Zinc-enhanced activity of an antimicrobial halogenated phenazine against Streptococcus mutans and other gram-positive bacteria
Jessica K. Kajfasz, Hannah B. Hosay, Qiwen Gao, Robert W. Huigens, José A. Lemos

TL;DR
A halogenated phenazine compound, HP-29, shows strong antimicrobial activity against Gram-positive bacteria, and its effect is greatly enhanced when combined with zinc.
Contribution
The discovery that zinc enhances the antimicrobial activity of HP-29 against Gram-positive bacteria, including Streptococcus mutans, is novel.
Findings
HP-29 inhibits the growth of Streptococcus mutans and other Gram-positive bacteria in a dose-dependent manner.
Zinc supplementation significantly enhances the antimicrobial effect of HP-29, while iron, cobalt, and manganese supplementation reduce it.
Transcriptome analysis reveals that HP-29 disrupts metal ion regulation, with zinc amplifying this disruption.
Abstract
Halogenated phenazine (HP) compounds have shown promise as antimicrobial agents, particularly against biofilm-associated Gram-positive pathogens. Among these compounds, HP-29 demonstrates potent activity against methicillin-resistant Staphylococcus aureus by inducing rapid iron starvation. As maintenance of trace metals homeostasis is critical for the survival of Streptococcus mutans, this study investigated the antimicrobial efficacy of HP-29 and the impact of metal supplementation on this major oral and occasional systemic pathogen. As anticipated, HP-29 inhibited S. mutans growth in a dose-dependent manner, with iron supplementation alleviating the antimicrobial effect. Cobalt, manganese, or nickel supplementation also mitigated the inhibitory activity of HP-29, but, unexpectedly, the addition of zinc greatly enhanced HP-29 antimicrobial activity. This zinc-driven potentiation of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
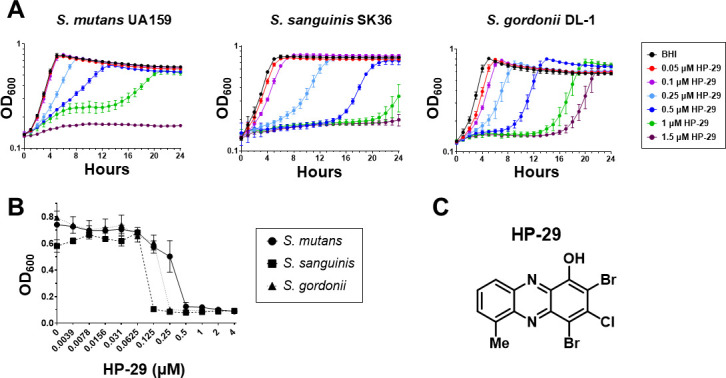 Fig 1
Fig 1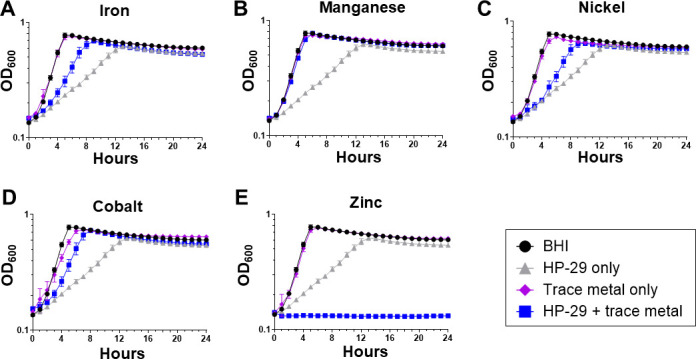 Fig 2
Fig 2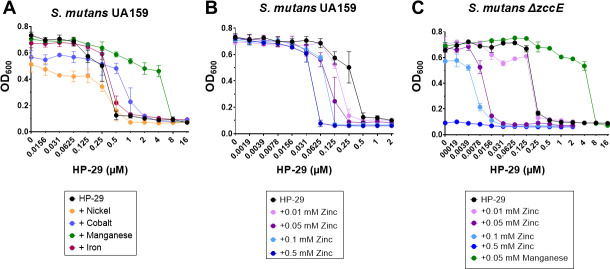 Fig 3
Fig 3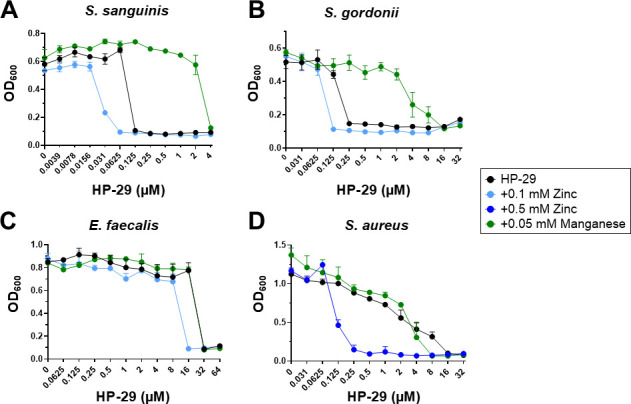 Fig 4
Fig 4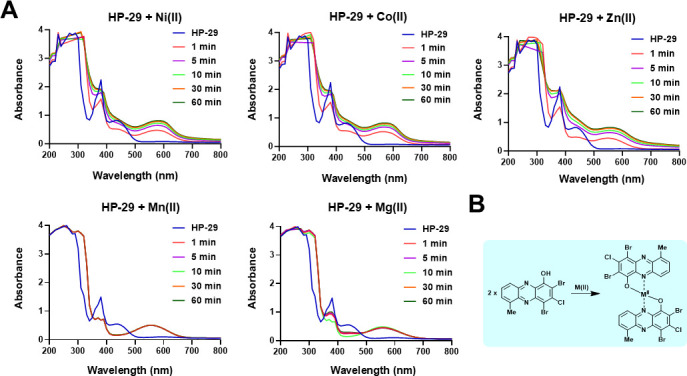 Fig 5
Fig 5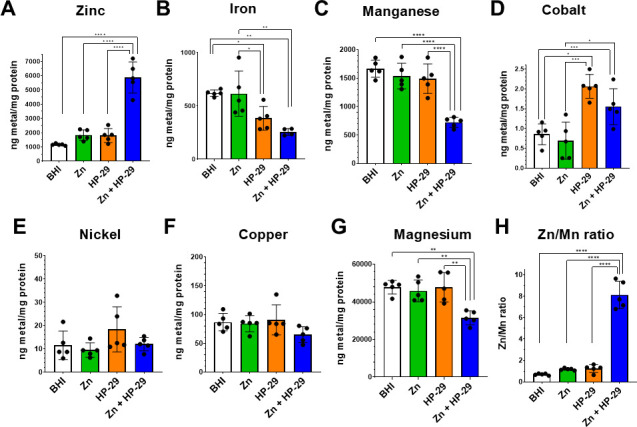 Fig 6
Fig 6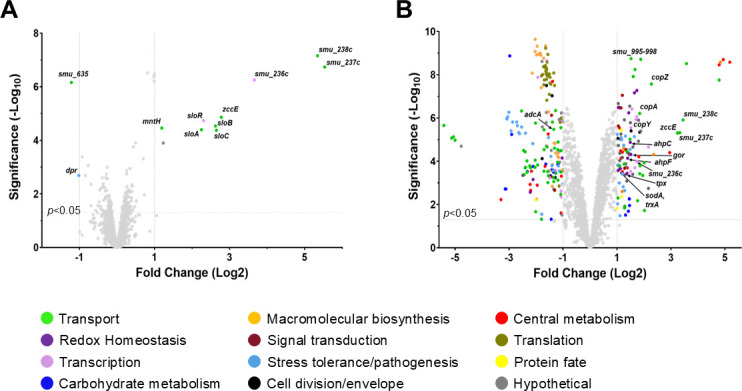 Fig 7
Fig 7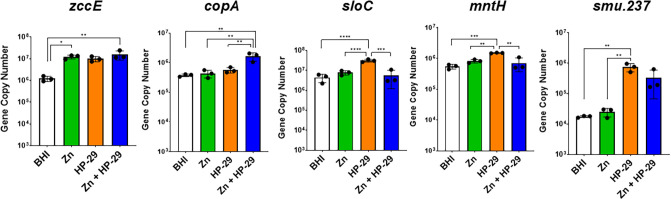 Fig 8
Fig 8| Medium supplementation | ||||||
|---|---|---|---|---|---|---|
| HP-29 only | 0.5 | 0.25 | 0.125 | 0.25 | 32 | 16 |
| HP-29 + 1 mM iron | 0.5 | |||||
| HP-29 + 50 µM manganese | 8 | 8 | 4 | 16 | 32 | 8 |
| HP-29 + 250 µM cobalt | 2 | |||||
| HP-29 + 500 µM nickel | 0.5 | |||||
| HP-29 + 10 µM zinc | 0.25 | 0.25 | ||||
| HP-29 + 50 µM zinc | 0.25 | 0.0156 | ||||
| HP-29 + 100 µM zinc | 0.125 | 0.0156 | 0.0625 | 0.125 | 16 | |
| HP-29 + 500 µM zinc | 0.0625 | 0.25 |
- —National Institute of Dental and Craniofacial Researchhttp://dx.doi.org/10.13039/100000072
- —National Institute of General Medical Scienceshttp://dx.doi.org/10.13039/100000057
