Fosfomycin at sub-minimum inhibitory concentration impairs biofilm and efflux pump activity in multidrug-resistant Klebsiella pneumoniae isolates
Marwa W. Moustafa, Tarek E. El-Banna, Fatma I. Sonbol, Maisra M. El-Bouseary

TL;DR
Fosfomycin at low doses can weaken biofilms and reduce efflux pump activity in drug-resistant Klebsiella pneumoniae, suggesting it could help fight antibiotic resistance.
Contribution
This study shows fosfomycin at sub-MIC levels impairs biofilm and efflux pump activity in MDR Klebsiella pneumoniae.
Findings
Sub-MIC fosfomycin significantly reduced biofilm formation in MDR K. pneumoniae isolates.
Fosfomycin downregulated biofilm-related genes (fimH, mrkD) and efflux pump genes (acrA, acrB).
SEM revealed morphological changes, including cauliflower-like deformation in treated isolates.
Abstract
Klebsiella pneumoniae plays a critical role in hospital-acquired infections, which pose a serious threat globally. Older antibiotics such as fosfomycin are being reconsidered to combat MDR K. pneumoniae. In healthcare settings, bacteria are exposed frequently to antibiotics at sub-inhibitory concentrations (sub-MICs), which may alter their virulent characteristics. The aim of this research was to investigate the impact of fosfomycin at sub-MICs on the virulence determinants of MDR K. pneumoniae. The effect of sub-MIC fosfomycin treatment on biofilm formation was evaluated by crystal violet assay and confocal laser scanning microscopy (CLSM). The cartwheel technique was employed to study efflux pump activity phenotypically. The expression of genes of biofilm (fimH, mrkD) and efflux pump (acrA, acrB) was determined using reverse transcription quantitative PCR (RT-qPCR). Scanning electron…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
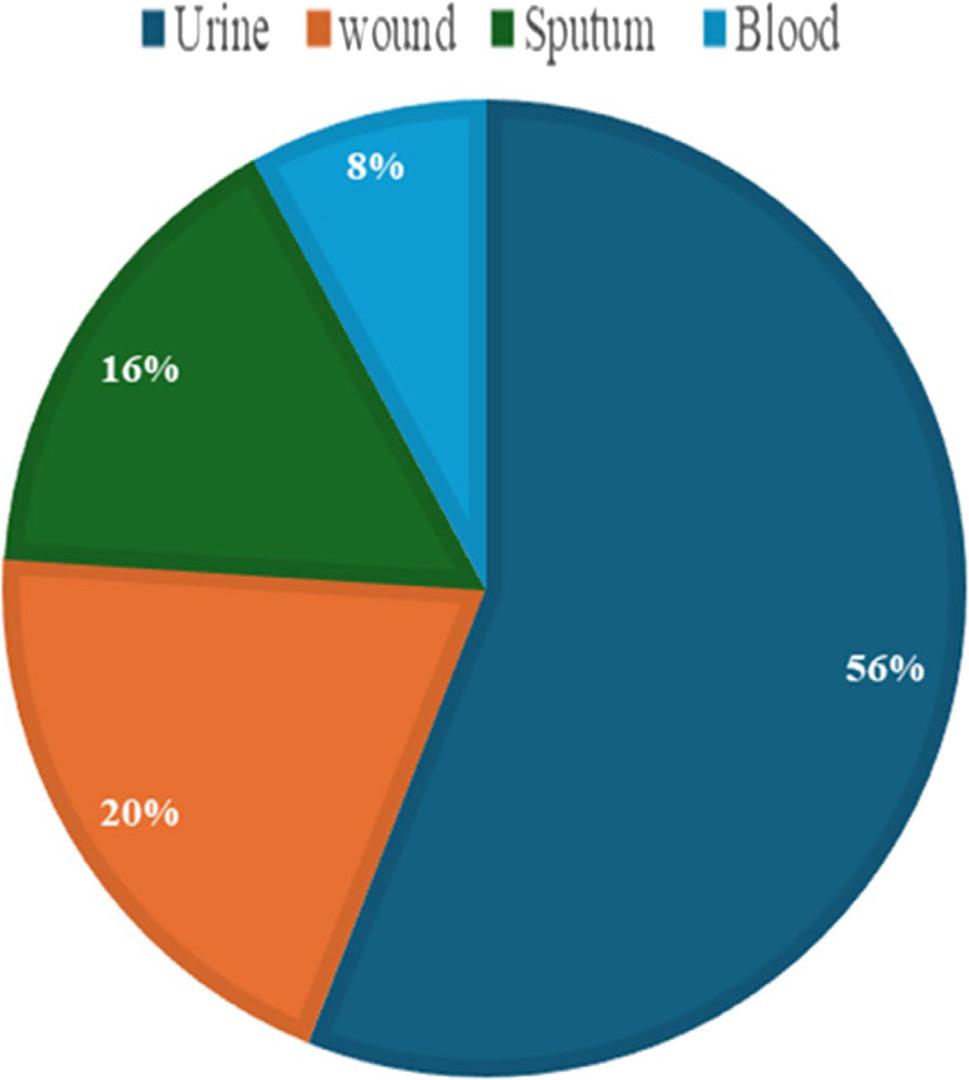 Figure 1
Figure 1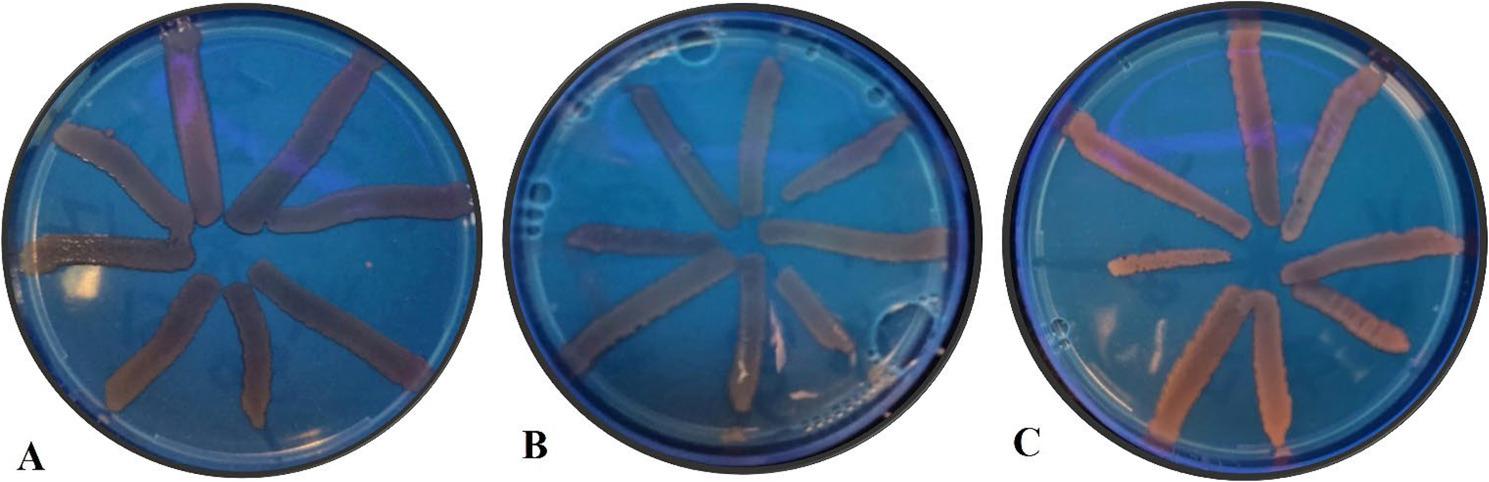 Figure 2
Figure 2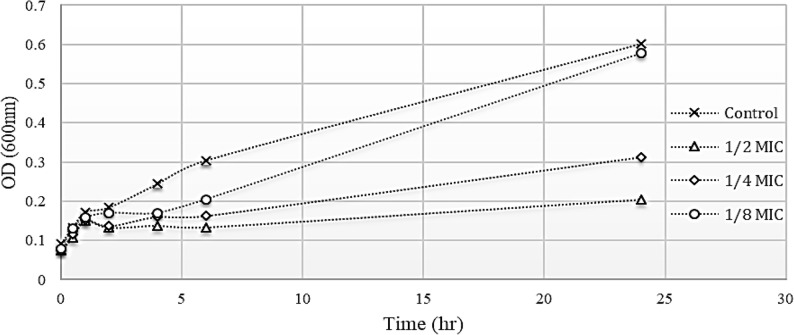 Figure 3
Figure 3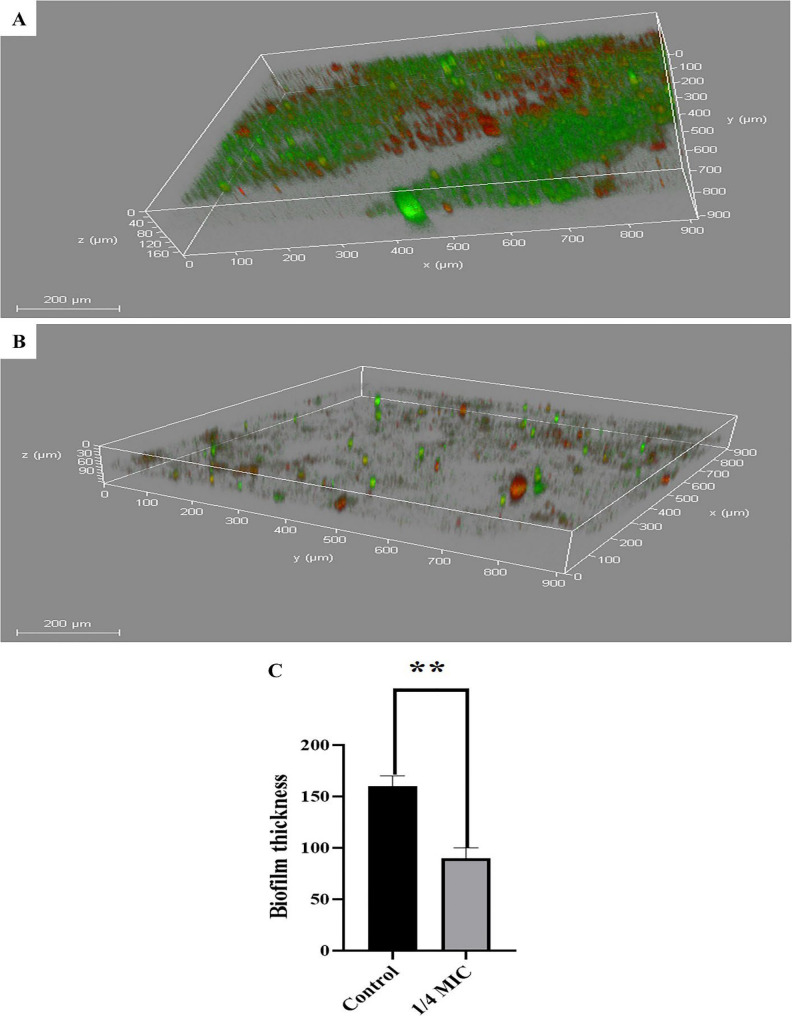 Figure 4
