Molecular insights into Valencene synthase (SgTPS-V) and its role in sesquiterpenoid biosynthesis
Mohammed Ali

TL;DR
This paper identifies and characterizes a Valencene synthase gene in Salvia guaranitica, revealing its role in producing sesquiterpenes.
Contribution
The study provides the first molecular characterization of Valencene synthase in Salvia guaranitica and its function in sesquiterpene biosynthesis.
Findings
SgTPS-V was identified as a Valencene synthase in Salvia guaranitica based on in silico and in vitro analyses.
Overexpression of SgTPS-V in Arabidopsis increased flowering and altered chemical profiles.
The enzyme catalyzes the production of Valencene and related sesquiterpenes from FPP.
Abstract
Salvia species produce a diverse array of terpenoids, particularly monoterpenes and sesquiterpenes, with applications in medicine, perfumery, and industry. However, the genes involved in sesquiterpene biosynthesis in Salvia guaranitica remain poorly characterized. This study centers on the characterization of the Valencene synthase SgTPS-V from S. guaranitica. In silico analyses of SgTPS-V revealed conserved domains and motifs characteristic of terpene synthases, including DDxxxD, RWW, RxR, and NSE/DTE, and the phylogenetic tree placed it within the TPS-a subfamily, which encodes mono- and sesquiterpene synthases. Overexpression of SgTPS-V in Arabidopsis thaliana resulted in enhanced flowering relative to the wild-type. Also, comparing the chemical profiles of the transgenic Arabidopsis plants to wild-types indicated that SgTPS-V catalyzes the synthesis of Valencene, (-)-Valencene,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1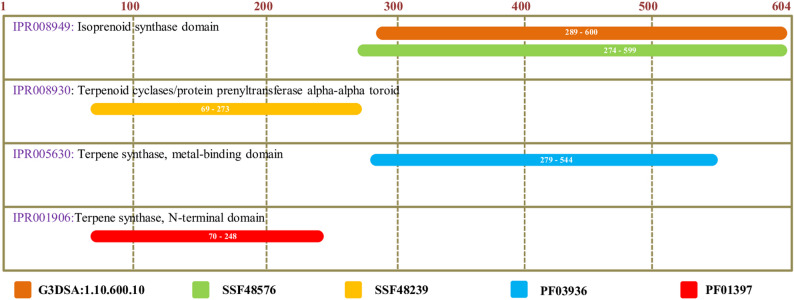 Figure 2
Figure 2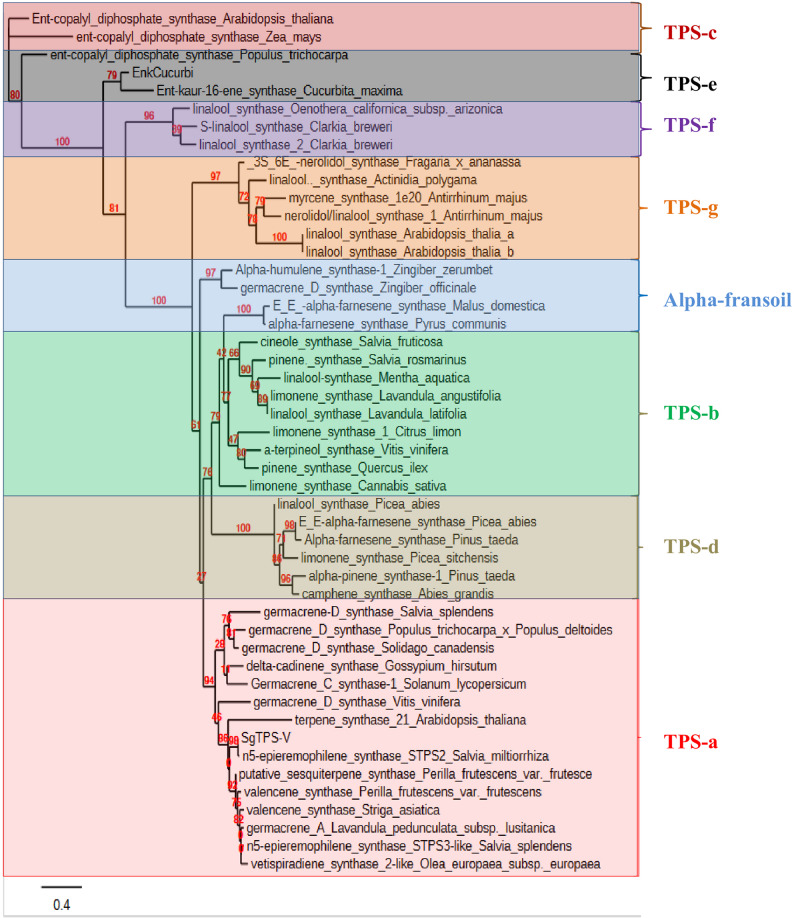 Figure 3
Figure 3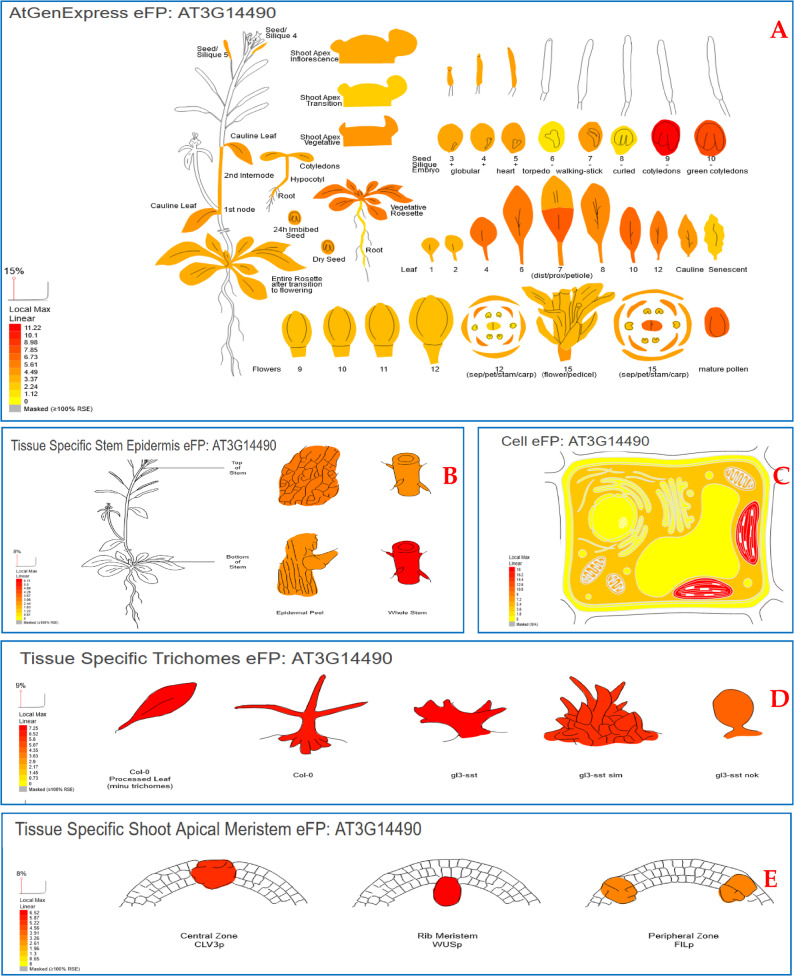 Figure 4
Figure 4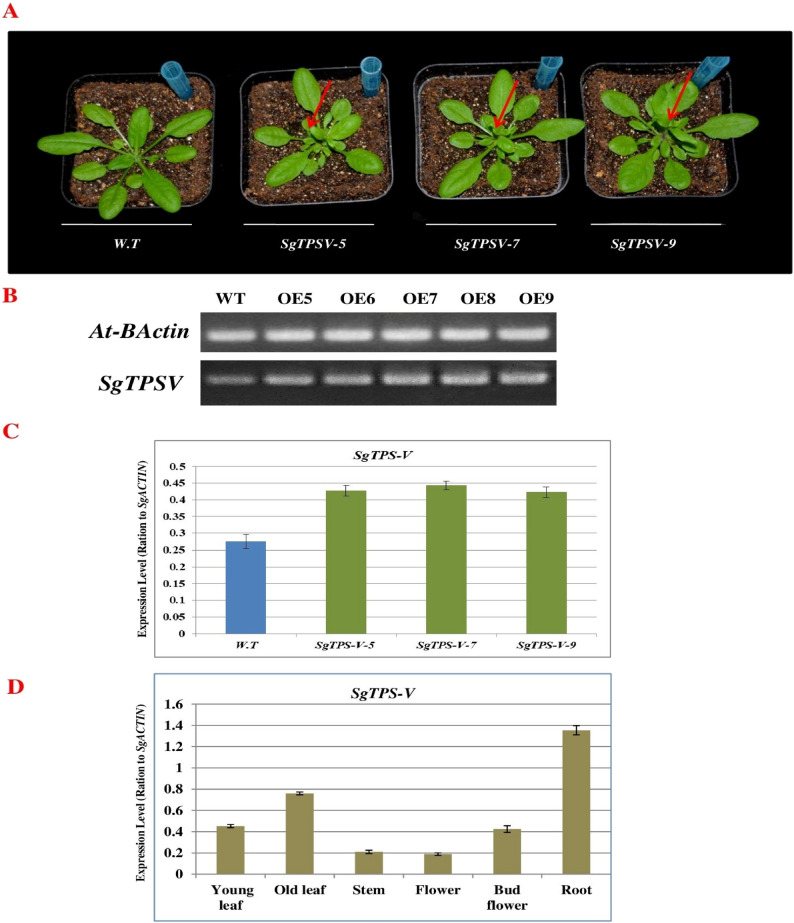 Figure 5
Figure 5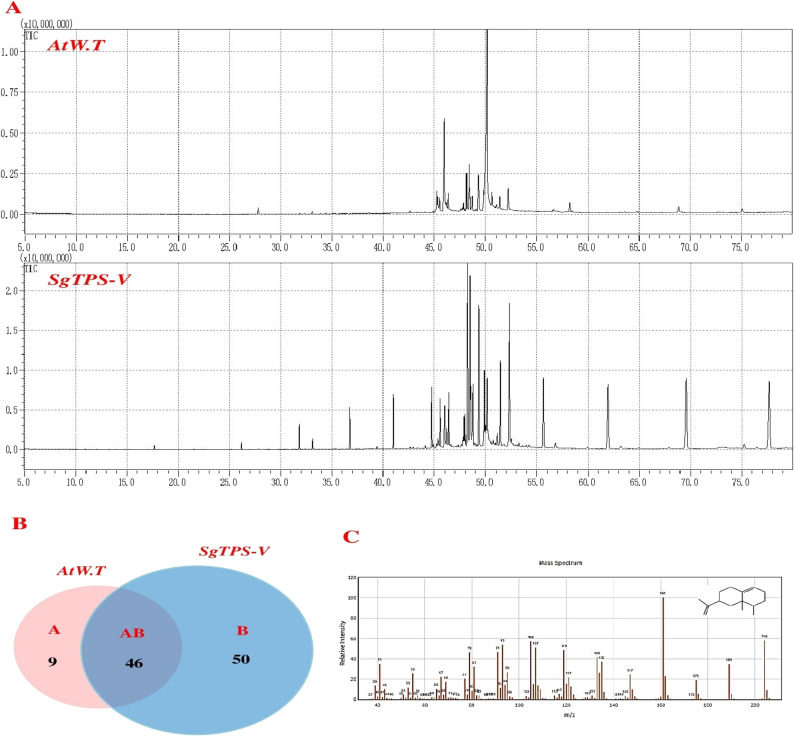 Figure 6
Figure 6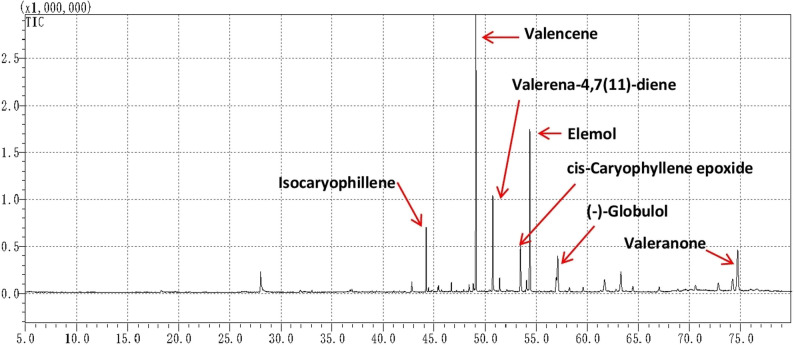 Figure 7
Figure 7- —Desert Research Center