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVanadium and Halogenation Chemistry · Nanoparticles: synthesis and applications · Trace Elements in Health
INTRODUCTION
The constant and fast emergence of multidrug (MDR) resistant bacteria is a topic of great concern, prompting efforts to develop new antibacterial strategies (1, 2). Recent work has shown that certain halogenated phenazine (HP) analogs can be highly effective against selected Gram-positive pathogens, including those with broad resistance to traditional antibiotics (3). This series of compounds was inspired by natural bacterial competition, in which strains of Pseudomonas aeruginosa were found to secrete pyocyanin, a phenazine antibiotic and virulence factor, that eliminated Staphylococcus aureus from the lungs of cystic fibrosis patients (4). Focused libraries of HPs have been synthesized and found to effectively kill Gram-positive pathogens, both in the planktonic and the biofilm state (5–10). Supporting previous observations that HP compounds bind divalent metal cations (6–10), transcriptional profiling of methicillin-resistant S. aureus (MRSA) biofilms following treatment with HP-14 (HP analog 14) revealed a rapid induction in expression of gene clusters associated with iron uptake (11). Continued efforts to explore new HP analogs through chemical synthesis and microbiological studies led to the identification of HP-29, which was highly effective against MRSA and Enterococcus faecalis among other major Gram-positive pathogens (10). When provided as a topically applied ointment, HP-29 treatment significantly reduced bacterial burden on wounds of mice infected with either E. faecalis or S. aureus (10).
Streptococcus mutans is a keystone pathogen of dental caries and one of the causative agents of infective endocarditis, a life-threatening infection of heart valve endothelium (12–15). As a member of the oral biofilm community, the S. mutans lifestyle demands the ability to adapt to large fluctuations in the availability and content of nutrients, which derive almost exclusively from the human host diet (16). Transition metals are essential micronutrients for all domains of life, as the function of about 40% of all enzymes is dependent upon a metal cofactor (17–19). Studies conducted by our group and others have shown that the ability to maintain trace metal homeostasis—achieved by proteins dedicated to the sensing, import, and efflux of metals—is a critical aspect of S. mutans pathophysiology (20–27). Several transport systems responsible for the import and efflux of trace metals in S. mutans have been characterized, including those for iron (feoABC, sloABC, smu.995-998) (20, 26), manganese (sloABC, mntH, mntE) (25, 26, 28), zinc (adcABC, zccE) (21, 22), and copper (copA) (27). As over-accumulation of metals is associated with toxicity, S. mutans relies on metal-sensing regulators (metalloregulators) to tightly govern metal uptake and efflux, including SloR (iron and manganese uptake), AdcR (zinc uptake), CopY (copper export), and ZccR (zinc export) (21, 22, 29–32).
