Genomic epidemiology of Helicobacter pylori in regions with high and low risk of gastric cancer, Colombia
Kevin Guzman, Danilo Igua, Harold Mauricio Casas Cruz, Alvaro Pazos, Arsenio Hidalgo

TL;DR
This study explores how different strains of Helicobacter pylori and environmental factors like altitude are linked to varying gastric cancer rates in Colombia.
Contribution
The study identifies two distinct H. pylori subpopulations associated with high and low gastric cancer risk in Colombia.
Findings
Gastric cancer mortality correlates positively with altitude in Nariño, Colombia.
Two H. pylori subpopulations, hspColombia_Andes and hspColombia_PacificCoast, are linked to high and low cancer risk, respectively.
The vacA and cagA genes are associated with gastric lesions and cancer risk prediction.
Abstract
Helicobacter pylori infects more than half of the world’s population and is the main risk factor for gastric cancer, although only a small percentage of those infected develop the disease. This disparity suggests the influence of bacterial, environmental, and host susceptibility factors. In Colombia, the department of Nariño presents a unique scenario: in both the Andean region and the Pacific coast, the prevalence of infection reaches 90%, but gastric cancer rates differ markedly (150/100,000 and 6/100,000 inhabitants, respectively), a phenomenon known as the “Colombian enigma.” This study analyzed gastric cancer mortality in 64 municipalities in Nariño, Colombia, using official epidemiological data and genome-based Helicobacter pylori cagA and vacA evolution and virulence. The results showed a positive correlation between altitude and gastric cancer mortality. Phylogenomically, two…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
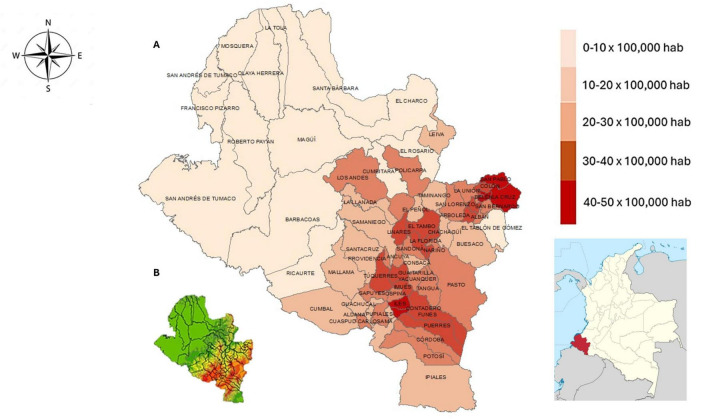 FIGURE 1
FIGURE 1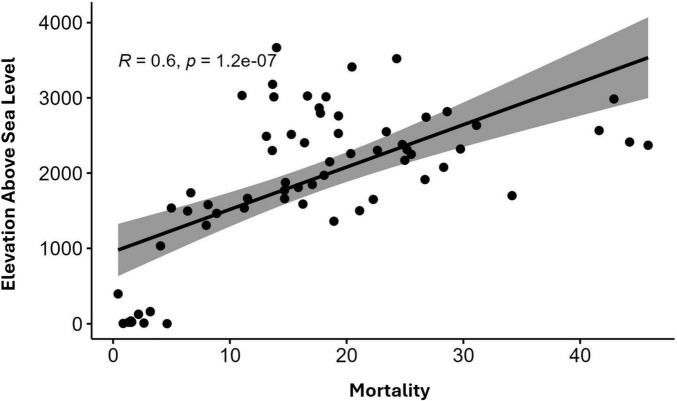 FIGURE 2
FIGURE 2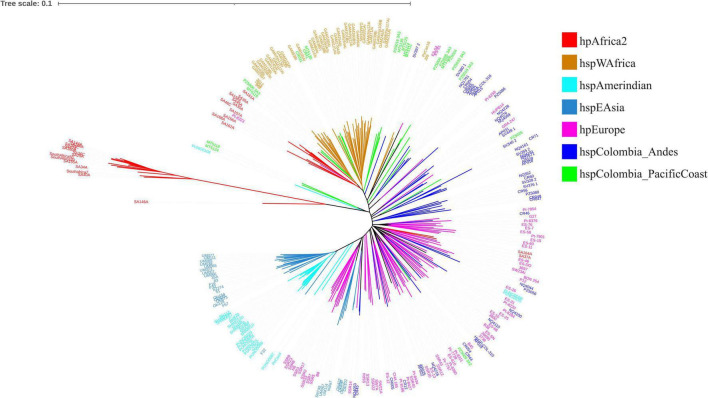 FIGURE 3
FIGURE 3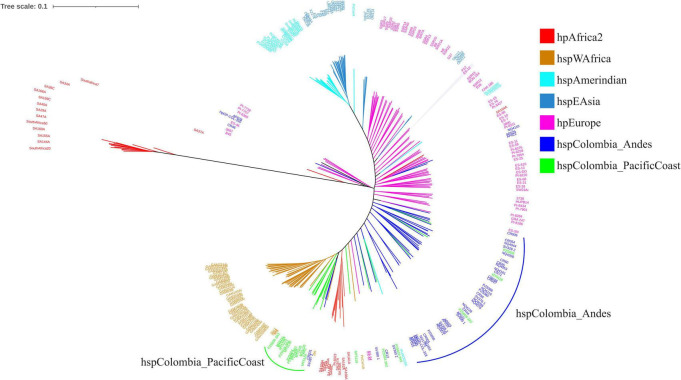 FIGURE 4
FIGURE 4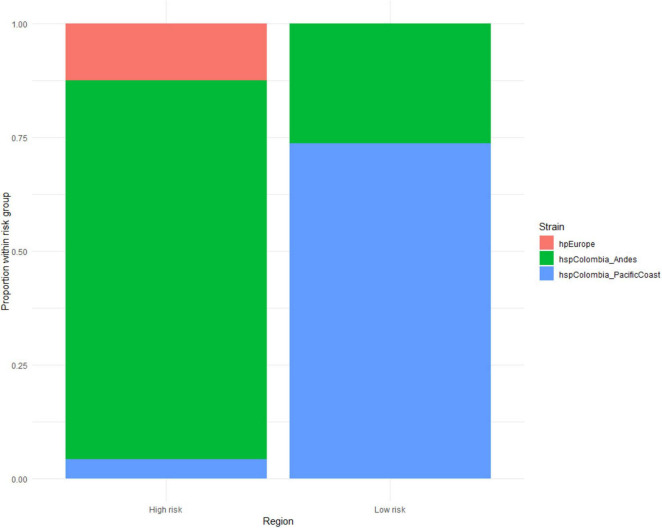 FIGURE 5
FIGURE 5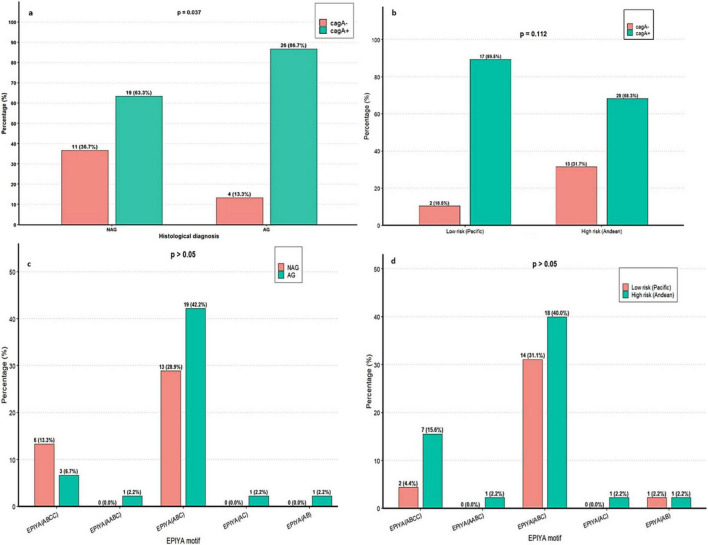 FIGURE 6
FIGURE 6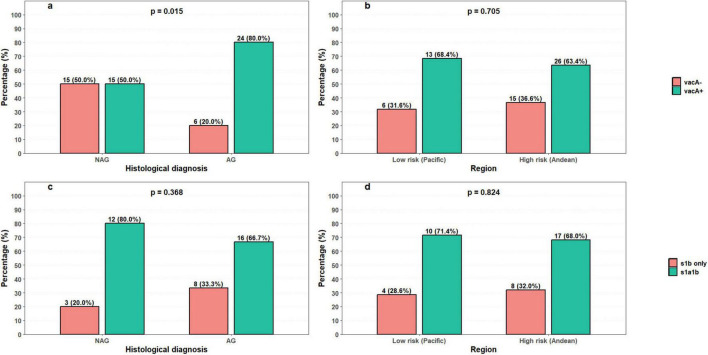 FIGURE 7
FIGURE 7| Isolated | Risk of gastric | Municipality | Isolated | Risk of gastric | Municipality |
|---|---|---|---|---|---|
| SV328_2 | High risk | Tuquerres | CR004 | High risk | Pasto |
| SV340_2 | High risk | Tuquerres | CR005 | High risk | Pasto |
| SV355_2 | High risk | Tuquerres | CR031 | High risk | Pasto |
| SV376_1 | High risk | Tuquerres | CR045 | High risk | Pasto |
| SV380_1 | High risk | Tuquerres | CR047 | High risk | Pasto |
