Total Chemical Synthesis of RNF4 by Sequential Native Chemical Ligation: C‑To‑N Versus N‑To‑C Strategies
Rajesh Pallava, Saed Bisher, Ashraf Brik

TL;DR
Scientists chemically synthesized the RNF4 protein using two methods, finding that one approach simplified the process and could help study its role in cancer.
Contribution
A reliable and simplified method for total chemical synthesis of RNF4 using N-to-C ligation is presented.
Findings
RNF4 was successfully synthesized using sequential native chemical ligation with five peptide fragments.
The N-to-C ligation strategy simplified the synthesis workflow and reduced purification steps.
The synthesis method addresses challenges in creating large proteins for biological research.
Abstract
RNF4, a RING-type E3 ubiquitin ligase, targets polySUMOylated proteins for ubiquitination and subsequent proteasomal degradation. The ability to chemically synthesize RNF4 will enable future studies of its structure and biological function, particularly its role in degrading the oncoprotein PML-RARα in acute promyelocytic leukemia. To achieve this, we performed a total chemical synthesis of RNF4 using sequential native chemical ligation. The presence of nine cysteine residues enables stepwise ligation of five peptide fragments to assemble the full-length protein. Two synthetic strategies were explored: the first employed a convergent C-to-N ligation, while the second used an N-to-C ligation. In the convergent C-to-N approach, cysteine residues were protected with acetamidomethyl groups to prevent side reactions during ligation, although this required multiple deprotection and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 1
1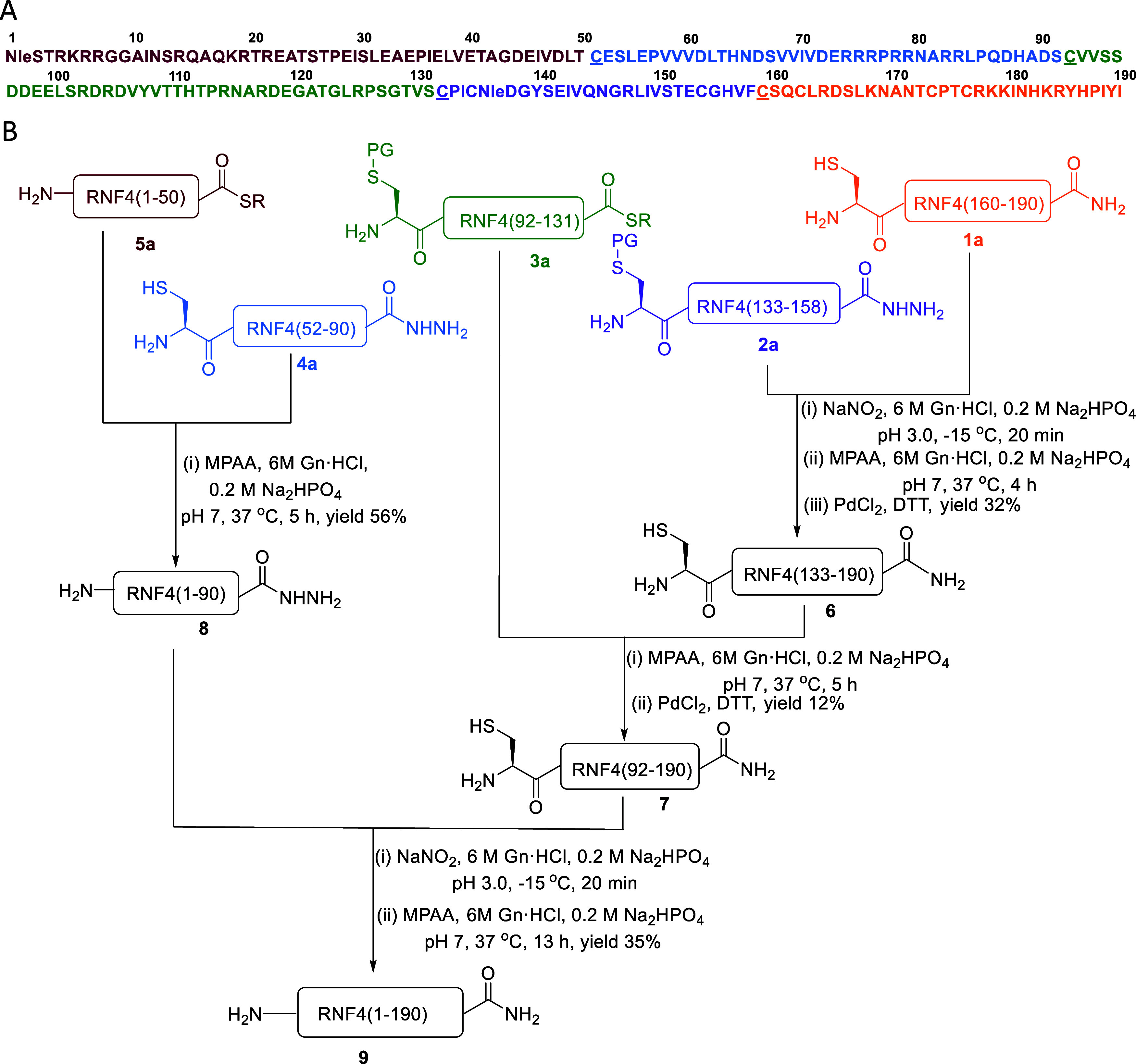 1
1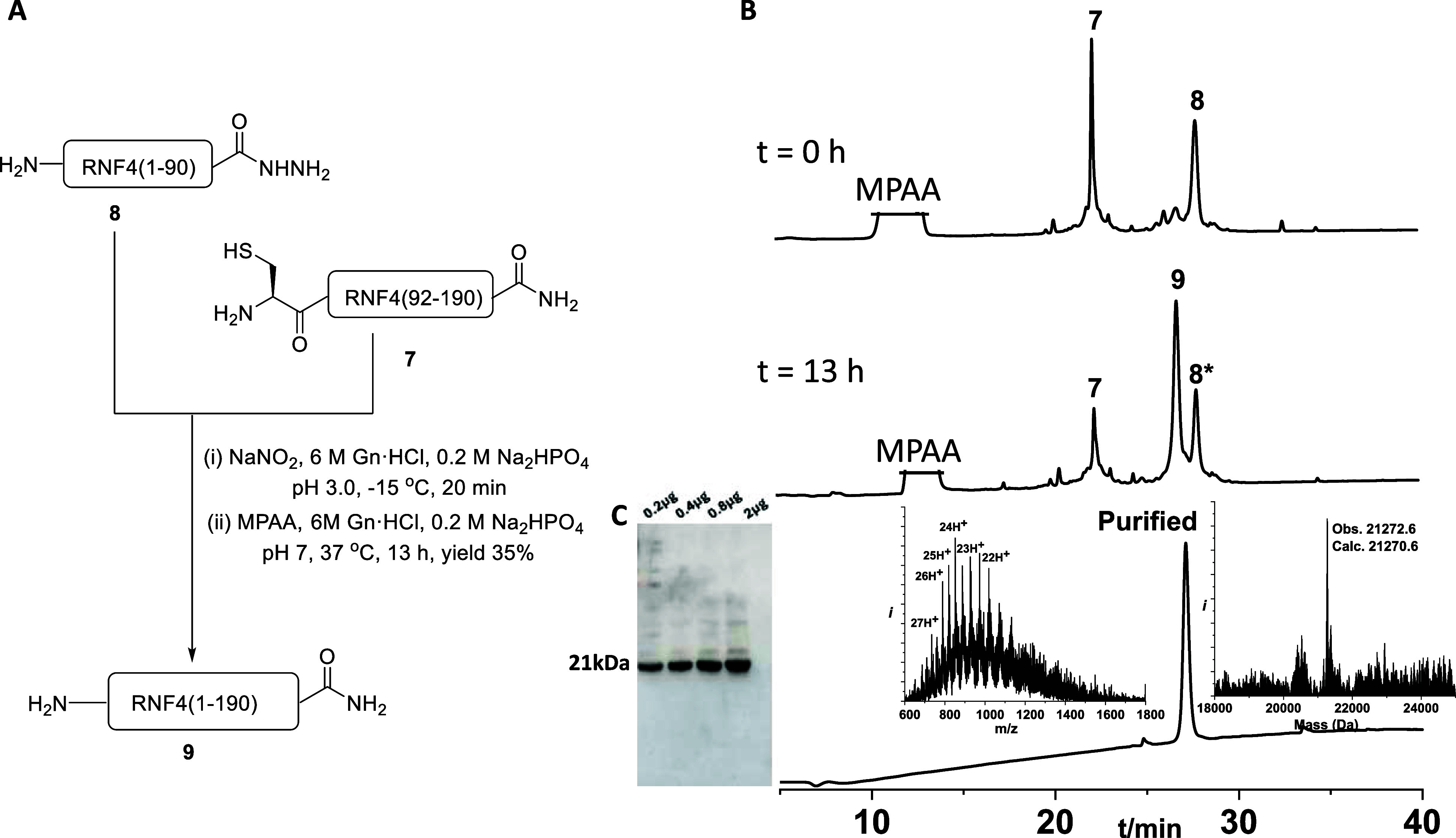 2
2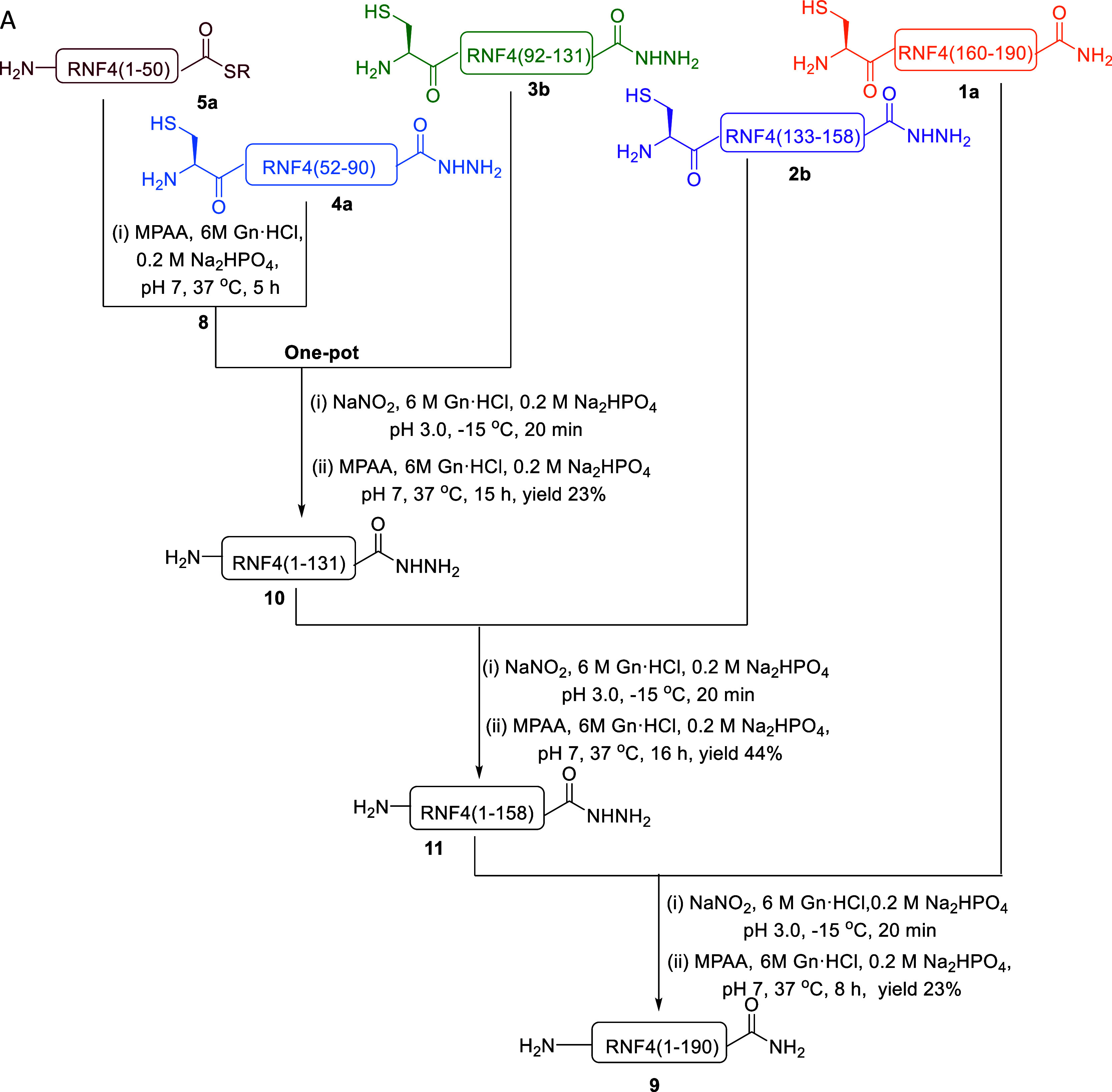 2
2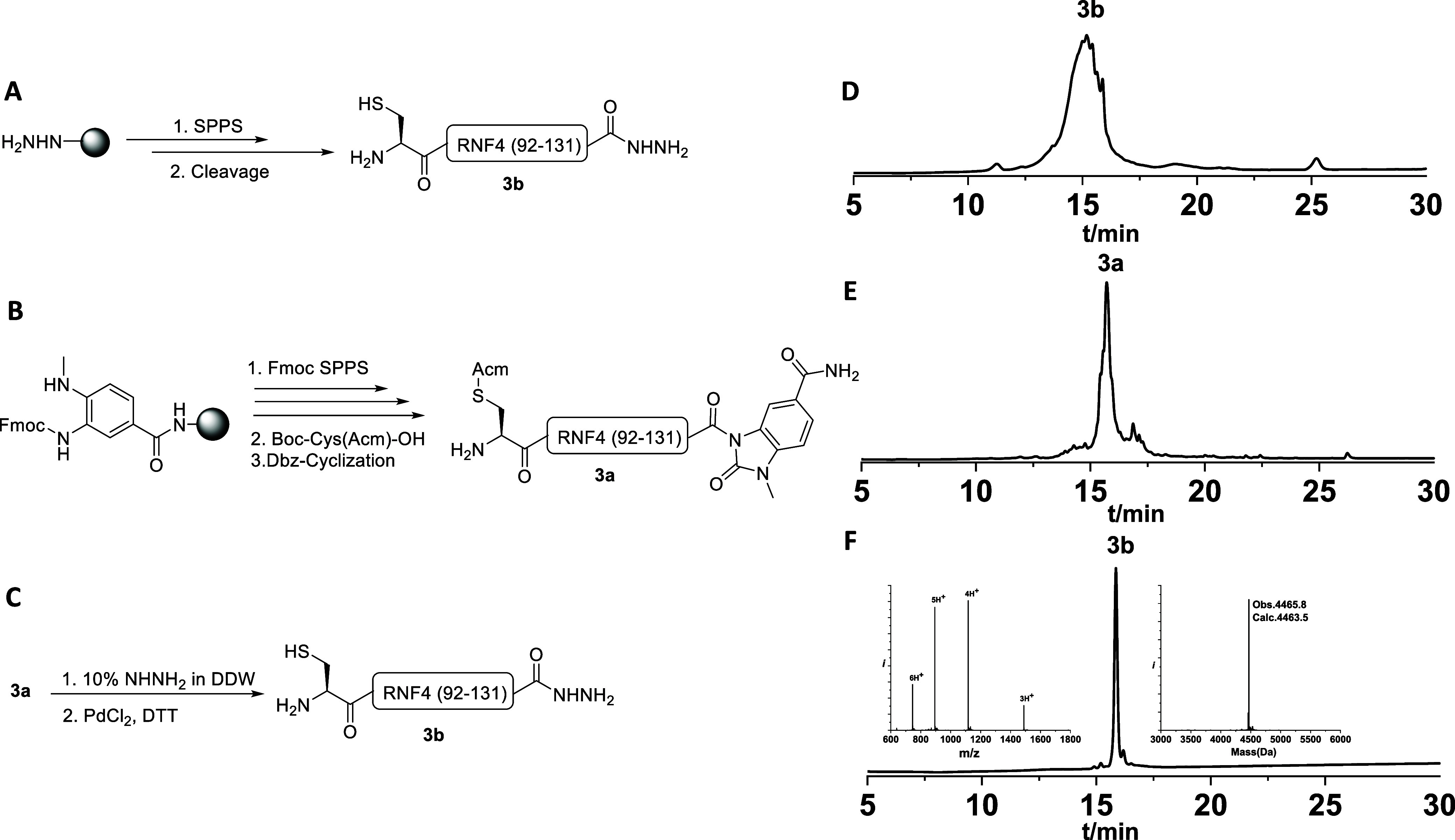 3
3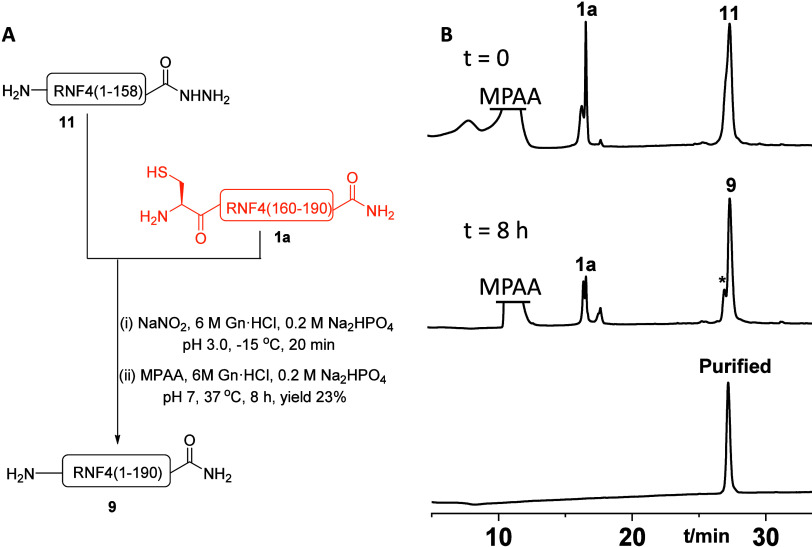 4
4- —Deutsche Forschungsgemeinschaft10.13039/501100001659
- —Ministry of Innovation, Science and Technology10.13039/501100001738
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsUbiquitin and proteasome pathways · Protein Degradation and Inhibitors · Chemical Synthesis and Analysis
Introduction
The Ring Finger Protein 4 (RNF4) is an E3 ubiquitin ligase that targets poly-SUMOylated proteins for ubiquitination and subsequent proteasomal degradation.? RNF4 was first identified in the late 1990^s^ and consists of 190 amino acid residues.? The N-terminal region of RNF4 contains a stretch of hydrophobic residues, valine (V), isoleucine (I), and leucine (L), which represent four well-characterized SUMO-interacting motifs (SIMs), designated as SIM1, SIM2, SIM3, and SIM4. These motifs specifically recognize polymeric SUMO chains, a crucial feature required for ubiquitination