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant biochemistry and biosynthesis · Sesquiterpenes and Asteraceae Studies · Traditional Chinese Medicine Analysis
Background
Salvia guaranitica L., an Asian plant, belongs to Lamiaceae family, and is distributed across various provinces in China [1]. Its tissues exhibit high concentrations volatile (essential) oils, including numerous terpene derivatives such as α-Pinene, Cis-α-terpineol, (-)-Camphor, β-Caryophyllene, laevo-β-Pinene, Germacrene-A, Thujone, Phytan, Squalene, Farnesan, Ledol, 1,8-cineol, and (E) Phytol [1, 2]. These compounds are beneficial for their anti-inflammatory, antibacterial, high-grade lubricant, antitumor, antiseptic, spasmolytic, antioxidant, analgesic, cardiovascular, anti-cholinesterase, and antidiabetic properties [1, 3–10].
Terpenes represent a significant category of secondary plant metabolites, encompassing over 60,000 distinct structures [11]. All of these compounds are derived from small five-carbon units via either the mevalonic acid (MVA) pathway or the methyl-D-erythritol phosphate (MEP) pathway. Subsequently, these block units are linearly coupled to generate geranyl diphosphate (GPP) and farnesyl diphosphate (FPP) through cyclization, rearrangement, besides various modifications [11, 12]. Diverse structures, including monoterpenes (C10H16), sesquiterpenes (C15H24), diterpenes (C20H32), sesterterpenes (C25H40), triterpenes (C30H48), sesquarterpenes (C35H56), and tetraterpenes (C40H64), were synthesized by various terpene synthase (TPS) enzymes utilizing geranyl diphosphate (GDP), farnesyl diphosphate (FDP), and geranylgeranyl diphosphate (GGDP) as substrates [9, 13–17]. Furthermore, numerous full and partial cDNAs for mono- (C10), sesqui- (C15), di- (C20), sester- (C25), tri- (C30), and tetraterpene synthase genes (C40) have been identified, cloned, and characterized from the Lamiaceae family, particularly within the genus Salvia [17–22]. The previously studied terpene synthase enzymes exhibit various putative motifs in their C-terminal and N-terminal domains, which are crucial for determining product specificity [1, 14, 20, 23–29].
