxidative stress.
Results
Effects of different silica concentrations on Fe(II) oxidation by cyanobacterial O2 under alternating day-night-cycles
We conducted Fe(II) oxidation experiments, under initially anoxic conditions, using Fe(II)(aq) concentrations ranging from 0.5 to 5 mM. Following inoculation with the cyanobacterial strain Synechococcus sp. PCC 7002, we monitored Fe(II), Fe(tot), cyanobacterially produced O_2_, dissolved Si, and cell counts over time (for sterile controls see Fig. S3). The results are plotted in Fig. 1 on the respective timescale of the active Fe(II) oxidation, as this sets the focus on the period when there was still Fe(II)(aq) in the system, which is the most relevant for determining toxicity effects and the latter modeling. In setups without amendment of additional Fe(II)(aq), all cultures exhibited a color change from clear to green within two days, regardless of SiO_2(aq)_ concentration (Fig. 1C), indicating cyanobacterial growth. In the ‘high silica’ setups compared to the ‘no silica’ setups, cell growth was initially more rapid but reached a slightly lower maximum cell density (Fig. 1C, Table 1). Under both experimental conditions, O_2_ concentrations peaked at 500 µM after 60 days (Fig. 1B). In the setups without additional Fe(II) amendment, the Fe(II)(aq) concentration remained relatively stable throughout the experiments, averaging approximately 50 µM in with and without both silica setups (Fig. 1A), a level attributable to the Fe(II)(aq) added as nutrient in the medium. In the ‘high silica’ setups, the initial SiO_2(aq)_ was ca. 1600 µM (Fig. S1A). However, after 30 days, solid silica precipitation started, ultimately reducing the SiO_2(aq)_ concentration to several hundred µM by the end of the experiment.Fig. 1. Geochemical data of Fe(II) oxidation experiments containing Synechococcus sp. PCC 7002.Panels A–C show the results of setups without amendment of Fe(II)(aq). Panels D–F show the results of setups with amendment of 500 µM Fe(II)(aq), panels G–I with 2500 µM Fe(II)(aq), and panels J–L with 5000 µM Fe(II)(aq). Filled symbols indicate setups with 2200 µM silica (‘high silica’), while empty symbols indicate setups without silica (‘no silica’). Fe(II) (gray), accumulated oxygen (blue), cell numbers (green) are plotted as average values from triplicates with the standard deviations shown as error bars. Please note that the time scale (x-axis) varies for the different plots because of the differences in the duration of active Fe(II) oxidation.Table 1. Geochemical data of the Fe(II) oxidation (FeOx) experimentsSilica [µM]Fe(II) [µM]FeOx rate µM/dayLength of FeOx [days]Maximum oxygen conc. [µM]Average maximum cell count [cells/mL]Green liquid observed after x days00**––450 after 60 days2.5 × 10^8^ after 87 days222000––**450 after 60 days1.7 × 10^8^ after 87 days20500101 ± 85450 after 60 days2.0 × 10^8^ after 87 days2220050096 ± 105450 after 60 days2.0 × 10^8^ after 87 days202500129 ± 521350 after 100 days2.3 × 10^8^ after 140 days-22002500174 ± 1117490 after 100 days1.7 × 10^8^ after 87 days310500077 ± 658290 after 140 days0.6 × 10^8^ after 140 days–2200500072 ± 2058100 after 140 days2 × 10^8^ after 140 days87The FeOx rate [µM/day] of the triplicates with the standard deviation, the length of active FeOx [days], the average maximum accumulated oxygen concentration [µM], the average maximum cell count [cells/mL,] and the amount of days after which the liquid turned green of the setups containing 0 µM added silica (‘no silica’) and 2200 µM added silica (‘high silica’) with amendment of 0–5000 µM Fe(II)(aq).
In experiments with amendment of 500 µM of Fe(II)(aq), all bottles turned green within two days, regardless of the SiO_2(aq)_ concentration (Fig. 1F). Growth curves were similar across setups, with no discernible effect of SiO_2(aq)_ (Fig. 1F, Fig. 2,Table 1), and maximum cell densities were reached after 87 days (Fig. 2). In both ‘no silica’ and ‘high silica’ setups, O_2_ accumulated in the headspace following complete Fe(II) oxidation, peaking at 450 µM after 60 days (Fig. 1E).Fig. 2. Geochemical data of Fe(II) oxidation experiments containing Synechococcus sp. PCC 7002 over the entire course of the experiment.Panels A–C show the results of setups without amendment of Fe(II)(aq). Panels D–F show the results of setups with amendment of 500 µM Fe(II)(aq), panels G–I with 2500 µM Fe(II)(aq), and panels J–L with 5000 µM Fe(II)(aq). Filled symbols indicate setups with 2200 µM silica (‘high silica’), while empty symbols indicate setups without silica (‘no silica’). Fe(II) (gray), accumulated oxygen (blue), cell numbers (green) are plotted as average values from triplicates with the standard deviations shown as error bars.
Within the first five days of the experiment, 80–90% of the added 500 µM Fe(II)(aq) was oxidized in both ‘no silica’ and ‘high silica’ setups (Fig. 1D). With and without silica, the Fe(II) oxidation rates were similar within the calculated error (Table 1). In the ‘no silica’ and ‘high silica’ setups, Fe(II) oxidation rates were similar at about 100 µM/day (Fig. 1D; Table 1). In the ‘high silica’ setups (Fig. 1B), silica precipitation began after 60 days, reducing SiO_2(aq)_ to approximately 100 µM.