and subsequent protein degradation.? In addition, RNF4 contains an arginine-rich motif (ARM; residues 73–82), which is essential for recognizing substrates that are simultaneously SUMOylated and phosphorylated.? The C-terminal region of RNF4 harbors a RING domain that mediates ubiquitin transfer from the E2 enzyme to the substrate. This domain adopts a unique cross-brace structure comprising a conserved arrangement of seven cysteine and one histidine residues that coordinate two Zn^2+^ ions, promoting RING dimerization, which is essential for efficient ubiquitin transfer activity.?
RNF4 is a SUMO-targeted ubiquitin ligase that links SUMOylation to ubiquitin-dependent proteasomal degradation and plays essential roles in genome stability, DNA repair, and cancer-related signaling.? RNF4 recognizes poly-SUMOylated substrates via SUMO-interacting motifs (SIMs) in its intrinsically disordered N-terminal region.? At the same time, its dimeric C-terminal RING domain activates the E2–ubiquitin conjugate to catalyze direct ubiquitin transfer without forming a covalent E3-ubiquitin intermediate.? In acute promyelocytic leukemia (APL), arsenic trioxide (As_2_O_3_) induces poly-SUMOylation of the PML protein, leading to RNF4 recruitment and proteasomal degradation of PML.? In addition to its role in protein degradation, RNF4 has also been implicated in the stabilization and transcriptional activation of several oncogenic proteins essential for cell survival, including c-Myc, c-Jun, β-catenin, and the Notch intracellular domain (N-ICD).?
Previous studies have shown that RNF4 activity is regulated by two post-translational modifications (PTMs): methylation and phosphorylation. Methylation of RNF4 at Arg164 by protein arginine methyltransferase 5 inhibits its interaction with PML-RARα, thereby stabilizing PML-RARα and promoting drug resistance.? Conversely, phosphorylation of RNF4 by cyclin-dependent kinases (CDKs) at Thr26 and Thr112 enhances its activity toward MDC1, which is required for its degradation and proper homologous recombination repair during S phase.? However, it remains unclear whether these PTMs affect RNF4 function within PML nuclear bodies (PML-NBs). The chemical synthesis of RNF4 could provide a powerful opportunity to investigate how these modifications regulate RNF4 activity in PML-NBs. Therefore, there is a clear need for a robust and reliable route to the total chemical synthesis of RNF4. To the best of our knowledge, no reports to date describe the total chemical synthesis or semisynthesis of RNF4. Until now, RNF4 has been obtained only through recombinant expression,? which is itself difficult, particularly due to handling issues and the need to introduce mutations to enable expression and support biological studies. ?,?
