Distribution of falciparum and non-falciparum malaria among symptomatic malaria patients in Dschang, West Region of Cameroon
Pacome V. K. Tchuenkam, Samuel J. White, Varun Potlapalli, Eva M. Keming, Meredith S. Muller, Darlin B. N. Kaunda, Oksana Kharabora, Rhoel R. Dinglasan, Jonathan B. Parr, Christopher B. Tume, Jessica T. Lin, Jonathan J. Juliano, Innocent M. Ali

TL;DR
This study found that both falciparum and non-falciparum malaria are common in Dschang, Cameroon, with mixed infections being frequent.
Contribution
The study provides updated prevalence data on all Plasmodium species, including P. ovale subspecies, in the West Region of Cameroon.
Findings
P. falciparum was the most common species, but non-falciparum malaria was also frequently detected.
Mixed infections with multiple Plasmodium species were found in 8.9% of cases.
Both P. ovale curtisi and P. ovale wallikeri subspecies were identified in the region.
Abstract
Malaria is a vector-borne parasitic disease that continues to be a global public health threat. Five different species of the genus Plasmodium (P. falciparum, P. malariae, P. ovale curtisi, P. ovale wallikeri, and P. vivax) cause malaria in Sub-Saharan Africa. Previous cross-sectional surveys from 2013 and 2017 indicated the circulation of P. vivax in the West region of Cameroon, prompting an investigation into the prevalence of all falciparum and non-falciparum malaria parasite species in this region. A cross-sectional facility-based study targeting both adult and children in which we recruited 431 clinically suspected cases of malaria from three health centres in the West region of Cameroon in 2020. Socio-demographic, clinical data, and dried blood spots (DBS) were collected from all consenting patients. Parasite DNA was extracted from DBS for real-time PCR amplification of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
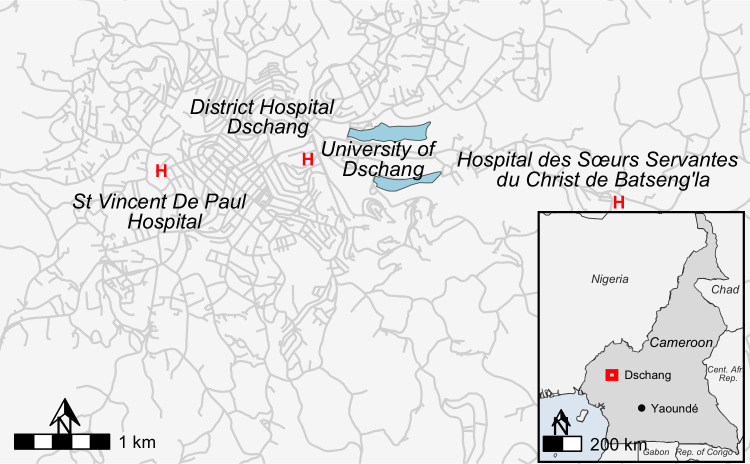 Fig 1
Fig 1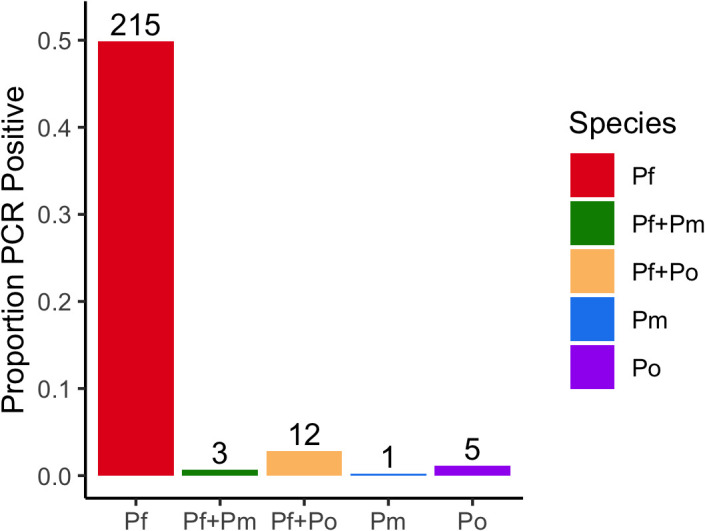 Fig 2
Fig 2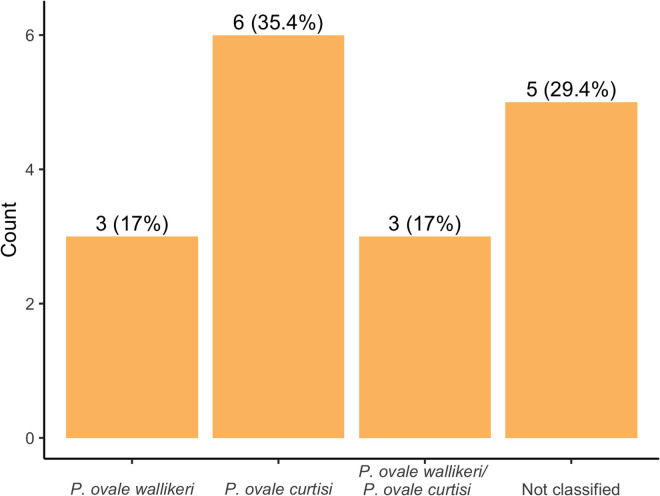 Fig 3
Fig 3- —http://dx.doi.org/10.13039/100006492Division of Intramural Research, National Institute of Allergy and Infectious Diseases
- —http://dx.doi.org/10.13039/100006492Division of Intramural Research, National Institute of Allergy and Infectious Diseases
- —http://dx.doi.org/10.13039/100006492Division of Intramural Research, National Institute of Allergy and Infectious Diseases
- —http://dx.doi.org/10.13039/100006492Division of Intramural Research, National Institute of Allergy and Infectious Diseases
