Encapsulation and Functional Activity of Lactobacillus reuteri Strains: Advances, Challenges, and Perspectives
León‐Espinosa Erika Berenice, Barrios‐Francisco Rigoberto, Colin‐Molina Abraham, Martínez‐Palma Nikte Yoliztli, Rentería‐Ortega Minerva

TL;DR
This paper reviews the functional benefits and encapsulation strategies for Limosilactobacillus reuteri, a probiotic with potential health benefits, and highlights challenges in its stability and application.
Contribution
The paper provides a comprehensive review of L. reuteri's functional properties and encapsulation methods, emphasizing strain-specific effects and future research needs.
Findings
L. reuteri exhibits immunomodulatory, antimicrobial, and anti-inflammatory properties.
Encapsulation methods like ionic gelation and electrospray improve strain survival during processing.
Strain-specific factors and coating materials influence release kinetics and viability.
Abstract
Over the past decade, probiotics have gone from been special health supplements to widely incorporated components in many foods and nutrition products. This has led to more careful checks of their safety, viability, and functional performance under realistic processing and consumption conditions. Within lactic acid bacteria, Limosilactobacillus reuteri (formerly Lactobacillus reuteri) has been extensively investigated for its immunomodulatory, antimicrobial, anti‐inflammatory, antioxidant, and metabolic properties as demonstrated across multiple in vivo and in vitro experimental models. Despite these functional attributes, its viability during food processing and gastrointestinal transit remains strongly influenced by the strain and the characteristics of the delivery matrix; as a result, encapsulation is an essential strategy to preserve cellular integrity and functionality. The…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
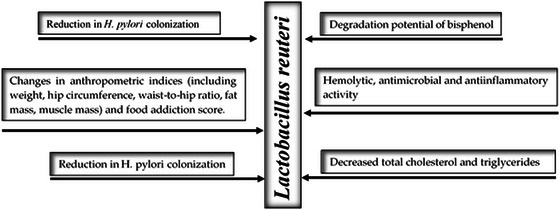 FIGURE 1
FIGURE 1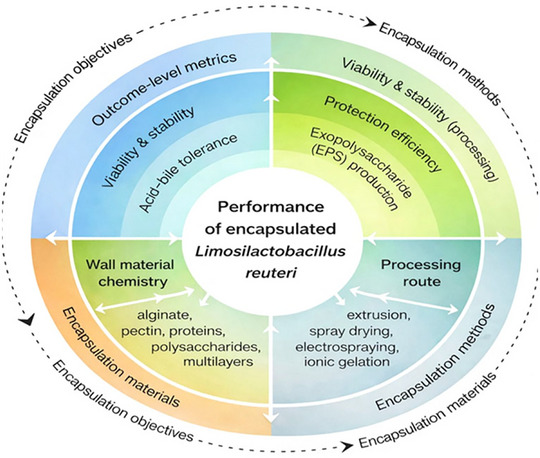 FIGURE 2
FIGURE 2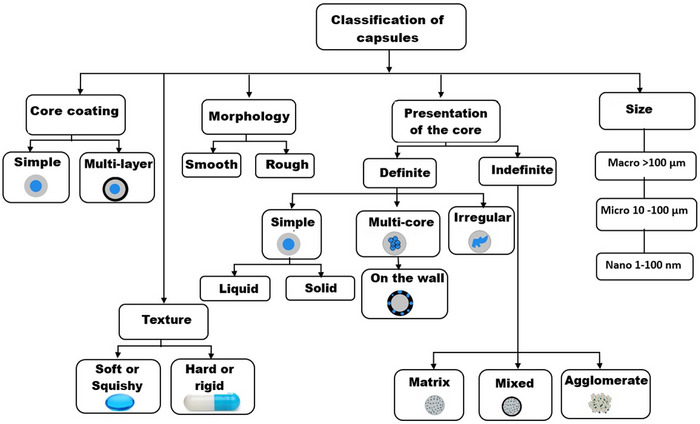 FIGURE 3
FIGURE 3 FIGURE 4
FIGURE 4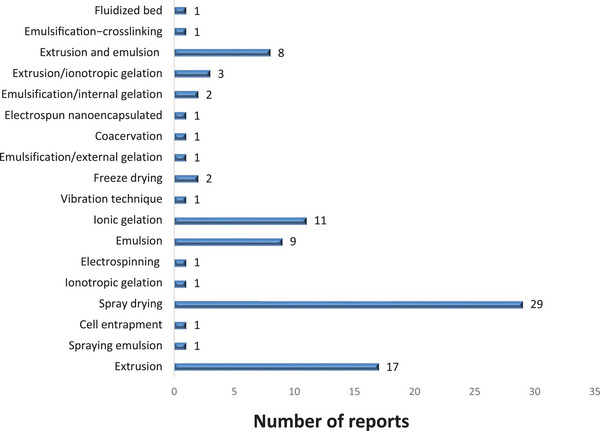 FIGURE 5
FIGURE 5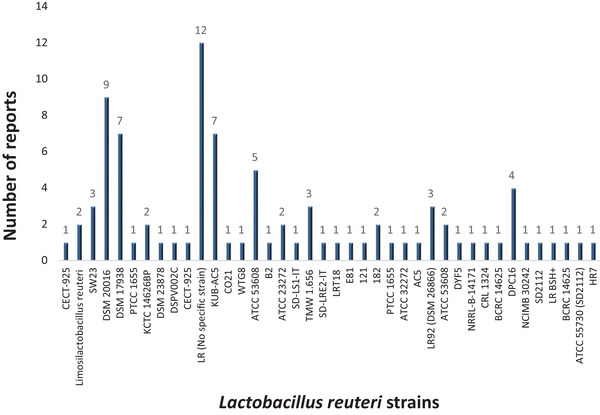 FIGURE 6
FIGURE 6| Strain | Wall material(s) | Encapsulation technique (key conditions) | Postprocessing viability/EE | Storage stability | GI survival/release behavior | Functional outcome | References |
|---|---|---|---|---|---|---|---|
|
| Metal–phenolic network (tannic acid + Fe3 +) + HA‐SH nanoencapsulation | Single‐cell nanoencapsulation (metal coordination + HA‐SH coating) | Maintains ∼6 log CFU/mL under simulated GI, heat, oxidative stress | Enhanced viability under storage and stress relative to free cells | Improved survival under simulated GI conditions | High tolerance to adverse environments; preserved viability and growth | Yang et al. ( |
|
| Metal–phenolic network (tannic acid + Fe3 +) + HA‐SH nanoencapsulation | | Single‐cell nanoencapsulation (metal coordination with Fe3 + and tannic acid followed by HA‐SH coating) | Maintains ∼6 log CFU/mL under simulated gastrointestinal, thermal and oxidative stress conditions | Enhanced viability under storage and stress conditions compared to free cells | Improved survival under simulated gastrointestinal conditions | High tolerance to adverse environments; preserved viability and growth performance | Yang et al. ( |