In experiments amended with 2500 µM of Fe(II)(aq), the bottles turned orange in the absence of added silica (Fig. 1I), contrasting with experiments with 0 and 500 µM added Fe(II)(aq). However, in the presence of silica, the 2500 µM Fe(II) bottles turned yellow by day 21 and green by day 31. The orange coloration in the ‘no silica’ setup suggests abiotic Fe(II) oxidation and the formation of Fe(III) oxyhydroxide minerals, likely caused by cyanobacterially produced O_2,_ but with limited cell growth and metabolic activity. By contrast, the green coloration in the ‘high silica’ setups indicates extensive cyanobacterial proliferation. It is important to interpret these color changes cautiously, as higher Fe(II) concentrations can intensify the orange hue regardless of biological activity, potentially confounding visual comparisons between setups.
Cell counts revealed that in the ‘no silica’ setups, cyanobacterial growth was delayed, with cell numbers increasing only after 25 days (Fig. 1I). Conversely, cell proliferation in the ‘high silica’ setups commenced immediately after inoculation and ultimately reached similar densities by day 150 (Fig. 1I). Silica availability had a marked influence on O_2_ accumulation. In the ‘no silica’ setups (Fig. 1H), O_2_ accumulated only after day 20 (ca. 50 µM by day 31). In comparison, in the ‘high silica’ setups (Fig. 1H) O_2_ accumulated as early as day 10, reaching 150 µM by day 31.
In the ‘no silica’ setups, the Fe(II) oxidation rate was lower compared to the ‘high silica’ setups (Fig. 1G; Table 1). Initial SiO_2(aq)_ concentration in the ‘high silica’ setups were 1500 µM (Fig. S1C, Fig. 1C) and decreased to 700 µM within the first 20 days. By day 90, variability emerged among the triplicate bottles: one replicate maintained a stable SiO_2(aq)_ concentration of 700 µM, whereas the other two showed continued declines to 300 µM and 100 µM, respectively.
In experiments amended with 5000 µM of Fe(II)(aq), the ‘no silica’ setups remained orange until day 87 (Fig. 1L), indicating Fe(III) oxyhydroxide formation and limited cyanobacterial growth. By contrast, the ‘high silica’ setups turned yellow by day 21 and green by day 87, reflecting more substantial cyanobacterial growth. Consistent with these color changes, total cell numbers were higher in the ‘high silica’ setups. In the ‘no silica’ setups, cell growth was delayed until Fe(II) oxidation was complete, after which cell densities increased to 5 × 10⁷ cells/mL (Fig. 1L). In one replicate, cyanobacteria eventually recovered, reaching 1.5 × 10⁸ cells/mL by day 140 Fig. 2. In the ‘high silica’ setups, final cell densities varied among the three replicates, reaching 2.5 × 10⁸, 1.5 × 10⁸, and 5 × 10⁷ cells/mL, respectively (Fig. 1L).
O_2_ accumulated in the ‘no silica’ setups after 60 days, reaching ca. 80 µM by day 87 (Fig. 1K), although replicate variability was high: one bottle showed no detectable O_2_, while another reached 300 µM O_2_ by day 140. In contrast, O_2_ concentrations in the ‘high silica’ setup increased after 57 days (Fig. 1K) and reached 200 µM by day 87. Two out of three replicates further reached 400 µM O_2_ by day 100, whereas in the third, in which only 60% of the Fe(II) was oxidized, showed no detectable O_2_.
The Fe(II) oxidation rates were similar between the ‘no silica’ and ‘high silica’ setups (Fig. 1J; Table 1), but were notably lower than the rates observed in experiments with 2500 µM Fe(II)(aq). In the ‘high silica’ setups, the initial SiO_2(aq)_ concentration of 1500 µM declined to 500 µM by day 31 (Fig. S1D), after which it either stabilized or continued to precipitate, reaching final concentrations between 100 and 200 µM.
Improvement of long-term cyanobacterial cell viability by silica
In addition to experiments focusing on cyanobacterial growth and activity i.e., cell numbers, O_2_ production, resulting Fe(II) oxidation and dissolved silica (see Fig. S1), we explored the role of silica in promoting long-term cell viability experiments by observing potential mitigation effects of the harmful effects of ROS generated in the presence of both Fe(II)(aq) and O_2_. In all setups, the presence of Fe(II)(aq) delayed both cell growth and O_2_ production (Fig. 2). In the 500 µM Fe(II)(aq) (Fig. 2 E), ‘high silica’ setups initially showed higher O_2_ accumulation during the first 70 days, with both setups converging at 350 µM by day 60. In the 2500 µM Fe(II) experiments (Fig. 2G–I), the presence of SiO_2(aq)_ led to significantly enhanced cyanobacterial activity: by day 150, O_2_ concentrations reached 500 µM and cell densities ca. 2.4 × 10^8^ cells/mL, compared to 350 µM O_2_ and 1.8 × 10^8^ cells/mL in the absence of SiO_2(aq). The beneficial effect of silica was even more pronounced in the 5000 µM Fe(II) setups. After 250 days, O_2 accumulation in the ‘high silica’ setups reached 250 µM, with cell densities of 2 × 10^8^ cells/mL, while the ‘no silica’ setups showed limited cell growth, reaching only 5 × 10^7^ cells/mL and 100 µM O_2_.
Formation of ROS in incubations of strain PCC 7002 with Fe(II)
Based on the lower cell growth and Fe(II) oxidation rates at higher Fe(II)(aq) concentrations (500, 2500–5000 µM), we hypothesized that cyanobacterially produced O_2_ reacted with Fe(II)(aq) to generate ROS, potentially leading to cellular stress or toxicity. However, our results also indicated that SiO_2(aq)_ mitigates these negative effects, as evidenced by higher cell densities, earlier O_2_ accumulation, and increased Fe(II) oxidation rates in silica-amended setups. To further evaluate the impact of varying Fe(II)(aq) and SiO_2(aq)_ concentrations on ROS production, we subsequently constructed a series of experiments using Synechococcus sp. PCC 7002 cell suspensions (5 × 10^9^ cells/mL) exposed to different Fe(II)(aq) concentrations (0, 500, 2500 or 5000 µM), both in the absence and presence of 2200 µM SiO_2(aq)_.