Figure 4 Figure 5
Figure 5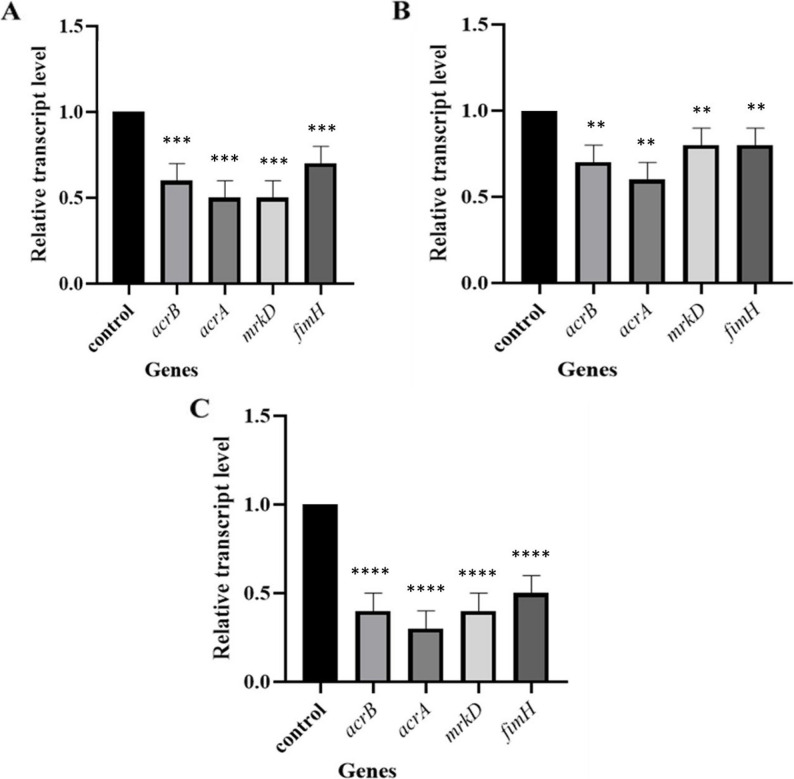 Figure 6
Figure 6- —Tanta University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Urinary Tract Infections Management · Bacterial biofilms and quorum sensing
Background
Klebsiella pneumoniae, a member of the Enterobacteriaceae family, is a Gram-negative and rod-shaped organism [1]. It ranks among the leading infectious agents associated with hospital-acquired infections, causing several conditions, including bacteremia, liver abscesses, pneumonia, and infections of the urinary tract [2].
Since K. pneumoniae accounts for approximately one-third of infections attributed to Gram-negative pathogens, it has emerged as a global threat [3]. In addition, K. pneumoniae has shown increasing resistance to numerous antibiotic classes. Over one-third of isolates reported in Europe are resistant to at least one major antibiotic class, with many exhibiting cross-resistance to aminoglycosides, cephalosporins, and fluoroquinolones, which makes them multidrug resistant [4].
Moreover, K. pneumoniae is also a reservoir for resistance genes, which can be yet another challenge because genes can be horizontally transferred to other Gram-negative species, which accelerates the spread of multidrug resistance [3, 5]. K. pneumoniae possesses multiple virulence factors, such as capsule polysaccharides and type 1 and 3 fimbriae, which are important for its ability to form biofilms [6]. Two major contributors of multidrug resistance are active efflux pump mechanisms and the ability to form biofilms [7].
Efflux pumps represent protein complexes that actively extrude toxic agents from the bacterial cells. For example, the AcrAB system in K. pneumoniae significantly contributes to the development of MDR strains, while the biofilm enhances the bacteria’s resistance to both immune responses and antibiotic treatment in MDR isolates [8]. Because the MDR pathogens spread tremendously while the development of new antibiotics effective against them progressed very slowly, physicians began using older antibiotics such as fosfomycin, which retains activity against a broad range of bacterial species.
