Alpha-synuclein overexpression triggers divergent cellular responses and post-translational modifications in SH-SY5Y and ReNcell VM models
Miraj Ud Din Momand, Petra Majerova, Diana Mjartinova, Natalia Maruskinova, Karolina Albertusova, Michael Dobrota, Lubica Fialova, Sara Stefankova, Petar Podlesniy, Muhammad Khalid Muhammadi, Miroslav Balaz, Dominika Fricova

TL;DR
This study shows how overexpressing alpha-synuclein in different human neuronal cells leads to varied effects on cell health and protein modifications, highlighting the importance of cell type in Parkinson's disease research.
Contribution
A modular in vitro platform with stable cell lines for studying alpha-synuclein overexpression in two human neuronal models.
Findings
A53T alpha-synuclein consistently causes cytotoxicity and mitochondrial dysfunction in both SH-SY5Y and ReNcell VM cells.
Wild-type alpha-synuclein has divergent effects on mitochondrial function depending on the cell type.
Post-translational modification patterns of alpha-synuclein vary significantly between cell types, even without aggregation.
Abstract
Alpha‑synuclein (α‑syn) overexpression models are widely used to unravel the molecular mechanisms of Parkinson’s disease (PD), particularly in light of the dose-dependent transition between its physiological and toxic roles. However, existing systems rely on inducible expression, lack robust dose stratification and comparative cellular contexts. Here, we developed and characterized a panel of stable neuronal cell lines in two human cellular models (SH‑SY5Y neuroblastoma cells and ReNcell VM neural progenitors) overexpressing GFP-tagged wild-type (WT) or A53T mutant α‑syn at low and high overexpression levels. Utilizing this framework, we demonstrated that A53T consistently induces cytotoxicity, oxidative stress and mitochondrial dysfunction in both cell types. In contrast, WT α‑syn had divergent effects depending on the cellular context. In SH‑SY5Y cells, it enhanced mitochondrial…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
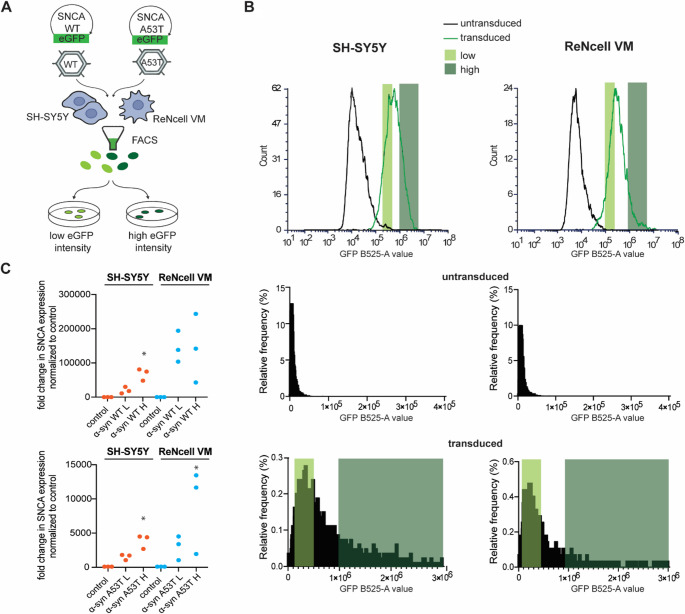 Figure 1
Figure 1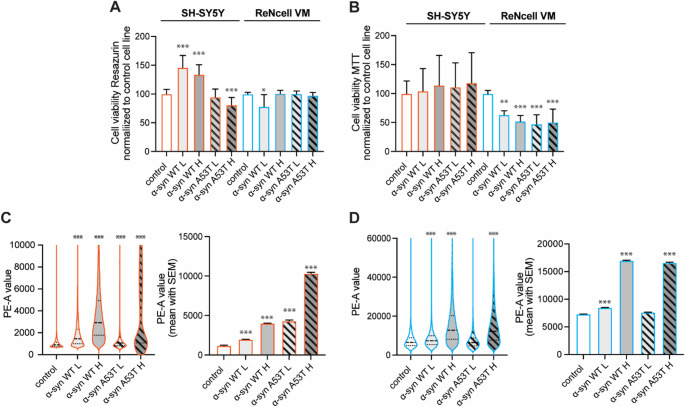 Figure 2
Figure 2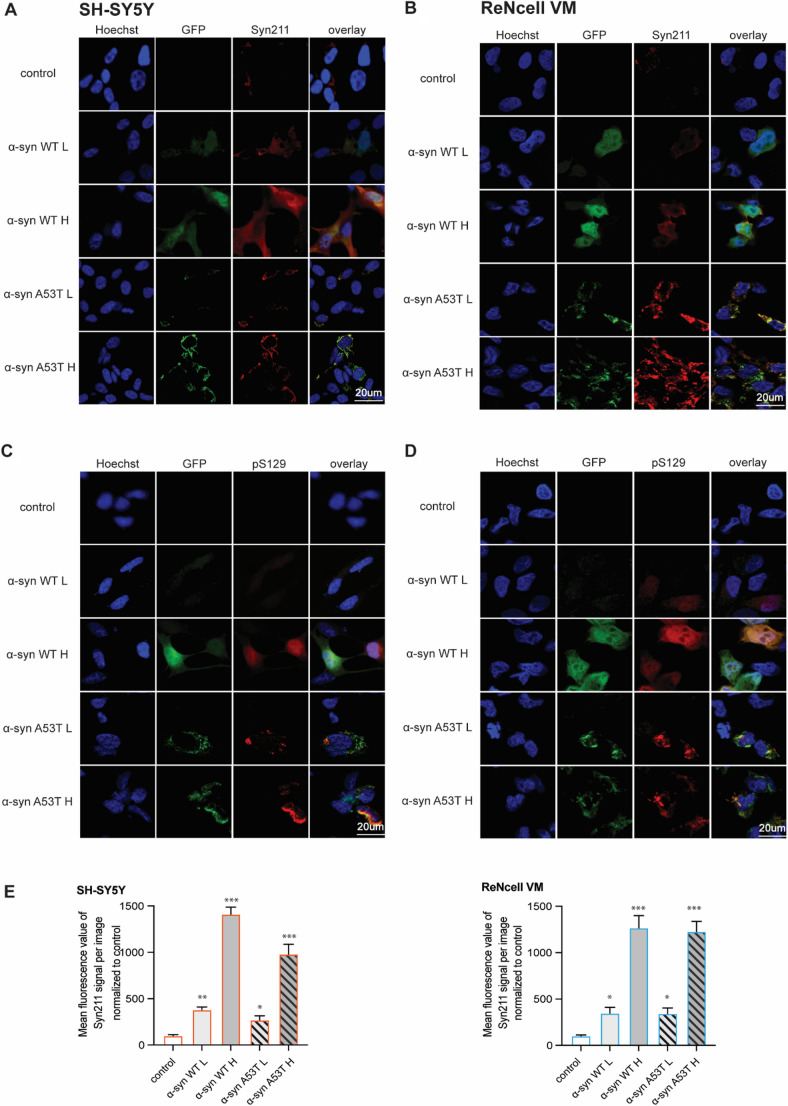 Figure 3
Figure 3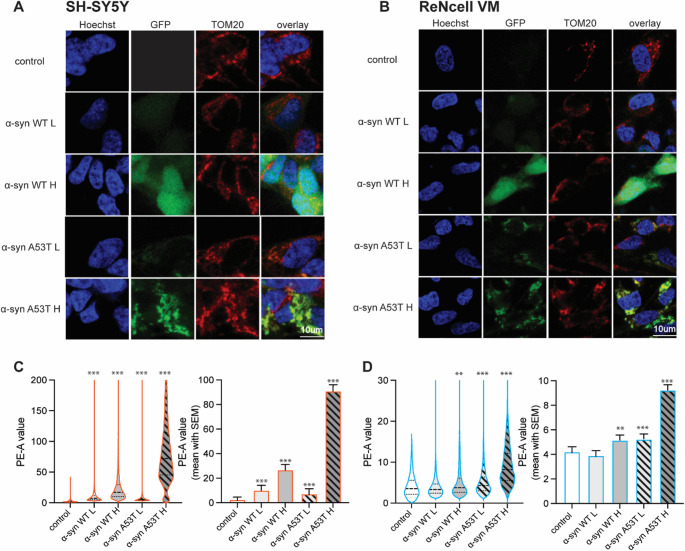 Figure 4
Figure 4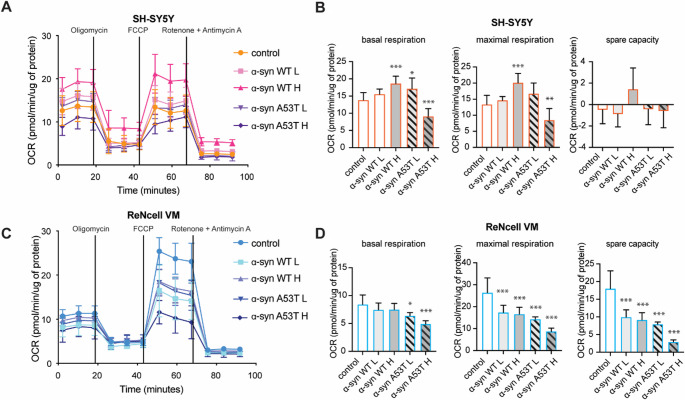 Figure 5
Figure 5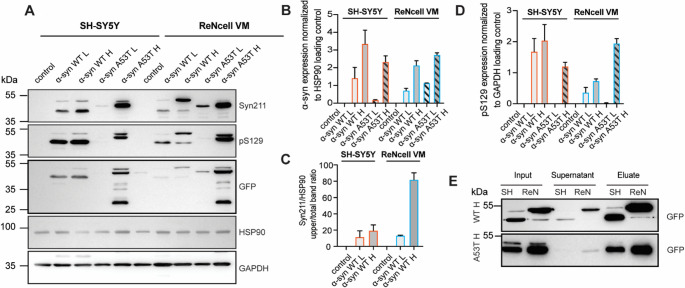 Figure 6
Figure 6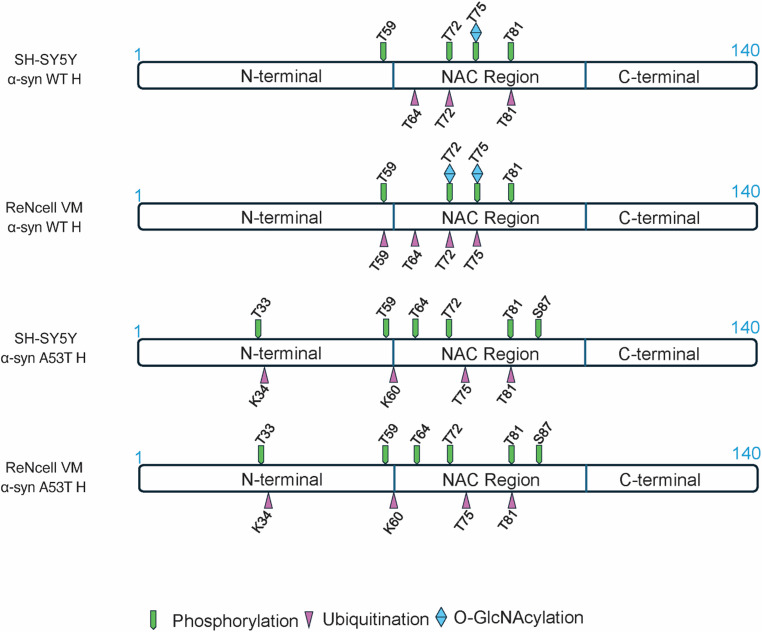 Figure 7
Figure 7- —Pavol Jozef Šafárik University in Košice
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParkinson's Disease Mechanisms and Treatments · Nuclear Receptors and Signaling · Neurological disorders and treatments
Introduction
Cell-based models remain essential for investigating alpha-synuclein (α‑syn)-related pathology in Parkinson’s disease (PD), as they allow controlled manipulation of expression levels and capture the dose-dependent shift in α‑syn function from physiological to pathological states. While physiological levels support normal neuronal activity, elevated expression as observed in gene multiplications linked to familial PD [1–3], drives toxicity through mechanisms such as mitochondrial dysfunction, impaired vesicle trafficking, and altered protein homeostasis [4, 5]. These models have employed diverse cellular backgrounds, ranging from non-neuronal HEK293T and H4 neuroglioma cells [6] to human-derived neuronal models such as SH-SY5Y neuroblastoma cells [7, 8], primary neurons [8], patient-derived fibroblasts, and induced pluripotent stem cells [9]. Among these models, SH-SY5Y neuroblastoma cells stably overexpressing wild-type (WT) or PD-related A53T mutant form of α-syn have emerged as a cornerstone for studying α-syn-mediated toxicity. Their widespread use has provided mechanistic insights into several key aspects of PD pathology. Notably, α-syn overexpression in SH-SY5Y cells has been shown to induce marked mitochondrial fragmentation, primarily through inhibition of mitochondrial fusion and promotion of fission processes, with A53T-expressing cells exhibiting more pronounced phenotypes [10]. α-syn overexpression in SH-SY5Y revealed impaired calcium homeostasis, particularly at mitochondria-associated endoplasmic reticulum membranes, where α-syn disrupts calcium transfer [11]. Further studies using SH-SY5Y cells have demonstrated that elevated α-syn impairs autophagic flux and lysosomal function, contributing to defective clearance of protein aggregates [12]. Additionally, α-syn-overexpressing SH-SY5Y cells display dysregulated iron metabolism, including altered iron uptake and homeostasis, which may contribute to oxidative stress [13]. Exosomes derived from α-syn overexpressing SH-SY5Y cells have been shown to cause impairment of autophagy in recipient cells [14]. Beyond functional impairments, these models have been instrumental in the characterization of α-syn post-translational modifications (PTMs) related to PD, especially phosphorylation, which is commonly observed in Lewy body inclusions in PD brains [15–17]. Despite their utility, SH-SY5Y cells carry limitations that constrain their physiological relevance. They represent a single cancer-derived lineage, often lack stratified expression models to test dosage-dependent effects, and have not been extensively compared to other human neuronal models.