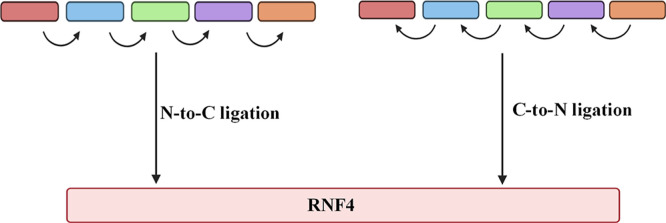
In this study, we report our initial efforts toward the chemical synthesis of the human RNF4 protein, which comprises 190 amino acid residues, including nine cysteines. We examine two synthetic strategies investigated: a C-to-N approach and an N-to-C approach (Figure). Based on the protein sequence, RNF4 was divided into five peptide fragments for each strategy, which were subsequently ligated by native chemical ligation (NCL) to get the full-length RNF4 protein. Initially, we pursued the C-to-N ligation strategy, employing Acetamidomethyl (Acm) protection of cysteine (Cys) residues in two peptide fragments to suppress side reactions during ligation. However, this approach required multiple Acm deprotection and purification steps, resulting in substantial material loss and a reduced overall yield of the RNF4 protein. In contrast, the N-to-C strategy employed cysteine-unprotected fragments and proceeded more efficiently, requiring fewer purification steps and ultimately providing an improved overall yield.
Schematic representation of N-to-C and C-to-N ligation strategies.
Results and Discussion
Synthetic strategies for preparing complex proteins are essential for generating novel proteins for diverse applications.? Solid-phase peptide synthesis (SPPS)? and chemoselective ligation technologies have revolutionized protein synthesis by enabling efficient assembly of proteins for structural and functional studies.? Native chemical ligation (NCL) is widely used to join unprotected peptide segments via a C-terminal thioester and an N-terminal cysteine.? To address the growing demand for challenging multistep protein syntheses, several advanced strategies have been developed to improve homogeneity and scalability.? Furthermore, the ligation methods, such as solid-supported ligation, convergent ligation, and one-pot approaches, and the fragment sequence strongly affect synthesis time, yield, and product purity. ?−? ? ? ? ? These methods enable the efficient synthesis of proteins with increasing size and complexity. To chemically prepare RNF4, we initially explored the C-to-N approach based on sequence considerations and the synthetic accessibility of the peptide fragments. This strategy allowed us to systematically evaluate the feasibility of sequential ligation starting from the C-terminal segments before adopting a convergent assembly strategy. The full-length sequence was divided into five peptide fragments: Fragment 1a Cys-RNF4(160–190); Fragment 2a Cys(Acm)-RNF4(133–158)-NHNH_2_; Fragment 3a Cys(Acm)-RNF4(92–131)-MMP; Fragment 4a Cys-RNF4(52–90)-NHNH_2_; and fragment 5a Nle-RNF4(2–50)-MMP (Scheme). Fragments 2a and 3a were synthesized with an N-terminal Cys(Acm) protecting group to prevent undesired intramolecular cyclization during NCL. Fragments 3a and 5a were prepared as thioesters using the N-acyl-benzimidazolinone (Nbz) strategy.? All peptide fragments were synthesized by Fmoc-SPPS, purified, and obtained in good yields (Supporting Information Figures S1–S7).
Total Chemical Synthesis of RNF4; (A) Primary Sequence of RNF4 with Ligation Sites Highlighted in Red; The Met-1 and Met-136 Residues Were Substituted With the Oxidation-Resistant Norleucine (Nle) Analogues; (B) Synthetic Route of RNF4 Using the C-To-N (Convergent) Ligation Strategy; PG- Acm(Acetamidomethyl), R = MMP(Methyl 3-Mercaptopropionate)
After preparing all peptide fragments, the synthesis was initiated by converting fragment 2a, Cys(Acm)-RNF4(133–158)-NHNH_2_, to the corresponding acyl azide using NaNO_2_ ? in 6 M Gn·HCl, 0.2 M Na_2_HPO_4_ buffer at pH 3.0 and −15 °C for 20 min. A solution of MPAA was prepared in a similar buffer at pH 7 and added to the reaction mixture to generate the thioester intermediate in situ. This activated species was then ligated with fragment 1a, Cys-RNF4(160–190), and the reaction mixture was incubated at 37 °C for 40 min. The reducing agent TCEP was then added, and the reaction was allowed to proceed for an additional 2 h (Scheme). The progress of the ligation was monitored by analytical HPLC, which confirmed the formation of the desired peptide 6 after 4 h. Subsequently, the peptide underwent smooth Acm deprotection with PdCl_2_ at 37 °C for 1 h, quenched with DTT, and was purified.? to give the ligated peptide 6 in ∼32% yield (Supporting Information Figure S8). Before the subsequent ligation, peptide 6 was dissolved in 6 M Gn·HCl, 0.2 M Na_2_HPO_4_, pH 7, containing MPAA and TCEP, and this solution was added to fragment 3a Cys (Acm)-RNF4(92–131)-MMP.
The mixture was incubated at 37 °C for 3 h, during which analytical HPLC confirmed formation of the desired ligated peptide 7. The N-terminal Acm protecting group was then deprotected with PdCl_2_ for 1 h, followed by quenching with DTT, washing, and HPLC purification to give ligated product 7 in ∼12% yield (Supporting Information Figure S9). The ligation of fragment 5a Nle-RNF4(2–50)-MMP with fragment 4a Cys-RNF4(52–90)-NHNH_2_ was performed using a similar strategy. Fragment 4a was dissolved in 6 M Gn·HCl, 0.2 M Na_2_HPO_4_, pH 7, containing TCEP and MPAA, and the resulting solution was added to fragment 5a. The mixture was incubated at 37 °C for 5 h and monitored by analytical HPLC. After completion, the crude mixture was purified, and peptide 8 was isolated (Supporting Information Figure S10) in 56% yield. We employed the same strategy for the final ligation between peptides 8 and 7 (FigureA). Peptide 8, RNF4(1–90)-NHNH_2_, was first converted to the corresponding acyl azide using NaNO_2_ in 6 M Gn·HCl, 0.2 M Na_2_HPO_4_ buffer, pH 3.0, at −15 °C for 20 min. A solution of MPAA was prepared in the same buffer, adjusted to pH 7, and added to the reaction mixture to generate the thioester intermediate in situ. This activated thioester was then ligated with peptide 7, Cys-RNF4(92–190). The reaction mixture was incubated at 37 °C for 40 min. TCEP was then added, and the reaction was allowed to proceed for an additional 4 h (Supporting Information Figure S11). Analytical HPLC confirmed formation of the full-length RNF4 product 9 after 5 h. The ligated polypeptide was isolated in ∼35% yield. Consequently, we performed Western blotting with an anti-RNF4 antibody to further verify the integrity of the final product and to demonstrate that our chemically synthesized RNF4 is correctly recognized by the anti-RNF4 antibody (FigureB,C).