In recent years, NGS-based RNA sequencing (RNA-Seq) has become an effective tool for providing information about the genes which are related with secondary metabolite pathways in various medicinal plants, especially plants that belong to the lamiaceae family [1, 20]. The information gained from RNA-Seq studies help researchers for identification and discovery of novel genes and their Key Enzymes [1, 20, 30]. Furthermore, the purpose of identifying these genes is not only to facilitate functional studies but also to develop biotechnology for improving the production of medicinal ingredients, plant growth and defense through metabolic engineering [1, 2, 9, 10, 30, 31]. Through our numerous previous studies, we have successfully identified, characterized, and quantified the expression levels of several genes which related to terpene synthesis in various plants (e.g., S. guaranitica, Salvia officinalis and Glycine max), based on the data obtained from RNA-Seq data analyses such as; SgCINS,* SgGPPS*,* SgFPPS*,* SgCMS*,* SgTPS-V*,* SgHUMS*,* SgNEOD-1*,SgNEOD-2,SgNEOD-3,SgTPS-1,SgTPS-3,SgTPS-6,,* SgTPS4*,* SgLINS-1*,SgLINS-2,* SgGLNS*,* SgGERIS*,* SgFARD*,* SoAMYS*,* SoFLDH*,* SoTPS3*,* SoTPS6*,* SoNEOD*,* SoLINS*,* SoSABS*,* SoGPS*,* SoCINS*,* GmFDPS* and GmGGPPS [1, 2, 9, 10, 30, 31].
Functional analysis of mono- and sesquiterpene synthase genes indicates that each terpene synthase gene typically produces specific substantial products. However, some of these genes possess the capability to synthesize varying amounts of products through deprotonation or cyclization of carbocations [1, 17, 20, 25–31]. Prior investigations elucidated the potential role of Salvia terpene synthases in terpene biosynthesis [1, 2, 17, 20, 29–32]. The article focus on the functional characterization of SgTPS-V, identified as the Valencene synthase gene, elucidating its biochemical role in sesquiterpene synthases in S. guaranitica.
Materials and methods
In silico Studies
The SgTPS-V was chosen as an authentic ortholog to other plant sesquiterpene syntheses [1]. Using the Arabidopsis transcript expression database, expression data for S. guaranitica sixty tissues were collected. The profiles of the transcript expression in different plant organs were created via eFP Browsers from Arabidopsis (eFP Browsers (http://bar.utoronto.ca/efp/cgi-bin/efpWeb.cgi). The profile expression pointers (arrows) show the degree of expression - red means high expression and yellow means lesser expression). Additionally, putative localizations of SgTPS-V from S. guaranitica was verified on the basis of Arabidopsis protein localization patterns at fourteen cell organelles to identify possible synthesis sites with Cell-eFP (http://bar.utoronto.ca/cell_efp/cgi-bin/cell_efp.cgi) eFP Browsers. Also, PROTPARAM Server (http://web.expasy.org/protparam/) elucidated the physiochemical characters of SgTPS-V. Moreover, iPSORT (http://ipsort.hgc.jp/), WolF PSORT Prediction (https://www.genscript.com/wolf-psort.html), TargetP-2.0 (https://services.healthtech.dtu.dk/services/TargetP-2.0/) and DeepLoc − 2.0 (https://services.healthtech.dtu.dk/services/DeepLoc-2.0/) were used to highlight SgTPS-V signal peptide based on the highest similarity scores with other plant signaling proteins. SgTPS-V was analyzed comparatively using the NCBI’s BLASTX to identify authentic homologs proteins (http://blast.ncbi.nlm.nih.gov/). For homology-based comparison, the online Clustal Omega was employed (https://www.ebi.ac.uk/Tools/msa/clustalo/). Also, a maximum-likelihood phylogenetic tree was constructed using the PhyML version 3.0 Server, with default settings and 1000-repetition bootstrapping to scrutinize the evolution of history of SgTPS-V with other orthologous plant TPS genes [32, 33]. And the protein sequences of functionally characterized TPS genes were retrieved from the National Center for Biotechnology Information (NCBI) database.
Plant material, cDNA library and cloning of the full-length region of cDNA
Complementary DNA (cDNA) was provided from our previous work on S. guaranitica [9], and the plant material was collected previously from the Wuhan Botanical Garden, Wuhan, China (Geo URI: geo:30.54445,114.42329). Concisely, TransZol Reagent was used to extract total RNA while cDNA was synthesized using the Super Mix kit ([1, 20, 34]. For gene cloning, the full-length cDNAs of SgTPS-V (GenBank: KX893974.1) was retrieved from NCBI for primer designing [1]. A pair of short and long degenerative primers was designed as short forward: 5′-ATGAGTGTTTTATTGTCAACAACTACT C-3′, short reverse: 5′-TTACGTTCTA TTATTGACACAAATAAAATA-3′, long forward: 5′-GGGGACAAGTTTGTACAAAAAAGCAGGCTTCATGAGTGTTTTAT TGTCA − 3′ and long reverse: 5′-GGGGACCACTTTGTACAAGAAAGCTGGGTTT ACGTTCTATTATTGAC-3′. Short primers and cDNA were used for the first PCR, after which the resultant PCR product was used along with long gene-specific primers. The final amplified fragments were then introduced in the pDONR221 vector (Invitrogen, Carlsbad, CA, USA) after purification [1, 20, 34]. Subsequently, the amplified SgTPS-V cDNA was transferred to the pB2GW7 vector (Invitrogen, Carlsbad, CA, USA). To authenticate the successful insertion, sequencing was performed using vector specific forward with SgTPS-V reverse primers for positive colonies selection, the recombinant vector preparation, transferred into bacteria, sequencing and cloning system were performed [1, 9, 20, 29–32].
Preparation of cDNA library from purified RNA
For Semi-quantitative RT-PCR and qRT-PCR, the total RNA from the leaves of wild type and transgenic A. thaliana were collected using Transzol Reagant (Focus Bioscience, Australia). The quantification and purity of the extracted RNA was assessed with NanoDrop™ 2000/2000c Spectrophotometers (Nanodrop technologies, Wilmington, DE, USA), and visualized on a 1.64% (W/V) agarose gel. Then, 1 µg RNA was used to synthesize cDNA via TransScript^®^ First-Strand cDNA Synthesis Super Mix Kit (TransGen Biotech Beijing, China) as of the manufacturer’s instructions [20, 34].