| SV397_2 | High risk | Tuquerres | CR048 | High risk | Pasto |
| SV449_1 | High risk | Tuquerres | CR054 | High risk | Pasto |
| PZ5056 | High risk | Tuquerres | AP029 | High risk | Florida |
| PZ5080 | High risk | Tuquerres | CR44 | High risk | Pasto |
| PZ5086 | High risk | Tuquerres | CR45 | High risk | Pasto |
| HpGP-COL-310 | High risk | Florida | CR56 | High risk | Pasto |
| HpGP-COL-318 | High risk | Florida | CR60 | High risk | Pasto |
| CR12 | High risk | Florida | CR71 | High risk | Pasto |
| NQ1671 | High risk | Nariño | CR46 | High risk | Samaniego |
| NQ1701 | High risk | Nariño | PZ5005_3A3 | Low risk | Tumaco |
| NQ352 | High risk | Nariño | PZ5006_3A3 | Low risk | Tumaco |
| NQ4191 | High risk | Nariño | PZ5009_3A2 | Low risk | Tumaco |
| NQ4228 | High risk | Nariño | PZ5016_3A3 | Low risk | Tumaco |
| NQ4216 | High risk | Nariño | PZ5019_3A3 | Low risk | Tumaco |
| NQ4200 | High risk | Nariño | PZ5033_3A2 | Low risk | Tumaco |
| NQ4161 | High risk | Nariño | PZ5004 | Low risk | Tumaco |
| NQ4110 | High risk | Nariño | PZ5024 | Low risk | Tumaco |
| NQ4099 | High risk | Nariño | PZ5026 | Low risk | Tumaco |
| NQ4076 | High risk | Nariño | MT5105 | Low risk | Tumaco |
| NQ4053 | High risk | Nariño | MT5111 | Low risk | Tumaco |
| NQ4044 | High risk | Nariño | MT5114 | Low risk | Tumaco |
| AP002 | High risk | Florida | MT5118 | Low risk | Tumaco |
| AP015 | High risk | Florida | MT5119 | Low risk | Tumaco |
| AP018 | High risk | Florida | MT5124 | Low risk | Tumaco |
| AP021 | High risk | Florida | MT5125 | Low risk | Tumaco |
| AP022 | High risk | Florida | MT5135 | Low risk | Tumaco |
| AP025 | High risk | Florida | MT5136 | Low risk | Tumaco |
| AP028 | High risk | Florida | CR41 | Low risk | Barbacoas |
| AP031 | High risk | Florida | |||
| Region | Motif/Primers | Primer sequence or amino acid | Size | References |
|---|---|---|---|---|
|
| EPIYA-A | EPIYAKVNKKKAGQ | 14 aa | ( |
| EPIYA-B | EPIY (A/T) QVAKKVNAKI | 15 aa | ||
| EPIYA-C | EPIYATIDDLGGP | 13 aa | ||
| EPIYA-D | EPIYATIDFDEANQAG | 16 aa | ||
|
| Forward | CTCTCGCTTTAGTAGGAGC | 213 pb | ( |
| Reverse | CTGCTTGAATGCGCCAAAC | |||
|
| Forward | AGCGCCATACCGCAAGAG | 187pb | |
| Reverse | CTGCTTGAATGCGCCAAAC | |||
|
| Forward | GCTAACACGCCAAATGATCC | 199 pb | |
| Reverse | CTGCTTGAATGCGCCAAAC | |||
|
| Forward | GGTCAAAATGCGGTCATGG | 290 pb | ( |
| Reverse | CCATTGGTACCTGTAGAAAC | |||
|
| Forward | GGAGCCCCAGGAAACATTG | 352 pb | |
| Reverse | CATAACTAGCGCCTTGCAC | |||
|
| Forward | GTTGGGATTGGGGGAATGCCG | 426 pb | ( |
| Reverse | TTAATTTAACGCTGTTTGAAG | |||
|
| Forward | GTTGGGATTGGGGGAATGCCG | 432 pb | |