In this investigation, we assessed the potential of HP-29 to serve as an antimicrobial agent against S. mutans. Growth of S. mutans was inhibited by HP-29 in a dose-dependent manner that was alleviated by supplementation with iron. Expanding the study to include other trace metals revealed that cobalt, manganese, and nickel could also alleviate HP-29 inhibitory activity. However, zinc supplementation greatly enhanced the antimicrobial efficacy of HP-29, an observation that was extended to other Gram-positive bacteria. Intracellular trace metal quantifications and transcriptome analysis of S. mutans cultures treated with HP-29 alone or combined with a non-inhibitory concentration of zinc revealed that HP-29 broadly disrupts trace metal homeostasis, and that this effect is further exacerbated upon the addition of zinc. This study confirms HP-29 as a potent antimicrobial agent against Gram-positive pathogens that disrupt intracellular metal homeostasis while also revealing the therapeutic potential of combining HPs with zinc to treat bacterial infections.
RESULTS
HP-29 is inhibitory to oral streptococci in a metal-dependent manner
To explore the antimicrobial potential of HP-29 for the prevention and treatment of S. mutans infections, we first tested the ability of S. mutans UA159 to grow in the presence of HP-29. Growth curves revealed a dose-dependent effect with 1.5 μM HP-29 completely inhibiting growth, with similar inhibition seen for the oral streptococcal species S. sanguinis and S. gordonii (Fig. 1A). MIC determinations performed using the broth microdilution method confirmed the efficacy of HP-29 against these streptococci, with S. sanguinis showing the greatest sensitivity (MIC of 0.125 μM) and S. mutans showing the highest tolerance (MIC of 0.5 μM) (Fig. 1B, Table 1). The chemical structure of HP-29 is shown in Fig. 1C.
The novel compound HP-29 inhibits the growth and survival of oral streptococci. The oral streptococcus strains S. mutans UA159, S. sanguinis SK36, and S. gordonii DL-1 were grown in BHI medium and exposed to the halogenated phenazine HP-29 in (A) growth curve assays or (B) minimal inhibitory concentration (MIC) assays. (C) Chemical structure of HP-29. Data represent averages and standard deviations of at least three independent experiments.
Previously, we have shown that brain-heart infusion (BHI) medium, used in the growth curve and MIC assays, contains low concentrations of the essential trace metals iron (~6 μM), zinc (~11 μM), and manganese (<1 µM) (25). Knowing that HP-29 triggers rapid iron starvation in S. aureus (10), we repeated the HP-29 growth kinetic assay in BHI supplemented with a sub-inhibitory concentration of iron, focusing first on the response of S. mutans. As expected, the addition of 1 mM FeSO_4_, the highest feasible concentration due to iron’s tendency to precipitate, partially rescued the growth inhibitory effect of HP-29 (Fig. 2A). Given that HP-29 can bind other metals in vitro (3, 10), we next sought to evaluate the effect of supplementation with sub-inhibitory concentrations of other divalent metals on HP-29 activity. Similar to iron, supplementation with manganese, nickel, or cobalt alleviated the inhibitory activity of HP-29 (Fig. 2B through D). Notably, while the addition of iron, nickel, or cobalt offered partial rescue of S. mutans UA159 growth, the addition of manganese resulted in a complete growth restoration. Unexpectedly, zinc supplementation caused the opposite effect, strongly potentiating the growth inhibitory effects of HP-29 (Fig. 2E).
Divalent metal cations can either rescue or exacerbate the ability of HP-29 to inhibit the growth of S. mutans. S. mutans UA159 was grown in BHI medium containing 0.5 μM HP-29. The medium was supplemented with the divalent metal cations (A) iron, 1 mM; (B) manganese, 0.5 mM; (C) nickel, 0.5 mM; (D) cobalt, 0.25 mM; or (E) zinc, 0.5 mM. Data represent averages and standard deviations of at least three independent experiments.
To confirm these results, we assessed the MIC of HP-29 when combined with each individual metal (Fig. 3A; Table 1). Supporting the trends observed in the growth curves, cobalt or manganese supplementation raised the HP-29 MIC from 0.5 μM to 2 μM or 8 μM, respectively. Despite the effects noted on growth curve assays, nickel or iron supplementation did not affect the HP-29 MIC. Most notably, the addition of 0.5 mM ZnSO_4_ lowered the HP-29 MIC to 0.0625 μM HP-29, an 8-fold increase in sensitivity compared to exposure to HP-29 alone (Table 1). Since the addition of zinc resulted in such a strong phenotype, a zinc titration was performed, revealing that as little as 0.01 mM ZnSO_4_ increased S. mutans sensitivity to HP-29 by 2-fold (Fig. 3B; Table 1). Previously, we showed that the zinc exporter ZccE mediates the high-zinc tolerance of S. mutans (21). Here, we show that the ΔzccE strain was slightly more sensitive to HP-29 than the parent strain in BHI (MIC of 0.25 µM for ΔzccE compared to 0.5 µM for UA159). As expected, the combination of HP-29 with sub-inhibitory concentrations of ZnSO_4_ was highly inhibitory to the growth of the ΔzccE strain. For instance, in the presence of 0.05 mM zinc, the MIC of HP-29 decreased 16-fold to 0.0156 μM (Fig. 3C; Table 1). As seen with the parent strain, manganese supplementation was highly effective at rescuing sensitivity of ΔzccE to HP-29, increasing the MIC by 32-fold (Fig. 3C; Table 1). Table S1 lists the final OD_600_ values obtained in the MIC assays.
Divalent metal cations can either rescue or potentiate the MIC of HP-29 against S. mutans. (A) HP-29 MIC assay for S. mutans UA159 with the addition of trace metals: 0.5 mM nickel, 0.25 mM cobalt, 0.05 mM manganese, and 1 mM iron. (B) A titration assay shows that as little as 0.01 mM zinc increases the sensitivity of S. mutans UA159 to HP-29. (C) The HP-29 MIC assays with supplementation of manganese or zinc were repeated with the S. mutans ΔzccE strain. Data represent averages and standard deviations of at least three independent experiments.