Fosfomycin acts as a bactericidal agent with a broad spectrum, effective against a range of Gram-positive organisms, such as Staphylococcus aureus and Enterococcus species, as well as Gram-negative bacteria such as Pseudomonas aeruginosa and K. pneumoniae [9]. Fosfomycin works by blocking the initial stage of the bacterial cell wall synthesis, inhibiting the production of the peptidoglycan precursor, uridine diphosphate N-acetyl muramic acid. Fosfomycin can also penetrate biofilms. Multiple studies using both in vitro and in vivo models have demonstrated that biofilms can be reduced or eliminated when fosfomycin is used alone or alongside other antibiotics. In P. aeruginosa and E. coli models, fosfomycin disrupted biofilms, especially when combined with agents like prulifloxacin or N-acetylcysteine [10, 11].
Beyond antibiotics’ activity at therapeutic concentrations, they can exert profound effects at sub-MICs, which refer to levels of concentration below the MIC threshold. Bacteria are frequently subjected to sub-inhibitory concentrations of antibiotics, commonly found in the environment due to agricultural practices and wastewater discharge, as well as in healthcare settings because of prophylactic low-dose use, improper dosing, or poor patient compliance [12]. Although sub-MICs do not inhibit the growth of bacteria, they can still alter significant bacterial functions and physico-chemical properties such as adhesiveness, surface hydrophobicity, and motility [13].
Therefore, extending our knowledge regarding the effects of sub-inhibitory concentrations of antibiotics on the behavior of bacteria is clinically important. Consequently, this study was designed to investigate how the sub-MIC of fosfomycin influences K. pneumoniae, focusing on virulence factors, efflux pumps, and biofilm formation.