In this study, we established a robust dual-cell model to address these limitations by using two human-derived neuronal lines: SH‑SY5Y and ReNcell VM. ReNcell VM, originating from human fetal ventral mesencephalon and immortalized via v‑myc, more closely resembles neural progenitors of the substantia nigra, a region critically affected in PD, making it a highly relevant model for comparison. We introduced stable overexpression of GFP-tagged WT or A53T α-syn via lentiviral vectors in both cell types and stratified the populations into low and high α-syn expressers, enabling dose-dependent comparisons. This multifaceted approach allowed us to directly compare α-syn-induced effects in two distinct neuronal cell types, investigate how WT and mutant A53T α-syn differentially impact cellular homeostasis, systematically assess dose-dependent responses to α-syn overload, a rarely addressed dimension in previous models. Our findings reveal that ReNcell VM cells exhibit pronounced sensitivity to α‑syn-induced mitochondrial dysfunction and display a distinct PTM profile compared to SH‑SY5Y cells. While previous models using SH‑SY5Y cells have highlighted phosphorylation at serine‑129 (pS129) as a hallmark of α‑syn pathology, our study demonstrates that this modification occurs more robustly and in a broader PTM landscape in ReNcell VM cells, even in the absence of overt aggregation. This suggests that specific cellular environments can influence conformational state and pathogenic transition of α‑syn. Collectively, these findings underscore the critical importance of cellular context, α‑syn variant, and overexpression dosage in shaping pathological outcomes, and they highlight the need for diversified models to more accurately capture early disease mechanisms in PD.
Materials and methods
Generation and maintenance of stable cell lines
Two cell lines used in this study were human neuroblastoma SH-SY5Y cells (Sigma-Aldrich, USA, 94030304) and human neural progenitor cell line ReNcell VM (Millipore, Germany, SCC008), generated by insertion of a v-myc oncogene into human fetal ventral mesencephalic cells. SH-SY5Y cells were maintained in DMEM medium (Gibco, USA, 11960044) supplemented with 10% FBS (Biosera, France, FB-1090/500), 2 mM L-glutamine (Biosera, France, XC-T1715/100), 1% penicillin/streptomycin (100 U/ml penicillin, 100 µg/ml streptomycin) (Biosera, France, LM-A4118). ReNcell VM cells were maintained in Matrigel (Corning, USA, 356234) coated plates with ReNcell NSC Maintenance Medium (Sigma-Aldrich, USA, SCM005) supplemented with 20 ng/ml fibroblast growth factor-2, 20 ng/ml epidermal growth factor, 1% penicillin/streptomycin. Cell lines stably overexpressing WT or A53T mutant α-syn were generated using eGFP-α-syn (referred to in text as GFP-α-syn) WT or A53T plasmids, a gift from David Rubinsztein (via Addgene, USA), through lentivirus transduction. Briefly, lentivirus particles were produced in HEK293T cells using standard third-generation packaging. HEK293T cells were transfected at ~ 70% confluency with the transfer plasmid (Addgene #36083 for eGFP, #40822 for WT α-syn, and #40823 for A53T α-syn), along with packaging plasmids psPAX2 (Addgene #12260) and pMD2.G (Addgene #12259) in a 4:3:1 ratio. Viral supernatants were collected at 48 h post-transfection, pooled, filtered through 0.45 μm filters. Functional viral titers were determined by serial dilution on SH-SY5Y and ReNcell VM cells, followed by flow cytometric quantification of GFP-positive cells at 48 h post-transduction. GFP expression levels were confirmed by both flow cytometry and fluorescence microscopy. To maintain defined expression profiles and minimize phenotypic drift, cells were used between passages 1–5 post-sorting. Experimental workflows were standardized across assays, cells were thawed from cryopreserved early-passage stocks (Day 0), cultured to 70–80% confluency by Day 3, and seeded into assay-specific plates or coverslips. Assays were performed on Day 5, once cells reached optimal confluency. This timeline was applied consistently for all assays and experimental replicates.