Genetic transformation
Arabidopsis thaliana ecotype Columbia-0 (Col-0) were cultivated according to Ali et al., 2018 and Mohammed et al., 2023a [1, 31]. Healthy pre-flower plants were selected for the floral-dip transformation experiment two months after growth. Agrobacterium tumefaciens strain GV101 received pB2GW7-SgTPS-V (expression vector) and pB2GW7 (empty vector). A. tumefaciens containing the expression vector was grown on Rif and Spc-supplemented solid LB medium. After screening, a positive colony was injected into 0.8 mL of liquid LB medium with the same antibiotics and components. Briefly, as stated by Ali et al. 2017; 2018 and Darwish et al. 2022) [1, 20, 35, 36], Agrobacterium cells were sub-cultured in Luria-Bertani medium or Lysogeny broth (LB medium) and incubated in a shaker incubator till they growth of 0.76 at the optical density at 600 nm (OD600). Then, a floral-dip inoculation medium with 5.2% sucrose and 0.055% Silwet was used to resuspend the pellet from Agrobacterium cell suspension. A. thaliana was individually introduced by immersing secondary inflorescence explants in fresh floral dip inoculation medium and applying gentle pressure to facilitate the uptake of A. tumefaciens vectors into the flowers’ gynoecia. A sequential culture regimen utilizing BASTA selection was applied to select the suitable plants from T0 and T1 and finally we successfully selected 12 transgenic A. thaliana transgenic lines from T2. Transgenic lines were investigated for leaf morphology, flowering time, and terpene metabolism.
Gene expression analysis
In verification of achieved gene transfer results, Semi-Quantitative RT-PCR (sqRT-PCR) was applied using an Eppendorf PCR system (Mastercycler Nexus PCR Machine from Eppendorf, Sydney, Australia). The housekeeping At-B-actin forward primer was 5’-GGCTGAGGCTGATGATATTC-3’ and reverse primer was 5’-CCTTCTGGTTCATCCBAAC-3’ to amplify 155 bp. However, SgTPS-V forward primer was 5’-GTTACTTCTGGGCGTTGG-3’ and the reverse one was 5’- CATCATCCCACCTCTGTA TTG − 3’ to amplify 168 bp. The PCR conditions were: 96 °C for 2 min, 94 °C for 35 s, 58.5 °C for 40 s, 72 °C for 30 Sect. (35 cycles), then a final extension at 72 °C for 10 min. Also, agarose gel was extra used to validate SgTPS-V expression analysis [37–40]. To analyze the expression of SgTPS-V at transgenic A. thaliana and wild type, qRT-PCR was performed using IQTM5 System, SYBR Green, and the cycler program as follows: 94 °C for 11 s, 58.5 °C for 30 s, and 72 °C for 20 s, then 66 °C for 5 s and 94 °C for 6 s. Moreover, the same qRT-PCR primers and condations were used to validate the expression of SgTPS-V gene under different tissues from S. guaranitica plant (e.g, young leaf, old leaf, stem, flower, bud flower and root). The expression levels were enumerated by comparing our target gene cycle thresholds (CTs) with the housekeeping gene SgACTIN using the 2^-ΔΔCt^ method (20, 31, 39). Values were offered as means ± SE of three different RNA pool replicates.
Terpenoids quantitative and qualitative analyses
GC-MS analysis was harnessed to compare terpenoids species in transgenic lines with wild type plants. For this, 12 leaves from each transgenic A. thaliana line or non-transgenic (three leaf from each plant) were homogenized in liquid nitrogen with a mortar and pestle, after which the plant material powder was directly soaked in n-hexane as a solvent in Amber storage bottles, 30 ml screw-top vials with silicone/PTFE septum lids (http://www.sigmaaldrich.com) were used to reduce loss of volatiles to the headspace then incubated with shaking at 37 °C and 200 rpm for 75 h. Afterward, the solvent was transferred using a glass pipette to a 10-ml glass centrifuge tube with screw-top vials with silicone/PTFE septum lids and centrifuged at 5,000 rpm for 10 min at 4 °C to remove plant debris [1,9, 20, 29, 30,]. A single microliter’s worth of the extract was put into a Shimadzu model GCMS-QP2010 Ultra and analyzed in triplicate. The Wiley-GC/MS-Library (10th Edition), the Volatile Organic Compounds (VOC) Analysis S/W software, and the NIST-Library (2014 edition) were used as references to identify various terpenoids [1, 2, 9, 20, 40–42].
Expression of Valencene synthase (SgTPS-V) recombinant proteins in Escherichia coli and enzyme activity assay
The full-length sequences of SgTPS-V gene was transferred to the pDEST17 Gateway^®^ destination vector. To authenticate the successful insertion, sequencing was performed using vector specific forward with SgTPS-V reverse primers for positive colonies selection, the recombinant vector preparation, transferred into Escherichia coli)E. coli( strain BL21 (DE3) bacteria, sequencing and cloning system were performed [1, 9, 20, 29–32]. The recombinant proteins were induced under various concentration from isopropyl-β-d-thiogalactopyranoside (IPTG) (e.g., 0.1 mM, 0.2 mM, 0.3 mM and 0.4 mM) at 26 °C for 15 h. After induction, the bacteria cells were harvested by centrifugation under cooling, then the bacteria cell pellets will resuspended in phosphate-buffered saline, and disrupted by sonication machine. The crude proteins were then extraction using BugBuster^®^ Ni-NTA His•Bind^®^ Purification Kit (Milipore) according to the manufacturer’s instructions. The purified proteins were collected and concentrated before enzyme assays and they were examined by SDS–PAGE. as described previously [43–46].
The Valencene synthase activity assays were conducted as described [43–46] with some modifications. In briefly, the reaction mixture consist from 25 mM HEPES buffer (pH 7.4), 15 mM MgCl2, 5 mM dithiothreitol, 50 µg purified protein, 2 mM FPP (Sigma-Aldrich) as substrate, in a totally volume of 100 µl. The reaction mixtures were incubated at 30 °C for 2 h. After incubation, extraction of reaction products relied on n-hexane 250 µl, and resulting extract 2 µl was sent to GC-MS analysis. GC-MS conditions were consistent with that described a bove at terpenoids quantitative and qualitative analyses.