Our results (Fig. 3, Fig. S2, Table S1) showed that in the absence of Fe(II)(aq), ROS fluorescence signals were generally very low, yet slightly elevated in the presence of SiO_2(aq)_ compared to setups without SiO_2(aq)_ (ca. 500 a.u. and 400 a.u., respectively) (Fig. 3). After the amendment of 500 µM Fe(II)(aq), the ROS fluorescence signal, with and without silica, was similar to the setups without Fe(II), suggesting no significant ROS formation at this Fe(II)(aq) concentration. However, in the presence of 2500 µM of Fe(II)(aq), we observed a pronounced increase in ROS fluorescence in the absence of silica (2000 a.u.), while the signal remained near baseline in silica-amended setups. A similar trend was observed at 5000 µM Fe(II)(aq), where ROS levels reached 1300 a.u. without silica, but remained negligible in the presence of silica (Fig. 3). These results suggest that high Fe(II) concentrations promote ROS generation, and that dissolved silica effectively suppresses ROS formation under these conditions.Fig. 3. Impact of silica on reactive oxygen species (ROS) formation at different iron concentrations.Average ROS fluorescence signals relative to an anoxic, abiotic control containing 5000 µM Fe(II)(aq) with and without 2200 µM SiO_2(aq). The experimental setups contained strain PCC 7002 cells, either 0, 500, 2500, or 5000 µM Fe(II)(aq)_ in the absence (blue) and presence of 2200 µM SiO_2(aq)_ (orange). Please note that the normalization of all measurements to the abiotic control can result in negative values. Error bars show the standard deviation calculated from quadruplicate setups.
Numerical modeling of the experimental O2 data
In order to simulate the total O_2_ production, we incorporated our raw laboratory data into a numerical model to simulate the total O_2_ production. To account for the observed plateau of O_2_ concentration at approximately 450 µM (Fig. 4A–E), we differentiated between ‘active’ cells (O_2_-producing; green solid line) and ‘inactive’ cells (non–O_2_-producing; calculated as the difference between total cells and measured active cells). The ‘inactive’ group included both dead cells and those estimated to have received insufficient light due to increased cell density during the experiment. While cultures were initially colorless, they gradually developed an intense dark green coloration, increasing turbidity, and thereby reducing the average light intensity available to each bacterial cell.Fig. 4. Modeled lab data of the experimental setups containing 500 µM iron without silica (top) and with 2200 µM silica (bottom).Panel A shows the modeled data of setups without amendment of silica or Fe(II)(aq), B shows data of setups without silica amendment but with 500 µM Fe(II)(aq). D Shows setups with 2200 µM silica without Fe(II)(aq), E shows setups with 2200 µM silica and 500 µM Fe(II)(aq). Panels C and F show the data zoomed into the first 15 days of panels B and E, respectively. Green (‘cell sim.total’), blue (‘DO sim.’) and black (‘Fe(II) sim.’) lines are the modeled total cell densities and oxygen concentrations, respectively, the green circles (‘cells meas.’), the blue crosses (‘DO meas.’) and the black crosses (‘FeII) meas.’) show the associated raw data points. The bright blue dashed line (‘cells sim. active’) shows the calculated concentration of active cells. The black solid lines (‘Fe(II) sim.’) shows the modeled Fe(II) curve.
We also calculated the total O_2_ production in our experiments, accounting for both the dissolved O_2_ in the liquid and free O_2_ present in the headspace. Based on the equilibrium relationship between the aqueous solution and headspace (see Supplementary Information, formula 9), we estimate that O_2_ production (including the headspace) is approximately 42 times greater than measurements based solely on dissolved O_2_. This correction is crucial for accurately quantifying true O_2_ production in our experimental setup.
Oxygen and Fe(II) distribution modeled for the ancient water column
To scale our laboratory-derived O_2_ production to early ocean conditions, we calculated the oxygen production per cell (r_photo_) from the first two days of the experiment, a period when no detectable O_2_ accumulated in the liquid phase and no measurable cell growth occurred (see “Methods” for details). We calculated the oxygen and Fe(II)aq distribution for three upwelling rates: 4, 95, and 473 m/yr. 4 m/yr represents the average upwelling velocity of modern oceans, while 95 m/yr and 473 m/yr are modeled average and enhanced oceanic upwelling velocities in the mid-Cretaceous North Atlantic Ocean, respectively^38^. Using an upwelling rate of 4 m yr⁻¹ (Fig. 5A–D), our model predicted O_2_ concentrations of at least 400 µM between 50 and 100 m depth in the water column, with the potential for substantially higher values. Given that the maximum O_2_ solubility in an open systems at room temperature and atmospheric pressure is ~300 µM^39,40^ (higher in closed systems), these results indicate that such upwelling rates would saturate and fully oxidize the photic zone, leading to complete oxidation of Fe(II)(aq) and excess O_2_ outgassing to the atmosphere.Fig. 51D steady-state model of oxygen distribution in the early ocean.Modeled oxygen (left, blue) and Fe(II) (right, gray) concentrations in the ancient ocean water column for an upwelling velocity of 4 (panels A–D), 95 (panels E–H) and 473 m/yr (panels I–L). Panels A, C, E, G, I, and K are calculated for hypothesized 10^5^ cells/mL in the early ocean, B, D, F, H, J, L for 10^6^ cells/mL. Panels A, B, E, F, I, and J are modeled based on the photosynthetic rate (r_photo_) calculated from the lowest cell count (6.5 × 10^5^ cells/mL) we measured in our experiments at d0; C, D, G, H, K, and L are based on the highest cell count (1.6 × 10^7^ cells/mL) we measured at d0.