Knowing that metal tolerance varies widely among bacteria (33, 34), the ability of divalent metal cations to rescue or potentiate the inhibitory effect of HP-29 in other oral streptococci (S. sanguinis and S. gordonii), E. faecalis, and S. aureus was evaluated. Here, we used only manganese or zinc as representatives of metals that either rescue or potentiate the antimicrobial activity of HP-29 in S. mutans. Like in S. mutans, manganese supplementation greatly increased the HP-29 MIC for S. sanguinis and S. gordonii, while zinc supplementation increased sensitivity by 2-fold for both strains (Fig. 4A and B). We also assessed the impact of manganese and zinc when combined with HP-29 against E. faecalis and S. aureus, organisms of particular medical concern to humans due to their association with antibiotic resistance (35–38). While manganese supplementation failed to alleviate the HP-29 sensitivity in E. faecalis, the addition of zinc increased its HP-29 MIC by 2-fold (from 32 μM to 16 μM) (Fig. 4C). In S. aureus, the addition of zinc caused a remarkable 64-fold increase in HP-29 sensitivity (from 16 μM to 0.25 μM), while manganese supplementation lowered the MIC from 16 to 8 μM (Fig. 4D).
Divalent metal cations can either rescue or exacerbate the MIC of HP-29 against a panel of bacterial strains. MIC assays were performed in the presence of manganese or zinc to test their ability to rescue or potentiate the sensitivity to HP-29 for (A) S. sanguinis, (B) S. gordonii, (C) E. faecalis, and (D) S. aureus. Data represent averages and standard deviations of at least three independent experiments.
In previous work, HP-29 has shown rapid binding to divalent iron in UV-vis spectroscopy experiments (10). Based on the rescue and potentiation profiles observed in the presence of different metals, we investigated the ability of HP-29 to bind other divalent metal cations. UV-vis spectroscopy revealed that the binding capacity of HP-29 extends to a range of divalent metal cations, including nickel, cobalt, manganese, magnesium, and zinc (Fig. 5A), with HP-29 binding to these metal cations in a 2:1 ratio (Fig. 5B).
HP-29 is able to bind to several divalent metal (II) cations. (A) UV-vis spectroscopy of HP-29 binding nickel (II), cobalt (II), zinc (II), manganese (II), and magnesium (II). (B) There is a 2:1 HP-29:metal (II) cation complex that forms from the direct metal binding of HP-29 in UV-vis experiments.
HP-29 causes a disruption of metal homeostasis in S. mutans that is exacerbated by the addition of zinc
The heightened potency of HP-29 in the presence of zinc, combined with the confirmation that HP-29 is capable of binding a range of divalent metal cations, introduced the possibility that the inhibitory effect of HPs may be associated with a more extensive disruption of metal homeostasis. To assess the broader impact of HP-29 on intracellular metal content, mid-exponential phase cultures of S. mutans UA159 were grown in BHI and exposed to sub-inhibitory concentrations of (i) 0.025 μM HP-29, (ii) 0.5 mM ZnSO_4_, or (iii) 0.025 μM HP-29 and 0.5 mM ZnSO_4_ for 90 min, with each treatment group individually compared to a control condition kept in BHI. As expected, treatment with zinc alone did not significantly affect intracellular zinc levels (Fig. 6A), as S. mutans can maintain zinc homeostasis at high concentrations of zinc due to the activity of the ZccE exporter (21). As a result, treatment with zinc did not impact the intracellular levels of the other metals (iron, manganese, cobalt, nickel, copper, and magnesium) tested (Fig. 6B through G). Treatment with HP-29 alone resulted in a significant decrease in intracellular iron content that dropped by ~40% when compared to the untreated control (Fig. 6B). These results provide the first direct evidence that HP-29 disrupts intracellular iron homeostasis in a bacterial species. While HP-29 did not affect the intracellular levels of zinc, manganese, nickel, copper, or magnesium, exposure to HP-29 caused a significant 2-fold increase in intracellular cobalt (Fig. 6D). In addition to iron, dual treatment with zinc and HP-29 revealed other important changes in intracellular metal content. Most strikingly, intracellular zinc soared in the dual-treatment condition to levels that were more than 3-fold greater than those seen in either of the single-compound treatments (Fig. 6A). Furthermore, dual treatment decreased intracellular manganese by ~50% and magnesium by ~30%, with no significant changes in cobalt, nickel, or copper content observed when compared to HP-29 alone (Fig. 6C through G). Notably, the dual treatment significantly disrupted the zinc-to-manganese ratio, reversing the typical balance that favors manganese by approximately 50% to instead favor zinc at an 8:1 ratio (Fig. 6H). This shift reflects the well-documented phenomenon in which zinc-induced manganese deficiency broadly impacts bacterial physiology (21, 39, 40).
*ICP-MS quantification of intracellular biometals reveals that exposure to HP-29 disrupts metal homeostasis in S. mutans. Mid-exponential phase cultures (OD600 = 0.4) of S. mutans UA159 were exposed to BHI alone or supplemented with 0.5 mM ZnSO4, 0.025 μM HP-29, or both for 90 min. After washing in PBS, the ICP-MS analysis was performed on the harvested cells to determine the intracellular content of biometals: (A) zinc, (B) iron, (C) manganese, (D) cobalt, (E) nickel, (F) copper, or (G) magnesium. (H) The intracellular zinc:manganese ratio in each condition. Data represent averages and standard deviations of five independent experiments. ****, P ≤ 0.0001; ***, P ≤ 0.001; **, P ≤ 0.01; , P ≤ 0.05.