- —http://dx.doi.org/10.13039/100006492Division of Intramural Research, National Institute of Allergy and Infectious Diseases
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMalaria Research and Control · Digital Imaging for Blood Diseases · Parasitic Diseases Research and Treatment
Introduction
Malaria is an infectious disease caused by parasites of the genus Plasmodium and transmitted by infected female Anopheles mosquitoes in tropical and subtropical countries [1]. Despite substantial control efforts, it remains one of the deadliest diseases in the world, with Africa suffering more than 95% of the burden. Several strategies are implemented across Africa to control malaria, including chemoprophylaxis, vector control, and appropriate diagnosis and treatment [2]. High quality malaria surveillance is a key part of the WHO malaria containment strategy and is an important piece to investigate the impact of the disease on the health status of the population [3]. Surveys addressing malaria diagnosis should be used as optimal control measures to determine the parasite species infecting individuals as treatment varies from one species to another [3]. To date, six species of malaria have been identified that cause disease in humans, Plasmodium falciparum, P. malariae, P. vivax, P. knowlesi, P. ovale wallikeri and P. ovale curtisi, of which 5 occur in Africa. P. falciparum and P. vivax are the most widely distributed world-wide with P. falciparum accounting for over 95% of the world’s cases and 99% of deaths from malaria. However, non-falciparum malaria is increasingly recognized as sources of clinical morbidity and mortality [4].
Historically, it was thought that P. vivax was largely absent from West and Central Africa due to a majority of the population of sub-Saharan Africa lacking the Duffy Antigen Receptor for Chemokines (DARC), the primary receptor for human red blood cell invasion [5]. However, recent findings have reported an apparent increase in non-falciparum malarias in Cameroon, [6–8] and other regions of Africa [9–11]. A study in Cameroon from 2017 found that 38.6% of all malaria cases in Dschang were P. vivax infections [6]. This is almost twice (20.4%) the prevalence that Ngassa et al. reported in 2016 in Douala and far greater (4%) than reported by Fru-cho in 2014 [12]. This inconsistency in the burden of P. vivax highlights the need to further characterise this species in West and Central Africa.