By increasing the upwelling rate to 95 m yr⁻¹, our model predicts that, at a cyanobacterial abundance of 10⁵ cells mL⁻¹ (Fig. 5E, G), Fe(II)(aq) would have reached the ocean surface. Oxygen concentrations at the surface would have reached between 0.04 and 0.4 µM maximum concentration. At a higher theoretical abundance of 10⁶ cells mL⁻¹ (Fig. 5F, H), however, the photic zone remained Fe(II)aq-free down to depths of ~200 m or ~125 m, respectively. In these high-cell-density scenarios, O_2_ accumulated to much higher concentrations within the upper 0–50 m of the water column, with modeled values reaching (200 µM) or exceeding (800 µM) maximum O_2_ saturation.
At an upwelling rate of 473 m/yr, Fe(II)(aq) reached the water surface in most scenarios (Fig. 5I, K, L). Under these conditions, O_2_ accumulation at the top of the water column was limited, ranging from 0.03 µM (Fig. 5K) to 0.8 µM (Fig. 5L). In scenario J, however, Fe(II)(aq) was absent only from the upper 50 m, allowing O_2_ to accumulate to 80 µM near the water surface. Overall, the results highlight the strong control of upwelling rate on O_2_ and Fe(II)(aq) dynamics. At low upwelling rates (4 m/yr), the photic zone was fully oxidized, whereas at 96 m/yr, O_2_ accumulated only near the surface. In contrast, most scenarios with rapid upwelling (473 m/yr) resulted in Fe(II)(aq) reaching the surface, suppressing O_2_ buildup and preventing full saturation.
Discussion
ROS formation and the mechanistic function of Si for ROS prevention
Our results demonstrate a clear relationship between Fe(II)(aq) concentrations and the extent of ROS formation in the absence of SiO_2(aq), generally supporting the findings of Swanner and colleagues^11,12^. Differences between our study and theirs can likely be attributed to variations in experimental setups, such as constant illumination versus day-night-cycles, and static versus agitated conditions (see above). Additionally, minor methodological variations, including the use of phosphate-buffered saline (PBS)^12^ versus TRIS buffer in our study, as well as differences in fluorometric detection systems, may have contributed to the observed differences. Nevertheless, despite the variations, our study demonstrates a protective effect of SiO_2(aq) against ROS-induced stress in cyanobacteria under conditions relevant to early Precambrian oceans. Even at high Fe(II)(aq) concentrations (2500 and 5000 µM), ROS levels remained below those observed in anoxic, abiotic controls when SiO_2(aq)_ was present (Fig. 3). Furthermore, we measured higher O_2_ immediately following Fe(II) oxidation in the presence of SiO_2(aq)_ at all Fe(II)(aq) concentrations (Fig. 1), suggesting enhanced cyanobacterial activity. Notably, in the 5000 µM Fe(II)(aq) experiments, SiO_2(aq)_ enabled long-term recovery and sustained growth of Synechococcus sp. PCC 7002, with cell densities reaching 2×10^8^ cells/mL after 150 days, compared with only 5 × 10^7^ cells/mL in the absence of SiO_2(aq)_.
The protective effect of SiO_2(aq)_ against ROS formation can be attributed to the ability of Si to bind Fe(II)(aq), forming Fe(II)-Si complexes^28,37^, that reduce Fe(II) reactivity with O_2_, thereby inhibiting or reducing Fenton-type reactions and lowering ROS production. The formation of amorphous Fe(II)-Si aggregates, as hypothesized in our experiments, correlates well with previous studies simulating Archean ocean conditions (1 mM SiO_2aq_, 1.1 mM Fe(II)(aq), pH of 6.5–7.5^41,42^;). This pH range is consistent with both our experimental conditions (pH 7) and estimates for circumneutral pH in early Precambrian oceans^43,44^. While Fe(II) within these aggregates remained susceptible to oxidation by O_2_, the resulting products likely precipitated as Fe(III)-Si-aggregates on the seafloor rather than as Fe(II)-Si phases (^37^).
Day-night-cycles reduce ROS formation compared to continuous light and static conditions
Our results showed that Fe(II)(aq) oxidation rates under day-night-cycle incubations were higher than those reported in a previous study using the same setup—including the same cyanobacterial strain (Synechococcus sp. PCC 7002), initial cell densities (10^6^–10^7^ cells/mL), similar Fe(II)(aq) and SiO_2(aq)_ concentrations, light intensities (300–500 lux), and identical glassware—but conducted under continuous illumination (^37^) (Table 2). In these continuous light experiments, the presence of 2200 µM silica resulted in remarkably higher cell counts and O_2_ concentrations compared to the silica-free equivalents^37^. The difference in Fe(II) oxidation rates between day-night cycles and continuous illumination was especially pronounced at the lowest Fe(II)(aq) concentration (500 µM) in the absence of SiO_2(aq), where Fe(II) oxidation rates reached 113 µM/day under day-night cycles versus only 32 µM/day under continuous light. The only exception occurred in the setup containing 2500 µM Fe(II)(aq)_ and 2200 µM SiO_2(aq)_, where Fe(II) oxidation rates of 129 and 168 µM/day were observed in day-night-cycles and under continuous illumination, respectively.Table 2. Fe(II) oxidation rates in day-night-cycles (left column, data from this paper); Fe(II) oxidation rates in continuous light right column, data from Dreher and colleagues^37^day-night-cycles**continuous lightSilica [µM]Fe(II) [µM]FeOx rate µM/dayFeOx rate µM/day0500101 ± 7.732 ± 1220050096 ± 9.730 ± 102500129 ± 5.1168 ± 222002500174 ± 10.8147 ± 150500077 ± 6.463 ± 102200500072 ± 20.240 ± 2
The effects of continuous light exposure on phototrophic organisms have been well documented^45^, with prolonged and continuous illumination linked to increased ROS formation and cellular damage. In the context of Fe(II) oxidation, Hegler and colleagues examined the influence of light intensity and wavelength under continuous illumination, reporting maximum Fe(II) oxidation rates of 4.5 mmol/L/day at 800 lux, with overall Fe(II) oxidation rates increasing with light intensity^46^. More recently, Nikeleit and colleagues demonstrated that day-night-cycle exposure enhanced Fe(II) oxidation rates in the anoxygenic phototroph Chlorobium ferrooxidans strain KoFox, suggesting a physiological benefit of diel variation^47^. Furthermore, day-night-cycles have been shown to stimulate biomass production, protein synthesis, and coenzyme Q10 production in anoxygenic phototrophic bacteria such as Erythrobacter sp. NAP1 and Rhodobacter sphaeroides^48,49^.