Analysis of intracellular metal content revealed that exposure to HP-29 and zinc disrupted the homeostasis of iron, manganese, and magnesium (Fig. 6). To determine whether supplementation with these metals could mitigate the growth inhibition induced by the HP-29/zinc combination, we supplemented cultures with iron, manganese, or magnesium. Although supplementation with iron or manganese restored the growth inhibition caused by HP-29 alone (Fig. 2), addition of any of these metals at a 1:1 test metal:zinc ratio failed to rescue the growth defect associated with dual HP-29 and zinc treatment (data not shown). The experiment was repeated using the highest concentrations of iron, manganese, or magnesium that remained soluble in combination with HP-29 and zinc, corresponding to test metal:zinc ratios of 2:1 (iron or manganese:zinc) and 20:1 (magnesium:zinc). Under these conditions, none of the metal supplements alleviated the growth inhibition elicited by the combination of HP-29 and zinc (Fig. S1). These results indicate that the dysregulation of intracellular metal homeostasis that results from the HP-29/zinc co-treatment is extensive and cannot be easily reversed by metal supplementation.
Transcriptome analysis following exposure to HP-29 clearly indicates trace metal stress
To obtain clues on the antimicrobial effects of HP-29 and to better understand the synergistic association of HP-29 with zinc, S. mutans UA159 cultures were grown to mid-log phase, treated for 30 min with 0.025 μM HP-29 or with 0.025 μM HP-29 and 0.5 mM ZnSO_4_, and then subjected to RNA sequencing (RNA-seq) analysis. When compared to untreated cells kept in BHI, only 12 genes showed significantly altered expression after applying a 2-fold linear cutoff (1-fold log_2_) (Fig. 7A; Table S2; P < 0.05). Yet, the identity of these 12 genes (10 up and 2 downregulated) reveals a compelling story of disrupted metal homeostasis. Among the upregulated genes was the entire sloABC operon and its cognate regulator sloR (4.7- to 6.3-fold linear; 2.3- to 2.6-fold log_2_), as well as mntH (2.3-fold linear; 1.2-fold log_2_). The sloABC operon encodes a highly conserved ABC transporter that mediates iron and manganese uptake, while mntH encodes an Nramp-type transporter that mediates manganese uptake (25, 26). In addition, the zinc exporter zccE was upregulated by 6.8-fold linear (2.8-fold log_2_), revealing that HP-29 triggers a high-zinc stress response even when cells are grown in a low-zinc medium such as BHI (25). The most highly upregulated genes were smu.236c, smu.237c, and smu.238c (from 13- to 46-fold linear; 3.6- to 5.5-fold log_2_), members of an uncharacterized three-gene operon comprising a TetR-type regulator (smu.236c) and permease (smu.237c) and ATP-binding (smu.238c) proteins of an ABC transporter (41). The only two downregulated genes were dpr (0.5-fold linear; −1.0-fold log_2_) and smu.635 (0.4-fold linear; −1.2-fold log_2_). The PerR-regulated dpr gene encodes a ferritin-like protein that acts as an Fe^2+^ sink, protecting cells from oxidative stress caused by Fenton chemistry (20, 42). The downregulation of dpr further suggests that HP-29 induces iron starvation. Additionally, smu.635 encodes an uncharacterized transmembrane protein. Although its function is unknown, it is also a member of the PerR regulon (43). Homology predictions suggest that smu.635 may be involved in manganese transport and homeostasis (41, 43).
Volcano plots of genes differentially expressed in S. mutans UA159 following a 30-minute exposure to (A) 0.025 µM HP-29 or (B) 0.025 µM HP-29 with 0.5 mM ZnSO4, compared to a BHI control condition. The x-axes indicate the log2 fold change in expression, while the y-axes indicate the significance. Selected genes of interest are noted. Colors are used to indicate the predicted function of the genes as listed. Genes that do not meet the threshold for significance (1-fold change, P < 0.05) are shown in light gray.
Exposure to the dual HP-29/zinc combination resulted in a much greater number of genes differentially expressed compared to the untreated condition, with 119 genes upregulated and 182 genes downregulated (Fig. 7B; Table S3, 1-fold cutoff (log_2_), P < 0.05). Not surprisingly, dual treatment increased zccE expression by 9.4-fold linear (3.2-fold log_2_), while transcription of adcA, which encodes the substrate-binding protein of the zinc uptake system AdcABC (22), was downregulated by 2.4-fold linear (−1.3-fold log_2_). The genes comprising the smu.995-998 operon, which we have previously described to be involved in iron uptake (20), also showed increased expression (2.9- to 3.7-fold linear; 1.5- to 1.9-fold log_2_) following dual treatment. Also significantly upregulated were the genes of the copYAZ operon (3.2- to 4.8-fold linear; 1.7- to 2.3-fold log_2_) that mediate copper export. Finally, the genes of the smu.236c-238c operon were also strongly upregulated (3.3- to 10.9-fold linear; 1.7- to 3.4-fold log_2_) following dual treatment. Of note, the smu.236c-238c operon was not impacted in our previous study of the S. mutans transcriptome following exposure to zinc (21), indicating that this induction is driven by HP-29. In total, 66 transport-associated genes were differentially expressed following the dual treatment exposure. As previously observed under zinc stress alone (21), several genes associated with oxidative stress tolerance were also upregulated upon dual treatment, including ahpCF, gor, gst, tpx, gloA, sodA, and trxA. For comparison purposes, Table S3, which contains the complete list of genes showing significant expression changes following treatment with both HP-29 and zinc, indicates which of these genes were also identified in our earlier study of the impact of zinc on the S. mutans transcriptome. Overall, the dual-treatment transcriptome analysis reinforces the notion that HP-29 causes broad disruption of trace metal homeostasis, and that the effect is exacerbated in the presence of elevated, but not inherently inhibitory, zinc concentrations.
To validate the results obtained from the RNA-seq analysis, qRT-PCR was utilized to examine the expression of selected metal transport genes after HP-29 (0.025 μM HP-29) and HP-29/zinc (0.025 μM HP-29 + 0.5 mM ZnSO_4_) exposure. This time, a zinc-only (0.5 mM ZnSO_4_) treatment was included, allowing direct comparisons of the impact of individual and dual treatments (Fig. 8). As anticipated, expression of zccE was significantly induced (~10-fold) in all three treatment conditions compared to the untreated controls, whereas only the combination of HP-29 and zinc resulted in a significant increase in expression of the copper exporter copA. The qRT-PCR profile of the dual transporter sloC and the manganese transporter mntH mirrored the RNA-seq results: HP-29 treatment alone led to significant upregulation of both transporters, whereas dual treatment did not. Finally, the elevated expression of smu.237c (ABC transport permease) following exposure to HP-29 or both HP-29 and zinc, but not zinc alone, was confirmed by qRT-PCR.