|
| Sodium alginate (1% w/v) as first layer + chitosan (1% w/v) as second layer on MCC carrier (double‐coated microcapsules) | Double coating by Wurster fluidized‐bed process (first layer: sodium alginate 0.5%–1.5% w/v; second layer: chitosan 0.5%–1.5% w/v or arabic gum 1.5%–6% w/v; optimized at alginate 1% + chitosan 1%) | Relative survival after simulated gastric conditions (pH 2, 1 h): 11.6% for double‐coated alginate–chitosan microcapsules; significantly higher than monolayer systems | Not evaluated during long‐term storage; focus on thermal and acid resistance | Improved resistance under simulated gastric conditions and heat treatment (80°C for 15 min: 7.31% survival; 30 min: 0.63%) compared to single‐layer microcapsules | Double‐layer architecture enhances protection against low pH and high temperature, demonstrating structure–function relationship between multilayer coating and probiotic survival | Zaghari et al. ( |
|
| Sodium alginate (Na‐alg) and sodium alginate–starch maleate (Na‐alg + SM) calcium‐crosslinked beads | Microencapsulation by extrusion–gelation: Na‐alg or Na‐alg + SM solutions dropped into CaCl2 (0.1 M) to form beads; optimization by RSM identified 1% Na‐alg + 1.5% SM as optimal formulation | Encapsulation yield: 70.2% (Na‐alg) and 88.4% (Na‐alg + SM); postencapsulation viability 7.80 and 7.88 log CFU/mL, respectively | Viability remained stable after microencapsulation; long‐term storage stability not specifically evaluated | Higher survival in simulated gastric juice (pH 2.5, 120 min): 8.88 log CFU/mL (Na‐alg + SM) vs. 8.76 log CFU/mL (Na‐alg) and 7.72 log CFU/mL for free cells | Alginate–starch maleate composite improves protection under acidic gastric conditions, demonstrating a clear structure–function relationship between biopolymer composition and probiotic survival | Popović et al. ( |
|
| Polysaccharide‐based bionanocomposite: inulin + pectin + sodium alginate matrix reinforced with MgO nanoparticles | Microencapsulation by extrusion–gelation into CaCl2 (0.1 M); polysaccharide prebiotic solution (1% w/v) containing MgO NPs (5 µg/mL), followed by microwave drying (400 W, 7.5 min) | Encapsulation efficiency 97.57%; viability after drying 99.37%; markedly higher survival compared to free cells | During 28 days storage, encapsulated cells showed reduced losses compared to free cells (2.56 log CFU/g at 4°C and 3.04 log CFU/g at 25°C prevented by encapsulation) | High survival under simulated gastrointestinal conditions (91.74%), whereas free cells lost 2.77 log CFU/g | Synergistic effect of prebiotic matrix and MgO nanoparticles enhances buffering capacity, limits acid and bile penetration, and improves probiotic survival and delivery to the colon | Mohamadzadeh et al. ( |
|
| Hempseed protein isolate (HPI) coencapsulated with Cyclocarya paliurus leaf extracts (protein–phenolic matrix) | Microencapsulation by protein–polyphenol complexation using HPI and C. paliurus leaf extracts at optimized 9:1 (w/w) ratio; drying to obtain stable powder | Encapsulation efficiency: 93.06%; post‐GITS viability 7.2 log CFU/g compared to 5.5 log CFU/g for free cells | Viability maintained above 6 log CFU/g after 120 days at −20°C, 4°C, and 25°C | Significantly higher survival after in vitro gastrointestinal tract simulation compared to free cells | Protein–phenolic interactions create compact matrices that reduce oxidation and acid penetration, enhancing probiotic stability and survivability during storage and GI transit | Lau et al. ( |