Cell sorting and flow cytometry analysis
Stably transduced cells were sorted based on GFP expression using CytoFLEX SRT (Beckman, USA). Cells were resuspended in phosphate-buffered saline (PBS) for sorting, GFP was excited at 488 nm, and emission was collected using a 525/40 nm filter. Cell lines were sorted into low and high overexpression populations. Sorted cells were collected in a tube containing growth medium and transferred to cell culture dishes. Successful sorting of GFP-positive cells was confirmed through microscopy. These cells were then expanded and used for subsequent experiments. To minimize variability and potential adaptation to α‑syn-induced stress, all cell lines were either freshly thawed or maintained in culture for no more than five passages. For the CellROX assay, cells were seeded and stained upon reaching 70% confluence with CellROX Orange reagent (Thermo Scientific, USA, C10443) at a final concentration of 2.5 µM for 30 min in growth conditions (humidified chamber with 37 °C, 5% CO₂). After 30 min, cells were detached, resuspended in PBS, and analyzed via flow cytometry using LSRFortessa (BD Biosciences, USA) with a 488 nm laser and 540/40 nm filter for GFP and 561 nm laser and 586/15 nm filter for CellROX Orange. For mitochondrial staining, cells were stained with 1 µM MitoTracker Red FM (Thermo Scientific, USA, M22425) solution for 30 min in growth conditions. After incubation and detachment, cells were resuspended in 500 µl PBS supplemented with 5% FBS. Cell suspensions were analyzed on a MACSQuant (Miltenyi Biotec, Germany) flow cytometry analyzer using a 488 nm laser and a 585/40 nm filter for GFP and a 561 nm laser and a 655/20 nm filter for MitoTracker. 10,000 cell events were probed and compared to unstained control cells. For the detection of activated caspase-3 and caspase‐7 proteins in apoptotic cells, cells were stained with CellEvent Caspase‐3/7 detection reagent red (Thermo Scientific, USA, C10430) for 1 h in growth conditions. After incubation and detachment, cells were resuspended in PBS. Cell suspensions were analyzed via flow cytometry using LSRFortessa (BD Biosciences, USA) with a 488 nm laser and 540/40 nm filter for GFP and a 561 nm excitation laser and 610/20 emission filter for Caspase‐3/7 detection red reagent. 10,000 cell events were probed and compared to unstained control cells.
Quantitative real-time polymerase chain reaction (qPCR)
Total RNA was extracted from cells using High Pure RNA Isolation Kit (Roche, Switzerland, 11828665001). cDNA was synthesized with 100 ng of total RNA using High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, USA, 4368814). Diluted cDNA (1:10) was used for gene expression analysis, each sample in triplicates mixed with 1x TaqMan Gene Expression Master Mix and relevant 1x TaqMan probes (Applied Biosystems, USA) Hs00240906_m1 or Hs04194366_g1 targeting SNCA or housekeeping gene (RPL13A) respectively. qPCR was performed on a QuantStudio 5 RT-PCR System (Applied Biosystems, USA). Reactions (10 µl) were performed using the following cycling program: hold at 50 °C for 2 min, initial denaturation at 95 °C for 10 min, followed by 40 cycles of denaturation for 15 s and annealing/extension at 60 °C for 1 min. Relative mRNA levels were calculated according to the 2 − ΔΔCt method [18].
Immunoblotting
Cells were lysed in RIPA or M-PER Mammalian Protein Extraction Reagent (Thermo Scientific, USA, 78503) supplemented with cOmplete protease inhibitor cocktail (Roche, Switzerland, 05056489001) on ice for 30 min, centrifuged at 13,000xg for 15 min, and quantified with Pierce BCA Protein Assay Kit (Thermo Scientific, USA, 23225). Equal amounts of protein were separated on SDS-PAGE and transferred to a nitrocellulose (Cytiva, USA, 45004001) or polyvinylidene fluoride (PVDF; Thermo Scientific, USA, 88518) membrane. For α-syn detection, the membranes were fixed with 0.4% paraformaldehyde (PFA) and then blocked with 5% milk. The membranes were probed overnight with the antibodies Syn211 (Invitrogen, USA, AHB0261), HSP90 (Cell Signaling Technology, USA, 4877), anti phospho-α-syn pS129 (Cell Signaling Technology, USA, 23706), GAPDH (Cell Signaling Technology, 5174, 97166), PARP (Cell Signaling Technology, 9542), TOM20 (Cell Signaling Technology, 42406), beta tubulin (Cell Signaling Technology, 2146) diluted 1:1000, and anti-GFP tag antibody (Proteintech, USA, 66002-1-Ig) diluted 1:5000. Membranes were then incubated with suitable secondary antibodies conjugated to horseradish peroxidase (Agilent, USA, P0447, P0448). The signal was detected using SuperSignal West Pico PLUS Chemiluminescent Substrate (Thermo Scientific, USA, 34580) on iBright 1500 Imaging System (Invitrogen, USA).
Immunocytochemical and mitochondrial staining
For immunocytochemistry staining, cells were washed with PBS, fixed in 4% PFA for 15 min, and permeabilized with 0.1% Triton X-100 for 30 min at room temperature. Non-specific binding was blocked with 2% BSA (Sigma-Aldrich, USA, 9048-46-8) for 1 h at room temperature and stained with primary antibody Syn211 (Invitrogen, USA, AHB0261), pS129 (Cell Signaling Technology, USA, 23706), and TOM20 (Cell Signaling Technology, USA, 42406) diluted 1:500. All primary antibodies were diluted in 0.1% BSA and incubated overnight at 4 °C. Cells were subsequently incubated with compatible secondary antibodies conjugated to Alexa-546 or Alexa-594 fluorophores (Invitrogen, USA, A-11030, A-11012) diluted 1:1000 for 1 h at room temperature. The nuclei were stained with Hoechst (Invitrogen, USA, 62249) for 10 min, and cells were mounted in Fluoromount-G mounting medium (Merck, USA, F4680). Stained samples were imaged with a Zeiss LSM 710 confocal microscope (Zeiss, Germany).
Cell viability analysis
Both SH-SY5Y and ReNcell VM cell lines were seeded in respective growth conditions in triplicates at a starting density of 10,000 cells/well in 96 well plates. Cell viability was assessed after cells reached around 70% confluence by adding MTT labeling reagent (Roche, Switzerland, 11465007001) with final concentration 0.5 mg/ml to each well and cells were incubated for 4 h in growth conditions. After 4 h, 100 µl of the solubilization buffer was added into each well and the plate was incubated overnight in growth conditions. The following day absorbance was measured with Varioskan plate reader (Thermo Scientific, USA) at wavelengths of 570 nm and 690 nm. For In Vitro Toxicology Resazurin-based assay (Merck, USA, TOX8) cells were seeded in a similar way. After reaching 70% confluence, resazurin dye solution was added to each well at an amount corresponding to 10% of the total well volume. Cells were incubated for 3 h in growth conditions and fluorescence was measured using a Varioskan multiplate reader with excitation at 560 nm and emission at 590 nm.
Immunoprecipitation and liquid chromatography with mass spectrometry (LC/MS) analysis
GFP-α-syn was immunoprecipitated using ChromoTek GFP-Trap Magnetic Agarose beads (Proteintech, USA). Briefly, 300 µg of total protein lysates were incubated overnight with 25 µl of beads on DYNAL Sample Mixer (Dynal Biotech, Norway) at 4 °C. The following day, beads were spun at 800 rpm for 30 s, supernatant was collected, beads were washed once, and the proteins were eluted in 100 µl of 0.2 M glycine, pH 2.5. pH was neutralized with Tris, pH 9.0, and successful pull-down was validated through immunoblotting with anti-GFP tag antibody. For LC/MS analysis, samples were dried using a SpeedVac and diluted in 8 M urea. Reduction was performed with 10 mM dithiothreitol (Sigma-Aldrich, USA, D9779) at 56 °C for 1 h. Alkylation was carried out with 15 mM iodoacetamide (Sigma-Aldrich, USA, I1149), protected from light for 30 min. Trypsin digestion (Promega, USA, V5280) was performed overnight at 37 °C. The resulting peptide mixtures were separated using an Acquity M-Class Ultra-High-Performance Liquid Chromatography system (Waters, USA), employing a nanoEase Symmetry C18 trap column (20 mm length, 180 μm diameter, 5 μm particle size; Waters, USA) for desalting, followed by separation on a nanoEase High Strength Silica T3 C18 analytical column (100 mm length, 75 μm diameter, 1.8 μm particle size). A 75-minute gradient of acetonitrile (5% to 40%) (Sigma-Aldrich, USA, 45983) containing 0.1% formic acid (Sigma-Aldrich, USA, 5.33002) was applied at a flow rate of 300 nL/min. Analysis was conducted using a Synapt G2-Si quadrupole time-of-flight mass spectrometer with ion mobility (Waters, USA). Data was acquired in a data-independent acquisition (DIA) mode at a spectral acquisition rate of one second. Peak detection and data processing were performed using PEAKS Studio 12.5 (Bioinformatics Solutions, Canada). Peptides were identified using a precursor mass tolerance of 20 ppm and a fragment ion mass tolerance of 0.2 Da. Peptide identification was performed against the human UniProt database supplemented with the GFP-α-syn fusion construct sequence. Peptide identifications were filtered at a 1% false discovery rate (FDR) and required minimum peptide length of 5 amino acids and at least 3 fragments per peptide. Post- translational modifications were assigned as follows: phosphorylation (+ 79.97 Da), ubiquitination (+ 114.04 Da), and O- GlcNAc (+ 203.20 Da). Fixed modification (carbamidomethylation) and variable modification (oxidation) were also included in the search criteria. Immunoprecipitation followed by LC-MS analysis was performed on three independent biological replicates, each obtained from separately cultured cell preparations. Each biological replicate was analyzed in duplicate LC-MS injections.