(A) General schematic of the final C-to-N (convergent) ligation between peptides 8 and 7; (B) analytical HPLC and ESI-MS of the final NCL product RNF4 (observed mass: 21,272.6 ± 2 Da; expected mass: 21,270.6 Da); (C) Western blot analysis using anti-RNF4 antibody, 8 = hydrolysis.*
After completing the RNF4 synthesis using the C-to-N ligation strategy, we found that repeated purification and Acm deprotection steps made the process time-consuming and reduced the overall yield. To address these limitations, we explored an alternative N-to-C ligation strategy to minimize purification steps, streamline the assembly, and improve the overall efficiency and yield.
We pursued the synthesis of RNF4 using an N-to-C ligation strategy based on hydrazide chemistry (Scheme). This approach utilizes cysteine-free peptide hydrazides that undergo chemoselective ligation with Cys-containing peptides to form native amide bonds. Importantly, peptide hydrazides are readily accessible via standard Fmoc-based solid-phase peptide synthesis (SPPS). For this purpose, we initially prepared three fragments using CTC-hydrazine resin.?
N-To-C Ligation Approach for the Total Chemical Synthesis of RNF4; (A) Synthetic Route of RNF4 Using the N-To-C Ligation Strategy, R = MMP (Methyl 3-Mercaptopropionate)
However, during the synthesis of fragment 3b using the CTC-hydrazine resin, we observed severe peptide aggregation that hampered further chain elongation. In light of our report and Hartrampf’s recent work? on the effect of the MeDbz linker on reducing aggregation during SPPS of complex peptides, we switched from CTC-hydrazine resin to a Dbz linker coupled to Rink amide resin. Using this strategy, fragment 3b (Cys-RNF4(92–131)) was successfully synthesized by SPPS with excellent purity. Subsequent Dbz-to-Nbz cyclization was performed, followed by conversion of the resulting Nbz moiety to the corresponding hydrazide using 10% hydrazine.? Finally, Acm deprotection with PdCl_2_ afforded the desired fragment 3b (FigureC)
Synthesis of the same peptide sequence using (A) solid-phase peptide synthesis using a CTC-hydrazine resin. (B) solid-phase peptide synthesis using a Dbz linker. (C) Conversion of peptide 3a to 3b via hydrazinolysis (NHNH2). (D) Crude analytical HPLC of the peptide synthesized on CTC-hydrazine resin. (E) Crude analytical HPLC of the peptide synthesized using the Dbz linker. (F) Analytical HPLC of purified peptide 3b.
With all fragments in hand, we initiated the synthesis of RNF4 using the N-to-C ligation strategy (Scheme). We first performed a one-pot ligation of three fragments: 5a, 4a, and 3b. Fragment 4a was dissolved in 6 M Gn·HCl, 0.2 M Na_2_HPO_4_, pH 7, containing TCEP and MPAA, and the resulting solution was added to fragment 5a. The reaction mixture was incubated at 37 °C and monitored by analytical HPLC, which confirmed the formation of the desired ligated peptide 8 after 5 h (Supporting Information Figure S12). Before the subsequent ligation, the reaction mixture was desalted, and the pH was adjusted to ∼3 using 2 N HCl. The intermediate 8 was then converted to the corresponding peptide thioester by treatment with NaNO_2_ in 6 M Gn·HCl, 0.2 M Na_2_HPO_4_ buffer at pH 3.0 at −15 °C for 20 min.
Following this activation, MPAA was added to 6 M Gn·HCl, 0.2 M Na_2_HPO_4_ buffer at pH 7 to generate the thioester intermediate in situ. The resulting activated thioester was then ligated with fragment 3b, Cys-RNF4(92–131)-NHNH_2_, and the reaction mixture was incubated at 37 °C for 40 min. TCEP was then added, and the ligation was allowed to proceed for an additional 15 h. Reaction progress was monitored by analytical HPLC, which confirmed formation of the desired ligated peptide 10, along with the hydrolysis byproduct. The crude reaction mixture was purified by preparative HPLC, affording the isolated peptide in 23% yield (Supporting Information Figure S12). The next ligation between peptide 10, RNF4(1–131)-NHNH_2_, and fragment 2b was carried out by first converting 10 to the corresponding acyl azide using NaNO_2_ in 6 M Gn·HCl, 0.2 M Na_2_HPO_4_ buffer at pH 3 at −15 °C for 20 min, followed by the addition of MPAA as described before. The resulting peptide thioester was then reacted with fragment 2b, and the reaction mixture was incubated at 37 °C for an additional 15 h. Analytical HPLC confirmed formation of the desired ligated peptide 11 (Scheme). Preparative HPLC was used to purify the crude reaction mixture, affording the isolated peptide in 44% yield (Supporting Information Figure S13). For the final ligation, we employed the same strategy, initiating the reaction between peptide 11 and 1a. Peptide 11 was first converted to the corresponding acyl azide using NaNO_2_ in 6 M Gn·HCl, 0.2 M Na_2_HPO_4_ buffer at pH 3.0 at −15 °C for 20 min, followed by the addition of MPAA as described before. The resulting peptide thioester was then reacted with fragment 1a, and the reaction mixture was incubated at 37 °C for an additional 8 h (FigureA). Reaction progress was monitored by analytical HPLC, which confirmed formation of the desired full-length RNF4 product 9. The crude reaction mixture was finally purified by preparative HPLC, affording the isolated product in 23% yield (FigureB).