*qRT-PCR analysis of genes associated with metal homeostasis in S. mutans UA159 following a 30-minute exposure to 0.025 µM HP-29, 0.5 mM ZnSO4, or both. zccE and copA encode proteins associated with zinc efflux. sloC and mntH encode proteins that function in iron (sloC) and manganese (sloC and mntH) import. smu.237 encodes a putative transporter of unknown function that is highly upregulated following exposure to HP-29. Data represent averages and standard deviations of at least three independent experiments. ****, P ≤ 0.0001; ***, P ≤ 0.001; **, P ≤ 0.01; , P ≤ 0.05.
DISCUSSION
Antimicrobial resistance is a major global concern, as introduction of antibiotics into clinical use is often (and quickly) followed by the emergence of MDR bacterial strains, placing an enormous burden on healthcare worldwide (2, 44). HPs were initially discovered through efforts inspired by phenazine antibiotics produced by P. aeruginosa in biological warfare against S. aureus (4, 6, 7, 9). One potent HP biofilm-eradicating agent discovered during initial efforts, HP-14, was shown to cause a rapid iron starvation in MRSA biofilms (11). Further structural modifications of the original HP scaffold led to the identification of HP-29, which demonstrates greater antibacterial and biofilm-killing activities (10).
Earlier studies have demonstrated that HP-29 is an effective antimicrobial against several bacterial species including S. aureus, S. epidermidis, E. faecalis, and S. pneumoniae (10). Due to the much greater efficacy of the HP compounds against Gram-positive bacteria, 10 to 100 times more effective when compared to Gram-negative bacteria (8), studies aimed at further developing these compounds as antimicrobial have focused on Gram-positive organisms. To date, neither HP-29 nor other HP analogs have been tested against the oral pathobiont S. mutans. Given our long-standing interest in the mechanisms of metal ion homeostasis in S. mutans and its link to virulence, this study investigated the impact of HP-29 on iron homeostasis in this major dental and occasional systemic pathogen. As we demonstrated the potential usefulness of HP-29 as an antimicrobial to combat S. mutans infections, we discovered that divalent metals, including cobalt, iron, manganese, and nickel, rescued S. mutans from HP-29–induced growth inhibition to varying degrees. Unexpectedly, sub-inhibitory concentrations of zinc significantly enhanced the antimicrobial activity of HP-29. This HP-29 plus zinc synergistic effect was also observed in E. faecalis and S. aureus, two major Gram-positive pathogens often associated with MDR infections, suggesting that combined HP-29 and zinc treatment may represent a novel therapeutic strategy for broad-spectrum bacterial control. However, testing its efficacy against additional bacterial pathogens and in animal infection models is warranted.
Through intracellular metal quantification and transcriptome analyses, we demonstrate that HP-29 broadly disrupts metal homeostasis, extending beyond the previously reported role in inducing iron starvation (10, 11). For the first time, we show that HP-29 treatment significantly reduces intracellular iron levels in bacteria. Additionally, our ICP-MS analysis reveals that HP-29 increases intracellular cobalt concentrations while exerting modest effects on zinc and manganese pools, slightly elevating zinc levels and reducing manganese levels. As ZccE has been shown to mediate tolerance to both zinc and cobalt (21), the increased transcription of zccE following HP-29 treatment could be a response to the influx of both metals. Perhaps the most compelling result was a striking 5-fold increase in intracellular zinc in cells treated with both HP-29 and zinc, compared to its untreated counterpart. This dual HP-29/zinc treatment also resulted in intracellular zinc quantities 3-fold greater than after treatment with either compound individually. A well-established consequence of high intracellular zinc stress is depletion of manganese (21, 39, 40, 45), and indeed, manganese levels dropped by approximately 50% in the dual-treated cells compared to single-treatment or untreated groups. As a result, these cells experienced a severe disruption in intracellular manganese:zinc balance, which is expected to negatively impact multiple cellular functions, including oxidative stress tolerance (21, 39, 40). In S. mutans and related bacteria, manganese contributes to oxidative stress survival by serving as a cofactor of the manganese-dependent superoxide dismutase (SOD) enzyme (46, 47) and, likely, by replacing iron as an enzymatic co-factor, thereby protecting iron-binding proteins from Fenton chemistry–induced damage. These findings provide a direct explanation as to why manganese supplementation alleviates the antimicrobial effects of HP-29. Furthermore, several genes associated with oxidative stress management were upregulated in dual-treated cells, supporting previous findings that zinc mismetallation is a key trigger of oxidative stress (21).
Although not a transition metal, magnesium levels were also measured following treatment. As the most abundant cation in living cells, magnesium plays a critical role in numerous cellular functions, including membrane stabilization, oligonucleotide folding, enzymatic cofactor activity, and participation in stress responses and virulence (48, 49). While individual treatments with HP-29 or zinc had no significant effect on intracellular magnesium levels, dual treatment resulted in a 30% decrease in magnesium content. This sharp decline is likely to impair multiple essential cellular processes, thereby contributing to the heightened susceptibility to HP-29. For example, studies in the model Gram-positive organism B. subtilis have shown that magnesium-depleted cells are unable to maintain normal protein translation and enter a state of stasis until magnesium homeostasis is restored (50).