|
| Pea protein microgel (MG)–reinforced low‐methoxyl pectin (LMP) hydrogel beads filled with pectic oligosaccharides (POS) | Two‐step encapsulation: preparation of pea protein microgels followed by entrapment into Ca2 +‐crosslinked LMP hydrogel beads containing 0%–0.4% POS (MG/LMP/POS system) | Encapsulation efficiency increased with POS concentration; MG/LMP/POS0.4 showed the highest retention of viable cells after processing and simulated digestion | MG‐reinforced synbiotic hydrogel beads showed higher thermal and storage stability than MG alone | Higher survival throughout simulated oral, gastric and intestinal phases; most cells released under simulated colonic conditions after 48 h | Microgel‐reinforced hydrogel architecture and POS incorporation create a compact, dual‐network matrix that reduces cell leakage, improves resistance to upper GI stress, and enables colon‐targeted release | Yi et al. ( |
|
| Kudzu starch–hemp protein complex coacervates (KS–HP CC) | Complex coacervation (KS:HP ratio 1:2 w/w, pH 5.0) followed by spray drying (inlet 160°C, outlet 93°C) | High postprocessing viability; KS–HP CC retained ∼89% viability after simulated gastrointestinal digestion, ∼83.2% after 30 days storage at 25°C, and ∼94.2% after thermal treatment (50°C, 10 min) | Significantly improved storage stability over free cells and single‐material KS or HP microcapsules during 30 days at room temperature | Enhanced survival during simulated gastric and intestinal digestion compared to free cells and single‐wall systems | Protein–polysaccharide coacervate structure provides a compact protective matrix, improving resistance to gastrointestinal, thermal, and storage stresses while preserving probiotic functionality | Hamdi et al. ( |
|
| Low‐acyl gellan gum (LAG) + High‐acyl gellan gum (HAG) + Ca2 + (binary polysaccharide system) | Internal ionic gelation (emulsification with Span 80, Ca2 + crosslinking, Box–Behnken optimization) | Encapsulation efficiency up to 95.5%; viability after encapsulation up to 97.43% (optimized conditions) | Stable viable counts; maintained ≥8.02 log CFU/mL after simulated GI exposure | Viability decreased from 9.50 to 8.46 log CFU/mL after SGF (2 h) and to 8.02 log CFU/mL after SIF (2 h); free cells dropped to undetectable levels | Enhanced protection against gastric acid and bile salts; suitable for functional food applications | González‐Cuello ( |
|
| Alginate and chitosan–alginate matrices | Vibrating technology (extrusion‐based encapsulation); alginate beads with or without chitosan coating; CaCl2 gelation; optional freeze‐drying | High encapsulation efficiency (∼97%); maintained viability after encapsulation and freeze‐drying (≈100% survival immediately after lyophilization); reduced log losses under thermal, osmotic, and oxidative stress compared to free cells | Improved stability during 28 days of storage at 4°C (≈1 log reduction vs. ≈3 log for free cells); better protection in Ringer and NaCl solutions | Significantly enhanced survival under simulated gastrointestinal conditions (gastric pH 2.5 with pepsin and intestinal bile salts); chitosan–alginate capsules showed superior protection against bile stress | Preserved functional activity, including reuterin production and antimicrobial activity; maintained probiotic functionality while improving technological robustness | De Prisco et al. ( |