During the light periods of day-night-cycles, cyanobacteria produce O_2_ most efficiently at light intensities between 30 and 50 µmol m² s^-1^ ^50–52^. In dark periods, their metabolic activity is severely limited, with neither O_2_ nor glucose produced; energy is instead derived from the degradation of internally stored glycogen^53^. Muhetaer and colleagues^52^ showed that the effects of continuous light depend on the strain’s specific circadian rhythm. For example, M. aeruginosa exhibited increased H_2_O_2_/ OD_730_ ratios and partial cell death after 8 days of continuous light (e.g., H_2_O_2_/ OD_730_ of 400 at 300 µmol m^–2^ s^–1^) compared to two days (H_2_O_2_/ OD_730_ of 230 at 300 µmol m^–2^ s^–1^), whereas the strain P. galeata showed opposite trends, suggesting significant inter-strain variability in stress responses to prolonged illumination^52^. It is thus plausible that day-night-cycles applied in our experiments were more ideal for the cyanobacteria’s circadian rhythm than continuous light exposure. This may explain the enhanced Fe(II) oxidation rates observed in our experiments compared to previous experiments conducted under continuous illumination, likely due to higher O_2_ production during the light periods.
In addition to day-night-cycles, shaking the incubations enhanced cyanobacterial growth. Multiple studies have shown that Synechococcus strains benefit from fluid movement during growth, as it promotes homogenization of the medium and improves access to nutrients, light, and CO_2_^54–56^. For example, Kuan and colleagues reported optimal growth rates for Synechococcus elongatus PCC7942 at shaking speeds of 100–150 rpm^57^. Our data are consistent with these findings, confirming the fluid agitation—analogous to conditions in the phototrophic zones of ancient oceans—stimulates cyanobacterial activity and, consequently, O_2_ production.
Beyond shaking, the configuration of incubation bottles also appears to influence cyanobacterial growth. More specifically, experiments with open bottles generally show faster doubling rates and higher cell densities^58^ compared to setups using closed bottles^11,12^. This difference may result from continuous headspace ventilation in open bottles, which facilitates O_2_ removal and potentially reduce ROS formation. However, comparisons between these studies should be made cautiously, as the Fe(II)(aq) concentrations used by Hermann and Gehringer were relatively low (20-120 µM)^58^, a range in which Swanner and colleagues also observed minimal ROS-related toxicity^11,12^.
Implications for the oxygen distribution in the seawater
From our laboratory-derived per-cell O_2_ production rates, we estimate that under low upwelling conditions (4 m yr⁻¹) (modern value) and a total Fe(II)(aq) concentration of 529 µM, the entire photic zone should be oxidized at cyanobacterial cell densities of approximately 10⁵–10⁶ cells mL⁻¹. At substantially higher upwelling rates (95 and 473 m yr⁻¹)^38^, overall O_2_ concentrations throughout the water column would be lower; nevertheless, cyanobacterial activity would still support O_2_ accumulation at the surface, forming a distinct oxygenated layer.
Incorporating the upwelling rates and Fe(II)(aq) concentrations applied in the studies of Swanner and colleagues^12^ into our models (see Fig. S6–S8), and applying our most conservative O_2_ production rate (r_photo_ of 0.122 µM/day), yields maximum O_2_ concentrations at least an order of magnitude greater than previously estimated (33 µM of maximum oxygen compared to 2–4 µM of Swanner and colleagues^12^). Moreover, the modest toxicity effects observed at 529 µM Fe(II)(aq) would be even less pronounced at lower concentrations (25–125 µM), as also reported by Swanner et al.^11,12^, suggesting that our model predictions may still underestimate true O_2_ production.
When applying this approach to specific scenarios, it produces results that diverge markedly from earlier models^12^. For example, with 25 µM Fe(II)(aq) and an upwelling rate of 95 m yr⁻¹, our model predicts O_2_ concentrations up to 30 µM and a Fe(II)(aq)-free zone extending to more than 200 m depth, whereas the earlier model estimated a maximum of 1–2 µM O_2_ and a Fe(II)(aq)-free zone less than 50 m deep. Under 25 µM Fe(II)(aq) and rapid upwelling (473 m yr⁻¹), both models predict Fe(II)(aq) transport to the surface, but our simulations still yield 2.5 µM O_2_ at the surface, while the earlier study predicted no measurable O_2_ accumulation. Finally, under low upwelling (4 m yr⁻¹) and higher Fe(II)(aq) concentrations (120 µM), our model produces 500 µM O_2_ with complete Fe(II)(aq) oxidation down to 400 m, in contrast to ~80 µM O_2_ and oxidation to only 250 m in the earlier model. Collectively, these findings demonstrate that using experimentally derived cyanobacterial O_2_ production rates results in substantially higher predicted O_2_ concentrations and deeper Fe(II)(aq)-free zones than previous modeling efforts have suggested.