Iron, manganese, and zinc are particularly important during infection, as the host actively limits pathogen access to these essential metals by producing metal-chelating proteins. In response, bacteria have evolved sophisticated mechanisms to scavenge these metals from host tissues (34, 51–53). Our transcriptome analysis strongly supported the conclusion that HP-29 broadly disrupts trace metal homeostasis. Focusing first on HP-29 exposure alone, we observed differential expression of a dozen genes (using a 2-fold linear change cutoff), with the majority clearly associated with metal transport and homeostasis. Notably, genes encoding manganese import systems, sloABC and mntH, were upregulated, while zccE was downregulated. When the cutoff was relaxed to include genes with a 1.5-fold linear change, we also detected upregulation (~1.8-fold) of the smu.995-998 operon, which is implicated in iron transport, as is the dual transporter sloABC (20). Collectively, this transcriptional response suggests that S. mutans activates multiple pathways to restore iron, manganese, and zinc homeostasis following HP-29 treatment. Despite the low concentration of zinc (0.5 mM ZnSO₄ vs 4 mM in our previous transcriptome study), the dual-treatment transcriptome still exhibits key features of the zinc stress response. These include upregulation of metal exporters zccE and copA, oxidative stress genes, and two major operons involved in lactose uptake and utilization. Notably, smu.236c-238c, an uncharacterized TetR regulator and two-gene ABC transporter lacking a cognate substrate-binding protein-encoding gene, was the most highly upregulated transcriptional unit in HP-29-treated cells and among the most upregulated in dual-treated cells. ABC transporters without substrate-binding proteins are not uncommon and have been linked to molecule export more often than import. Further investigation into the potential role of smu.236c-238c in metal transport is warranted.
The concept of zinc synergizing with antimicrobials has been a topic of interest in recent years. For example, the hydroxyquinoline compound PBT2, a zinc ionophore that has reached phase 2 clinical trials to treat Alzheimer’s and Huntington’s diseases, in combination with zinc (PBT2-Zn), was shown to exhibit antibacterial activity against Streptococcus pyogenes, MRSA, and VRE, and to act synergistically with other antibiotics, reversing antibiotic resistance (54). ICP-MS analysis of bacteria treated with PBT2-Zn revealed widespread disruption of metal homeostasis, with increases in zinc, iron, manganese, and copper, pointing to a major dysregulation of metabolic systems that contribute to virulence (54). In a recent study, the combination of zinc with the natural product carvacrol was shown to act synergistically with clinically relevant antibiotics, protecting mice from Pseudomonas aeruginosa lung infection (55). Furthermore, the association of a sub-family of antimicrobial peptides (AMP) with zinc (termed metalloAMPs) has been shown to potentiate the antimicrobial activity of the AMPs (56). Although the mechanisms underlying zinc’s synergistic effects with antimicrobials remain under investigation, our findings, together with these previous studies, suggest that zinc mismetallation broadly disrupts bacterial homeostasis.
In summary, we expand the breadth of Gram-positive pathogens susceptible to HPs, specifically HP-29, to include three species of oral streptococci. While confirming that HP-29 induces iron starvation, we further demonstrate that it disrupts broader aspects of metal homeostasis, as evidenced by altered intracellular cobalt and manganese levels. Additionally, unlike iron and manganese, which can rescue HP-29 sensitivity, zinc acts synergistically with HP-29. This suggests that a combined HP-zinc therapy may offer a promising strategy for treating infections caused by Gram-positive pathogens.
MATERIALS AND METHODS
Bacterial strains and growth conditions
The bacterial strains used in this study (S. mutans UA159, S. gordonii DL-1, S. sanguinis SK36, E. faecalis OG1RF) were obtained from the American Type Culture Collection (ATCC), with the exception of S. aureus RN4220, obtained from BEI Resources, and S. mutans ΔzccE, which was generated as previously described (21). Oral streptococci were routinely grown in BHI broth at 37°C in a 5% CO_2_ atmosphere. For physiologic analyses, bacterial inocula were prepared from overnight cultures, sub-cultured 1:20 into fresh media, and grown to mid-logarithmic phase (OD_600_ of 0.4), then diluted 1:25 into the indicated medium (BHI ± HP-29 and ± divalent metals) in a microtiter plate. Growth was monitored using the BioScreenC growth reader (Growth Curves USA) at 37°C, with each well overlayed with sterile mineral oil to minimize oxidative stress once the plate was shaken prior to OD_600_ readings. For RNA-Seq analysis, replicate cultures of S. mutans UA159 were grown as described above to an OD_600_ of 0.4 and separated into four aliquots: (i) BHI control, (ii) 0.5 mM ZnSO_4_, (iii) 0.025 μM HP-29, and (iv) 0.5 mM ZnSO_4_ + 0.025μM HP-29. These samples were incubated for an additional 30 min, harvested by centrifugation, and bacterial pellets were resuspended in 1 mL of RNA Protect Bacterial Reagent (Qiagen). Following another centrifugation cycle, the supernatants were discarded, and the pellets were stored at −80°C until use. For ICP-MS analysis, the cultures were grown as described above for RNA-Seq analysis but were incubated for 90 min following the addition of ZnSO_4_ and/or HP-29. The bacterial cells were harvested by centrifugation and then washed twice in PBS. After a final round of centrifugation, the harvested cell pellets were stored at −20°C until use. Similar methods were used for growth of E. faecalis and S. aureus, but these strains were incubated at 37°C in an aerobic environment.
MIC assays
The minimum inhibitory concentration (MIC) of HP-29 was determined by a broth microdilution method using 2-fold serial dilutions of HP-29 of ≥95% purity (21). Briefly, mid-logarithmic phase cultures (OD_600_ of 0.4, 1.5 × 10^9^ CFU/mL) were diluted 1:25 in the indicated medium. Ninety-six-well plates were incubated at 37°C in the appropriate atmosphere for 24 h, and the concentration of HP-29 at which the absorbance values were 15% or less of the control condition was determined to be the MIC. Assays were performed with a minimum of three biological replicates. Student’s t-test was performed to verify the significance of the results.