P. ovale is often detected as a mixed species infection in areas with high malaria transmission intensity, including Cameroon [13,14]. The low parasite counts, and mixed infections commonly result in misdiagnosis of P. ovale malaria, and its morphological similarities with P. vivax make microscopic identification challenging [15]. Adding to the difficulty in species identification, there are two morphologically indistinguishable but genetically distinct species of P. ovale; Plasmodium ovale curtisi and Plasmodium ovale wallikeri [16]. In Cameroon, P. ovale curtisi was first reported in Douala by Foko et al in 2021 [17] in mixed infections with P. falciparum, but no occurrence of P. ovale wallikeri has been reported. According to Antonio-Nkondjio and colleagues, P. malariae represent 1% of infection cases in Cameroon by microscopy [18]. This likely underrepresents the true burden as recent studies using molecular tools indicated 17% out of 236 blood samples analysed [19] in a forest area and close to 20% (both in mono and mixed infections) in the Adamaoua region contained P. malariae [4]. These studies suggest the need to deploy molecular diagnostics to improve non-falciparum species detection [18].
Cameroon primarily uses rapid diagnostic tests (RDT) that only detect P. falciparum antigens, providing no data on the burden of non-falciparum malaria [20,21]. Diagnoses of non-falciparum infections are difficult, especially as current RDTs do not reliably or specifically detect P. malariae, P. ovale spp., and P. vivax. Also, the life cycles of P. ovale spp. and P. vivax include dormant liver stages, called hypnozoites, that can cause relapse weeks to months after primary infection and are not detectable by current diagnostics. With the aim to strengthen control measures and design specific tools to combat malaria, the main objective of this work was to characterise the burden of non-falciparum malaria in Dschang and surrounding neighbourhoods by leveraging sensitive molecular diagnostics.
Methods
Ethics statement
The study was reviewed and approved by the institutional review board (IRB) of the Cameroon Baptist Convention Health Board (FWA00002077), Protocol IRB2019−40. Written informed consent was administered in French or English based on participant preference, via an independent translator (who also spoke the local language Yemba) as needed. For children, informed consent was obtained from a parent or guardian. Humans Subjects Research-exempt, de-identified dried blood spots were sent to the University of North Carolina for molecular testing. The study complied with the declaration of Helsinki.
Study site and study design
The Dschang Health District encompasses 22 health areas and covers 1060 km^2^. It is a tropical, semi-urban environment at an elevation of 1380-1400m above sea level with a rainy season that occurs between mid-March and mid-October. This study was a prospective hospital-based cross-sectional survey in the three main health facilities in the Dschang Health District in the West region of Cameroon: Dschang District Hospital, St. Vincent Catholic Hospital, and Hopital des Soeur Servante du Christ de Batseng’la (Fig 1). Sample collection was undertaken from June 12^th^ to September 8^th^, 2020, following a random convenience sample of individuals willing to participate. This study was designed to characterise the species of parasites causing clinical malaria in this region of Cameroon.
Clinical sites in Dschang, Cameroon.Country border shape files available from geoBoundaries at William and Mary (https://www.geoboundaries.org/index.html) available through CC BY 4.0 license. Highway and bodies of water shape files from Humanitarian OpenStreetMap https://www.hotosm.org/ available through Open Database License.
Patient enrolment and Sample collection
In total, 431 patients were enrolled (Table 1). The inclusion criteria of the study included female and male of all ages with fever (axillary temperature 37.5 °C or self-reported history of fever) in past 24 hours without observable signs and symptoms suggestive of severe malaria. These signs included the history of vomiting, multiple convulsions, or impaired consciousness [22]. All those who signed the consent form were enrolled in the study. Participants were excluded if they had used (self-report) anti-malarial medicine in the past 14 days as mRDTs may remain positive after therapy due to persistent antigenemia. Dried blood spots (DBS) were prepared using left over blood samples by spotting 3–4 drops on a Whatman filter paper (N°03) and left to dry overnight away from sunlight. Samples were then sealed in a zip lock bag with a desiccant and stored at −20°C until DNA extraction.
Table 1: Presentation of socio-demographic data with respect to malaria outcome.