Mitochondrial bioenergetics profiling using Seahorse XF Pro analyzer
Mitochondrial respiration was assessed using the Seahorse XF96 Extracellular Flux Analyzer (Agilent, USA). SH-SY5Y and ReNcell VM cell lines were seeded at a density of 30,000 cells per well in Seahorse XF96 cell culture microplates (Agilent, USA, 103794-100) coated with Matrigel and incubated overnight in standard growth conditions. The sensor cartridge was hydrated in Seahorse XF Calibrant (Agilent, USA, 100840-000) and placed in a 37 °C non-CO_2_ incubator overnight. On the day of the assay, the culture medium was replaced with Seahorse XF DMEM assay medium (Agilent, USA, 103680-100) supplemented with 10 mM glucose (Agilent, USA, 103577-100), 1 mM pyruvate (Agilent, USA, 103578-100), and 2 mM glutamine (Agilent, USA, 103579-100), adjusted to pH 7.4. Cells were incubated at 37 °C in a non-CO_2_ incubator for 60 min. The compounds were diluted in complete Seahorse XF DMEM medium and loaded into the appropriate ports of the sensor cartridge to reach final concentrations of 1.5 µM oligomycin, 2 µM FCCP, and 0.5 µM rotenone/antimycin A. The cartridge was then placed in the Seahorse XF96 Analyzer for calibration for 20 min. Subsequently, the sensor cartridge was replaced with the cell culture plate and measurement of oxygen consumption rate (OCR) was performed. OCR was analyzed using Wave software (Agilent Technologies, Inc, USA). Data were normalized to total protein content using DC assay.
Statistical analysis
All quantitative data are expressed as mean ± standard error of the mean (SEM), geometric mean or median, as indicated. Each experiment was independently repeated three times. Statistical significance was determined using unpaired two-tailed t test, one-way ANOVA with Dunnett’s correction, or Kruskal–Wallis test with Dunn’s post hoc test, as appropriate. Values of p < 0.05 (), p < 0.01 (), and p < 0.001() were considered statistically significant; p ≥ 0.05 was considered not significant. All statistical analyses were performed using GraphPad Prism version 10.5.0 (774). All figures were created using GraphPad Prism and Adobe Illustrator 2023.
Results
Generated stable α‑syn overexpression neuronal cell models in SH-SY5Y and ReNcell VM cell lines exhibit consistent α-syn (WT/A53T) overexpression
To systematically investigate α‑syn variant’s and dosage-dependent effects across distinct neuronal backgrounds, we developed a comprehensive panel of neuronal cell lines overexpressing α‑syn through lentiviral transduction and fluorescence-activated cell sorting (FACS). Lentiviral constructs encoding either WT or A53T mutant α‑syn fused to GFP leading to N-terminal fusion were generated and used to transduce two human neuronal cell lines: SH-SY5Y neuroblastoma cells and ReNcell VM ventral midbrain neural stem cells (Fig. 1A). Following transduction, cells were sorted by flow cytometry based on GFP fluorescence intensity, enabling precise selection of populations with defined overexpression levels designated as low (L) and high (H) overexpressers. To control for potential effects of GFP expression itself, we generated matched GFP-only low (L) and high (H) overexpressing cell lines in both cell types using identical viral vector backbone containing only GFP and identical sorting criteria (Supplementary Fig. S1). Flow cytometric analysis demonstrated successful transduction efficiency in both cell lines, with clear separation between untransduced and transduced populations based on GFP fluorescence intensity. The transduced populations displayed distributions enabling effective gating strategies for low and high overexpression level selection in both SH-SY5Y and ReNcell VM cells. Untransduced control populations exhibited minimal background fluorescence, confirming the specificity of the sorting methodology (Fig. 1B). This strategy yielded eight genetically defined α‑syn overexpressing cell lines: SH-SY5Y (α-syn WT L, α-syn WT H, α-syn A53T L, α-syn A53T H) and ReNcell VM (α-syn WT L, α-syn WT H, α-syn A53T L, α-syn A53T H), complemented by untransduced control lines for each cell type. qPCR analysis confirmed robust overexpression of SNCA mRNA across all generated cell lines (Fig. 1C). Both SH‑SY5Y and ReNcell VM lines overexpressing WT and A53T α‑syn showed significant increases in SNCA transcript levels compared to their respective controls. Notably, high-expressing groups consistently exhibited several-fold greater mRNA abundance than their low-expressing counterparts, validating the success of the GFP-based sorting strategy. However, we detected variability in SNCA mRNA expression among biological replicates that had undergone extended passaging, likely due to passage-related changes in lentiviral integration efficiency or transcript regulation. To minimize these inconsistencies, all subsequent experiments were performed using freshly thawed cells and limited to early passage numbers. Thus, despite this transcriptional variability, total α‑syn protein levels remained consistent across experimental replicates, as confirmed by immunofluorescence (Fig. 3) and immunoblotting analysis (Fig. 6).
Fig. 1. Generated stable α‑syn overexpression neuronal cell models in SH-SY5Y and ReNcell VM cell lines exhibit consistent α-syn (WT/A53T) overexpression. (A) Brief schematic overview of the workflow used to generate stable cell lines in SH-SY5Y/ReNcell VM cells overexpressing GFP-tagged α-syn WT or A53T mutant via lentivirus transduction. Transduced cells were selected based on GFP expression with FACS. (B) Representative FACS plots illustrating the gating strategy adopted to separate GFP positive and GFP negative populations. GFP positive cells were further separated into low and high overexpression distinguished based on the fluorescent intensity of GFP. (C) qPCR analysis of SNCA mRNA expression levels in sorted cell lines in comparison with non-transduced SH-SY5Y/ReNcell VM cells, normalized to housekeeping gene RPL13A and shown here as fold change relative to the control. Dots represent three independent biological replicates, each involving separate passages, cell seeding and RNA isolations. Statistical significance was determined using an unpaired two-tailed t-test, significance indicated as p < 0.05 (), p < 0.01 (), and p < 0.001(); non-significant p > 0.05
α-syn overexpression differentially affects cell viability and reactive oxygen species production in neuronal cell models
To assess the cellular effects of α‑syn overexpression, we employed two complementary viability assays with distinct mechanistic principles: resazurin reduction and MTT metabolism (Fig. 2A, B). Although both report on cellular reducing capacity, they differ in enzymatic contributors and dynamic range. Resazurin reduction reflects the activity of a broad range of cytosolic and mitochondrial oxidoreductases, while MTT reduction has classically been attributed to mitochondrial dehydrogenase activity. As such, the assays provide partially overlapping but complementary readouts of cellular metabolic status. The resazurin assay revealed striking cell type-specific and mutation-dependent responses. SH-SY5Y cells demonstrated beneficial metabolic effects with WT α‑syn overexpression, with both low and high levels showing increased metabolic activity, reaching approximately 120–150% of control values. Conversely, the A53T variants exhibited opposite effects, displaying reduced cellular viability in both the low and high overexpression groups, indicating that the pathogenic mutation transforms α‑syn from metabolically beneficial to detrimental in this cellular context. ReNcell VM cells maintained viability levels comparable to controls across all α‑syn overexpression conditions in the resazurin assay, indicating minimal impact on overall cellular metabolic capacity regardless of variant type or overexpression level (Fig. 2A). The MTT assay provided contrasting results, highlighting the assay-specific sensitivity to α‑syn overexpression and revealing differential mitochondrial vulnerability profiles. SH-SY5Y cells showed mitochondrial resilience, with no significant alterations in MTT reduction capacity across all α‑syn variants and overexpression levels, maintaining mitochondrial function similar to controls. However, ReNcell VM cells demonstrated pronounced mitochondrial dysfunction, with reduced MTT metabolism observed across all α‑syn overexpressing populations. Both WT and A53T variants at all overexpression levels showed decreased mitochondrial metabolic capacity compared to controls, indicating that ReNcell VM cells are inherently more susceptible to α‑syn-induced mitochondrial impairment (Fig. 2B). Importantly, neither low nor high GFP expression significantly altered viability in resazurin or MTT assay in either cell line (Supplementary Fig. S1D–E), indicating that GFP alone does not account for the viability differences observed in α-syn overexpressing cells.
Fig. 2α-syn overexpression differentially affects cell viability and ROS production in neuronal cell models. The effects of α-syn (WT/A53T) overexpression on cell viability were assessed using (A) resazurin and (B) MTT assays in SH-SY5Y and ReNcell VM cell lines. Viability is presented here as a percentage relative to untransduced control cells. Group comparisons were analyzed by one-way ANOVA followed by Dunnett’s correction for multiple comparisons. (C–D) Intracellular reactive oxygen species (ROS) production was measured in α-syn (WT/A53T) overexpressing SH-SY5Y (C) and ReNcell VM (D) cells using CellROX Orange staining followed by flow cytometry revealing elevated ROS levels compared to untransduced controls. Violin plots and corresponding bar graphs summarizing mean fluorescence intensity are shown displaying the distribution/shift and mean ± SEM of CellROX fluorescence intensity. Group comparisons were performed using the Kruskal–Wallis test with Dunn’s post hoc correction for multiple comparisons. Statistical significance is indicated as p < 0.05 (), p < 0.01 (), and p < 0.01(); non-significant p > 0.05
Reactive oxygen species analysis using CellROX staining and flow cytometric quantification revealed distinct cellular oxidative stress responses to α‑syn overexpression (Fig. 2C, D). In SH-SY5Y cells, reactive oxygen species (ROS) production demonstrated a graduated mutation-dependent pattern, with substantial progressive increases from WT low overexpression through A53T high overexpression conditions. The most pronounced ROS elevation occurred in A53T high overexpressers, indicating that both mutation status and overexpression level contribute to oxidative burden in these cells (Fig. 2C). ReNcell VM cells exhibited broader sensitivity to α‑syn-induced oxidative stress, with both WT and A53T variants inducing elevated ROS production compared to controls. The oxidative stress response was most prominent in high overexpression conditions regardless of variant type. Notably, ReNcell VM cells demonstrated substantially higher ROS production responses compared to SH-SY5Y cells across comparable overexpression conditions, indicating enhanced susceptibility to α‑syn-induced oxidative stress (Fig. 2D).