Results
Cloning and sequence analysis of the full-length SgTPS-V
The entire open reading frame of the SgTPS-V, comprising 1829 base pairs, encodes a protein of 604 aa, which is anticipated to have a theoretical IP of 5.35 and a molecular weight of 70.70 kDa. The hypothesized amino acid sequence of the SgTPS-V features a signal peptide that is shorter than those found in monoterpene synthases (600–650 aa) and sesquiterpene synthases (550–580 aa). Furthermore, our target protein sequence aligns with numerous sesquiterpene synthases due to the presence of a 30 amino acid long target sequence (MSVLL STTTQ PKFGI FRHIY TMSTV NSNSF) at the N-terminal. The application of the ‘iPSORT’ program, WolF PSORT Prediction, TargetP-2.0 and DeepLoc − 2.0 databases have elucidated that SgTPS-V protein have mainly localized in the Plastid, which maybe because the origin of FPP and its biosynthetic processes are in these organelles. The BLASTX analysis presented in Table 1 reveals that SgTPS-V exhibits a remarkable identity ranging from 91.45% to ≥ 66.60% with its homologous sesquiterpene synthase protein derived from Salvia splendens and various plant species.Table 1NCBI BLASTX results of SgTPSV to retrieve authentic homologous proteinsNCBI Accession^a^ DescriptionOrganismE valueIdentity (%)Accession lengthXP_042060897.1(-)−5-epieremophilene synthase STPS3-likeSalvia splendens091.45%550XP_042060899.1(-)−5-epieremophilene synthase STPS3-likeSalvia splendens084.39%551AAX16076.1Putative sesquiterpene synthasePerilla frutescens var. frutescens069.56%550AGN72800.1Germacrene ALavandula pedunculata subsp. lusitanica070.93%547XP_022850114.1Vetispiradiene synthase 2-likeOlea europaea var. sylvestris068.27%554QIQ55988.1Putative terpene synthase 2Eremophila denticulata subsp. trisulcata068.06%557XP_011073749.1Vetispiradiene synthase 3Sesamum indicum067.70%554QNC49793.1Terpene synthase 12aLeucophyllum frutescens066.61%553A0A1W6GW18.1RecName: (-)−5-epieremophilene synthaseSalvia miltiorrhiza068.01%546A0A1W6GW06.1RecName: (-)−5-epieremophilene synthaseSalvia miltiorrhiza068.01%546AGN72803.1Germacrene ALavandula stoechas069.39%542AAX16077.1Valencene synthasePerilla frutescens var. frutescens067.22%550A0A1W6GW32.1RecName: (-)−5-epieremophilene synthaseSalvia miltiorrhiza067.65%546AGN72806.1Germacrene ALavandula viridis069.26%543CAA3024998.1Vetispiradiene synthase 2-likeOlea europaea subsp. europaea066.67%555GER46991.1Valencene synthaseStriga asiatica066.60%556^a^Description—homology search using BLASTX
The alignment of SgTPS-V with other authentic orthologous sequences facilitated the inference of its potential function. According to this prediction, the SgTPS-V exhibits a variety of motifs, including DDEVYD (residues 121–126), RWW (residues 271–273), RxR (residues 286–288), RxR (residues 307–309), DDxxD (residues 323–327), NSE/DTE (residues 466–474), and DxxxD (residues 508–512). These motifs are prevalent in analogous sesquiterpene synthases associated with Valencene synthase [1, 9, 20, 27, 29, 47, 48] (Fig. 1).Fig. 1. Multiple sequence alignment. The deduced amino acid sequence of SgTPS-V was aligned with homologues identified from the BLASTX analysis. The conserved motifs DDEVYD, RWW, RxR, DDxxD, NSE/DTE and DxxxD are marked. SgTPS-V: Valencene synthase from *S. guaranitica *plant
In comparison to other sesquiterpene synthases, SgTPS-V is characterized by five distinct domains, as delineated by the InterPro protein sequence analysis and classification database (InterPro: https://www.ebi.ac.uk/interpro/). The SgTPS-V protein comprises five distinct domains belonging to the terpene synthase family: the Terpenoid cyclases/protein prenyltransferase alpha-alpha toroid (IPR008930: spanning residues 69 to 273), the Terpene synthase N-terminal domain (IPR001906: spanning residues 70 to 248), the Terpene synthase metal-binding domain (IPR005630: spanning residues 279 to 544), the Farnesyl Diphosphate Synthase (1.10.600.10: spanning residues 289 to 600), and the Isoprenoid synthase domain (IPR008949: spanning residues 274 to 599) (see Fig. 2). Furthermore, the SgTPS-V has been categorized within the TPS-a subfamily of angiosperm sesquiterpene synthases, as indicated by the results of the phylogenetic analysis (Fig. 3).
Fig. 2. Putative domain analysis for SgTPS-V using the InterPro protein sequence analysis & classification (https://www.ebi.ac.uk/interpro/) database. SgTPS-V protein sequence has four protein family domains
Fig. 3. Phylogenetic tree of SgTPS-V with selected terpene synthases from other plants. Seven previously identified TPS subfamilies (Tps-a to Tps-g) were chosen based on Degenhardt et al., 2009; Abdelhameed et al., 2024; Esraa et al., 2022; Nasir et al., 2021 [49–52]. The alignment was performed using the PhyML server. The numbers indicated are the actual bootstrap values of the branches
In Silico expression profiling and subcellular localization of SgTPS-V
A BlastP search was conducted using the SgTPS-V protein sequence as a query to investigate the hypothesized tissue expression pattern of SgTPS-V in the A. thaliana genome at the Phytozome database (https://phytozome.jgi.doe.gov/pz/portal.html#!search? show=BLAST&method=Org_Athaliana). This study found other proteins closely associated with the SgTPS-V sequences, particularly (AT3G14490), which had a high BLAST score and e-value of 351 and 1.62e-113, respectively. The hypothesized tissue expression patterns of the SgTPS-V in Arabidopsis, revealed by our data, were analyzed across forty-five tissues using the BAR database (http://bar.utoronto.ca/efp/cgi-bin/efpWeb.cgi) (Fig. 4A). The Arabidopsis eFP Browsers indicated that SgTPS-V (AT3G14490) was predominantly expressed in various tissues, particularly in Seeds Stage 9 without Siliques (11.22), Seeds Stage 10 without Siliques (8.03), Leaf 7, Proximal Half (7.28), Mature Pollen (7.07), Flower Stage 15, Carpels (6.73), Rosette Leaf 12 (6.72), and Rosette Leaf 4 (6.37) (Fig. 4a and Supplementary Table S1). Conversely, this gene exhibited significant expression in tissue-specific stem epidermis, particularly at the apex of the stem (3.28), followed by the entire stem, apex of the stem (2.72), stem epidermis, base of the stem (2.69), and the entire stem, base of the stem (6.11) (Fig. 4B and Supplementary Table S1).
Furthermore, Arabidopsis Cell Electronic Fluorescent Pictograph tool (eFP; http://bar.utoronto.ca/cell_efp/cgi-bin/cell_efp.cgi) was used to predict the putative subcellular localizations of our SgTPS-V (AT3G14490) gene according to the protein localization of different cell organelles in the Arabidopsis. The subcellular localization profiles showed that SgTPS-V protein was highly expressed and presented in the Plastid (see Fig. 4C). Also, it was present with highly expressed in all tissue specific trichomes (Fig. 4D and Supplementary Table S1). Ultimately, our target gene exhibited high expression in the tissue-specific shoot apical meristem, particularly in the Central Zone (5.38), followed by the Rib Meristem (6.52) and the Peripheral Zone (3.16) (Fig. 4E and Supplementary Table S1).