*(A) General schematic of the final N-to-C ligation between peptides 11 and 1a; (B) analytical HPLC and ESI-MS of the final NCL product RNF4. Represent unidentified mass.
Although the full-length protein was assembled and recognized by the Anti-RNF4 antibody, we first attempted to solubilize it in aqueous buffers to investigate its secondary structure. RNF4 was dissolved in 8 M Gn·HCl and then dialyzed against a degassed buffer containing 20 mM Tris, 1 mM NaCl, 1 mM DTT, and 50 μM ZnCl_2_. Multiple alternative buffer solvents, such as sodium phosphate and HEPES, were also explored in the presence of Zn^2+^ at varying μM concentrations, various redox reagents, and glycerol, but none improved refolding. We further explored a segmental folding strategy using peptide 7, but this approach likewise failed to yield successful folding. However, in all cases, RNF4 precipitated, formed highly aggregated species, failed to fold correctly, and exhibited a low Circular Dichroism signal, indicating limited solubility and intrinsic instability in aqueous solution. We attributed this to the structurally disordered region in RNF4, prompting us to explore alternative strategies to stabilize it, which we are currently pursuing.
Despite this, both synthetic approaches, the C-to-N and the N-to-C, were successful; however, they differed in efficiency. The C-to-N strategy was considerably more time-consuming for synthesizing RNF4, primarily because of the requirement for Acm cysteine deprotection and the associated multiple purification steps after intermediate transformations. These additional steps not only prolonged the overall synthesis but also led to cumulative material losses, reducing the overall yield. Using the C-to-N approach, full-length RNF4 could ultimately be isolated, yielding ∼1.3% (1.2 mg) based on the yield of each ligation. In contrast, the N-to-C approach proved more efficient because it did not require Cys protection, thereby eliminating the need for deprotection steps. This strategy enabled a more streamlined workflow, with fewer reaction steps and significantly fewer purification cycles, affording the isolated product with an improved overall yield of ∼2.3% (0.45 mg).
Conclusions
We accomplished the total chemical synthesis of RNF4 from five peptide fragments using both C-to-N (convergent) and N-to-C ligation strategies. Based on two independent syntheses, our results demonstrate that the N-to-C approach offers advantages for complex protein targets, with reduced reaction time, fewer purification steps, and fewer than the C-to-N (convergent) strategy. To the best of our knowledge, this is the first study to systematically evaluate synthetic routes for preparing the RNF4 protein. These findings provide valuable guidelines for the synthesis of modified RNF4 analogues, such as methylated and phosphorylated variants, and offer a versatile, scalable strategy for generating RNF4 derivatives for biochemical and biomedical investigations.
Experimentalsection
General
Reagents
Fmoc-SPPS was performed either manually in Teflon-filtered syringes purchased from Torviq or using an automated peptide synthesizer (CS336X, CSBIO). 2-Chlorotrityl chloride (2-CTC) resin was purchased from Chem-Impex. All protected amino acids were purchased from Chem-Impex and Iris-Biotech. The activating reagents [(6-c 6-chlorobenzotriazolyl)oxy(dimethylamino)methylidene]dimethylazaniumhexafluorophosphate (HCTU), 1-[bis(dimethylamino)methylene]-1H-1,2,3-triazolo [4,5-b] pyridinium 3-oxid hexafluorophosphate (HATU), and 1- 1-hydroxybenzotriazole monohydrate (HOBt) were purchased from Luxembourg Bio Technologies. All solvents, including N,N-dimethylformamide (DMF), dichloromethane (DCM), acetonitrile (ACN), N,N-diisopropylethylamine (DIEA), Piperidine, Diethyl ether (Et_2_O), and Trifluoroacetic acid (TFA), were purchased from Bio-Lab. Triisopropyl silane (TIPS) was purchased from Sigma-Aldrich. Paraformaldehyde, 4% in PBS, was purchased from Affymetrix. 4–20% MOPS gel (MP42G12) was purchased from Merck. Additional miscellaneous chemicals were purchased from Merck, Strem Chemicals, and Alfa Aesar.
Caution
TFA, DMF, DCM, ACN, Et_2_O, and coupling reagents (HCTU, HATU, and HOBt) are hazardous chemicals with high toxicity and potential health risks. All procedures involving these reagents were performed in a well-ventilated fume hood, with appropriate personal protective equipment (PPE) including lab coats, chemical-resistant gloves, and safety goggles.