| Reverse | GATCAACGCTCTGATTTGA |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHelicobacter pylori-related gastroenterology studies · Tuberculosis Research and Epidemiology · Gastric Cancer Management and Outcomes
Introduction
Helicobacter pylori (H. pylori) is a gram-negative bacterium that infects more than half of the world’s population, with an estimated 4.4 billion people carrying it, although its prevalence in adults has been declining in recent decades (Chen Y. C. et al., 2024; Bashir and Khan, 2023) The infection is mainly acquired in childhood and persists throughout life if it is left untreated, contributing to chronic gastric pathologies and increasing the risk of precursor lesions and gastric cancer (Borka Balas et al., 2022). The discovery of H. pylori and its causal relationship with chronic gastritis and peptic ulcers was made in the 1980s by Robin Warren and Barry Marshall. For this finding, which revolutionized the treatment and understanding of H. pylori infection and stomach cancer, they received the Nobel Prize in Medicine in 2005 (Marshall, 2008; Otero-Regino, 2022).
H. pylori infection is considered the main risk factor for the development of gastric cancer, although the presence of the bacterium is necessary but not sufficient for this process to occur: only between 1 and 3% of infected individuals develop gastric cancer during their lifetime (Guzmán and Pazos, 2023; Ishaq and Nunn, 2015; Mbulaiteye et al., 2009). This low progression rate suggests the influence of dietary and environmental factors and the genetic susceptibility of the host (Acosta-Astaiza et al., 2023).
H. pylori has virulence genes, notably vacA and cagA, which have been directly associated with the pathogenesis and progression of gastric damage. The cagA gene encodes an oncoprotein that, interferes with multiple cellular regulatory pathways and causes morphological and functional changes in gastric epithelial cells, facilitating neoplastic processes (Acosta-Astaiza et al., 2023; Martínez Leyva et al., 2021; Salvatori et al., 2023). vacA produces a cytotoxin that causes cell vacuolization, apoptosis, and alters epithelial homeostasis. Some allelic variants, such as s1/m1, are linked to greater tissue damage and oncological risk (Acosta-Astaiza et al., 2023; Martínez Leyva et al., 2021; Salvatori et al., 2023). The interaction of these genetic factors (vacA and cagA) with the chronic inflammatory response promotes a sequence of lesions that begins with chronic gastritis, atrophy, intestinal metaplasia, dysplasia, and finally gastric adenocarcinoma (Piazuelo and Correa, 2013).