To further assess cellular vulnerability associated with α-syn overexpression, we evaluated markers of apoptosis in both SH-SY5Y and ReNcell VM models. Immunoblot analysis of total and cleaved PARP revealed mild to moderate induction of PARP cleavage in some high-expression α-syn conditions compared to controls (Supplementary Fig. S2A). Given that cleavage of PARP by caspase-3 and − 7 represents a key apoptotic hallmark, we employed a complementary flow cytometric approach using CellEvent Caspase-3/7 detection. Quantification of the percentage of caspase-positive cells in the total population, as well as the mean fluorescence intensity among apoptotic cells, demonstrated a modest but statistically significant increase in caspase activity in α-syn high (especially A53T) expression groups relative to controls (Supplementary Fig. S2B–D). Notably, GFP-only expressing controls did not exhibit similar caspase activation, suggesting that the observed effects were specific to α-syn overexpression rather than attributable to the fluorescent tag alone. These findings support the presence of early-stage apoptotic stress under high α-syn load, without overt loss of viability, and align with the expected phenotypic trajectory in early Parkinson’s disease model systems.
α‑syn localization and phosphorylation are variant and dose-dependent but do not differ between the neuronal cell models
To assess the subcellular distribution and pathological modification of α‑syn in our cellular models, we performed immunocytochemistry using Syn211 antibody and phospho-specific anti‑pS129 antibody across SH‑SY5Y and ReNcell VM backgrounds, with variable overexpression levels of WT or A53T α‑syn (Fig. 3). Immunofluorescence microscopy utilizing the Syn211 antibody (Fig. 3A, B), which specifically recognizes amino acid residues 121–125 of α‑syn, validated successful overexpression of GFP-α‑syn fusion constructs across all generated cell lines. The analysis demonstrated robust colocalization between GFP fluorescence and Syn211 immunoreactivity in both SH-SY5Y and ReNcell VM cells, confirming fusion protein integrity and detectability. Control cells exhibited minimal staining with the Syn211 antibody, establishing the specificity of α‑syn detection. Quantification of Syn211 signal intensity relative to control cells confirmed a marked increase in α‑syn protein levels across all overexpressing lines (Fig. 3E). Both SH-SY5Y and ReNcell VM cell lines overexpressing WT α‑syn demonstrated a predominantly diffuse, homogeneous cytoplasmic distribution pattern independent of overexpression levels (Fig. 3A, B). Low expression WT cells displayed evenly distributed fluorescence throughout the cytoplasm. High overexpression WT variants maintained this diffuse localization pattern, exhibiting increased fluorescence intensity without formation of discrete intracellular aggregates or punctate structures. Cells overexpressing the A53T pathogenic mutation exhibited markedly distinct subcellular localization characterized by formation of prominent punctate structures. Both low and high overexpression A53T variants demonstrated cytoplasmic inclusions with aggregate-like morphology across both cell lines (Fig. 3A, B). These punctate structures appeared as discrete, intensely fluorescent foci distributed throughout the cytoplasm, readily distinguishable from the diffuse pattern observed in WT overexpressing cells. Immunofluorescence analysis utilizing pS129-specific antibodies revealed modification patterns that closely mirrored the subcellular distribution observed with Syn211 staining (Fig. 3C, D). Phosphorylated α‑syn at serine 129 represents a well-characterized pathological marker in PD and synucleinopathies. WT α‑syn overexpressing cells in both SH-SY5Y and ReNcell VM backgrounds demonstrated diffuse cytoplasmic pS129 immunoreactivity corresponding to the homogeneous distribution pattern observed with total α‑syn detection. Low overexpression WT groups exhibited modest pS129 signal intensity, while high overexpression WT variants showed proportionally increased pS129 staining intensity throughout the cytoplasm, maintaining the diffuse localization pattern without formation of discrete phosphorylated aggregates (Fig. 3A-D). Cells overexpressing A53T mutant α‑syn exhibited markedly enhanced pS129 immunoreactivity with a distinct punctate distribution pattern (Fig. 3C, D). Both low and high overexpression A53T variants demonstrated intense pS129 staining that strongly co-localized with the aggregate-like punctate structures observed in total α‑syn detection. The pS129 modification patterns remained consistent across both SH-SY5Y and ReNcell VM cell types, with no discernible differences in staining intensity or distribution between the neuronal backgrounds.
Fig. 3α‑syn localization and phosphorylation are variant- and dose-dependent but do not differ between the neuronal cell models. Immunocytochemistry staining was performed to assess the subcellular distribution and pathological modification status of WT/A53T α-syn in SH-SY5Y and ReNcell VM cell lines. Total α-syn was detected using the Syn211 antibody in (A) SH-SY5Y and (B) ReNcell VM cells that robustly colocalized with GFP, while phosphorylated α-syn at serine 129 (pS129) was visualized using phospho-specific antibody pS129 in SH-SY5Y (C) and ReNcell VM (D) cells. Overexpression-level and variant-specific differences in subcellular distribution were observed, with A53T higher overexpression associated with more intense and distinct punctate signals. Nuclei were counterstained with Hoechst. (E) Microscopic signal intensity was quantified using ImageJ software by measuring the mean fluorescence intensity within individual cells, with background subtraction applied uniformly across images. At least 10 cells per condition were analyzed, and values were normalized to the mean intensity of non-overexpressing control cells to assess relative α‑syn protein overexpression levels. Group comparisons were analyzed using one-way ANOVA with Dunnett’s correction for multiple comparisons with significance indicated asp < 0.05 (), p < 0.01 (), and p < 0.001(); non-significant p > 0.05
α-syn overexpression alters mitochondrial integrity in a variant-, dose- and cell-type dependent manner in neuronal cell models
Following our observation of cell type-specific mitochondrial dysfunction in MTT assays, we investigated the potential of the underlying mitochondrial alterations induced by α-syn overexpression using TOM20 staining and quantitative flow cytometric analysis using MitoTracker dye across generated SH-SY5Y and ReNcell VM cell line panel (Fig. 4). This analysis revealed profound cell type-specific and variant-dependent differences in mitochondrial properties between the neuronal models. SH-SY5Y cells overexpressing α-syn exhibited an altered TOM20 staining pattern compared to control cells (especially in high overexpressing population), characterized by a more compact and punctate mitochondrial appearance in representative images (Fig. 4A). This was further supported by quantification, where the increased MitoTracker signal reflected an increase in mitochondrial density (Fig. 4C. In ReNcell VM cells we did not observe morphological changes between WT α-syn overexpressing cells and control, but quantification of MitoTracker signal showed increased fluorescence intensity compared to controls in high overexpression groups (Fig. 4D). ReNcell VM cells highly overexpressing A53T mutant α-syn demonstrated analogous alterations in mitochondrial architecture as observed in SH-SY5Y cell lines and a shift toward fragmented and granular pattern (Fig. 4B). High-overexpression conditions of A53T resulted in extensive mitochondrial clustering, and loss of filamentous structure. These morphological disruptions were quantitatively confirmed by significant increases in mitochondrial signal density, especially in the A53T H group (Fig. 4D). GFP-only overexpressing controls displayed unaltered TOM20 immunofluorescence patterns (Supplementary Fig. S1 B–C), confirming that the mitochondrial phenotypes in α-syn-expressing cells are not attributable to GFP toxicity. To address the changes in mitochondrial mass, we analyzed the levels of TOM20 by Western blot (Supplementary Figure S1 C). In SH-SY5Y cells, we observed relatively weak TOM20 signal in the control group, but levels were consistently maintained across GFP only- and α-syn overexpressing lines, suggesting that α-syn overexpression did not lead to major changes in mitochondrial abundance. In ReNcell VM cells, TOM20 levels were robust and uniform across all conditions, further supporting the conclusion that mitochondrial mass remains largely unchanged under the tested conditions.