Detection of malaria parasites in clinical samples
One 6-mm whole punch of each DBS was used for molecular testing and multiple blank DBS punches were done after each sample to prevent contamination. Parasite DNA was extracted from DBS punch using Chelex®-100 (Bio-Rad Laboratories, CA, USA), extraction [23]. Malaria positivity was assessed using quantitative real-time PCR (qPCR) for P. falciparum, P. vivax, P. ovale spp., and P. malariae using species-specific 18S rRNA gene fragment probes (S1 Table). All plates contained negative controls and positive controls consisting of mocked blood spots containing the desired target (MRA 177 178 179 and 180, BEI Resources, Manassas, VA). Non template control consisting of molecular grade water was used to monitor for contamination. Samples were tested only once. All reactions were carried out on a Bio-Rad CFX Connect Real-Time PCR Detection System. ROCHE FastStart Universal Probe Master mix (catalog #4913957001) was used alongside published primer sequences. Each reaction had a total volume of 10µl and was run for 45 cycles (S1 Table) and the limit of detection for each species detection was a cycle threshold (Ct) < 44 cycles.
Detection of P. ovale wallikeri and P. ovale curtisi
A combination of both nested PCR (nPCR) and qPCR was used for the detection of P. ovale spp. as described by Potlapalli et al. [24]. Briefly, the nPCR was performed using the specific primers rPLU1 and rPLU5 targeting a 1200 bp gene fragment for nest 1 and the combination ROVA1 and ROVA2 for P. ovale curtisi; ROVA1v and ROVA2v for P.ovale wallikeri. For the qPCR, a modified protocol from Perandin et al. [25] and Calderaro et al, [26] targeting the small unit of the 18S rRNA was used. The reaction was carried out using Fast Start Universal Probe Master mix (ROX, Roche) and published primer and probe concentrations on a Bio-Rad CFX Connect Real-Time PCR Detection System. Both assays were run in parallel to 50 cycles, instead of the originally published 45 and 55 cycles, and the limit of detection was fixed at Ct < 49. For P. ovale curtisi amplification, OVA-F (TTTTGAAGAATACATTAGGATACAATTAATG) and OVA-R (CATCGTTCCTCTAAGAAGCTTTACAAT) were used along with OVA (VIC) probe (CCTTTTCCCTATTCTACTTAATTCGCAATTCATG). For P. ovale wallikeri amplification, OVA-Fv (TTTTGAAG AATATATTAGGATACATTATAG) and OVA-R were used along with Ovav (FAM) probe (CCTTTTCCCTTTTCTACTTAATTCGCTATTATG). A common annealing temperature of 52.8 °C was chosen as this yielded similar Ct for detection of the target plasmid copy concentrations [24].
Statistical analysis
Clinical and socio-demographic data were analysed with Chi-square analysis for categorical variables. A Chi-square test was conducted to compare categorical variables to the outcome of interest. Fisher’s exact test was conducted for variables with less than 5 observations when appropriate. Bivariate associations between Plasmodium infection and socio-demographic data were investigated using R (4.4.1).
Results
Socio-demographic characteristics and clinical parameters
Of the total participants enrolled**,** 59.3% (255/431) were from the District Hospital Dschang (HDD) and the rest enrolled from Hopital des Soeur servant du Christ de Batsengla (HSSCB, n = 106 (24.6%)) and Hopital Saint Vincent (HSV, n = 70 (16.2%)). Table 1 summarizes the key socio-demographic characteristics of the study population and association with Plasmodium infection status. In the study population, 58.1% (253/431) were female while 15/253 (5.9%) of the women were pregnant at the time of enrolment. The median age was 26 years (IQR: 20.7–31.3), and children less than 10 years represented 10% of the total study population. Close to 63% of the study population had either secondary or tertiary education. With regards to occupation, only 9% had a job in the formal sector. The majority were students (61%) as Dschang is a university town. While 86% reported travel out of the region (within Cameroon) in the past year at least once, most participants reported to have been in Dschang in the previous three months (77.9%). Most of the patients (82.3%) did not have a water source within 1 km of their homes. About 10% of study participants reported keeping domestic animals at home. More than half of the study population (54.5%) reportedly owned bed nets. About the same proportion of all study participants reported sleeping under a bed net the previous night (55.2%). Of these socio-demographic data, only sex trended toward an association with malaria positivity, with males more likely than females to test malaria positive with a p-value of 0.06. Multivariate logistic regression testing age, sex and level of education to malaria infection did not show more significance with all p-value above 0.05 (S2 Table)
Table 2 shows the symptoms at presentation. Among the 431 participants enrolled in this study, 311 reported symptoms beyond fever or history of fever and the most commons symptoms were abdominal pain or diarrhoea (35.7%), headache (29.5%), and general fatigue (28.3%). A minority of patients had cough, gastric pain, or other symptoms.