Fig. 4. Visualization the putative an “electronic fluorescent pictograph” browsers for exploring the putative tissue expression and cell localization of SgTPS-V (AT3G14490) gene, based on Arabidopsis gene expression and protein localization at different tissues and cell organs. a Expression data at different tissues from seedling to flowering stages. b Expression data of tissue specific stem epidermis at top and bottom. c Expression data at different cell organs. d Expression data tissue at specific trichomes. e Tissue Specific Shoot Apical Meristem. The color box represents the expression scale (the more intense red color the more gene expression)
Characterization of SgTPS-V Activity in transgenic Arabidopsis thaliana leaves and validation of the SgTPS-V gene expression patterns by quantitative RT-PCR
The SgTPS-V was cloned from S. guaranitica to examine its impact on the phenotypes of A. thaliana plants after 36 days of growth. Subsequently, A. thaliana was employed as a transient bio-expression system to overexpress SgTPS-V. We employed the A. tumefaciens-GV101 containing the vector pB2GW7-SgTPS-V, regulated by the 35 S-promoter, to produce transgenic A. thaliana plants that constitutively overexpress the SgTPS-V (Fig. 5A). The positive transforming lines were then validated using both BASTA reagent and semi-quantitative RT-PCR of the cDNA (Fig. 5B). QRT-PCR was used to inspect the transcription levels of SgTPS-V at transgenic A. thaliana and wild type. (Fig. 5C). From the qRT-PCR analysis results, it was found that the highest expression levels were observed in various transgenic A. thaliana lines in compared with the wild type (Fig. 5C). Moreover, qRT-PCR was used to validate the expression patterns of SgTPS-V gene across different tissues of S. guaranitica such as; young leaf, old leaf, stem, flower, bud flower and root, to understand their expression profiles within the previous various tissues samples (Fig. 5D). Transgenic A. thaliana plants exhibited an increase in leaf width and an earlier onset of flowering stem growth compared to the wild type plants.
Fig. 5. Overexpression of SgTPS-V gene in transgenic Arabidopsis. A Comparison of the phenotypes of the transgenic A. thaliana and wild type A. thaliana. The red arrow (↘) indicates the flowers at the transgenic A. thaliana. B Semiquantitative RT-PCR to confirm the expression of terpenoid genes. C Quantitative RT-PCR validation of expression of SgTPS-V gene at transgenic A. thaliana and wild type. D Quantitative RT-PCR validation of expression of SgTPS-V gene under different tissues of S. guaranitica Plant
Profiling terpene contents
To investigate the impact of SgTPS-V overexpression in the leaves of A. thaliana, the qualitative and quantitative alterations in terpene profiles were monitored through GC-MS analysis. The findings indicated that multiple terpenes exhibited a significant increase in transgenic A. thaliana leaves with SgTPS-V overexpression, in comparison to the control, as detailed in Table 2 and illustrated in Fig. 6A. In leaves of A. thaliana plants upregulating SgTPS-V, sesquiterpene compounds constituted the predominant group at 48.08%, followed by diterpene compounds at 8.42% and monoterpene compounds at 0.51%. Additionally, one triterpene compound was represented at 0.15%. In non-transgenic A. thaliana (control), sesquiterpene compounds constituted the predominant group at 14.85%, followed by diterpene compounds at 10.26% and monoterpene compounds at 0.1%, see (Table 2). The hexane extracts from transgenic and non-transgenic A. thaliana exhibit distinct, shared, and predominant compounds (Table 2 and Fig. 6B).
Fig. 6. Typical GC-MS mass spectrographs for terpenoids from leaf of A. thaliana plants. A GC-MS Peak of the essential oil, (B) Two-way Venn diagram to show the number of unique and common compounds in the essential oil extracts from the transgenic and non-transgenic A. thaliana. (C) The Mass spectrum (electron ionization) and structure of valenene
Table 2. Principal terpenoids detected in Transgenic A. thaliana leaves that overexpress SgTPS-V N CompoundsR.T (min.)FormulaMolecular Mass(g mol-1)Type% Peak areaAtW.TSgTPS-V1(8)Annulene5.963C8H8104.14910.060.032Undecane17.649C11H24156.3080.010.213α-Terpinenyl acetate26.194C12H20O2196.286Mono0.010.44Patchulane31.833C15H26206.367Sesqui0.041.355Trans-β-Ionone32.304C13H20O192.29730.060.036Topanol33.074C15H24O220.3505Sesqui0.180.587All-trans-Geranylgeraniol35.132C20H34O290.4834Diter0.038p-Mentha-6,8-dien-2-ol, cis37.027C10H16O152.2334Mono0.099Isovaleric acid p-tolyl ester37.369C12H16O2192.2542Mono0.0110Guaia-1(10),11-diene44.898C15H24204.3511Sesqui0.090.1311β-cedren-9-α-ol45.113C15H24O220.3505Sesqui0.280.1312Cedrenol47.234C15H24O220.3505Sesqui0.1813β-Carotene47.66C40H56536.8726Triter0.1514Ledol47.774C15H26O222.3663Sesqui0.240.5515(+)-Valencene47.873C15H24204.3511Sesqui0.661.7816Kolavenol acetate47.989C22H36O2332.52Diter0.150.6317Geranylgeraniol48.136C22H36O2332.52Diter3.8818Valencene48.394C15H24204.3511Sesqui4.0913.2419(-)-Valencene48.464C15H24204.3511Sesqui1.169.1320Valencene (isomer I)48.692C15H24204.3511Sesqui1.873.25214,8,13-Duvatriene-1,3-Diol49.272C20H34O2306.4828Diter2.6322Phytol49.319C20H40O296.531Diter3.67.623Viridiflorine49.825C15H24204.3511Sesqui2.114.0424Linolenic acid50.164C18H30O2278.429644.781.13254,8,13-Duvatriene-1,3-Diol50.5C20H34O2306.4828Diter0.1626(+)-Ledol51.048C15H26O222.3663Sesqui0.410.6627(-)-Ledol52.208C15H26O222.3663Sesqui2.957.8328Epiglobulol52.523C15H26O222.3663Sesqui0.4229Ledol, isomer 152.677C15H26O222.3663Sesqui0.1230trans-caryophyllene oxide53.221C15H24O220.3505Sesqui0.231α-Elemol53.954C15H26O222.3663Sesquit0.132Elemol54.147C15H26O222.3663Sesqui0.090.1233Z-Citronellyl tiglate55.724C15H26O2238.3657Sesqui0.093.834Ledane56.633C15H26206.3669Sesqui0.290.0335β-Elemol56.961C15H26O222.3663Sesqui0.30.2636Isovaleric anhydride73.242C10H18O3186.2481Mono0.0837(-)-Myrtenol78.184C10H16O152.2334Mono0.02Total % of monoterpene0.10.51Total % of sesquiterpene14.8548.08Total % of diterpene10.268.42Total % of triterpene0.15
Measurements of enzyme activity