Oxygenic photosynthesis on early Earth proceeded without harmful ROS stress to cyanobacteria
Our experiments involving cyanobacterial incubations at high Fe(II)(aq) concentrations in the absence of SiO_2(aq)_ are consistent with previous studies demonstrating that O_2_-producing cyanobacteria were exposed to ROS under early Earth conditions^11,12^. However, our results suggest that ROS were not necessarily a major physiological stressor for cyanobacteria during this time. Dreher and colleagues^36^ reported increasing Fe(II)(aq) toxicity with increasing Fe(II)(aq) concentrations, but observed markedly enhanced bacterial growth in the presence of silica. Based on these results, we hypothesize that silica reacts with Fe(II)(aq), thus distinctly lowering Fe(II) reactivity and preventing Fe(II)-induced ROS formation. First, in experiments with iron and silica concentrations relevant to Archean oceans (500 µM Fe(II)(aq) and 2200 µM SiO_2(aq)^18,19^), we observed rapid cell growth, high O_2 production, and complete Fe(II)(aq) oxidation under simulated day-night-cycles (Fig. 2D–F). Second, we found that SiO_2(aq)_ effectively suppressed ROS formation at Fe(II)(aq) concentrations as high as 5000 µM (Fig. 2G–L and Fig. 3). These findings imply that cyanobacteria inhabiting coastal environments with episodically elevated Fe(II)(aq) concentrations (several mM^8,59^) could have remained viable and active—particularly in contrast to the lower Fe(II)(aq) concentrations generally inferred for bulk seawater (0.02–500 µM respectively^14,35,60–62^).
The observation that ROS formation was inhibited even at high concentrations of both SiO_2(aq)_ (2200 µM) and Fe(II)(aq) (5000 µM) suggests that these chemical conditions imposed minimal stress to cyanobacteria inhabiting early Precambrian oceans. This finding broadly supports the findings of Swanner and colleagues^11,12^, although in our experiments, lower Fe(II)(aq) concentrations did not induce significant ROS formation or toxicity. Taken together, our lab experiments, supported by modeling, indicate that ROS toxicity was likely not a major constraint on cyanobacterial growth or proliferation in early marine environments. Consequently, ROS stress alone is unlikely to explain the temporal lag between the emergence of oxygenic cyanobacteria ( > 3 Ga) and the onset of the GOE (2.5–2.3 Ga)^31,63–65^.
Our results add to the growing body of evidence demonstrating the geomicrobiological significance of Si-Fe interactions in early Precambrian oceans. For instance, SiO_2(aq)_ binds to reactive surface sites of Fe-bearing minerals such as pyrite (FeS_2_), stabilizing it against oxidative dissolution and inhibiting ROS formation, a process that might have been relevant in the Archean^66,67^. Indeed, silica coatings on pyrite have been shown to reduce surface-bound sulfate generation during oxidative weathering by up to 91%^68^, indicating an increased stability of the pyrite. Furthermore, Fe(III)-silica aggregates can serve as adsorption sites for Fe(II) via outer-sphere complexation^69^, promoting scavenging of Fe(II)(aq) and therefore suppressing ROS production. Our results expand this knowledge by demonstrating that SiO_2(aq)_ can inhibit ROS formation directly in the water column by binding to Fe(II)(aq), thereby contributing to a more favorable chemical environment for cyanobacterial activity in early oceans.
Methods
Cultivation of microorganisms and growth conditions
The cyanobacterium Synechococcus sp. PCC 7002, provided by Gen Enomoto from the University of Freiburg, was cultivated in liquid oxic A+ medium. This medium was prepared using the following stock solutions: Stock 1 (2 mL) (1:2 dilution, containing 3.89 g FeCl₃·6H₂O, 34.3 g H₃BO₃, 4.3 g MgCl₂·4H₂O, 0.315 g ZnCl₂, 0.03 g MoO₃, 0.003 g CuSO₄·5H₂O, and 0.0122 g CoCl₂·6H₂O in Milli-Q water to 1 L), stock 2 (10 mL) (100 g Trizma base adjusted to pH 8.2 with HCl in Milli-Q water to 1 L), stock 3 (100 mL) (0.5 g KH₂PO₄, 10 g NaNO₃, 24.4 g MgSO₄, 180 g NaCl, and 6 g KCl in Milli-Q water to 1 L), stock 4 (10 mL) (0.3 g Na₂EDTA in Milli-Q water to 100 mL), stock 5 (10 mL) (2.8 g CaCl₂ in Milli-Q water to 100 mL). The PCC 7002 culture was grown on a shaker at 60 rpm to a cell density of approximately 10⁸ cells/mL at 25 °C in an Erlenmeyer flask, under a 40-watt halogen light bulb providing an intensity of 300–500 lux.