Table 2: Symptoms other than fever reported by participants.
Frequency of Plasmodium species, distribution, and associated symptomatology
Of the 431 patients enrolled, we observed an overall malaria prevalence by PCR of 54.8% [95% CI: 50.1–59.8] (236/431). Infection status of malaria-infected participants is summarized in Fig 2. Of these 431, we observed that 230/431 (53.36% [95% CI: 48.7–58.5]) were positive for P. falciparum, 17/431 (4.0% [95% CI: 2.2–5.6]) were positive for Plasmodium ovale spp. and 4/431 (0.9% [95% CI: 0.2–1.7]) were infected with P. malariae. No cases of Plasmodium vivax were identified. The remaining 195 of 431 (45.2% [95% CI: 40.6–50.3]) febrile illness samples were negative for any Plasmodium species by PCR. Mixed infections were common. Of all malaria-positive individuals (n = 236), 8.9% [95% CI: 5.9–12.6] (21/236) contained at least one non-falciparum species, with 7.2% [95% CI: 4.2–10.8] (17/236) and 1.7% [95% CI: 0.0–5.3] (4/236) containing P. ovale spp. and P. malariae, respectively. We observed that among 17 samples that were positive for P. ovale, 5/17 were mono-infections (29% [95% CI: 11.8–50.9]) and 12/17 (71% [95% CI: 52.9–92.07]) were mixed infections with P. falciparum. Whereas only a single sample was a P. malariae mono-infection (1/4, 25%) and the remaining 3/4 (75%) were mixed infections with P. falciparum.
Molecular identification and distribution of Plasmodium spp.(P. falciparum vs. others).
Of the 17 P. ovale positive samples by real-time PCR, 12 were successfully classified to the species level. Six (35.3% [95% CI: 17.6–63.4]) were P. ovale curtisi (Poc) and three (17.7% [95% CI: 0.0–45.7]) P. ovale wallikeri (Pow) Fig 3. Mixed infections of Pow/Poc were identified in three (17.7% [95% CI: 0.0–45.7]) samples (Fig 2). Although most (11/17; 64.7%) of these P. ovale positives were from Saint Vincent De Paul hospital, they were widely distributed across several quarters of Dschang. Moreover, 70.6% (12/17) of those infected with P. ovale spp were female. The most commonly reported symptoms amongst the P. ovale positives were headache 29.4% (5/17) followed by general fatigue 23.5% (4/17). Thirteen (76.5%) of these patients were identified to live within 1-km of lakes. All the P. ovale mono infections came from 20- to 23-year-old patients with headaches (3/5; 60%) as this was the most represented symptom. No significant association was observed between Plasmodium species infections and clinical parameters and documented symptoms.
Distribution of P. ovale species.
Discussion
We investigated the prevalence of non-falciparum malaria in Dschang, West region, Cameroon using highly sensitive molecular testing among individuals reporting to clinics with symptoms consistent with uncomplicated malaria. We showed that non-falciparum malaria is circulating within Dschang, resulting in clinical malaria cases, the majority of which are associated with mixed infections with P. falciparum and only a handful were due to non-falciparum mono-infections.