Materials and methods
Isolation and identification of Klebsiella pneumoniae
The clinical isolates were obtained from tertiary settings in Damanhour and Alexandria, Egypt, from August 2022 to September 2023. They were identified by culturing them on MacConkey agar (Oxoid, UK) plates containing fosfomycin breakpoints, targeting the collection of fosfomycin-resistant K. pneumoniae and subjecting them to routine biochemical assays, including triple sugar iron agar, indole, methyl red, Voges-Proskauer, and citrate utilization, and their identities were confirmed using VITEK^®^ 2 (BioMérieux, France) [14].
Antibiotic susceptibility testing
The Kirby-Bauer disc diffusion method determined the susceptibility pattern of Klebsiella pneumoniae isolates to various antibiotics, and Escherichia coli (ATCC 25922) served as the reference strain. The tested antibiotic discs (Oxoid, UK) were ciprofloxacin (CIP), amoxicillin (AMX), amoxicillin-clavulanate (AMC), cefazolin (CZ), tobramycin (TOB), gentamicin (GEN), fosfomycin (FF, glucose-6-phosphate), doxycycline (DO), imipenem (IMP), cefotaxime (CTX), amikacin (AK), and azithromycin (AZM), and the concentrations per disc were [5, 10], (20/10) [10, 15],, (200 + 50) [10, 15],, and [16] µg, respectively. The results were classified according to the CLSI, 2021 [16].
The multiple antibiotic resistance index (MAR) is employed to measure the levels of antibiotic resistance through this equation:
\documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$\begin{aligned} &\mathbf{M}\mathbf{A}\mathbf{R}\:\mathbf{i}\mathbf{n}\mathbf{d}\mathbf{e}\mathbf{x}\\&=\:\frac{\mathrm{N}\mathrm{u}\mathrm{m}\mathrm{b}\mathrm{e}\mathrm{r}\:\mathrm{o}\mathrm{f}\:\mathrm{a}\mathrm{n}\mathrm{t}\mathrm{i}\mathrm{b}\mathrm{i}\mathrm{o}\mathrm{t}\mathrm{i}\mathrm{c}\mathrm{s}\:\mathrm{t}\mathrm{o}\:\mathrm{w}\mathrm{h}\mathrm{i}\mathrm{c}\mathrm{h}\:\mathrm{t}\mathrm{h}\mathrm{e}\:\mathrm{i}\mathrm{s}\mathrm{o}\mathrm{l}\mathrm{a}\mathrm{t}\mathrm{e}\:\mathrm{i}\mathrm{s}\:\mathrm{r}\mathrm{e}\mathrm{s}\mathrm{i}\mathrm{s}\mathrm{t}\mathrm{a}\mathrm{n}\mathrm{t}\:}{\mathrm{T}\mathrm{o}\mathrm{t}\mathrm{a}\mathrm{l}\:\mathrm{n}\mathrm{u}\mathrm{m}\mathrm{b}\mathrm{e}\mathrm{r}\:\mathrm{o}\mathrm{f}\:\mathrm{t}\mathrm{e}\mathrm{s}\mathrm{t}\mathrm{e}\mathrm{d}\:\mathrm{a}\mathrm{n}\mathrm{t}\mathrm{i}\mathrm{b}\mathrm{i}\mathrm{o}\mathrm{t}\mathrm{i}\mathrm{c}\mathrm{s}\:} \end{aligned}$$\end{document}If the value > 0.2, it indicates that the isolate comes from a high-risk source with frequent antibiotic exposure [17, 18].
Determination of fosfomycin Minimum InhibitoryConcentration (MIC)
MICs of the clinical isolates were determined by agar dilution, which is the sole standardized method for testing the MIC of fosfomycin (Sigma Aldrich). Muller-Hinton agar (Oxoid, UK) plates were prepared with the addition of 25 µg/mL of glucose-6-phosphate (G6P) (Santa Cruz) as recommended by CLSI 2021 guidelines, which were also used to interpret the results: susceptible (≤ 64 µg/mL), intermediate (128 µg/mL), and resistant (≥ 256 µg/mL). Fosfomycin concentrations ranged from 0.5 to 1024 µg/mL. All subsequent experiments involving fosfomycin included G6P at this concentration.
Screening of biofilm production
The ability of Biofilm formation was assessed for all Klebsiella pneumoniae isolates. Overnight cultures (16–20 h) of the test isolates in Lauria Bertani broth (Oxoid, UK) were diluted in LB medium (1:100) to assess biofilm formation. Aliquots (100 µL) were inoculated into 96-well microtiter plates in triplicate. Wells containing only LB medium acted as negative controls. Following static incubation (37 °C, 24 h) carefully removing the culture, and phosphate-buffered saline (PBS) was used to rinse each well gently to eliminate non-adherent cells. Then the biofilm underwent fixation with methanol for 30 min and was stained with 2% crystal violet for 15 min. Excess dye was rinsed off, and plates were air-dried for 10–15 min. To quantify biofilm formation, the bound stain was solubilized in glacial acetic acid (33%), and absorbance was measured at 590 nm using an ELISA reader (Sunrise TM, TECAN, Switzerland) [19].
The 3 standard deviations (SD) above the average of OD of the negative control is known as the cut-off value (ODc). ODc = Average of optical density OD_avg_ + 3 × SD of OD_avg_. Isolates were divided into four categories: non-biofilm producer (OD ≤ ODc), weak (ODc < OD ≤ 2ODc), moderate (2ODc < OD ≤ 4ODc), and strong biofilm producer (OD > 4ODc). according to Stepanovic et al., [20].