Currently, H. pylori is classified into eight main modern populations, based on their geographic distribution and genetic diversity: hpAfrica1, hpAfrica2, hpNEAfrica, hpEurope, hpEastAsia, hpNorthAsia, hpAsia2, and hpSahul (Moodley and Linz, 2009; Moodley et al., 2021; Yamaoka et al., 2023; Guzman K. A. et al., 2024). These populations have specific subpopulations that reflect evolutionary and migratory patterns of human populations (Moodley and Linz, 2009; Moodley et al., 2021; Yamaoka et al., 2023; Guzman K. A. et al., 2024). This phylogeographic diversity is key to understanding the evolution and adaptability of H. pylori, as well as its differential impact on gastric pathogenesis among different populations.
Understanding the genetic diversity of H. pylori and its populations is key to explaining the differences in risk in diverse regions. In this regard, the department of Nariño represents a particular case in Colombia where, despite a high prevalence of H. pylori infection in its Andean and Pacific regions (90%), the incidence of gastric cancer varies considerably between the two areas. This disparity, with high incidence rates in the Andean region (150/100,000 inhabitants) and low rates on the Pacific coast (6/100,000 inhabitants), known as the “Colombian enigma,” is an ideal model for studying the interaction between genetic and environmental bacterial factors that modulate disease risk (Guzmán and Pazos, 2023; Piazuelo and Correa, 2013). Therefore, identifying the genetic characteristics of local H. pylori populations, together with epidemiological analysis, can provide key information for the development of prevention, detection, and treatment strategies tailored to the specific needs of these populations.
Materials and methods
Mortality data
To estimate the mortality rate from gastric cancer in the department of Nariño, data provided by the Nariño Departmental Health Institute^1^ were collected and analyzed. These data included official mortality records associated with gastric cancer in the 64 municipalities of the department corresponding to the period 2011–2020.
The altitude data were obtained through the national cartography application provided by the Agustín Codazzi Geographical Institute (IGAC).^2^ Subsequently, a mortality rate map was created for the entire department using ArcGIS v3.0 software. Mortality data were georeferenced by municipality, allowing for the visualization of the spatial distribution of gastric cancer mortality in Nariño. This cartographic approach facilitated the identification of areas with higher and lower mortality rates, contributing to a detailed regional analysis.
Obtaining bioinformatic data
The initial bioinformatic data used in this study come from complete H. pylori genomes sequenced in different regions of the world, which are available in PubMLST. To identify housekeeping genes, sequences annotated in the PubMLST database^3^ (Jolley et al., 2018) were used. A total of 67 isolates were used for the department, 48 genomes from municipalities with a high risk of gastric cancer, while for the Pacific coast in the municipalities of Tumaco and Barbacoas, 19 H. pylori isolates were used (Table 1).
Multilocus sequence typing analysis
To identify the housekeeping genes atpA, efp, mutY, ppa, trpC, ureI, and yphC in the genomes, annotation was performed using the PubMLST database (see text footnote 3; Jolley et al., 2018). The concatenated sequences were aligned with Muscle (Edgar, 2004) and the phylogenetic analysis was constructed using a Neighbor-joining method, based on a T92 + G + I evolutionary model in MEGA 12 (Kumar et al., 2024). A bootstrap analysis with 1,000 replicates was performed to assess the robustness of the phylogenetic tree, which was subsequently visualized and edited in iTol^4^ (Letunic and Bork, 2024).
Phylogenomic analysis based on the core genome of Helicobacter pylori
All complete genome sequences were imported into the BIGSdb database (Jolley et al., 2018), which is dedicated to isolated bacterial genome sequences. Subsequently, a gene-by-gene alignment was performed using the CDS sequences of H. pylori strain 26695 as a reference, and the alignments were exported from the database. The matrix obtained from the genome comparator generated by BIGSdb was used to construct the phylogenetic tree using the maximum likelihood method with the GTR + G + I model in the IQ-Tree software (Nguyen et al., 2015). The resulting phylogenetic tree was visualized and edited in iTol (see text footnote 4; Letunic and Bork, 2024).
Genotyping of the cagA and vacA oncogenes of Helicobacter pylori from the department of Nariño
Of the 67 previously used H. pylori genomes, 60 complete genomes available for Nariño were downloaded from the PubMLST database (see text footnote 3) because they contained complete histopathology metadata. Of the 60 isolates, 41 originated from municipalities in the Andean region of Nariño a high gastric cancer risk area (Pasto, Túquerres and La Florida) and 19 from municipalities in the Pacific region, a low gastric cancer risk area (Tumaco and Barbacoas). Isolates were categorized according to histopathological diagnosis into two groups: of the 41 isolates from the Andean region, 20 were from patients with non-atrophic gastritis (NAG) and 21 from patients with atrophic gastritis (AG); of the 19 isolates from the Pacific region, 10 were from patients with NAG and 9 from patients with AG, yielding a total of 30 patients for each histological diagnosis.