P. falciparum accounted for more than 90% of all malaria cases. This is consistent with other studies across Sub-Saharan Africa [3,27–29] and in Cameroon [30–33]. Even though P. falciparum prefers young reticulocytes, it has the ability to invade all red blood cell (RBC) age classes, increasing the probability of having higher P. falciparum parasite density, thereby reducing RBC availability for other Plasmodium species [34]. The low prevalence of P. malariae in this study does not align with other studies carried out in Cameroon. Indeed, Nguiffo-Nguete et al. [4] reported a high proportion of P. malariae infections in Adamaoua; reporting 2.5% P. malariae mono-infections as compared to 0.4% in our study and 17% P. falciparum/P. malariae co-infections compared to 1.3% in our study. A possible explanation could be the difference in transmission patterns in both epidemiological zones as well as the sample collection period [17,35]. In these studies, the authors looked at the transmission pattern of malaria species in several regions of Cameroon ranging from the Sahelian region to the forest humid zone passing across the tropical area and the great highlands.
In this study, P. vivax was not identified, which contrasts with previous studies in Cameroon [8,12,36] and in Dschang in particular [6,7]. The study conducted by Ngassa et al. [36] targeted 4 regions of Cameroon (Centre, Littoral, South, and East) and recruited symptomatic hospital cases. They reported at least one P. vivax infection either as a mono- or mixed infection with P. falciparum in all study sites. Triple infection with P. malariae as well as mono- and mixed infections was also reported by Fru-Cho et al. [12] in Bolifamba in the Southwest region of Cameroon. These reported levels of P. vivax are relatively low compared to levels reported by Russo et al. who sampled febrile patients attending the hospital (35% of all malaria infections in Dschang (West region of Cameroon)), suggesting a potential P. vivax hotspot that time [6]. Although we did not find P. vivax in our study, this may have been due to timing of sampling or other unmeasured variables. The previous reports support the need for wider sampling, not only in hospital cases, but also asymptomatic community cases.
Overall, we had a combined prevalence of 4% (17/431) for P. ovale spp. with 5 mono-infections and 12 co-infections with P. falciparum. We documented the presence and distribution of both species of P. ovale in Dschang either as single infections or in mixed infections. This study is consistent with reports from Kojom-Foko and colleagues [37] who were the first to report the presence of P. ovale curtisi in Douala, Cameroon [30]. Our results corroborate findings from other studies conducted in countries neighbouring Cameroon, such as Equatorial Guinea [38], the Republic of Congo [39], Nigeria and Gabon [40], which reported the presence of both species of ovale. The detection of both P. ovale species in Dschang highlights the need to better describe the epidemiology and transmission of these species among populations in Cameroon.
The relapsing malarias, P. ovale spp. and P. vivax deserve further study in Cameroon due to the need for understanding relapses and whether hypnozoite treatment with primaquine is needed. Equally important is the need to address burden of relapsing malaria in Cameroon due to their potential role in life-threatening malaria complications and severe thrombocytopenia [18]. This will prompt decision makers to strengthened diagnostic strategies with the use of G6PD testing and to make primaquine readily available for the treatment. Together, these factors necessitate improved surveillance and diagnostic capabilities to accurately detect and differentiate the various species and subspecies of Plasmodium in Cameroon.
There are limitations to this study. We may have missed dormant P. vivax and P. ovale spp. infections due to their ability to form hypnozoites [41]. Furthermore, the hospital-based approach for the collection of our samples limited us to clinical cases suspected of malaria infection, and non-falciparum infections may result in less severe symptoms reducing the likelihood of seeking care, although previous studies of P. vivax from Dschang identified cases from symptomatic persons [6]. Population-targeted sampling of asymptomatic individuals was not conducted, which may differ from symptomatic case prevalence estimates.