Phenotypic detection of efflux pump
The cartwheel technique was employed to phenotypically assess efflux pump activity in K. pneumoniae isolates. Mueller–Hinton agar plates were supplemented with varying ethidium bromide (EtBr) concentrations (ranging from 0.5 to 2 µg/mL). Bacterial suspensions were adjusted to 0.5 McFarland with a turbidity assay. The isolates were streaked from the center to the margin on EtBr-containing plates and negative control (without EtBr) plates. They were incubated for an entire night at 37 °C. A UV transilluminator (SpectroLine Model CM-10 Fluorescence Analysis Cabinet) was used to examine the plates. Isolates were assessed for efflux pump activity based on their fluorescence patterns [21].
Growth curve assay
A growth curve assay was performed to assess the impact of sub-MIC fosfomycin on bacterial growth. The isolates were cultured in LB broth with and without ½, ¼, and ⅛ MICs of fosfomycin at 37 °C. Bacterial growth was evaluated by measuring absorbance at specific time intervals (0, 30, 60, 120, 240, 360 min, and 24 h) [22, 23].
The effect of the sub-MIC of fosfomycin on the Inhibition of biofilm formation
The impact of sub-MIC of fosfomycin on biofilm formation by K. pneumoniae isolates was assessed using the crystal violet assay as described previously [19, 20]. Bacterial cultures were inoculated in LB medium, into microtiter plates with and without ¼ MIC of fosfomycin, to assess its inhibition of biofilm growth. The wells having no fosfomycin served as negative controls.
Assessment of biofilm structure and viability by confocal laser scanning microscopy (CLSM)
The effect of the sub-MIC of fosfomycin on K. pneumoniae biofilm and cells was evaluated using CLSM. Selected bacterial suspensions (10⁶ CFU/mL), both untreated and treated with 1/4 MIC fosfomycin, were cultured in LB broth on sterile glass coverslips placed in 6-well culture plates. After incubation for 18 h (37 °C) under static conditions, non-adherent cells were washed with PBS.
After that, the biofilm was stained with each of acridine orange (AO) and propidium iodide (PI) (Sigma, USA) for 15 min in the dark. PI stains dead cells red, while living cells are stained green by AO. Following staining, the biofilm structure and viability were analyzed using CLSM (DMi8, Leica Microsystems, USA) [24].
Scanning electron microscopy
A representative isolate was cultivated in LB broth with and without exposure to the sub-MIC of fosfomycin for an entire night at 37 °C. The resulted pellets were processed for additional analysis, after washing several times with PBS. The pellets were resuspended, fixed at room temperature for two hours using 2.5% glutaraldehyde in PBS buffer (pH 7.4), and then postfixed for one hour (4 °C) using 1% OsO4 in PBS buffer (pH 7.4). After dehydrating ethanol and allowing it to air dry, a drop of prepared bacterial suspension was placed onto a microscope slide mounted on metal stubs and coated with gold using a sputter coater (Akashi Seisakusho, Japan). The samples were then visualized under a scanning electron microscope (SEM) [25].
Gene expression in various bacteria
The total RNA was extracted from both untreated and treated K. pneumoniae isolates with ¼ MIC of fosfomycin using TRIzols Reagent (15596026, Life Technologies, USA) following the manufacturer’s instructions. The extracted RNA was then reverse transcribed into single-stranded complementary DNA (cDNA) using the QuantiTect^®^ Reverse Transcription Kit (Qiagen, USA).
Quantitative real-time PCR (RT-qPCR) was conducted on a Rotor-Gene Q system (Qiagen, USA) using Maxima SYBR Green/Fluorescein qPCR Master Mix (Thermo Scientific, USA). Each 25 µL reaction contained 300 nM of each primer and 30 ng of cDNA. All samples were run in duplicate. Expression levels of target genes were normalized to rpoD, which is the housekeeping gene, using the ∆Ct method, and relative gene expression was calculated by the 2^–∆∆Ct method [26], with untreated bacteria used as the calibrator (Table 1).
Table 1. Quantitative real-time PCR primer sequencesGeneSequenceAnnealing Temp. (ºC)Reference acrB F-CAATACGGAAGAGTTTGGCAR-CAGACGAACCTGGGAACC58[27] acrA F-GTCCTCAGGTCAGTGGCATTAR-ATTGCTCTGCTGCGCCGTTG60[28] mrkD F-CCACCAACTATTCCCTCGAAR- ATGGAACCCACATCGACATT58[29] fimH F-TCCACAGTCGCCAACGCTTCR-GCTCAGAATCAACATCGGTAAC58[30] rpoD (Housekeeping gene)F-TCCGGTGCATATGATTGAGAR-ATACGCTCAGCCAGCTCTTC60[15]
Statistical analysis
In the present study, we employed ANOVA (one-way) and t-test for statistical comparisons and the p-value < 0.05 was considered significant. All experiments were performed in triplicate, and the results were presented as mean ± SD. Data were analyzed and visualized using GraphPad Prism version 8.
Results
Identification of Klebsiella pneumoniae
The isolates were Gram-negative, rod-shaped, and produced lactose-fermenting, pink mucoid colonies on MacConkey agar. They were positive for Voges-Proskauer and citrate utilization while being negative for indole and methyl red tests. They also gave acid over acid and gas in the triple sugar iron test. The identity of all isolates was confirmed at the species level using the VITEK^®^ 2 system. The number of fosfomycin-resistant K. pneumoniae (n = 50) that were collected from urine, wound, sputum, and blood was 28,10,8,4, respectively (Fig. 1).