To express SgTPS-V proteins in E. coli, the full-length sequences of SgTPS-V genes were amplified with Gateway method and then sub cloned into the pDEST17 Gateway^®^ destination vector. Then the recombinant proteins induced various concentrations from IPTG and from our results we found the optimal induction concentration and condition was 0.4 mM at 26 °C for 15 h. Furthermore, the crude proteins were then extraction using BugBuster^®^ Ni-NTA His•Bind^®^ Purification Kit (Milipore) and the purified proteins were isolated and electrophoresed by SDS–PAGE see Supplementary Fig. S1. Moreover, enzymatic assays of the crude SgTPS-V E. coli extracts showed that a fully functional protein was being produced and the reaction products were extracted in 250 µl of hexane and the reaction products were running using GC-MS for analysis. And from GC-MS analysis we found the major compound produce by SgTPS-V enzyme in vitro assay is Valencene (38.01%) at 48.923RT, followed by Elemol (12.06%) at 54.147 RT, Valerena-4,7(11)-diene (9.47%) at 50.65 RT, Isocaryophillene (7.12%) at 44.15 RT, cis-Caryophyllene epoxide (5.1%) at 53.221 RT, Valeranone (4.35%) at 74.53RT, and (-)-Globulol (3.86%) at 57.025 RT (see Fig. 7).Fig. 7GC-MS of the products generated in vitro by recombinant SgTPS-V protein. In vitro enzymatic activity analysis of SgTPS-V protein using FPP as substrate Products catalyzed by SgTPS-V protein using FPP as substrate were subjected to GC-MS. Peaks were identified as terpenes by comparing mass spectra with various library such as Wiley-GC/MS-Library (10th Edition), the Volatile Organic Compounds (VOC) Analysis S/W software, and the NIST-Library (2014 edition), that used as references to identify various terpenoids
Discussion
The complete cDNA of the SgTPS-V was identified and extracted from the pool of leaves of S. guaranitica, utilizing in silico sequence comparison with other orthologs from S. splendens,* Lavandula pedunculata* subsp. lusitanica, Salvia miltiorrhiza, L. stoechas, and L. viridis. In comparison to other sesquiterpene synthases, the SgTPS-V contains five domains, as identified by the InterPro protein sequence analysis and classification database (InterPro: https://www.ebi.ac.uk/interpro/). The first domain is the Terpenoid cyclases/-protein prenyltransferase alpha-alpha-toroid (IPR008930: residues 69–273), followed by the Terpene synthase, N-terminal domain (IPR001906: residues 70–248). The third to fifth domains include the Terpene synthase, metal-binding domain (IPR005630: residues 279–544), Farnesyl Diphosphate Synthase (1.10.600.10: residues 289–600), and the Isoprenoid synthase domain (IPR008949: residues 274–599) (Fig. 2). A comparison of the putative SgTPS-V with other orthologous proteins revealed several highly conserved motifs, including DDxxD (residues 323–327) and DxxxD (residues 508–512). These conserved motifs are located close to the active site. Therefore, they may stabilize the binding of the substrate and the inorganic cofactor [1, 20, 27, 48, 49, 53, 54] (Fig. 1).
Additionally, these conserved motifs stabilize a tri-nuclear magnesium cluster [27, 53, 55], which in turn catalyzes the formation of the C15-substrate-FPP at the hydrophobic substrate-binding-pocket [27, 49, 56]. Additionally, RWW (residues 271–273), RxR (residues 286–288), and RxR (residues 307–309) are essential for product cyclization in class-III-TPS proteins [48, 57, 58] (Fi. 1). The in-silico studies identified also additional conserved region motifs in the SgTPS-V, specifically the DDEVYD (residues 121–126) and NSE/DTE (residues 466–474) regions, which are prevalent in other sesquiterpene synthases. Each protein sequence within the terpene synthase family contains one or two conserved domains and motifs [1, 17, 20]. The phylogram was constructed to examine the phylogenetic relationships among SgTPS-V and other sesquiterpene synthases, utilizing the neighbor-joining method for optimal parameter selection. The results indicate that SgTPS-V belongs to the TPS-a subfamily, which is capable of encoding sesquiterpenes synthase. This result supports the previously noted ability of SgTPS-V to produce various types of terpenes [1, 20] (Fig. 3).
The in-silico expression patterns of SgTPS-V across forty-five tissues were investigated harnessing the high similarity between SgTPS-V and its homologs sequence from A. thaliana (AT3G14490). The SgTPS-V in-silico expression patterns were consistent with findings from Ali et al. (2017, 2018, 2022a) [1, 9, 20], which indicated that the majority of TPS genes (e.g., SoFLDH,* SgTPSV*,* SgGERIS*,* SoLINS-2*,* GmTPS-21*,* SgFARD*,* SoNEOD*, and SoHUMS) from S. officinalis, Glycine max, and S. guaranitica exhibited high transcripts in seeds, leaves, mature pollen, stems, trichomes, and flowers. The proposed subcellular localization of the SgTPS-V protein indicates that it is primarily found in the plastids, mitochondria, and cytosol. The findings align with those of [1, 9, 20, 29–32, 59–66](Fig. 4).
To investigate the role of SgTPS-V in A. thaliana Columbia-0 (Col-0), we overexpressed SgTPS-V in A. thaliana integrating Agrobacterium with the pB2GW7-SgTPS-V vector. The SgTPS-V expression in successful transformed lines was validated through sqRT-PCR (Fig. 5). The transgenic lines exhibited significantly elevated SgTPS-V expression levels versus WT plants, supporting successful introduction of the transgene. Three independent transgenic lines, designated OE-SgTPS-V-5, OE-SgTPS-V-7, and OE-SgTPS-V-9, were selected for terpene analysis. Morphological assessments revealed that these recombinant plants exhibited an accelerated reproductive shoot formation relative to the wild-type (WT) (Fig. 5). In addtion, we used qRT-PCR to elucidate the expression level of SgTPS-V gene across different tissues of S. guaranitica such as; young leaf, old leaf, stem, flower, bud flower and root, and from our results we found that the SgTPS-V gene was highly expression at root tissue, followed by old leaf, young leaf, bud flower, stem and flower. And this previous results are in line with our previous results [1].
These results corroborate previous research by Ali et al., 2017, 2018, 2022a [1, 9, 20], which demonstrated that the upregulation of terpene synthase genes (TPS), such as SgGPS,* SoCINS*,* SgFPPS*,* SoTPS6*,* SoLINS*,* SgLINS*,* SoFLDH*,* SgGPS*,* SoSABS*, and SoNEOD from S. guaranitica and S. officinalis in A. thaliana and Nicotiana tabacum, enhanced vegetative and reproductive development against the WT plants. Prior research indicates that various TPS family members play roles in distinct cellular mechanisms, for example, 1,−8-cineole synthase acting as a monoterpene synthase, Z–γ-bisabolene synthase functioning as a sesquiterpene synthase, rhizathalene synthase (AtTPS08) contributing to diterpene biosynthesis, and β-amyrin and thalianol synthases participating in triterpene formation [61–64, 67–71].
Moreover, the co-expression of TPS genes in various cells, tissues, and organs highlights their role in shaping diverse plant phenotypes, reinforcing their significance in plant ontogeny [61–64, 68–72]. Gas-chromatography-mass spectrometry (GC-MS) identified terpenes produced following SgTPS-V overexpression. The resulting transformant lines exhibited distinct peaks corresponding to mono-, sesqui-, and diterpenes, with the percentage peak area indicating the types and relative abundance of these compounds. Terpene identification was carried out using mass spectral libraries, including the Wiley GC/MS and NIST Library, in addition to comparative analysis with wild-type A. thaliana extracts.