Setup of Fe(II) oxidation experiments
The Fe(II) oxidation experiments aimed to culture the planktonic cyanobacterium Synechococcus sp. PCC 7002 in the presence of 0, 500, 2500, and 5000 µM Fe(II), along with 0 and 2200 µM dissolved monomeric silica. These experiments were set up in triplicate bottles each to analyze cell growth and accumulated oxygen concentration after complete Fe(II) oxidation by the O_2_ produced by the cyanobacteria. The bottles were kept at room temperature on a shaker at 60 rpm with day-night-cycles of 8/16 h at intensities of 300–500 lux. Each experiment was conducted in 250 mL Schott bottles containing 100 mL of liquid medium. Anoxic artificial seawater medium (ASW) was prepared with the following composition: 17.3 g NaCl, 8.6 g MgCl₂·6H₂O, 0.025 g MgSO₄·7H₂O, 0.99 g CaCl₂·2H₂O, 0.39 g KCl, 0.059 g KBr, 0.05 g KH₂PO₄, 0.25 g NH₄Cl, and 2.5 g NaHCO₃ per 1 L of Milli-Q (MQ) water. Salts, the bicarbonate buffer (30 mM), and the monomeric silica stock solution were separately purged with N₂ (or N₂/CO₂ at 90:10 % for the buffer) and autoclaved. After autoclaving, the salt solution in a Widdel flask was flushed with N₂/CO₂ while still hot (~70–80 °C), and the sterile silica solution was immediately added, resulting in the formation of white flocs. The medium was cooled to 20 °C, the bicarbonate buffer was added, and the pH was adjusted to 7.0 using 1 M anoxic HCl. Due to precipitate formation, the medium was transferred to 2 L Schott bottles and kept at room temperature for two days. It was then filtered through a 0.22 µm PES bottle-top filter inside a glovebox. Vitamins and trace element solutions were added under sterile conditions, and the pH was re-adjusted to 7.0. The prepared medium was aliquoted into sterile 250 mL Schott bottles, with each bottle containing a total volume of 100 mL, which included 5 mL of washed cyanobacteria and the respective FeCl₂ solution. During this process, a constant stream of N₂/CO₂ was maintained. Oxygen concentrations were measured, and any bottles with more than 3 µM oxygen were flushed with N₂/CO₂ again. Subsequently, FeCl₂ was added from an anoxic and sterile 100 µM or 1 M stock solution. The pH values were rechecked and remained stable at 7.0. Cyanobacteria were washed three times with ASW before being added to the medium as a 5% inoculum to achieve a final cell density of 10⁶ cells/mL. The experiments were initially fully anoxic, so all oxygen detected during the course of the experiments was produced solely by the inoculated cyanobacteria.
Quantification of Fe(II) and Fe(III) by the spectrophotometric Ferrozine assay
The ferrozine assay is a spectrophotometric method used to quantify the concentration of Fe(II). Samples were first dissolved and diluted in 1 M HCl to achieve a concentration of up to 1 mM Fe. For Fe(II) analysis, 80 µL of 1 M HCl was pipetted into a 96-well plate, followed by the addition of 20 µL of the sample. After a 15-min incubation, 100 µL of ferrozine solution (0.1 w/v) was added. The mixture was allowed to complex for 5 min before being analyzed using a Multiskan™ GO Microplate Spectrophotometer (Thermo Fisher Scientific, USA). Fe(II) is forming a purple complex with ferrozine molecules, which absorbs light at 562 nm. For total iron (Fe_(tot)_) determination, the procedure was modified by replacing the 1 M HCl with 80 µL of 10% (v/v) hydroxylammonium chloride (HAHCl) before adding the sample. This step was followed by a longer incubation period of 30 min to ensure the complete reduction of Fe(III) to Fe(II). To prevent degradation, both the ferrozine and HAHCl solutions were stored and incubated in the dark, as they are highly light-sensitive. Different standards between 0 and 1000 µM were used to calculate the respective iron concentration.
Quantification of dissolved silica by the molybdenum blue method
Dissolved silica, including monomeric silica and colloidal silicic acid (Si-Si-colloids), was quantified by the Molybdenum Blue Method^70^. The samples were filtered (0.22 µm, PES) and diluted in MQ to 1 mL. Subsequently, 40 µL of an ammonium heptamolybdate tetrahydrate (6.33 g) together with 50 mL of 4.5 M sulfuric acid solution were added to form a blue complex with the silica. To avoid complexing with phosphate, 40 µL of a 10 g/100 mL MQ oxalic acid solution was added. Lastly, 20 µL of 0.16 M ascorbic acid was added as reductant. After 30–60 min of incubation, the blue silica-molybdenum complexes were quantified at 810 nm. Different standards between 1 and 100 µM were used to calculate the silica concentration. Due to the filtration of the samples (0.22 µm) and removal of Si precipitates, the measured remaining silica fraction is named ‘dissolved silica’.
Oxygen quantification by optode sensors
Free dissolved oxygen was quantified using a luminophore optode foil from PreSens. Small 3 by 3 mm pieces of foil were affixed with silicone glue within the lower 2 cm of the bottles to ensure direct contact with the liquid phase. The interaction between the optode foil (luminophore) and oxygen molecules (quencher) results in an energy transfer that reduces the luminescence signal of the foil. The oxygen concentration in the liquid phase was then calculated using the Stern-Volmer equation. A temperature sensor was placed in a co-incubated Schott bottle under identical experimental conditions to account for temperature variations. Each container and optode foil setup underwent a fresh two-point calibration before measurements. The calibration included one data point at full oxygen saturation and another one under oxygen-free conditions. For the first calibration point, 100 mL of pure ASW medium was added to a 250 mL Schott bottle containing the optode foil and stirred vigorously until complete O₂ saturation was achieved. For the second calibration point, the reductant sodium dithionite (Na₂S₂O₃) was added to the same bottle to eliminate all remaining dissolved oxygen.
Intracellular ROS measurements
The membrane-permeable dye CM-H_2_DCFDA (5-(and-6)-chloromethyl-2′,7′-dichlorodihydrofluorescein diacetate, acetyl ester; Life Technologies GmbH) was used to quantify intracellular reactive oxygen species. Log-phase cultures were harvested via centrifugation and washed twice with TRIS buffer (pH 7.0). Cells were adjusted to a concentration of 5 × 10^8^ cells/mL in TRIS buffer and degassed in the dark using a 90:10 N_2_/CO_2_ gas mixture. FeCl_2_ was added from anoxic stock solutions to achieve final concentrations of 0, 500, 2500, and 5000 µM We then added pH-adjusted silica from a metasilicate stock solution, pre-heated to 90 °C in a water bath to monomerize the silica, leading to a final concentrations of 0 and 2200 µM in duplicates in two different experimental runs. The suspensions were shaken for one hour. After incubation, cells were resuspended in fresh buffer containing 5 µM CM-H_2_DCFDA and incubated in the dark for 30 min. Following a second resuspension in fresh buffer, fluorescence emission at 519 nm (excitation at 490 ± 5 nm) was recorded using the infinite 200 PRO Fluorometer from Tecan Lifesciences. Background fluorescence from untreated cells and Fe minerals was subtracted to ensure accuracy.