Fig. 1. Distribution of K. pneumoniae isolates recovered from different clinical specimens. Percentages were calculated based on the total number of isolates (n = 50)
Antimicrobial susceptibility pattern
A total of 50 K. pneumoniae isolates exhibited varying degrees of resistance to the tested antibiotics. Their resistance percentages were as follows: imipenem (58%), ciprofloxacin (60%), amoxicillin (100%), amoxicillin–clavulanic acid (100%), cefazolin (94%), tobramycin (32%), gentamicin (46%), azithromycin (70%), doxycycline (38%), cefotaxime (70%), amikacin (74%), and fosfomycin (100%) (Table 2).
Table 2. It displays the antimicrobial resistance patterns of the tested Klebsiella pneu**moniae isolates against the selected antibiotics. The multiple antibiotic resistance (MAR) index values varied between 0.3 and 1. [17, 18]Pattern codeResistance markerIsolate codeNumberMAR index I AMX-AMC-CZ-FFK4, K38, KUK30.33 II AMX- AMC- CZ- FF-AKK23, K24, and KE130.416 III AAMX-AMC-CZ-AZM-DO-FFK26, K4020.5BAMX-AMC-CZ-AZM-CTX-FFKBK1CAMX-AMC-CZ-FF-DO-AKK81 IV ACIP-AMX-AMC-CZ-FF-GEN-CTXK54, K6, K2830.58BAMX-AMC-CZ-GEN-AZM-FF-CTXK53, K272CAMX-AMC-CZ-AZM-FF-DO-AKKE2, K14A, K16A3 V AAMX-AMC-CZ-FF-TOB-GEN-AZM-CTXK52, K2920.667BAMX-AMC-CZ-AZM-FF-DO-CTX-AKK91CAMX-AMC-CIP-AZM-FF-IMP-CTX-AKK7A, K1A, K13A3DCIP-AMX-AMC-CZ-IMP-CTX-AK-FFK10, K212 VI ACIP-AMX-AMC-CZ-AZM-FF-IMP-CTX-AKK13, K9A20.75BCIP-AMX-AMC-CZ-TOB-FF-IMP-CTX- AKK7, K3 &K483 VII ACIP-AMX-AMC-CZ-AZM-DO-IMP-CTX-FF-AKK17, K14&K4930.833BCIP-AMX- AMC- CZ-GEN- AZM- FF- DO- IMP- AKK22, K12A, K53CAMX-AMC-CZ- GEN- AZM- FF- DO- IMP- CTX- AKK20, K122 VIII CIP- AMX- AMC- CZ-TOB-GEN-AZM-FF-IMP-CTX-AKK50, K51, K6A, K5A, K3A, K8A, K11A.70.916 X CIP-AMX-AMC-CZ-TOB-GEN-AZM-FF-DO-IMP-CTX-AKK25, K11, K18, K19.41AMX amoxicillin, AK amikacin, AMC amoxicillin-clavulanate, AZM azithromycin, CIP ciprofloxacin, CTX cefotaxime, CZ cefazolin, DO doxycycline, FF fosfomycin, GEN gentamicin, IMP imipenem, and TOB tobramycin
Determination of fosfomycin Minimum Inhibitory Concentrations (MIC)
The MIC values against fosfomycin ranged from 256 to 1024 µg/mL. The majority exhibited (38 isolates) an MIC of 256 µg/mL, while 9 and 3 isolates were found to have MICs of 512 and 1024 µg/mL, respectively (Table 3).
Table 3MIC values for K. pneumoniae isolates against fosfomycinMIC value (µg/ml)Number of isolates (%)Isolate code 25638 (76%)k17, k11, K20, K53, K6A, K12A, K21, K9A, K13A, K9, K52, K48, K28, K8, K24, K40, K23, K4, K38, K8A, K14A, KE1, K7A, K1A, K13, K7, K26, K54, K10, K22, K5A, K3A, K50, KUK, KBK, KE2, K19, K51. 5129 (18%)K14, K5, K16A, K29, K18, K12, K11A, K27, K6. 10243 (6%)K25, K49, K3
Quantification of biofilm production
The results revealed varying capacities for biofilm formation among K. pneumoniae isolates. Eleven isolates (22%) were classified as strong producers, 23 (46%) exhibited moderate ability of biofilm production, and 16 (32%) were identified as weak biofilm producers.
Phenotypic detection of efflux pump
The results showed that all K. pneumoniae isolates had efflux pumps using the cartwheel method. No fluorescence was observed at 1.5 µg/mL of ethidium bromide (EtBr), which indicated that the bacterial isolates efficiently efflux the EtBr dye. However, at higher concentrations of EtBr (2 µg/mL), fluorescence was observed, pointing out the accumulation of intracellular EtBr due to the inhibition of the efflux pump (Fig. 2).
Fig. 2. The cartwheel method was used to determine efflux pump activity. A Plate without EtBr; negative control, showed no fluorescence. B Plate containing EtBr (1.5 µg/mL) showed no fluorescence. C Plate containing EtBr (2 µg/mL) showed fluorescence
Growth curve assay
The impact of ½, ¼, and ⅛ of the MIC of fosfomycin on 19 selected K. pneumoniae isolates (each representing a different reported resistance pattern) was evaluated (Fig. 3). The results showed that ½ MIC of fosfomycin affected bacterial growth markedly, while the effect of the ⅛ MIC was negligible. In contrast, ¼ MIC exhibited moderate activity. Consequently, ¼ MIC was selected for further investigation.