Metal binding to HP-29 (UV-vis)
A mixture of DMSO (970 μL) and HP-29 (30 μL of a 10 mM DMSO stock) was added to a 1.5 mL cuvette. In a separate cuvette, DMSO (955 μL), HP-29 (30 μL of a 10 mM DMSO stock), and metal (II) cation (15 μL of a 10 mM water solution) were added and thoroughly mixed. Then, spectral scanning was performed from 200 to 800 nm in 10 nm increments at various time points (1, 5, 10, 30, and 60 min). All divalent metal salt solutions were freshly made and added to the cuvette immediately. The elevated absorbance of the HP sample between 500 and 700 nm after the addition of divalent metal indicates rapid binding between HP-29 and the metals. Metal salts used in these studies included ZnSO_4_, NiSO_4_, CoSO_4_, MgSO_4_, and MnSO_4_∙H_2_O.
ICP-MS analysis
The intracellular metal content of S. mutans UA159 was determined via inductively coupled plasma mass spectrometry (ICP-MS) performed at the University of Florida Analytical Toxicology Core Laboratory (ATCL). The cell pellets were resuspended in HNO_3_ and incubated at 100°C for 30 min. Metal concentrations were determined using an Agilent 7900 ICP MS equipped with in-line internal standard addition. Data acquisition was accomplished in helium gas mode. Calibration ranges for all metals were 0–10,000 ng/mL, and all linear regression r^2^ values were 1.0000. Metal concentrations were normalized to total protein content, which was determined using the bicinchoninic acid (BCA) assay (Pierce). The data shown were collected from five biological replicates. Analysis of variance (ANOVA) was performed to assess the significance of the results.
RNA sequencing analysis
Total RNA from three biological replicates was isolated from homogenized S. mutans cell lysates by acid-phenol:chloroform extractions, as previously described (43). Briefly, nucleic acid obtained after homogenization was digested with Ambion DNaseI (Thermo Fisher) and then purified using the RNeasy kit (Qiagen), which included an on-column DNase digestion according to the manufacturer’s instructions. Sample quality and quantity were assessed using an Agilent 2200 Tape Station at the University of Florida Interdisciplinary Center for Biotechnology Research (ICBR). For RNA-Seq analysis, purified samples were sent to SeqCenter (Pittsburgh, PA) for DNase treatment. rRNA depletion was performed using the Ribo-Zero Plus Microbiome kit (Illumina) to generate RNA libraries. Following cDNA synthesis, libraries were subjected to RNA deep sequencing using the Illumina NovaSeq platform. Read quantification was performed using the featureCounts2 function of Subread. Quality control and adapter trimmer were performed with bcl-convert. Read mapping was performed with HISAT2 using the S. mutans UA159 genome (GenBank accession number NC_004350.2). Read counts loaded into R were normalized using edgeR’s Trimmed Mean of M values (TMM) algorithm. Subsequent values were then converted to counts per million (CPM). Differential expression analysis was performed using edgeR’s glmQLFTest function.
Targeted gene expression analysis was performed by quantifying mRNA using quantitative reverse transcriptase real-time PCR (qRT-PCR), according to an established protocol (24), with gene-specific primers listed in Table S4. ANOVA was performed to assess the significance of the results.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Antimicrobial Resistance Collaborators. 2022. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet 399:629–655. doi:10.1016/S 0140-6736(21)02724-037500506 · doi ↗ · pubmed ↗
- 2Sun S. 2025. Emerging antibiotic resistance by various novel proteins/enzymes. Eur J Clin Microbiol Infect Dis 44:1551–1566. doi:10.1007/s 10096-025-05126-440232578 PMC 12241128 · doi ↗ · pubmed ↗
- 3Liu K, Xiao T, Yang H, Chen M, Gao Q, Brummel BR, Ding Y, Huigens RW. 2023. Design, synthesis and evaluation of halogenated phenazine antibacterial prodrugs targeting nitroreductase enzymes for activation. RSC Med Chem 14:1472–1481. doi:10.1039/D 3MD 00204 G 37593580 PMC 10429720 · doi ↗ · pubmed ↗
- 4Machan ZA, Pitt TL, White W, Watson D, Taylor GW, Cole PJ, Wilson R. 1991. Interaction between Pseudomonas aeruginosa and Staphylococcus aureus: description of an anti-staphylococcal substance. J Med Microbiol 34:213–217. doi:10.1099/00222615-34-4-2131902262 · doi ↗ · pubmed ↗
- 5Garrison AT, Abouelhassan Y, Kallifidas D, Bai F, Ukhanova M, Mai V, Jin S, Luesch H, Huigens RW 3rd. 2015. Halogenated phenazines that potently eradicate biofilms, MRSA persister cells in non-biofilm cultures, and Mycobacterium tuberculosis. Angew Chem Int Ed Engl 54:14819–14823. doi:10.1002/anie.20150815526480852 · doi ↗ · pubmed ↗
- 6Garrison AT, Abouelhassan Y, Kallifidas D, Tan H, Kim YS, Jin S, Luesch H, Huigens RW 3rd. 2018. An efficient Buchwald-Hartwig/reductive cyclization for the scaffold diversification of halogenated phenazines: potent antibacterial targeting, biofilm eradication, and prodrug exploration. J Med Chem 61:3962–3983. doi:10.1021/acs.jmedchem.7b 0190329638121 · doi ↗ · pubmed ↗
- 7Garrison AT, Abouelhassan Y, Norwood VM 4th, Kallifidas D, Bai F, Nguyen MT, Rolfe M, Burch GM, Jin S, Luesch H, Huigens RW 3rd. 2016. Structure-activity relationships of a diverse class of halogenated phenazines that targets persistent, antibiotic-tolerant bacterial biofilms and Mycobacterium tuberculosis. J Med Chem 59:3808–3825. doi:10.1021/acs.jmedchem.5b 0200427018907 · doi ↗ · pubmed ↗
- 8Liu K, Brivio M, Xiao T, Norwood VM IV, Kim YS, Jin S, Papagni A, Vaghi L, Huigens RW III. 2022. Modular synthetic routes to fluorine-containing halogenated phenazine and acridine agents that induce rapid iron starvation in methicillin-resistant Staphylococcus aureus biofilms. ACS Infect Dis 8:280–295. doi:10.1021/acsinfecdis.1c 0040235089005 PMC 9004446 · doi ↗ · pubmed ↗