Cell numbers by hemocytometry
The cell density was visually counted under a Leica CTR 5500 microscope using a Neubauer-improved hemocytometer from Hirschmann. Particularly, 10 µL of sample was pipetted between sample holder and coverslip. At a magnification of 400x, the cells were counted in the smallest squares until a final count of 100 cells. 180 µL cells were fixed by the addition of 20 µL 21% PFA solution. The minerals were dissolved by the addition 200 µL of 100 mM Fe(II)-EDAS solution and 600 µL of ca. 0.8 M oxalic acid The cell count per small square was then multiplied by the dilution factor and by 2.5 × 10^7^ as volume factor to get the real cell count in mL/L.
MATLAB model from lab experimental data
We modeled our lab experimental data to quantify total oxygen production in the system in MATLAB (version R2023b), considering both dissolved oxygen and the contribution from the headspace. Based on the equilibrium partitioning between the aqueous solution in the head space based on the Henry-Law (see SI formula 9), oxygen production including the headspace is approximately 42 times greater than what is measurable in the dissolved liquid phase alone. Additionally, increasing cell density caused greater turbidity in the liquid phase, which led to shading effects reducing the overall photosynthetic rate per cell. We accounted for this by modeling the proportion of active versus inactive cells in the system. Detailed methods and the corresponding MATLAB code are provided in the Supplementary Information (SI).
MATLAB model to mimic the oxygen and Fe(II) distribution in the ancient ocean
To simulate oxygen distribution in ancient ocean water columns, we used the photosynthetic rate (r_photo_) derived from our lab experiments under the most realistic conditions—500 µM Fe(II) and 2200 µM silica. We divided the experiment into two phases: Phase 1, when dissolved Fe²⁺ was still present, mimicking early ocean conditions, and Phase 2, after Fe²⁺ oxidation, to analyze the long-term bacterial response following Fe²⁺ exposure. From Phase 1, we calculated the daily oxygen production per cell (r_photo_). According to reaction (5), one molecule of O₂ can oxidize four Fe²⁺ ions.
\documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$${{{\rm{O}}}}_{2}+4{{{\rm{Fe}}}}^{2+}+8{{\rm{O}}}{{{\rm{H}}}}^{-}\leftrightarrow 4{{\rm{FeOOH}}}+2{{{\rm{H}}}}_{2}{{\rm{O}}}$$\end{document}We determined \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$${r}_{{photo}}$$\end{document} from the first two days of the experiment, during which no free oxygen was detected, and cyanobacteria were still in the lag phase without measurable growth, allowing the simplification that all produced oxygen was used for Fe(II) oxidation and the cell count stayed the same. Starting with 529 µM Fe(II) at day 0, 150 µM was oxidized over two days, corresponding to a rate of 75 µM/day Fe(II) oxidation, which equates to 18.75 µmol O₂/day based on reaction (5). Due to uncertainty in manual cell counts, we calculated \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$${r}_{{photo}}$$\end{document} for two different cell densities observed in our experiments: 6.48 × 10⁶ cells/mL (experimental cells 1) and 1.6 × 10⁷ cells/mL (experimental cells 2). Reported early ocean cyanobacterial concentrations range between 10⁵cells/mL (target cells 1) and 10⁶ cells/mL (target cells 2), so we modeled four scenarios combining these experimental and target cell densities to calculate \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$${r}_{{photo}}$$\end{document} values (see Table 3) using formula 6. Based on the study of Swanner and colleagues^12^, we further estimated oxygen distribution in the early ocean water column for upwelling rates of 4, 94, and 473 m/year.
\documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$18.75\,\frac{{{\rm{\mu }}}M}{{day}} * \frac{{target}\,{cells}}{{experimental}\,{cells}\,}={r}_{{photo}}$$\end{document}Table 3. Photosynthetic rate (oxygen production per cell/ day) of the lab experiments calculated from 6.48 × 10⁶ cells/mL (‘experimental cells 1’) and 1.6 × 10⁷ cells/mL (‘experimental cells 2’) for 10⁵ cells/mL (target cells 1) and 10⁶ cells/mL (target cells 2) in the early ocean \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$${r}_{{photo}}$$\end{document} Experimental cells 1Experimental cells 2Target cells 10.289 µM/day2.89 µM/dayTarget cells 20.112 µM/day1.12 µM/day
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Supplementary information
Supplementary information Peer Review file Reporting Summary
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kasting, J. F., Schopf, J. W. & Klein, C. Models relating to Proterozoic atmospheric and oceanic chemistry. The Proterozoic Biosphere: A Multidisciplinary Study, 1185–1187 (1992).
- 2Swanner, E. D. et al. Physiology, Fe (II) oxidation, and Fe mineral formation by a marine planktonic cyanobacterium grown under ferruginous conditions. Front. Earth Sci.3, 60 (2015).
- 3Trabucho Alexandre, J. et al. The mid‐Cretaceous North Atlantic nutrient trap: black shales and OA Es. Paleoceanography 25 (2010).
- 4Wydrzynski, T. J. & Hillier, W. Molecular Solar Fuels (Royal Society of Chemistry, 2011).
- 5Herrmann, A. J. & Gehringer, M. M. A low-cost automized anaerobic chamber for long-term growth experiments and sample handling. Hardware X, 10, e 00237 (2021).
- 6Mel’nik, Y. P. Precambrian Banded-Iron-Formations. (Elsevier, 1982).