List of the Protected Amino
Acids Used in Peptide Synthesis
Fmoc-Ala-OH, Fmoc-Asp(OtBu)–OH, Fmoc-Gly-OH, Fmoc-Glu(OtBu)–OH, Fmoc-His(Trt)–OH, FmocIle-OH, Fmoc-Leu-OH, Fmoc-Phe-OH, Fmoc-Asn(Trt)–OH, Fmoc-Gln(Trt)–OH, Fmoc-Arg(Pbf)–OH, Fmoc- Lys(Boc)–OH, Fmoc-Pro-OH, Fmoc-Tyr(tBu)–OH, Fmoc-Ser(tBu)–OH, Fmoc-Thr(tBu)–OH, Fmoc-Asp(OtBu)- OH, Fmoc-Cys(Trt)–OH, Fmoc-Nle–OH, Fmoc-Val-OH, Fmoc-Asp(OtBu)Thr(ψMe, MePro)–OH, Fmoc-(Dmb)Gly-OH, Fmoc-Leu-Ser(ψMe, MePro)–OH.
Peptide Synthesis, Purification, and Analysis
RNF4 fragments were synthesized on Rink amide MBHA and 2-chlorotrityl chloride (CTC) resins using a synthesizer (CSBio CS336X automated peptide synthesizer). After swelling in DMF for 1 h, Fmoc deprotection was achieved by adding 20% piperidine in DMF for 2 × 5 min cycles. The resin was washed three times with DMF, and amino acid? (AA) was added for 45 min (4 eq AA, 4 eq HCTU, 8 eq DIPEA). When twin couplings occurred, the coupling period decreased to 2 × 30 min. The coupling phase for dipeptides involved 2.5 eq. AA, 2.5 eq. HATU, and 5 eq. DIPEA. Underlined AAs were coupled as dipeptides, while bold AAs were coupled twice. The N-terminal amino acid in the sequence was used in its Boc- and Acm-protected forms. The peptide was cleaved from the resin after 3 h in a cleavage solution (95% TFA, 2.5% H_2_O, 2.5% TIPS). The peptide was then precipitated with cold diethyl ether. After centrifugation (4000 rpm, 15 min, 4 °C), the pellet was dissolved in 50% ACN in H_2_O and lyophilized.
The lyophilized peptide was dissolved in 50% ACN in H_2_O and purified by preparative high-performance liquid chromatography (HPLC) on a Dionex Ultimate 3000 system (Thermo Scientific). H_2_O + 0.05% TFA and ACN +0.05% TFA were used as buffers A and B, respectively. Gradients from 0 to 65% B over 10–45 min were used at a flow rate of 15 mL/min. The peptide mass was confirmed by LCQ Fleet Ion Trap (Thermo Scientific), and purity was assessed by analytical HPLC (0–60% B in 30 min, 1.2 mL/min), using the same solvents as for preparative HPLC.
For resin N-**MeNbz formation^23^ **, the peptide-N-MeDbz-resin was treated with 4-nitrophenyl chloroformate (5 equiv) in DCM while shaking for 1 h at room temperature, repeated 3 times. The resin was drained and treated with a 0.5 M DIEA solution in DMF for 30 min to complete Nbz formation, repeated 3 times. Finally, the resin was washed with DCM and dried under vacuum.
Supplementary Material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Sriramachandran A. M.Dohmen R. J.SUMO-targeted ubiquitin ligases Biochim. Biophys. Acta Gen. Subj.20141843758510.1016/j.bbamcr.2013.08.02224018209 · doi ↗ · pubmed ↗
- 2a Chiariotti L.Benvenuto G.Fedele M.Santoro M.Simeone A.Fusco A.Bruni C. B.Identification and Characterization of a Novel RING-Finger Gene (RNF 4) Mapping at 4p 16.3Genomics 19984725826510.1006/geno.1997.51059479498 · doi ↗ · pubmed ↗
- 3a Tatham M. H.Geoffroy M. C.Shen L.Plechanovova A.Hat-tersley N.Jaffray E. G.Palvimo J. J.Hay R. T.RNF 4 is a poly-SUMO-specific E 3 ubiquitin ligase required for arsenic-induced PML degradation Nat. Cell Biol.20081053854610.1038/ncb 171618408734 · doi ↗ · pubmed ↗
- 4a Kuo C. Y.Li X.Kong X. Q.Luo C.Chang C. C.Chung Y.Shih H. M.Li K. K.Ann D. K.An Arginine-Rich Motif of Ring Finger Protein 4 (RNF 4) Oversees the Recruitment and Degradation of the Phosphorylated and SUM Oylated Krüppel-Associated Box Domain-Associated Protein 1 (KAP 1)/TRIM 28 Protein during Genotoxic Stress. J Biol. Chem.2014289207572077210.1074/jbc.M 114.555672 PMC 411028524907272 · doi ↗ · pubmed ↗
- 5Plechanovova A.Jaffray E. G.Tatham M. H.Naismith J. H.Hay R. T.Structure of a RING E 3 ligase and ubiquitin-loaded E 2 primed for catalysis Nature 201248911512010.1038/nature 1137622842904 PMC 3442243 · doi ↗ · pubmed ↗
- 6Her J.Zheng H.Bunting S. F.RNF 4 sustains Myc-driven tumorigenesis by facilitating DNA replication J. Clin. Invest.202413410 e 16741910.1172/jci 16741938530355 PMC 11093604 · doi ↗ · pubmed ↗
- 7a Pickart C. M.Eddins M. J.Ubiquitin: structures, functions, mechanisms Biochim. Biophys. Acta Gen. Subj.20041695557210.1016/j.bbamcr.2004.09.01915571809 · doi ↗ · pubmed ↗
- 8a Lallemand-Breitenbach V.Jeanne M.Benhenda S.Nasr R.Lei M.Peres L.Zhou J.Zhu J.Raught B.de ThéH.Arsenic degrades PML or PML-RARα through a SUMO-triggered RNF 4/ubiquitin-mediated pathway Nat. Cell Biol.20081054755510.1038/ncb 171718408733 · doi ↗ · pubmed ↗