Reference sequences for the virulence genes cagA and vacA were obtained from the consolidated annotations in the Virulence Factor Database (VFDB),^5^ using H. pylori strain 26695 as the reference genome (CagA: WP_000180747.1; VacA: WP_000405496.1) (Hatakeyama, 2017; Zhou et al., 2025). Initially, the presence or absence of each gene was assessed individually using the BLAST tool integrated in PubMLST. BLAST was run with default parameters; a gene was considered present when the alignment showed ≥ 90% identity and ≥ 85% coverage relative to the 26695 reference (Muñoz et al., 2025). For cagA-positive strains, sequences were exported in FASTA format, translated to amino acids, open reading frame (ORF) integrity was checked, and alignments were performed in MEGA 12 to locate EPIYA motifs (Kumar et al., 2024), using the amino-acid sequences presented in Table 2 for strain 26695 as reference (Rodríguez Gómez et al., 2020). For *vacA-*positive strains, allele typing was performed using the in silico PCR option integrated in PubMLST by entering the primers described in Table 2.
Statistical analysis
Statistical analysis was performed in RStudio v. 4.1.1. For the correlation analysis, given that the mortality data did not show a normal distribution (Kolmogorov-Smirnov test), correlations were performed for height above sea level and mortality using Spearman’s non-parametric coefficient, taking a value of p < 0.05 as statistically significant. To evaluate the relationship between the distribution of H. pylori strains and risk based on gastric cancer mortality data (high vs. low) for the municipalities studied, Fisher’s exact test was applied, with p < 0.05 as significant. In the analyses for the vacA and cagA oncogenes, chi-square tests were applied when the expected frequencies were ≥ 5 and Fisher’s exact test in cases with lower counts. Odds ratios (OR) and 95% confidence.
Results
Mortality rates were high in municipalities in the Andean region, ranging from 20 to 50 per 100,000 inhabitants. In contrast, municipalities on the Pacific coast had considerably lower mortality rates, ranging from 0 to 10 per 100,000 inhabitants, as shown in Figures 1A,B illustrates the altitude of the municipalities: the areas in green correspond to those located at sea level, while the colors ranging from yellow to red represent municipalities with greater altitude above sea level (Figure 1).
Spatial distribution of mortality and altitude across the 64 municipalities of the department of Nariño, Colombia. (A) Mortality map showing lower values in municipalities along the Pacific coast (0-10 per 100,000 inhabitants) and higher values in municipalities located in the Andean region (10-50 per 100,000 inhabitants). (B) Altitude map of the municipalities, where green areas represent locations at sea level, and colors ranging from yellow to red indicate municipalities situated at higher elevations above sea level.
The relationship between altitude above sea level and mortality rates in the 64 municipalities of the department of Nariño was analyzed. Spearman’s correlation analysis yielded a p < 0.05, establishing a direct correlation between altitude and mortality from gastric cancer in the department of Nariño. The finding indicates that, in the department of Nariño, the higher the altitude above sea level, the higher the mortality rate from gastric cancer. This suggests that altitude is acting as a factor associated with the risk of developing stomach cancer (Figure 2).
Correlation analysis between altitude on the y-axis and mortality on the x-axis in the 64 municipalities of the department of Nariño, Colombia.
When performing MLST analysis on H. pylori isolates from the department of Nariño, the formation of clusters corresponding to the hpAfrica2, hspWAfrica, hspAmerindian, and hspEAsia strains was observed, while isolates of European origin were grouped with both strains from the Andean region and those from the Pacific coast. Interestingly, some isolates from the Pacific coast grouped specifically with strains from West Africa. These findings demonstrate the European and African migration process that occurred 500 years ago to the American continent, bringing with it this genetic load in microorganisms, especially the hpEurope and hspWAfrica strains (Figure 3).
MLST phylogenetic tree of Helicobacter pylori isolated from the department of Nariño with isolates from other regions of the world.
When analyzing the phylogenomic tree, it was observed that the lineages presented a more defined grouping. The most ancestral group, which formed the root of the tree, corresponded to hpAfrica2, and close to this, European isolates and isolates from the Andean region of Nariño were identified, suggesting the preservation of an ancestral homology with these strains. Unlike what was evidenced by MLST, in this tree the hspAmerindian isolates were clearly differentiated from the Asian isolates. Likewise, in contrast to the MLST results, the isolates from the Andean region formed an independent lineage called hspColombia_Andes, while on the Pacific coast another distinct lineage was identified, called hspColombia_PacificCoast, which differed from the hspWAfrica isolates. In both lineages, isolates from both the Andean region and the Pacific coast were also observed to overlap, suggesting the existence of gene flow between these populations, which are only 150 km apart (Figure 4).
Phylogenomic tree of Helicobacter pylori isolates obtained from the Andean region and Pacific coast of the department of Nariño, Colombia.
In the Andean region, classified as high risk, the hspColombia_Andes population predominated (n = 40), with a lower representation of hpEurope strains (n = 6) and a marginal proportion of hspColombia_PacificCoast (n = 2). In contrast, in the Pacific coastal region, characterized by low risk, most isolates belonged to the hspColombia_PacificCoast population (n = 14), while the frequency of hspColombia_Andes strains (n = 5) was low and hpEurope was practically absent. Statistical analysis using Fisher’s exact test showed a significant association between strain distribution and geographical risk of gastric cancer (p < 0.05), suggesting that bacterial population structure plays a decisive role in the epidemiological differences observed between the two regions (Figure 5).