|
| Chia seed mucilage + sodium alginate | Electrohydrodynamic spraying (EHDA) and dripping mode; ionic crosslinking with Ca2 +; nonthermal encapsulation | Viability ≈ 9.9 log CFU/mL after encapsulation and lyophilization; EE = 99.0 ± 0.01% (EHDA) and 99.0 ± 0.16% (DM) | High stability after freeze‐drying; maintained viability during powder formulation | Encapsulation matrix protected cells under simulated gastrointestinal conditions, improving survival compared to free cells | Successful incorporation into whey‐based functional powders; preserved probiotic viability and functional potential | Cid‐Córdoba et al. ( |
|
| Alginate beads coated with chitosan (alginate–chitosan microcapsules) | Extrusion–gelation technique: cell–alginate suspension extruded into CaCl2 solution to form beads, followed by chitosan coating via electrostatic interaction | Encapsulation efficiency > 90%; encapsulated cells showed significantly higher viability than free cells after encapsulation and thermal treatment | Improved stability during storage at 4°C; encapsulated cells showed lower viability loss compared to free cells over storage period | Encapsulated cells exhibited significantly higher survival under simulated gastrointestinal conditions (pH 2.0 with pepsin and bile salts) compared to free cells | Alginate–chitosan coating provided an effective physical barrier against acid and bile, enhancing probiotic survival and suitability for functional food applications | Chitprasert et al. ( |
|
| Chia seed mucilage + sodium alginate (binary polysaccharide hydrogel matrix) | Electrohydrodynamic spraying (EHDA); ionic crosslinking with CaCl2; nonthermal encapsulation followed by freeze‐drying | High postprocessing viability; encapsulation efficiency ≈ 98%–99%; viable counts remained > 9 log CFU/mL after encapsulation and lyophilization | Good storage stability of dried powders; encapsulated cells maintained viability during storage compared to free cells | Encapsulated cells showed significantly higher survival under simulated gastrointestinal conditions (gastric and intestinal phases) than free cells | Hydrogel microstructure provided effective protection against acidic and bile environments, enabling application in functional food systems | Cerón‐Córdoba et al. ( |
|
| Food matrix/system | Encapsulation strategy | Functional activity or experimentally evaluated | Outcome on functional activity | References |
|---|---|---|---|---|---|
|
| Legume‐based spread | Not applied | Yes | High probiotic survival during refrigerated storage, supporting functional performance in plant‐based matrices | Barboza Martínez et al. ( |
|
| Cold‐smoked salmon | Not applied | Yes (antimicrobial) | Reuterin production effectively inhibited Listeria monocytogenes | Montiel et al. ( |
|
| Model system | Alginate‐based encapsulation | Yes (antimicrobial) | Encapsulation preserved antimicrobial activity by protecting viable cells and entrapped metabolites | Juárez‐Tomás et al. ( |
|
| Powdered food model | Spray drying with lactose and ascorbic acid | Yes (antioxidant) | Coencapsulation preserved antioxidant activity and improved stability during storage | Rodklongtan and Chitprasert ( |
|