Fig. 3. Growth curve of one of the isolates that was cultured in the absence and the presence of ½, ¼, and ⅛ of MICs of fosfomycin
The effect of the sub-MIC of fosfomycin on biofilm formation
It was observed that ¼ MIC of fosfomycin resulted in statistically significant inhibition of biofilm formation (p < 0.05) compared with their negative (untreated) controls in all tested isolates (n = 19) (See supplementary Table S1). The degree of inhibition varied among the isolates, with the percentage of reduction ranging from 37.8% to 76.1% (Table 4).
Table 4. Effect of the treatment by ¼ MIC of fosfomycin on the Inhibition of biofilm formationCategories ofBiofilm ProductionPre-Treatment(without ¼ MIC of fosfomycin)Post-treatment(with ¼ MIC of fosfomycin)Strong6 (31.57%)1 (5.2%)Moderate10 (52.63%)8 (42.105%)Weak3 (15.7%)10 (52.6%)
Confocal laser scanning microscopy
CLSM was employed to investigate the impact of ¼ MIC of fosfomycin on K. pneumoniae. The results showed a significant reduction in overall thickness and density with increased red fluorescence in some areas, which refers to dead cells. However, the untreated biofilm producer isolate (K17) demonstrated a dense and well-structured architecture with pronounced green fluorescence, signifying a substantial proportion of viable cells within the biofilm matrix (Fig. 4).
Fig. 4CLSM of K. pneumoniae (K17) (A) Untreated biofilm. B Treated biofilm with ¼ MIC of fosfomycin, (C) Biofilm thickness (µm) with and without fosfomycin treatment. The error bars indicate standard deviations. Statistical significance was evaluated using a paired t-test, and (**) represents a statistical difference (p < 0.01)
Scanning electron microscope
The scanning electron microscope images exhibited significant morphological differences between the untreated isolate and after treating it with ¼ MIC of fosfomycin (Fig. 5). Concerning the untreated isolate, the bacterial cells were densely packed and had typical smooth and rod-shaped morphology. On the other hand, the isolate treated with ¼ MIC of fosfomycin was sparsely dispersed and exhibited roughening or irregular rod-shaped and wrinkling of cell surface features with “cauliflower-like distortion”. Overall, fosfomycin at sub-MIC significantly disrupted both bacterial aggregation and biofilm structural integrity.
Fig. 5SEM of K. pneumoniae (K17). A Untreated cells appeared as rod-shaped bacilli. B Treated cells with ¼ MIC of fosfomycin revealed distorted morphology
The impact of the sub-MIC of fosfomycin on the gene expression
The effect of ¼ MIC of fosfomycin on gene expression of representative isolates (K17, K20, K11) was studied using quantitative real-time PCR. The criteria for the selection of these isolates were MDR and having varying capacities for biofilm formation. Isolate K17 had the ability to form a strong biofilm, isolate K20 showed moderate biofilm formation, while isolate K11 formed a weak biofilm.
These findings reveal that the sub-MIC of fosfomycin significantly downregulated the gene expression of efflux pump activity and biofilm formation, potentially contributing to decreased virulence and antibiotic resistance.
The degree of the downregulation among isolates was variable. Notably, the moderate biofilm-forming isolate (K20) showed the greatest overall suppression, particularly in acrA (70%) and acrB (60%). All changes were statistically significant (p < 0.05). The expression of key virulence- and resistance-associated genes after sub-MIC fosfomycin exposure is presented in Fig. 6.
Fig. 6. Relative transcription levels of the following selected genes, acrA, acrB, mrkD, and fimH, in three K. pneumoniae. A K17 isolate, (B) K11 isolate, and (C) K20 isolate, after treatment with ¼ MIC of fosfomycin. The gene expression levels were normalized relative to rpoB (Housekeeping gene) and presented as fold change relative to the control, which is set to 1. The error bars indicate standard deviations. The statistical significance was determined by one-way ANOVA (each in comparison to the control group). The (), (), and (***) represent statistical significance (p < 0.01), (p < 0.001), and (p < 0.0001), respectively
Discussion
Klebsiella pneumoniae is increasingly recognized for its characteristic of resistance to multiple classes of antibiotics, including several last-line treatments. This pathogen represents a significant challenge in the hospital setting, since infections are often associated with serious complications and high mortality rates [3, 31]. Due to these high infection rates, an increase in MDR K. pneumoniae has become an urgent priority for global health [8].
The most common mechanisms of resistance reported for MDR K. pneumoniae against various antibiotic classes are biofilm formation and efflux pumps. Key virulence factors, such as fimbriae (types 1 and 3) and capsule, confer increased pathogenicity to K. pneumoniae and play an important role in biofilm development [6]. Biofilm is an assembly of microorganisms adhering to biotic surfaces, such as mucosal surfaces of the respiratory, urinary, and gastrointestinal systems, or non-living surfaces, including catheters and endotracheal tubes. Fimbriae (types 1 and 3) contribute to different aspects of bacterial colonization and persistence, with type 1 responsible for the development of intracellular bacterial communities and type 3 essential for biofilm development on both living and non-living surfaces [32–34].
Considering our observed results related to the marked biofilm reduction under ¼ MIC of fosfomycin. This observation was previously observed by Pallam et al., who reported that a two-fold sub-MIC of fosfomycin disrupted the biofilm of K. pneumoniae [35]. Another study also reported that ¼ MIC of fosfomycin inhibited biofilm formation in 83.33% of E. coli [36]. A noteworthy observation is that the isolates with MAR index > 0.5 and strong biofilm producers exhibited the most marked reductions in biofilm formation.
Moreover, quantitative PCR analysis revealed a significant downregulation in the expression of fimH and mrkD. Furthermore, according to a study by Guofeng Dong et al., the ¼ MIC of ciprofloxacin significantly suppressed Fim genes [22]. The sub-MIC of amikacin also decreased the expression of *mrkD *in K. pneumoniae, as reported by Rahmati et al., [37].