Relative frequencies of Helicobacter pylori strains identified in the regions studied. Each bar corresponds to a geographical region in the Andes and Pacific Coast, while the colors indicate the proportional distribution of the different lineages.
In the analysis of vacA and cagA, cagA was detected in 45 strains overall (75%; 45/60). Regarding histopathological diagnosis, cagA was detected in 86.7% (26/30) of strains isolated from AG, compared with 63.3% (19/30) of strains from NAG (Figure 6a). This association was statistically significant (p = 0.037; OR = 3.76; 95% CI, 1.04–13.60), indicating that the odds of strains from AG being cagA-positive were 3.76 times the odds for strains from NAG. By region, cagA prevalence was higher in the low-risk region (89.5%; 17/19) than in the high-risk region (68.3%; 28/41), although this difference did not reach statistical significance (Figure 6b).
Association of cagA and EPIYA motifs with histological diagnosis and geographic region. (a) Presence of cagA by histological diagnosis. (b) Presence of cagA by region. (c) Distribution of EPIYA motifs by histological diagnosis. (d) Distribution of EPIYA motifs by region.
In the 45 cagA-positive strains, the EPIYA (ABC) pattern was the most frequent in both groups: 42.2% in AG (19/45) and 28.9% in NAG (13/45), with no significant differences (Figure 6c). At the regional level, this pattern also predominated in both the low-risk region (31.1%; 14/45) and the high-risk region (40.0%; 18/45), with no statistical significance (Figure 6d). Although no statistically relevant differences were identified, the distribution suggests possible variations according to lesion type and region.
The vacA gene was detected in 39 strains overall (65%; 39/60). In strains associated with AG, vacA was detected in 80.0% of isolates (24/30), compared with 50.0% (15/30) in NAG. This association was statistically significant (p = 0.015; OR = 4.0; 95% CI, 1.27–12.6), indicating that the odds of strains from AG being vacA-positive were 4.0 times the odds for strains from NAG (Figure 7a). By region, vacA was detected in 68.4% (13/19) of strains from low-risk areas and 63.4% (26/41) of strains from high-risk areas, with no significant association (Figure 7b).
Association of the vacA gene and its allelic variants with histological diagnosis and geographic region. (a) Presence of vacA by histological diagnosis. (b) Presence of vacA by region. (c) Distribution of vacA alleles by histological diagnosis. (d) Distribution of vacA alleles by region.
In allele characterization of the vacA-positive strains, the m1 and i1 regions were detected in all isolates, while the s1a1b combination was observed in 66.7% (16/24) of strains from AG and 80.0% (12/15) of strains from NAG, with no significant differences (Figure 7c). Similarly, the s1a1b allele combination was more frequent in the low-risk region (71.4%; 10/14) than in the high-risk region (68.0%; 17/25), although without statistical differences (Figure 7d).
Discussion
H. pylori is one of the most ancient microorganisms associated with Homo sapiens. Its infection is estimated to have occurred approximately 130,000 years ago, and for the past 60,000 years, it has accompanied humans from East Africa, giving rise to different bacterial populations in each settlement (Moodley et al., 2012; Linz et al., 2007). Only about 500 years ago, the arrival of Europeans and Africans to the American continent introduced a new set of pathogens, including H. pylori. The encounter between the hspAmerindian, hpEurope, hpAfrica2, and hspWAfrica strains gave rise to new, independent lineages of H. pylori in the Americas (Thorell et al., 2017; Guzman et al., 2023; Guzman K. A. et al., 2024).
The two new H. pylori subpopulations suggest that the bacteria are undergoing a constant process of evolution and adaptation to the new gastric environment, the host’s immune response, and the geographical conditions in which these populations are found (Thorell et al., 2017; Guzman et al., 2023; Guzman K. A. et al., 2024). In the Andes region, which has a higher risk, most isolates corresponded to a predominant local population, hspColombia_Andes, while European-origin strains and those typical of the Pacific coast appeared in much smaller proportions. In contrast, in the Pacific region, classified as low risk, the opposite pattern was observed, with a strong representation of the area’s native population (hspColombia_PacificCoast) and a low presence of isolates characteristic of the Andes, while European strains were practically absent. Statistical analysis confirmed that this differential distribution is not random but is significantly associated with the geographical risk of gastric cancer (p < 0.05). Likewise, the finding of strains with genetic overlap in both areas suggests the existence of population exchange facilitated by geographical proximity, which maintains a dynamic and diverse gene pool within the species.
These results contrast with previous MLST studies, which found that strains from the Pacific coast were of African and European origin, while in the Andean region, particularly in Túquerres, the strains were of European origin. This coevolutionary mismatch was identified as the cause of gastric lesions and gastric cancer in the Andean region, whereas on the Pacific coast, the explanation for the low incidence and mortality from gastric cancer was related to the “African enigma,” associated with the host–pathogen coevolutionary process (de Sablet et al., 2011; Kodaman et al., 2014).