| Mango pulp‐based beverage | Spray drying with maltodextrin and inulin | No | Encapsulation improved probiotic stability, supporting preservation of functional potential | Gámez et al. ( |
|
| Meat‐based system | Not applied | Yes (antioxidant) | Postbiotic application enhanced antioxidant capacity of the meat matrix | Jalali et al. ( |
|
| Functional activity | References |
|---|---|---|
| FCQHC8L6 y FYNDL13 | Anti‐inflammatory and Antimicrobial | Lin et al. ( |
| BR120 | Probiotic, anti‐inflammatories, radical scavengers, acid tolerance, bile tolerance and intestinal adhesion | Ju et al. ( |
| ATCC 53608 | Antimicrobial activity against viruses, fungi, protozoa and gram‐positive and gram‐negative bacteria | Ma et al. ( |
| ATCC PTA5289 | Inhibits the acid tolerance response in oral bacteria | Boisen et al. ( |
| DSM 17938 | Colonize the gastrointestinal tract effectively and shorten the duration of watery diarrhea associated with rotavirus infection time significantly | Sun et al. ( |
| NCIMB 30242 | Reduced total cholesterol (TC) | Liu et al. ( |
| DSM 20016 y DSM 17938 | Antimicrobial characteristics, such as antifungal activity and a broad‐spectrum of activity against Gram‐positive and negative bacteria | Rodrigues et al. ( |
| DSM17648 | Reduction in H. pylori colonization | |
| DSM 17938 | Antimicrobial activity | Zhang et al. ( |
| ATCC 53608 | Antimicrobial activity | Soltani Lak et al. ( |
| DSM 17648 | Reduction in H. pylori colonization | Yang et al. ( |
| DSM 17648 | Reduction in H. pylori colonization | Dargenio et al. ( |
| ATCC 23272 | Probiotic | Karimi et al. ( |
| KUB‐AC5 | Probiotic | Diệp Huy Vũ et al. ( |
| B2 | Antimicrobial activity and Acid resistance and Bile Salt Tolerance | Popović et al. ( |
| B‐14171. | Inhibit the growth of pathogenic bacteria and improve the immune system of the host. | Mis Solval et al. ( |
| E81 | Effect against H. pylori infection | Ceylan et al. ( |
| ATCC 55730 | Degradation potential of bisphenol | Ju et al. ( |
| B‐14171 | Antimicrobial | Mu et al. ( |
| ATCC 55730 y ATCC PTA 5282 | Antimicrobial and anti‐inflammatory properties | Kaur et al. ( |
| ATCC 55730 | Treatment of diarrhea, hernia and intestinal backflow | Indrio et al. ( |
| DSM 20016T; ATCC 23272 | Biotherapeutic potential | Juárez Tomás et al. ( |
| DSM17648 | Reduction in H. pylori colonization | Holz et al. ( |
| ATCC PTA 6475 y ATCC PTA 5289 | Antimicrobial activity | Jones and Versalovic ( |
| NCIMB 30242 | Reduced total cholesterol (TC) | Casas and Dobrogosz ( |
| CRL 1098 | Decreased total cholesterol and triglycerides | Taranto et al. ( |
| Core materials | Encapsulation efficiencies (EE) | References |
|---|---|---|
| Protein isolates | 93.06 | Lau et al. ( |
| Beads of alginate hydrogel embedded with gellan fluid gel | 86.64 | Najafpour et al. ( |
| Whey proteins and gum Arabic | 82.11 | Atta et al. ( |
| Mucin and hempseed protein isolate | Not reported | Lau and Quek ( |
| Hydrogel formed by hyaluronic acid modified with thiol groups of L‐cysteine | 38.15 | Yang et al. ( |
| Polysaccharide‐based bionanocomposite. Inulin, polydextrose, and pectin were utilized as prebiotics, and magnesium oxide nanoparticles as reinforcing agent in the microgel structure | 99.37 | Mohamadzadeh et al. ( |
| Inulin and maltodextrin | 43.27 | Ceron‐Cordoba et al. ( |
| Whey protein isolate or sodium caseinate (Cas), followed by pectin | 92 | Rodríguez et al. ( |