The GC/MS results (Table 2; Fig. 6) revealed an emergence of two peaks at a retention time of 48.394 and 48.464 min, which were identified as Valencene and (-)-Valencene based on spectral matching with reference databases. However, the comparative analysis of hexane extracts from transgenic and non-transgenic A. thaliana revealed distinct, shared, and predominant compounds. The transgenic extract contained unique compounds, along with 46 compounds that were also present in the non-transgenic extract. In the context, the transgenic extract exhibited 50 unique compounds and 11 compounds from them are terpenoids (e.g., All-trans-Geranylgeraniol, Isovaleric acid p-tolyl ester, Cedrenol, β-Carotene, 4,8,13-Duvatriene-1,3-Diol, Epiglobulol, Ledol, isomer 1, trans-caryophyllene oxide, α-Elemol, Isovaleric anhydride and (-)-Myrtenol) (Fig. 6B and supplementary Table 2). The production of Valencene, (-)-Valencene, Valencene (isomer I), (+)-Valencene, Viridiflorine, (-)-Ledol and Patchulane as a sesquiterpene in transgenic A. thaliana is consistent with previous reports [1, 48]. Terpene synthases are known to catalyze the synthesis of multiple terpenes, including carene synthases, (±)-linalool synthases, cineole, myrcene, β-amyrin, and terpinolene synthases [25–28, 73–75]. Therefore, SgTPS-V is suggested to facilitate valencene and other isoformas biosynthesis via the isoprenoid pathway, a well-characterized mechanism in sesquiterpene production.
In the context, for measurements of enzyme activity the E. coli BL21 (DE3) cells expressing SgTPS-V protein using the expression vector pDEST17 were harvested and lysed. Upon incubation with (2E,6E)-farnesyl diphosphate (FPP) as a substrate, SgTPS-V catalyzed the formation of Valencene, and other sesquiterpenes such as; Elemol, Valerena-4,7(11)-diene, Isocaryophillene, cis-Caryophyllene epoxide, Valeranone, and (-)-Globulol that were detected by GC-MS analysis (Fig. 7). The identities of these previous compound peaks were confirmed by comparisons with retention times (R.T; min) and the mass spectra (Molecular Mass; g mol-1) of various libraries. Based on results the major product for SgTPS-V was designated as a Valencene, see Fig. 7. And from our results we found the SgTPS-V belongs to the class of sesquiterpene enzymes which have the ability to catalyze the formation of Valencene with higher levels. At the end, these results confirmed the significant epistatic relationship that was found between the release of Valencene as a sesquiterpene and SgTPS-V gene expression.
In the end, economics studies refer to the great market demand for valencene and its global annual demand exceeds 10,000 kg with market value estimated to grow to 9.2 million dollars by 2033 [76, 77]. Valencene is widely used in the food, beverage, flavor, and a cosmetic fragrance ingredient with extensive applications [76, 77]. In the last decade, valencene supply was primarily relies on extraction from natural sources (e.g., fresh citrus,, woody orange notes, grapefruit peel, other citrus fruits, Myrica rubra, Cyperus rotundus and Alpinia oxyphylla Miq) or chemical synthesis, but the extraction process is affected by some limitations [76, 78]. In Addition, in plant Valencene can also be readily converted to (+)-nootkatone by chemical or biological means, which is not only a commonly used flavor compound in soft drinks but also can be used as mosquito repellant [76, 78]. Several studies used the valencene compound level as an indicator of good quality of orange fruit, orange oil, fruit maturation, fruit development period and the level of the citrus greening disease [79, 80]. Furthermore, in plant valencene synthase can be affected by the plant hormone stimuli such as ethylene, development stages, irrigation conditions, and synthesis of other volatile terpenoid products [81, 82]. Moreover, the increased demand for valencene has attracted considerable attention from researchers to develop ways for increasing Valencene production by transgenic plants or by novel microbial cell factories for more efficient and sustainable production modes. As a result, this study provides a clearer vision through study of the function of the SgTPS-V gene from S. guaranitica as a new source of valencene synthese.
Conclusion
In this study, we successfully cloned and characterized SgTPS-V, a valencene synthase gene from S. guaranitica, a medicinal plant with significant pharmacological properties. Overexpression of SgTPS-V in A. thaliana accelerated flowering in transgenic lines (OE-SgTPS-V−5, OE-SgTPS-V−7, and OE-SgTPS-V−9) and led to increased valencene production, confirming that A. thaliana can synthesize valencene via the mevalonate (MVK) pathway of sesquiterpene biosynthesis. Computational analysis indicated that SgTPS-V displayed significant expression levels across multiple tissues and its product can be found in plastids and mitochondria, underscoring its role in terpene biosynthesis. These findings not only show that A. thaliana is a robust model for functional studies of sesquiterpene synthases but also provide a foundation for enhancing ornamental properties in the target species via synthetic biology.
Supplementary Information
Supplementary Material 1.
Supplementary Material 2.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kamatoua GPP, Makungab NP, Ramogolab WPN, Viljoen AM. South African salvia species: A review of biological activities and phytochemistry. J Ethnopharmacol. 200;119:664–72.10.1016/j.jep.2008.06.03018640254 · doi ↗ · pubmed ↗
- 2Takano A, Okada H. Phylogenetic relationships among subgenera, species, and varieties of Japanese salvia L. (Lamiaceae). J Plant Res.2011; (124): 245–52.10.1007/s 10265-010-0367-920628783 · doi ↗ · pubmed ↗
- 3Fateme AM, Mohammad HF, Abdolhossein R, Ali Z, Maryam S. Volatile constituents of Salvia compressa and Logochilus macranthus, two labiatae herbs growing wild in Iran. Res J Recent Sci 2013; (2): 66–8.
- 4Zhenqing B, Wenrui L, Yanyan J, Zhiyong Y, Jie J, Wenli H, Pengguo X, Zongsuo L. The ethylene response factor Sm ERF 6 co-regulates the transcription of Sm CPS 1 and Sm KSL 1 and is involved in Tanshinone biosynthesis in Salvia miltiorrhiza hairy roots. Planta. 2018; (248): 243–55.10.1007/s 00425-018-2884-z 29704055 · doi ↗ · pubmed ↗
- 5Yoko I, David RG, Eyal F, Efraim L, Eran P. Characterization of geraniol synthase from the peltate glands of sweet Basil. Plant Physiol. 2004; (134): 370–9.10.1104/pp.103.032946 PMC 31631614657409 · doi ↗ · pubmed ↗
- 6Shimada T, Endo T, Fujii H, Hara M, Omura M. Isolation and characterization of (E)-beta-ocimene and 1, 8 cineole synthases in Citrus Unshiu Marc. Plant Sci. 2005;(168):987–95.
- 7Abbas F, Yanguo K, Rangcai Y, Yanping F. Functional characterization and expression analysis of two terpene synthases involved in floral scent formation in Lilium. ‘Siberia ’ Planta. 2019;(249):71–93.10.1007/s 00425-018-3006-730218384 · doi ↗ · pubmed ↗
- 8Lucker J, El Tamer MK, Schwab W, Verstappen FW, van der Plas LH, Bouwmeester HJ, Verhoeven HA. Monoterpene biosynthesis in lemon (Citrus limon). c DNA isolation and functional analysis of four monoterpene synthases. Eur J Biochem. 2002; (269):3160–71.10.1046/j.1432-1033.2002.02985.x 12084056 · doi ↗ · pubmed ↗