The remarkable difference in gastric cancer mortality rates between the Andean region (high mortality) and the Pacific coast (low mortality), despite the high prevalence of infection in both areas (90%) (Guzmán and Pazos, 2023), confirms the well-known “Colombian enigma.” This disparity reinforces the hypothesis that the mere presence of H. pylori is not sufficient to explain cancer development and that environmental factors such as altitude play an important role, as evidenced by the statistically significant correlation between altitude and mortality. Altitude may influence physiological mechanisms, exposure to different environmental factors, or lifestyle patterns that, in turn, modulate cancer risk.
Moreover, the analyses show that cagA and vacA are significantly associated with atrophic gastritis compared to non-atrophic gastritis; this finding is consistent with previous studies reporting an association between cagA/vacA genotypes and preneoplastic lesions in Andean populations (Guzmán J. et al., 2024; Bustos-Fraga et al., 2023; Takahashi-Kanemitsu et al., 2020). The cagA oncogene modulates the immune response and innate inflammatory pathways by influencing mitochondrial integrity and the inflammasome. Recent studies have shown that cagA can activate the NLRP3 inflammasome, increasing ROS, IL-1β/IL-18, and promoting migration and invasion (Chen D. et al., 2024; Zhang et al., 2022). Similarly, in adaptive immunity, cagA promotes evasion of effector T cell responses by increasing PD-L1 levels in gastric cell-derived exosomes, inhibiting CD8 + proliferation and function, thereby promoting local tolerance and gastric lesion progression (Wang et al., 2023). However, the relationship between the mere presence of cagA and clinical risk is nuanced by that human factors such as ancestry, diet, high salt consumption, and evolutionary lineage with H. pylori modulate the likelihood of progression to severe lesions (Guzmán and Pazos, 2023).
For its part, vacA induces and dysregulates mitophagy and autophagy, causes alterations in lysosomal trafficking, and can prevent the lysosomal degradation of bacterial and cellular proteins (Wang et al., 2022; Zhu et al., 2017), facilitating the intracellular accumulation of cagA and thus amplifying its oncopathogenic effects, a synergistic effect in which it enhances the activity and persistence of cagA in epithelial cells, amplifying chronic inflammation and precancerous damage (Abdullah et al., 2019). In addition, it can actively interfere with the maturation and function of antigen-presenting cells or directly with T cells by suppressing IL-23 and inducing IL-10/*TGF-*β, promoting Treg differentiation and a tolerogenic microenvironment that weakens the adaptive response (Altobelli et al., 2019).
In contrast, no significant association was detected between regional origin and the overall frequency of these genes, a finding consistent with studies that found no consistent geographical differences between areas of proven risk in Colombia (Trujillo et al., 2014), although it contrasts with multiple studies reporting that H. pylori strains from regions of Colombia with high rates of gastric cancer express higher levels of cagA and vacA than strains from regions with lower rates of gastric cancer (Bravo et al., 2002; Sicinschi et al., 2010; Loh et al., 2011). The absence of an association between these virulence genes and geographic region may be explained by the phylogenetic overlap between strains classified as hspColombia_Andes and hspColombia_PacificCoast, which is indicative of gene flow across regions. Such admixture is consistent with coinfection or multiple colonization of distinct gastric niches by strains of different ancestry within the same host, a dynamic that can disrupt the expected geographic distribution of these genes (de Sablet et al., 2011; Guzmán and Pazos, 2023). Moreover, host susceptibility factors, particularly polymorphisms or mutations in cytokine genes such as IL-1B and IL-10, can increase an individual’s predisposition to severe gastric disease irrespective of geographic origin (Camargo et al., 2006).
The molecular characterization of the predominance of the EPIYA-C motif in both strains associated with atrophic gastritis and those from the high-risk region is consistent with recent descriptions of Colombian isolates that document ABC patterns as frequent in the region (Muñoz et al., 2025; Rodríguez Gómez et al., 2020); At the same time, other studies have linked a higher number of EPIYA-C repeats with greater oncogenic capacity in cell assays and with more severe histology (Quiroga et al., 2010), and a recent study in Latin America showed that microvariants in the cag pathogenicity island may nuance the association between cagA and disease, suggesting that the presence of EPIYA-C alone may not be sufficient to increase risk in the absence of additional repeats (Rizzato et al., 2020).
The presence of the vacA m1 and i1 alleles variants associated with increased cytotoxicity in all vacA-positive isolates, together with the predominance of the s1a1b combination, yet the absence of a significant association with lesion type or geographic origin, may be explained by high local genetic diversity that facilitates coinfection by strains of distinct ancestry. When combined with H. pylori’s elevated rate of homologous recombination via horizontal gene transfer, this diversity can promote the formation of genetic mosaics (mixed s1a1b genotypes) and favor the emergence and persistence of more virulent allele combinations (m1i1), thereby enhancing bacterial fitness. Collectively, these processes may modulate clinical outcomes (Akeel et al., 2019; García-Ortíz et al., 2011; Gutiérrez-Escobar et al., 2019; Wroblewski et al., 2010).