| Chitosan and Ca‐alginate beads | 92.5 | Jeznienė et al. ( |
| Whey protein concentrate and gum arabic | 67 | Teymoori et al. ( |
| Sodium alginate and tomato seed mucilage | 91.54 | Ganje et al. ( |
| Alginate or pectin followed by polymeric coating (with whey protein concentrate or chitosan) | 91 | Mosquera‐Vivas et al. ( |
| Alginate‐inulin | Not reported | Yadav et al. ( |
| Rice protein, pea protein and inulin | 96.99 | Mudgil et al. ( |
| Gelatin and pregelatinized starch, maltodextrin and Cheese whey permeate | 50.53 | Zimmermann et al. ( |
| Binary matrix of inulin and maltodextrin | 91.35 | Cerón Córdoba et al. ( |
| Cruciferin/alginate | Not reported | Akbari et al. ( |
| Chitosan‐coated alginate‐inulin matrix | 95.85 | Parsana et al. ( |
| Skim milk and infant formula | 90 | Alves Gragnani Vido et al. ( |
| Binary matrix composed of inulin and maltodextrin | 89.76 | Jurado‐Gámez et al. ( |
| Skim milk, sodium glutamate, polyvinylpyrrolidone, maltodextrin and gelatin | Not reported | Wang et al. ( |
| Sodium alginate and sesame seed oil | 89.22 | Javed et al. ( |
| Alginate‐konjac gum | Not reported | Rodrigues et al. ( |
| Acacia gum | Not reported | do Nascimento et al. ( |
| Alginate | 97 | Abuqwider et al. ( |
| Pequi oil | 96 | Cedran et al. ( |
| fish skin gelatin | Zhang et al. ( | |
| Sodium hyaluronate | 79 | Li et al. ( |
| Isolated lignin complex from whey protein (WPI) and lignin | 100 | Diệp Huy Vũ et al. ( |
| Hydrophobic surfactant into the MCT | 84 | Marefati et al. ( |
| pomegranate ( | 46 | Bron et al. ( |
| Sunflower oil, Skim milk powder | 98.93 | Kil et al. ( |
| novel enzymatically modified resistant starch type III | 86.73 | Khan et al. ( |
| yellow passion fruit ( | 99 | Santos Monteiro et al. ( |
| PVA | 78 | Ceylan et al. ( |
| Gum arabic | 63 | Guergoletto et al. ( |
| gelatin | 24.4 | Guergoletto et al. ( |
| Holy basil essential oil | 47.73 | Rodklongtan and Chitprasert ( |
| Milk powder | 74.91 | Liu et al. ( |
| xanthan–gellan gum | 72,5 | Juárez Tomás et al. ( |
| Aluminum carboxymethyl cellulose–rice bran | 99.71 | Chitprasert et al. ( |
| Palm oil was of food grade | 99 | Chitprasert et al. ( |
| Mixtures of sodium alginate with other biopolymers | 83.08 | Akbari et al. ( |
| Sodium alginate | 70 | Ebrahimi Monfared et al. ( |
| Maltodextrin | 57 | Guergoletto et al. ( |
| Blends of maltodextrin with other biopolymers | 89.75 | Gámez et al. ( |
| Whey protein isolate, sucrose | 68.4 | Sompach et al. ( |
| Whey | 0.54 | Jantzen et al. ( |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsProbiotics and Fermented Foods · Microencapsulation and Drying Processes · Microbial Metabolites in Food Biotechnology
Introduction
1
The growing demand of foods that provide health benefits has accelerated the development of formulations fortified with probiotic microorganisms (Damián et al. 2022; Das et al. 2016). Within this group, strains of Limosilactobacillus reuteri have garnered sufficient interest due to their specific functions, such as immunomodulation, production of antimicrobial metabolites, for example, reuterin, reduction of total cholesterol and triglycerides, inhibition of Helicobacter pylori, and their ability to colonize different host niches.