Fig. 4α-syn overexpression alters mitochondrial integrity in a variant-, dose- and cell-type dependent manner in neuronal cell models. Mitochondrial morphology and integrity were assessed in SH-SY5Y and ReNcell VM cell lines overexpressing GFP-tagged α-syn (WT/A53T). (A, B) Representative confocal images show TOM20 staining in SH-SY5Y (A) and ReNcell VM (B) cells, revealing qualitative changes in mitochondrial distribution and signal intensity across α-syn variants and overexpression levels. (C, D) Quantitative assessment of mitochondrial integrity was performed by flow cytometry following MitoTracker staining in SH-SY5Y (C) and ReNcell VM (D) cells. Data are shown as violin plots and bar graphs representing the mean fluorescence intensity of MitoTracker with Dunn’s correction for multiple comparisons. Statistical significance is indicated as p < 0.05 (), p < 0.01 (), and p < 0.001(); non-significant p > 0.05. Column graphs show the mean ± SEM as an alternative representation of staining intensity shifts
Real-time functional mitochondrial metabolism analysis reveals cell type-specific metabolic responses to different variants and dose of α-syn
To further investigate the mitochondrial functional consequences of α‑syn overexpression in SH‑SY5Y and ReNcell VM cells, we performed Seahorse XF Cell Mito Stress Test analysis, which provides dynamic profiling of OCR across multiple mitochondrial parameters (Fig. 5). In SH‑SY5Y cells, overexpression of WT α‑syn resulted in a marked increase in mitochondrial respiration relative to control cells (Fig. 5A, B). Both basal and maximal OCR values were elevated in α‑syn WT low and WT high groups, indicating an adaptive upregulation of oxidative metabolism. The α‑syn WT high group demonstrated the highest mitochondrial activity. Nevertheless, this enhanced respiratory profile was not accompanied by significantly elevated spare respiratory capacity. In contrast, SH‑SY5Y cells expressing high levels of A53T mutant α‑syn displayed a distinct pattern. Both basal and maximal OCR were reduced compared to WT-overexpressing counterparts (Fig. 5A, B). In contrast, ReNcell VM cells exhibited a consistent decline in mitochondrial function across all α‑syn overexpressing groups (Fig. 5C, D). WT α‑syn overexpression led to a significant reduction in basal OCR. Maximal respiration and spare respiratory capacity were similarly diminished, indicating compromised ability to respond to energetic demands. Notably, ReNcell VM cells expressing A53T mutant α‑syn exhibited the most profound deficits, with further reductions across all respiratory parameters, consistent with pronounced mitochondrial dysfunction. Across both WT and A53T variants, basal OCR declined in comparison to control, and spare respiratory capacity was nearly abrogated in high-expressing mutant groups.
Fig. 5. Real-time functional mitochondrial metabolism analysis reveals cell type-specific metabolic responses to different variants and doses of α-syn. Mitochondrial respiration was assessed in SH-SY5Y (A, B) and ReNcell VM (C, D) neuronal cells overexpressing (WT/A53T) α-syn using the Seahorse XF Cell Mito Stress Test. (A, C) Representative OCR traces depict mitochondrial respiratory profiles in SH-SY5Y (A) and ReNcell VM (C) cells following sequential injections of mitochondrial inhibitors and uncouplers. (B, D) Quantification of basal respiration, maximal respiration, and spare respiratory capacity are shown as bar graphs for SH-SY5Y (B) and ReNcell VM (D) cells. Group comparisons were analyzed using one-way ANOVA with Dunnett’s correction for multiple comparisons with significance indicated as p < 0.05 (), p < 0.01 (), and p < 0.001(); non-significant p > 0.05
Basal and maximal respiration rates in GFP-only overexpressing lines were comparable with untransduced controls, demonstrating that GFP expression does not impair mitochondrial oxidative capacity (Supplementary Fig. S1F). We observed subtle fluctuations in OCR parameters (maximal respiration in SH‑SY5Y GFP H and basal respiration in ReNcell VM GFP H). However, in SH‑SY5Y cells, the effect was not as pronounced as in the corresponding α-syn overexpressing lines, while in ReNcell VM cells, the trend appeared opposite to that observed in α-syn conditions.
α-syn overexpression and post-translational processing differ between SH-SY5Y and ReNcell VM cell lines
To validate the dosage-dependent overexpression and investigate biochemical processing of α-syn in our cell models, we performed immunoblotting on total protein lysates from SH-SY5Y and ReNcell VM cells stably overexpressing GFP-WT/A53T α-syn (Fig. 6). Syn211 antibody detected a clear, gradual increase in α-syn levels from control to low and high overexpression groups in both SH-SY5Y and ReNcell VM cell lines, confirming the dose-dependent overexpression of α-syn (Fig. 6A, B). In addition to the expected ~ 42 kDa band corresponding to GFP-α-syn, a second band migrating approximately 10–12 kDa higher was observed with more prominent band in ReNcell VM WT α‑syn high while in case of SH-SY5Y WT α‑syn high the lower band remained dominant. This higher molecular weight band was consistently more prominent in ReNcell VM cells compared to SH-SY5Y cells, suggesting differential processing or PTM of α-syn between the two cell types. ReNcell VM WT α‑syn high cells exhibited a significantly higher proportion of the upper band relative to SH-SY5Y WT α‑syn high cells and quantitative analysis of the upper to total band ratio further confirmed this difference (Fig. 6C). This cell type-specific difference in band pattern was not observed in the A53T overexpressing lines, where comparable bands were observed across the two cell lines. Probing with a phospho-specific antibody against pS129 revealed increased α-syn phosphorylation with increased overexpression levels in both SH-SY5Y and ReNcell VM cells (Fig. 6A, D). Together, these findings indicate that α-syn overexpression induces distinct biochemical signatures in a cell type- and overexpression level-dependent manner. Based on the difference in band patterns observed especially in the WT α‑syn high overexpressing cell lines, we hypothesized that cell type-specific post-translational processing may have a role in the observed band shift. To further investigate this, we immunoprecipitated GFP-α‑syn from SH‑SY5Y and ReNcell VM cells followed by validation of successful pulldown via immunoblotting (Fig. 6E). Both GFP-WT and GFP-A53T α‑syn were efficiently enriched, making it suitable for downstream mass spectrometry analysis (Fig. 7).
Fig. 6α-syn overexpression immunoblot profiles differ between SH-SY5Y and ReNcell VM cell lines. (A) Representative immunoblots showing GFP-tagged α-syn overexpression in SH-SY5Y and ReNcell VM cell lines. Two bands corresponding to GFP-α-syn were detected at approximately 40 kDa and 55 kDa (with Syn211 antibody). Cells with high GFP-α-syn overexpression exhibited markedly increased protein levels compared to low expressers, whereas no signal was observed in untransduced controls. Immunoblot analysis of phosphorylated α-syn at serine 129 (pS129) revealed enhanced phosphorylation in cells with higher GFP-α-syn overexpression. Bands were detected at the same molecular weights as GFP-α-syn. HSP90 (90 kDa) or GAPDH (36 kDa) served as loading control. Quantification of band intensities was performed using ImageJ and normalized to loading controls (HSP90 or GAPDH). Data are presented as mean ± SEM from independent experiments. (B) Total α‑syn levels, (C) ratio of upper α‑syn band to total α‑syn, and (D) levels of phosphorylated α‑syn (pS129) normalized to the HSP90 loading control are shown. Statistical comparison was not performed for this figure because endogenous α-syn levels in the control group were below the detection limit of our assay. As a result, quantifiable measurements were only obtained in the α-syn overexpression condition. Since the control values could not be reliably measured, statistical comparisons or multiple group analyses were not applicable. (E) Validation of immunoprecipitation efficiency of GFP-α-syn with elute being subsequently used in mass spectrometry analysis
Fig. 7. Mass spectrometry identifies NAC region–focused and cell type–specific post-translational modifications of WT but not A53T α-syn LC/MS analysis was performed on GFP-α-syn immunoprecipitated from SH-SY5Y and ReNcell VM cells. As shown in figure, sequence covered in the analysis was predominantly in NAC region (61–95) and adjacent N-terminal sites. Multiple PTMs including phosphorylation, ubiquitination, and O-GlcNAcylation were identified, with modifications concentrated mainly on threonine residues.
Mass spectrometry identifies NAC region–focused and cell type–specific post-translational modifications of WT but not A53T α-syn
To investigate the potential molecular basis underlying the distinct α-syn band patterns observed in immunoblotting, we performed mass spectrometry analysis on GFP-tagged α-syn immunoprecipitated from SH-SY5Y and ReNcell VM cells overexpressing either WT or A53T α-syn at high levels. PTMs were mapped across the α-syn sequence. We have selected the PTMs present on residues with peptides detected in both cell types, whether modified or unmodified, to ensure confident cross-sample comparison (Fig. 7). Details of overall abundance and the percentages of modified peptides are provided in the table (Table 1). Sequence coverage was concentrated within the NAC region (61–95) and adjacent N-terminal residues. Within this region, multiple PTMs were identified, including phosphorylation, ubiquitination, and O-GlcNAcylation. Notably, WT α-syn exhibited several cell type–specific differences in modification patterns across six threonine sites. T59 was phosphorylated in both SH-SY5Y and ReNcell VM cells, but was ubiquitinated exclusively in ReNcell VM. T64 was ubiquitinated in both cell lines. T72 showed phosphorylation and ubiquitination in both cell types, while O-GlcNAcylation at this site was only found in ReNcell VM. T75 was phosphorylated and O-GlcNAcylated in both cell lines, but ubiquitination at this residue was again exclusive to ReNcell VM. In contrast, T81 was phosphorylated in both cell types but ubiquitinated only in SH-SY5Y (Fig. 7). These findings reveal a shared phosphorylation profile across backgrounds, but striking divergence in ubiquitination and O-GlcNAcylation, with ReNcell VM cells displaying a broader and more complex PTM profile. The co-occurrence of multiple PTMs at single residues, particularly T72 and T75 suggests potential modification crosstalk and dynamic regulation, which may influence α-syn folding, accumulation, or interactions. Interestingly, we detected ubiquitination on threonine residues, a non-canonical modification rarely reported for α-syn. The presence of threonine ubiquitination across both cell types, with cell type–specific differences in occurrence, indicates the possibility of atypical ubiquitin linkages or non-lysine–based enzymatic targeting under overexpression conditions. In contrast to the variability seen in WT α-syn, the A53T mutant showed identical PTM profiles in both SH-SY5Y and ReNcell VM cells (Fig. 7). All detected residues were modified in the same manner regardless of background, suggesting that the A53T mutation may limit α-syn’s ability to undergo environment-dependent modifications. This loss of PTM plasticity could reflect altered accessibility or reduced interaction with modifying enzymes, potentially contributing to the mutant’s enhanced pathogenicity and reduced clearance observed in disease models. Together, these data reveal that α-syn undergoes diverse, context-dependent PTMs in its NAC and N-terminal regions, with WT protein showing greater sensitivity to the cellular environment than the A53T mutant.