Furthermore, efflux pumps are another important mechanism that is responsible for resistance in K. pneumoniae. Efflux pumps help maintain bacterial homeostasis by expelling harmful substances such as antibiotics and dyes, which leads to a decrease in the intracellular drug concentration and contributes to antimicrobial resistance [38]. AcrAB is a member of RND family efflux system strongly implicated in multidrug resistance in K. pneumoniae [8, 38]. Expression of acrA and acrB genes were found to be downregulated upon treatment with ¼ MIC fosfomycin, which suggests that subinhibitory fosfomycin concentrations may decrease the activity of efflux pumps and, thus, reduce antimicrobial resistance. Such a sub-MIC effect of antibiotics has been reported recently. Colistin at sub-MIC was shown to behave as an efflux pump inhibitor by inhibiting the AcrAB pump in K. pneumoniae [39]. According to Gil-Gil et al.., who reported that sub-MIC of fosfomycin decreased the expression of the SmeYZ efflux pump in Stenotrophomonas maltophilia [40]. Another study reported that the sub-mic of fosfomycin downregulated virulence-related genes, transport, nucleic acid biosynthesis, and energy metabolism pathways in Staphylococcus aureus [41]. Based on these studies, we propose that when fosfomycin was used at the sub-MIC in K. pneumoniae, it not only interfered with cell wall synthesis but also triggered a widespread starvation response. As fosfomycin is an analogue of phosphoenol pyruvate (PEP), and inhibits MurA, resulting in accumulation of PEP and activating the cell wall stress response, which may result in downregulation of energy-consuming processes such as efflux pumps. To the best of our knowledge, this is the first study to investigate the effect of sub-MIC fosfomycin on efflux pump gene expression in K. pneumoniae.
The “cauliflower-like” surface alterations that were observed in the treated isolate by SEM, previously reported by [42], where the sub-MIC of the fosfomycin caused a similar change in morphology in the Staphylococcus aureus. Overall, these findings suggest that even sub-inhibitory concentrations of fosfomycin can disturb biofilm integrity and attenuate efflux pump systems, highlighting its possible role as an adjunct approach against MDR K. pneumoniae infections.
Additional investigations are necessary to explain the molecular mechanisms underlying the decrease in the activities of efflux pumps and biofilm formation after treatment with fosfomycin at sub-MIC levels. Our results also show that there is a strain-dependent response, since isolates reacted differently, despite the sample size being too small to draw wide general conclusions. In-depth analysis with more isolates would give more information on gene expression under fosfomycin pressure. Therefore, further studies should involve a higher diversity of isolates to confirm our findings and to establish whether the observed effects are constant among tested strains. Moreover, OD measurements were used to assess bacterial growth. However, this approach can’t differentiate between live and dead cells, nor overcome possible cell clumping effects that may reduce its accuracy. As a result, we suggest further studies to include time kill assays or colony-forming unit count. Finally, in vivo and clinical studies are needed to confirm the clinical relevance of these findings and to determine the therapeutic potential of sub-MIC fosfomycin to combat the serious infections caused by K. pneumoniae.
Conclusion
This study demonstrates that sub-inhibitory concentrations (¼ MIC) of fosfomycin significantly interfere with biofilm formation, downregulate essential virulence genes (fimH,* mrkD*), and reduce efflux pump gene expression (acrA,* acrB*) in multidrug-resistant Klebsiella pneumoniae isolates. Morphological analysis indicated a marked alteration of morphology. Although a limited number of isolates underwent gene expression analysis, these results may indicate that fosfomycin, even at a concentration lower than the MIC, could disrupt biofilm integrity and might act as a promising adjuvant therapy against MDR infections. Further clinical studies are necessary with more representative isolate samples to validate these findings.
Supplementary Information
Supplementary Material 1
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1European Centre for Disease Prevention and Control (ECDC). Surveillance of antimicrobial resistance in Europe 2018. Stockholm: ECDC; 2018. https://www.ecdc.europa.eu/en/publications-data/surveillance-antimicrobial-resistance-europe-2018
- 2Riwu KHP, Effendi MH, Rantam FA, Khairullah AR, Widodo A. A review: virulence factors of Klebsiella pneumonia as emerging infection on the food chain. Vet World. 2022;2172–2179. 10.14202/vetworld.2022.2172-2179 PMC 963138436341059 · doi ↗ · pubmed ↗
- 3Osman EA, El-Amin N, Adrees EAE, Al-Hassan L, Mukhtar M. Comparing conventional, biochemical and genotypic methods for accurate identification of Klebsiella pneumoniae in Sudan. Access Microbiol. 2020;2(3). [Cited 2025 Apr 21] Available from: https://www.microbiologyresearch.org/content/journal/acmi/10.1099/acmi.0.00009610.1099/acmi.0.000096 PMC 747031232974573 · doi ↗ · pubmed ↗
- 4Krumperman PH. Multiple antibiotic resistance indexing of Escherichia coli to identify High-Risk sources of fecal contamination of foodst. APPL Env MICROBIOL. 1983;46. (1): 165:170 10.1128/aem.46.1.165-170.1983 PMC 2392836351743 · doi ↗ · pubmed ↗
- 5Clegg S, Murphy CN. Epidemiology and Virulence of Klebsiella pneumoniae. Mulvey MA, Stapleton AE, Klumpp DJ, editors. Microbiol Spectr. 2016;4(1):4.1.06 10.1128/microbiolspec.UTI-0005-201226999397 · doi ↗ · pubmed ↗