Despite its multiple benefits, the application of L. reuteri in food matrices remains complex, as the microorganism is sensitive to food processing conditions, for example, heating, dehydration, and oxygen exposure, but also to gastrointestinal challenges such as pH, bile salts, and digestive enzymes (Algaithi et al. 2022). These limitations tend to reduce the viability and functional activity of L. reuteri, compromising its efficacy when administered through food matrices.
In this sense, encapsulation has emerged as a key strategy to improve the stability, survival, and targeted release of L. reuteri, both in food and throughout the gastrointestinal tract (Anal and Singh 2007; Cook et al. 2012). Several studies have explored the use of different coating materials, for example, alginate, pectin, proteins, starch, and polysaccharides, as well as different encapsulation methods such as extrusion, emulsification, ionic gelation, electrospraying, coacervation, and spray drying, with the aim of improving probiotic functionality (Algaithi et al. 2022; Rokka and Rantamaki 2010; Nazzarro et al. 2012).
However, despite the increasing number of experimental studies, literature still presents several gaps in this area. Nowadays, most research focuses on the individual strains, isolated functional activity, or specific encapsulation approaches, without adequately integrating how strain‐dependent characteristics align with encapsulation design or how these interactions may be translated into technological performance in real word food systems (Gbassi and Vandamme 2012; Rokka and Rantamaki 2010). According to the above, this review aims to integrate current knowledge on the functional properties of L. reuteri with the role of encapsulation strategies in preserving its viability during processing and gastrointestinal transit, considering how strain characteristics, encapsulation methods and coating materials impact functional activity and technological viability in food matrices
Background on Probiotics and Viability in Food Systems
1.1
Globally, there has been an increasing interest in foods that provide health‐promoting benefits beyond basic nutrition, along with a growing demand for transparency regarding ingredients, additives, and the inclusion of functional components such as probiotic microorganisms. In this context, various validation approaches such as standardizer in vitro gastric digestion models have been developed to assess digestibility, bioaccessibility, stability, and release of nutrients and bioactive compounds. These methodologies have been extensively used to assess the survival and stability of probiotic microorganisms under simulated gastrointestinal conditions, thereby providing valuable insights into the challenges associated with their incorporation into food matrices (Brodkorb et al. 2019; Minekus et al. 2014).
A fundamental requirement for probiotics to exert their intended physiological effects is their capacity to reach the intestine in sufficient numbers while maintaining viability during food processing, storage, and gastrointestinal transit (FAO/WHO 2002; Tripathi and Giri 2014). However, exposure to heat, mechanical stress, oxygen, moisture, acidity, and bile salts can substantially decrease cell survival. Consequently, preserving probiotic viability remains one of the major technological challenges in the development of functional foods (Tripathi and Giri 2014).