Furthermore, the development of gastric cancer in Colombian populations is determined by a complex interaction between helminth co-infection, immune response, environmental factors, diet, and proximity to volcanoes. H. pylori infection induces a proinflammatory Th1-type immune response characterized by the production of cytokines such as IL-10-1082AG, which promote inflammation and gastric damag (Piazuelo et al., 2008; Correa, 2013; Torres et al., 2013). However, helminth co-infection can modulate this response, shifting it toward an anti-inflammatory Th2 profile, thereby reducing tissue damage and potentially decreasing cancer risk (Piazuelo et al., 2008). From an environmental perspective, altitude and exposure to volcanic particles in the Andean region generate additional pro-oxidative and inflammatory conditions (Correa, 2013; Torres et al., 2013). Added to this is a diet predominantly based on potatoes and beans, with low consumption of fruits and vegetables, which promotes chronic gastric inflammation (Piazuelo et al., 2008). This scenario contrasts with that of the Pacific coast, where the diet is more varied and rich in fruits, vegetables, and seafood, which is associated with lower gastric cancer mortality (Guzmán and Pazos, 2023; Torres et al., 2013).
This study presents several limitations that should be considered when interpreting the results. First, although 67 complete Helicobacter pylori genomes were included, sequence availability depended on public databases, which may have introduced selection bias and limited the representativeness of certain geographic areas. In addition, the number of isolates per municipality was not homogeneous, particularly in low-risk regions, which could affect the robustness of comparative analyses between bacterial subpopulations. The histopathological information used was obtained from records associated with the available genomes, making it impossible to standardize and verify the clinical conditions across all patients.
Conclusion
H. pylori is a continuously evolving microorganism whose coevolution with humans has given rise to new subpopulations, including hspColombia_Andes and hspColombia_PacificCoast. This genetic diversity is associated with the marked difference in gastric cancer mortality between the Andean region and the Pacific coast, reinforcing the role of bacterial population structure.
The pronounced difference in gastric cancer mortality rates between the Andean and Pacific regions, despite the high prevalence of infection in both, supports the so-called “Colombian enigma” and demonstrates that the presence of H. pylori alone is not sufficient to explain carcinogenesis. Environmental factors such as altitude play a decisive role in modulating gastric cancer risk.
The analysis of the vacA and cagA oncogenes confirms their association with the progression of gastric lesions, positioning them as potential genetic biomarkers for predicting progression to more advanced stages and the risk of gastric cancer. Although vacA alleles and the EPIYA motifs of the cagA gene showed no relationship with lesion severity in this study, the sample size represents a limitation that could be addressed in future research.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abdullah M. Greenfield L. K. Bronte-Tinkew D. Capurro M. I. Rizzuti D. Jones N. L. (2019). Vac A promotes Cag A accumulation in gastric epithelial cells during Helicobacter pylori infection. Sci. Rep. 9:38. 10.1038/s 41598-018-37095-4 30631092 PMC 6328614 · doi ↗ · pubmed ↗
- 2Acosta-Astaiza C. López-Sandoval A. Bonilla-Chaves J. Valdes-Valdes A. Romo-Romero W. (2023). Helicobacter pylori virulence genotypes and their relationship with precursor lesions of gastric malignancy and histological parameters in infected patients in Colombia. Rev. Peru Med. Exp. Salud Publica 40 348–353. 10.17843/rpmesp.2023.403.12858 37991039 PMC 10953661 · doi ↗ · pubmed ↗
- 3Akeel M. Shehata A. Elhafey A. Elmakki E. Aboshouk T. Ageely H. (2019). Helicobacter pylori vac A, cag A and ice A genotypes in dyspeptic patients from the southwestern region of Saudi Arabia: distribution and association with clinical outcomes and histopathological changes. BMC Gastroenterol. 19:16. 10.1186/s 12876-019-0934-z 30683054 PMC 6346553 · doi ↗ · pubmed ↗
- 4Altobelli A. Bauer M. Velez K. Cover T. L. Müller A. (2019). Helicobacter pylori Vac A targets myeloid cells in the gastric lamina propria to promote peripherally induced regulatory T-cell differentiation and persistent infection. m Bio 10 e 261–e 219. 10.1128/m Bio.00261-19 30890606 PMC 6426600 · doi ↗ · pubmed ↗
- 5Atherton J. C. (1998). H. pylori virulence factors. Br. Med. Bull. 54 105–120. 10.1093/oxfordjournals.bmb.a 011662 9604436 · doi ↗ · pubmed ↗
- 6Bashir S. K. Khan M. B. (2023). Overview of Helicobacter pylori infection, prevalence, risk factors, and its prevention. Adv. Gut Microbiome Res. 2023:9747027. 10.1155/2023/9747027 · doi ↗
- 7Borka Balas R. MeliţL. E. Mărginean C. O. (2022). Worldwide prevalence and risk factors of Helicobacter pylori infection in children. Children 9:1359. 10.3390/children 9091359 36138669 PMC 9498111 · doi ↗ · pubmed ↗
- 8Bravo L. E. van Doom L. J. Realpe J. L. Correa P. (2002). Virulence-associated genotypes of Helicobacter pylori: Do they explain the African enigma? Am J Gastroenterol. 97 2839–2842. 10.1111/j.1572-0241.2002.07031.x 12425557 · doi ↗ · pubmed ↗