Table 1α-syn PTMs detected in LC/MS analysisSH-SY5Y α-syn WT HReNCell VM α-syn WT HAmino acidPTMPeptides% of total PeptidesAmino acidPTMPeptides% of total PeptidesT59P2 of 1612.5T59P2 of 306.666667T72P6 of 2030T72P6 of 3417.64706T75P4 of 1822.22222T75P6 of 3417.64706T81P1 of 425T81P1 of 425T59Ub0 of 160T59Ub2 of 306.666667T64Ub2 of 2010T64Ub2 of 345.882353T72Ub2 of 2010T72Ub10 of 3429.41176T75Ub0 of 180T75Ub6 of 3417.64706T81Ub1 of 425T81Ub0 of 40T72O-GlcNAc0 of 250T72O-GlcNAc2 of 1414.28571T75O-GlcNAc2 of 287.142857T75O-GlcNAc2 of 1513.33333Amino acidPTMPeptides% of total PeptidesAmino acidPTMPeptides% of total PeptidesT33P2 of 2100T33P2 of 2100T59P2 of 825T59P2 of 922.22222T64P2 of 1811.11111T64P2 of 1711.76471T72P3 of 1915.78947T72P3 of 1816.66667T75P0 of 180T75P0 of 180T81P1 of 616.66667T81P1 of 616.66667S87P1 of 616.66667S87P1 of 616.66667K34Ub2 of 2100K34Ub2 of 2100K60Ub2 of 825K60Ub2 of 825T72Ub0 of 190T72Ub0 of 180T75Ub2 of 1811.11111T75Ub2 of 1811.11111T81Ub1 of 616.66667T81Ub1 of 616.66667
While multiple post-translational modifications (PTMs) were identified in both WT and A53T α-syn across cell lines, including ubiquitination, acetylation, and phosphorylation at several residues, phosphorylation at Ser129 was not detected by LC–MS/MS. This absence likely reflects the low sequence coverage of the highly acidic C-terminal domain of α-syn, where Ser129 resides, as well as the typically low abundance of this modification in unenriched samples. Importantly, the presence of pS129 was consistently confirmed by both immunofluorescence and Western blotting (Figs. 3C and D and 6 A and D), highlighting the limitations of untargeted LC–MS for low-stoichiometry phospho-sites.
Discussion
Model system design: relevance, stability, and technical advantages
Overexpression-based cellular models have been instrumental in advancing our understanding of α‑syn-mediated toxicity in PD. However, the majority of these models rely on inducible expression systems, which can introduce confounding factors such as artificial stress responses and mitochondrial dysfunction. Tetracycline- or doxycycline-based induction, in particular, has been shown to impair mitochondrial protein synthesis, reduce oxygen consumption, and promote mitochondrial fragmentation at commonly used concentrations (0.01–1 µg/mL), thereby compromising experimental outcomes [19–21]. Moreover, doxycycline exposure leads to mito-nuclear protein imbalance and shifts cellular metabolism away from oxidative phosphorylation toward glycolysis, further complicating analyses of mitochondrial and metabolic phenotypes [20, 22, 23]. These well-documented off-target effects limit the interpretability of results in studies where mitochondrial function is a key endpoint. To overcome these limitations, we established a panel of stably expressing cell lines overexpressing either WT or A53T mutant α‑syn in two neuronal backgrounds SH‑SY5Y neuroblastoma cells and ReNcell VM ventral midbrain progenitor cells. This dual-cell model enabled us to directly compare the effects of α‑syn variants, dosage, and cellular context under consistent experimental conditions. Our stable expression system provided several advantages. It maintained consistent α‑syn protein levels across low passage numbers (passages 3–5), avoiding variability associated with extended passaging or transient transfection. Although some variability in mRNA levels was observed across biological replicates, protein levels remained stable, consistent with previous reports showing a decoupling between α‑syn mRNA and protein expression [24]. This highlights the necessity of directly assessing protein levels when interpreting toxicity or functional outcomes in α‑syn models. Importantly, the expression levels achieved in our system (approximately 3–10-fold above endogenous) fall within the pathophysiological range reported in human PD and synucleinopathies, thereby enhancing the translational relevance of our findings [24]. In contrast to inducible systems, our model minimizes artificial metabolic perturbations, allowing for a clearer interpretation of the intrinsic cellular effects of α‑syn.
Given that high-level expression of GFP fusion proteins can impose cellular stress or off-target effects, we systematically evaluated GFP-only control lines to distinguish α-syn specific phenotypes. As shown in Supplementary Fig. S1, GFP expression, matched to α-syn-GFP intensities, did not significantly alter cell viability, TOM20 abundance, or overall mitochondrial morphology between the generated cell lines. While minor fluctuations in mitochondrial respiration were observed, such as elevated maximal respiration in SH-SY5Y GFP H and increased basal respiration in ReNcell VM GFP H, these effects were less pronounced and followed patterns distinct from those seen in α-syn overexpressing counterparts. These data support the conclusion that the observed mitochondrial and cellular phenotypes in our model are attributable to α-syn overexpression, rather than nonspecific effects of the GFP tag.
Comparative cell biology: cell type-specific vulnerability
A53T mutation: pathogenic effects across neural contexts
Our data revealed that the A53T mutation induces remarkably consistent toxic phenotypes across both SH-SY5Y and ReNcell VM cell models, characterized by pronounced ROS elevation, a mutation-driven shift from diffuse to punctate α-syn. High-expression A53T populations also displayed mitochondrial dysfunction, indicating that the A53T variant fundamentally alters α-syn’s mitochondrial interactions. This is in line with studies showing that A53T enhances the ability of α‑syn to assemble into oligomers that directly interact with mitochondrial membranes, disrupting normal mitochondrial function and dynamics [25]. The increased MitoTracker staining in A53T-overexpressing cells may reflect mitochondrial hyperpolarization resulting from impaired ATP synthase function or altered mitochondrial membrane permeability, representing an early pathogenic event in A53T-mediated neurotoxicity. This convergence aligns with extensive evidence documenting the robust, context-independent pathogenicity of A53T α-syn in neuronal systems [26–28]. The A53T mutation enhances oligomerization of α‑syn even at physiological expression levels [25, 29], accelerates fibril formation rates, and has been linked to autosomal dominant early-onset PD with more severe clinical presentations [30]. The mutation fundamentally alters dynamics of α-syn aggregation through enhanced membrane binding [31] and formation of toxic oligomeric intermediates [32, 33]. These oligomers demonstrate region-specific conformational differences, with inclusion-bearing regions harboring oxidized oligomers that accelerate aggregation and cause neuronal degeneration [33]. The consistent pathogenic effects observed across our cellular models support the dominant toxicity of A53T variants, which override cell type-specific protective mechanisms through gain-of-toxic-function mechanisms [32]. Interestingly, despite comparable WT α-syn overexpression levels, only A53T triggered the formation of punctate aggregates. This pattern was independent of expression level and consistently observed across both cell lines, strongly indicating that the A53T mutation possesses an intrinsic aggregation-promoting effect that fundamentally alters conformational behavior of α‑syn [32, 33]. This observation aligns with protein engineering studies showing that mutations specifically destabilize native protein states or accelerate conversion of partially folded conformations into oligomeric structures. The A53T mutation enhances oligomers and inclusion formation and membrane interactions [25, 31], creating toxic species that can disrupt cellular function even at modest expression levels where WT α‑syn protein remains diffusely distributed [34, 35]. Importantly, few studies have systematically compared A53T effects across different human neural cell types. Previous comparative work primarily examined SH-SY5Y versus PC12 cells, focusing on autophagy impairment rather than comprehensive metabolic profiling [36]. Our dual-cell system highlights the consistent pathogenic impact of the A53T mutation across distinct neuronal backgrounds, thereby enhancing the relevance and applicability of our findings to broader PD modelling.