Conclusion
We characterised the prevalence of Plasmodium species, particularly the non-falciparum species, among individuals presenting to clinic in Dschang in 2020. The results show that non-falciparum malaria species circulate in Dschang in non-negligible proportions, including both species of Plasmodium ovale spp. Non-falciparum malaria remains a concern locally in Dschang, and expanding community-level surveillance will help clarify its burden and transmission. In addition, much remains to be learned about P. ovale curtisi and P. ovale wallikeri biology and epidemiology in sub-Saharan Africa. These include the roles they play in relapsing malaria, the dynamics of coinfections with other species as well as how they may be evolving in response to current malaria control efforts designed to target P. falciparum. Our findings suggest that the degree to which these closely related but sympatric species co-circulate within their human hosts may be under-appreciated. Further studies of non-falciparum malaria, in both febrile and asymptomatic individuals, across Cameroon will help us better understand these neglected pathogens and develop control measures to eliminate all malaria.
Supporting information
S1 TablePrimer sequences and reaction conditions.(DOCX)
S2 TableMultivariate logistic regression of sociodemographic factors (Gender, Age, level of education) associated with malaria infection.(DOCX)
S1 FileInclusivity-in-global-research-questionnaire-filled-TKVP.(DOCX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Tchuenkam PVK, Ngum LN, Ali IM, Chedjou JPK, Nji AM, Netongo PM, et al. Plasmodium falciparumdhps and dhfr markers of resistance to sulfadoxine-pyrimethamine five years (2016-2020) after the implementation of seasonal malaria chemoprevention in Cameroon. Wellcome Open Res. 2024;9:323. doi: 10.12688/wellcomeopenres.22347.1 39649624 PMC 11624434 · doi ↗ · pubmed ↗
- 2GnéméA, Guelbéogo WM, Riehle MM, Tiono AB, Diarra A, KabréGB, et al. Plasmodium species occurrence, temporal distribution and interaction in a child-aged population in rural Burkina Faso. Malar J. 2013;12:67. doi: 10.1186/1475-2875-12-67 23421809 PMC 3583752 · doi ↗ · pubmed ↗
- 3Nguiffo-Nguete D, Nongley Nkemngo F, Ndo C, Agbor J-P, Boussougou-Sambe ST, Salako Djogbénou L, et al. Plasmodium malariae contributes to high levels of malaria transmission in a forest-savannah transition area in Cameroon. Parasit Vectors. 2023;16(1):31. doi: 10.1186/s 13071-022-05635-7 36698132 PMC 9875470 · doi ↗ · pubmed ↗
- 4Howes RE, Reiner RC Jr, Battle KE, Longbottom J, Mappin B, Ordanovich D, et al. Plasmodium vivax Transmission in Africa. P Lo S Negl Trop Dis. 2015;9(11):e 0004222. doi: 10.1371/journal.pntd.0004222 26587988 PMC 4654493 · doi ↗ · pubmed ↗
- 5Russo G, Faggioni G, Paganotti GM, Djeunang Dongho GB, Pomponi A, De Santis R, et al. Molecular evidence of Plasmodium vivax infection in Duffy negative symptomatic individuals from Dschang, West Cameroon. Malar J. 2017;16(1):74. doi: 10.1186/s 12936-017-1722-2 28196496 PMC 5309988 · doi ↗ · pubmed ↗
- 6Djeunang Dongho GB, Gunalan K, L’Episcopia M, Paganotti GM, Menegon M, Efeutmecheh Sangong R, et al. Plasmodium vivax Infections Detected in a Large Number of Febrile Duffy-Negative Africans in Dschang, Cameroon. Am J Trop Med Hyg. 2021;104(3):987–92. doi: 10.4269/ajtmh.20-1255 33534776 PMC 7941824 · doi ↗ · pubmed ↗
- 7Ngassa Mbenda HG, Gouado I, Das A. An additional observation of Plasmodium vivax malaria infection in Duffy-negative individuals from Cameroon. J Infect Dev Ctries. 2016;10(06):682–6.27367020 10.3855/jidc.7554 · doi ↗ · pubmed ↗
- 8Brazeau NF, Mitchell CL, Morgan AP, Deutsch-Feldman M, Watson OJ, Thwai KL, et al. The epidemiology of Plasmodium vivax among adults in the Democratic Republic of the Congo. Nat Commun. 2021;12(1):4169. doi: 10.1038/s 41467-021-24216-3 34234124 PMC 8263614 · doi ↗ · pubmed ↗