WT α-syn: neuroprotection and context-sensitivity
The protective effects of WT α-syn overexpression observed in SH-SY5Y cells align with mounting evidence for α-syn’s physiological neuroprotective functions [37]. WT α-syn has been demonstrated to protect against oxidative damage, dopamine toxicity, and various neurotoxic insults through its co-chaperone functions and role in maintaining synaptic vesicle homeostasis [38, 39]. The enhanced mitochondrial function, increased viability, and improved bioenergetic capacity observed in our SH-SY5Y WT α-syn overexpressers likely reflect α-syn’s interaction with ATP synthase, where monomeric α-syn localizes to mitochondria and aids ATP synthase efficiency. The physiological role of monomeric α-syn in mitochondrial bioenergetics is supported by knockout studies where loss of α-syn resulted in uncoupled mitochondrial respiration and reduced ATP production [39, 40]. SH-SY5Y WT α-syn overexpressers showed 40–60% increase in basal respiration, 2–3-fold higher maximal respiratory capacity, and substantially elevated spare respiratory capacity, indicating robust mitochondrial reserve to respond to energetic demands. This mitochondrial enhancement correlates with increased cellular viability and enhanced MitoTracker staining, supporting WT α-syn’s potential neuroprotective role through direct interactions with ATP synthase or other mitochondrial components. Additionally, α-syn can rescue neurodegeneration and motor impairment resulting from cysteine-string protein-α deficiency, demonstrating its physiological neuroprotective function at synapses [38]. Our observation of elevated pS129 levels in SH-SY5Y WT high α-syn overexpressers, without associated toxicity, supports recent paradigm-shifting findings that pS129 phosphorylation occurs after initial protein deposition and serves a protective function [41]. This phosphorylation appears to represent a cellular response to manage increased protein burden rather than a pathogenic modification. Multiple lines of evidence demonstrate that pS129 inhibits α-syn fibril formation and seeded aggregation while reducing cellular toxicity [41]. Recent work has established that pS129 is physiologically triggered by neuronal activity to positively regulate synaptic transmission [42, 43], serving as an activity-dependent trigger for protein-protein interactions necessary for the function of α-syn at synapses. In vivo studies demonstrated that the non-phosphorylated form of S129 exacerbates α-syn-induced nigral pathology, whereas S129 phosphorylation eliminates α-syn-induced nigrostriatal degeneration [44]. These findings challenge older paradigms wherein pS129 was viewed solely as a pathological marker.
The contrasting toxicity observed in ReNcell VM cells suggests fundamental differences in cellular stress response mechanisms and highlights the cell type-specific nature of α-syn effects. ReNcell VM cells, derived from ventral midbrain neural progenitors, show proteomic landscapes relevant to dopaminergic neuron development and may better recapitulate the vulnerability of dopaminergic neurons in PD [45]. ReNcell VM cells exhibited consistent mitochondrial dysfunction across all α-syn variants, characterized by reduced basal respiration (20–40% decreases), decreased maximal capacity, and diminished spare respiratory capacity. This pattern of mitochondrial impairment resembles the dysfunction, respiratory collapse, and selective neuronal death observed in PD midbrain dopaminergic neurons [46], highlighting ReNcell VM as a sensitive and disease-relevant model system. The mitochondrial vulnerability in ReNcell VM cells may reflect differences in mitochondrial composition, antioxidant capacity, or protein quality control mechanisms compared to SH-SY5Y cells. This vulnerability pattern corresponds closely to observations in patient-derived neurons and human PD post-mortem brain tissue. The enhanced PTM patterns observed in ReNcell VM WT α-syn high overexpressers, evidenced by altered electrophoretic mobility, may reflect increased ubiquitination at non-canonical sites. Under proteotoxic stress conditions, cells can exhibit promiscuous ubiquitination patterns, potentially including modification at sites like T75 through non-canonical mechanisms involving E2/E3 pairs capable of serine/threonine ubiquitination. This could represent a compensatory response where cells attempt to manage protein overload through alternative degradation pathways, potentially disrupting protective modifications like O-GlcNAcylation at T75.
In line with previous reports [47, 48] linking α-syn overexpression to early cellular stress responses, our analysis revealed modest activation of apoptotic pathways, particularly in high-expression α-syn models. Cleaved PARP, a classical marker of caspase-mediated apoptosis and DNA damage response, was elevated in some conditions, and further validated by caspase-3/7 activation assessed via flow cytometry. Importantly, GFP-expressing control lines did not exhibit comparable increases, indicating that the observed apoptotic activation was not a general consequence of protein overexpression or fluorescent tagging, but rather specific to α-syn-related cellular burden. Although the magnitude of apoptotic signaling was modest and did not translate into global viability loss, these findings are consistent with subtle mitochondrial dysfunction and redox imbalance observed elsewhere in our study. This suggests that α-syn accumulation sensitizes cells to apoptotic stress even in the absence of overt cytotoxicity, reflecting a vulnerable pre-degenerative state characteristic of early Parkinson’s disease pathology. Moreover, our ability to detect such effects within a narrow, well-controlled post-sorting timeframe reinforces the sensitivity and disease relevance of our stable expression model.
Differential post-translational modification landscapes reflect cell-type-specific vulnerability
The distinct PTM profiles observed between SH‑SY5Y and ReNcell VM cells offer valuable insight into mechanisms underlying differential neuronal vulnerability. In Western blot analyses, ReNcell VM cells overexpressing WT α‑syn at high levels consistently displayed prominent upper molecular weight bands, absent in SH‑SY5Y counterparts, suggesting extensive PTMs. These shifts likely reflect a complex modification profile, including enhanced ubiquitination or phosphorylation, representing an adaptive response to increased α‑syn burden. Such modifications may serve dual roles: protective, anti-aggregation functions such as O‑GlcNAcylation and certain phosphorylation events or as markers of cellular stress that promote protein degradation, such as ubiquitination. The balance between these opposing PTM types plays a critical role in regulating α‑syn solubility, clearance, and toxicity. This evolving concept of a context-dependent “PTM code” proposes that cell-specific modification landscapes can dictate whether neurons resist α‑syn pathology or enter a pathogenic state. Ubiquitination of α-syn is a a well-documented post-translational modification, reported at multiple lysine residues (e.g., K12, K21, K23, K45, K96, K102) in both cellular models and in Lewy bodies from Parkinson’s disease brains [49, 50]. It is widely recognized to play a role in the degradation of α-syn via the ubiquitin-proteasome system and possibly in aggregate clearance via autophagy. What appears novel in our dataset is not the presence of ubiquitination itself, but rather the expression level– and cell type–specific prominence of a distinct higher molecular weight α-syn species in ReNcell VM cells overexpressing WT α-syn at high levels. Interestingly, A53T-overexpressing cells did not show comparable differences in PTM profiles between the two cell types. This uniformity further underscores the pathological dominance of the A53T variant and suggests its potential capacity to escape or bypass normal post-translational regulation.
Clinical relevance and ReNcell VM model validity
The enhanced susceptibility of ReNcell VM cells to α-syn-mediated toxicity may better recapitulate the vulnerability of dopaminergic neurons in PD. ReNcell VM cells, derived from ventral midbrain neural progenitors, represent a reproducible and easy-to-propagate cell culture system for studying neural differentiation with direct relevance to dopaminergic neuron biology [45]. These cells can differentiate into all three neural lineages (neurons, astrocytes, oligodendrocytes) and have been used for PD modeling [45, 51]. Studies using ReNcell VM cells have established functional links between PD-associated genes like PINK1 and neural degeneration, demonstrating their utility as physiologically relevant tools for studying PD mechanisms [51, 52]. The extensive PTMs observed in ReNcell VM cells mirror findings from human post-mortem brain tissue, where complex PTM patterns are associated with synucleinopathy progression [53]. The proteomic landscapes of differentiated ReNcell VM cells show characteristics relevant to dopaminergic neuron development and vulnerability [45]. The differential responses between cell types suggest that therapeutic strategies must consider cellular context. The protective effects observed in SH-SY5Y cells with WT α-syn overexpression support approaches aimed at enhancing α-syn’s physiological functions rather than simply reducing protein levels. Conversely, the vulnerability patterns in ReNcell VM cells suggest that interventions targeting protein quality control mechanisms and PTM pathways may be particularly relevant for vulnerable neuronal populations.
Conclusion
Our comparative, longitudinal analysis using a stably engineered dual-cell model establishes cell type as a critical determinant of α-syn-driven pathology, independent of simple protein abundance. These findings demonstrate that α-syn’s cellular effects are highly dependent on both genetic context (WT vs. A53T) and cellular environment (SH-SY5Y vs. ReNcell VM), with the same protein capable of conferring protection or toxicity depending on the specific neuronal context. The consistent pathogenic effects of A53T across cell types support its dominant toxicity. The differential responses to WT α-syn overexpression reveal cell type-specific vulnerability mechanisms that may explain the selective neurodegeneration patterns observed in PD. The enhanced PTMs observed in ReNcell VM cells represent stress-induced compensatory responses that ultimately contribute to cellular dysfunction, highlighting the importance of PTM landscape, mitochondrial adaptation, and cellular quality control in dictating neuronal resilience or vulnerability. Similarly, the protective metabolic effects of WT α-syn in specific cellular contexts support approaches aimed at enhancing α-syn’s physiological functions. These results reinforce the need for targeted, context-aware PD cellular modelling that accounts for neuronal population-specific vulnerabilities and provide a novel platform for further mechanistic and therapeutic investigations.
Electronic Supplementary Material
Below is the link to the electronic supplementary material.
Supplementary Material 1
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Luger AL et al (2018) Doxycycline impairs mitochondrial function and protects human glioma cells from Hypoxia-Induced cell death: implications of using Tet-Inducible systems. Int J Mol Sci 19(5). 10.3390/ijms 1905150410.3390/ijms 19051504 PMC 598370429772845 · doi ↗ · pubmed ↗
- 2Wust RCI et al (2021) The antibiotic Doxycycline impairs cardiac mitochondrial and contractile function. Int J Mol Sci 22(8). 10.3390/ijms 2208410010.3390/ijms 22084100 PMC 807136233921053 · doi ↗ · pubmed ↗
- 3Barnes J et al (2025) Soluble and insoluble lysates from the human A 53T mutant alpha-Synuclein Transgenic mouse model induces alpha-Synucleinopathy independent of injection site. Int J Mol Sci 26(13). 10.3390/ijms 2613625410.3390/ijms 26136254 PMC 1224984040650031 · doi ↗ · pubmed ↗