State of the Art on Encapsulated Limosilactobacillus reuteri
1.2
Among the assorted genera used in probiotic applications, it can be found Lactobacillus and Bifidobacterium; however, L. reuteri stands out due to its unique biological and functional properties (Algaithi et al. 2022). Particularly, LR is an intestinal symbiont that can colonize gastrointestinal tract in various mammals, including humans. It is tolerant to gastric acidity and bile, but also produces many of the essential compounds responsible for gut health (Liu et al. 2016; Yu et al. 2023).
Nevertheless, in recent decades, there has been a considerable decrease in the presence of L. reuteri in humans, which have been correlated with an increase in the incidence of inflammatory diseases (e.g., ulcerative colitis, Crohn's disease, and indeterminate colitis). For the above, research works attending the development of functional foods, particularly those containing probiotic strains, are imperative for addressing some of the public health issues mentioned.
Over the past decade, research on L. reuteri encapsulation has expanded substantially, motivated by the need to enhance cell survival during food processing, storage, and gastrointestinal transit (Anal and Singh 2007; De Prisco and Mauriello 2016; Tripathi and Giri 2014). Various encapsulation materials, such as alginate, pectin, starch derivatives, chitosan, whey proteins, mucilage, inulin, and multilayer coatings, have been investigated alongside methods including extrusion, emulsification, ionic gelation, spray drying, and electrospraying (Anal and Singh 2007; De Prisco and Mauriello 2016). While these approaches have consistently demonstrated improvements in viability; the extent of protection is highly dependent on the chemical composition of the wall material, capsule structure, drying conditions, and strain‐specific physiological characteristics.
A key challenge in current research is the strain‐dependent response of L. reuteri. Strains such as DSM 17938, DSM 20016, CRL 1324, ATCC 55730, and others show significant differences in acid–bile tolerance, adhesion, exopolysaccharide (EPS) production, reuterin synthesis, and dehydration sensitivity (Britton et al. 2014; Mu et al. 2018a; Walter et al. 2011). Despite these differences, many studies assess encapsulation effects on specific functions without directly linking these outcomes to material selection or capsule architecture. This limits the ability to establish structure–function relationships or identify optimal material–strain combinations (De Prisco and Mauriello 2016; Tripathi and Giri 2014).
Although previous reviews have explored either the probiotic activity or encapsulation strategies of L. reuteri, only a few numbers of they have provided an integrated analysis that connects specific strains, coating materials, encapsulation methods, and their technological and health‐related implications. Most existing reviews either focus narrowly on clinical aspects or provide general overviews of probiotic encapsulation without discriminating against bacterial species or materials. Table 1 addresses gaps in the general overview by providing an integrative comparison of representative encapsulation strategies for L. reuteri. It organizes studies by wall material and processing technique, and highlights encapsulation efficiency (EE), postprocessing viability, gastrointestinal survival, and functional performance.
TABLE 1: Integrative comparison of encapsulation strategies for Limosilactobacillus reuteri: Materials, processing methods, and quantitative functional outcomes.
<table><col align="left"/><col align="center"/><col align="center"/><col align="center"/><col align="center"/><col align="center"/><col align="center"/><col align="center"/><thead><tr><th align="left" rowspan="1" colspan="1">Strain</th><th align="center" rowspan="1" colspan="1">Wall material(s)</th><th align="center" rowspan="1" colspan="1">Encapsulation technique (key conditions)</th><th align="center" rowspan="1" colspan="1">Postprocessing viability/EE</th><th align="center" rowspan="1" colspan="1">Storage stability</th><th align="center" rowspan="1" colspan="1">GI survival/release behavior</th><th align="center" rowspan="1" colspan="1">Functional outcome</th><th align="center" rowspan="1" colspan="1">References</th></tr></thead><tbody><tr><td align="left" rowspan="1" colspan="1"> <italic>Limosilactobacillus reuteri</italic> HR7</td><td align="center" rowspan="1" colspan="1">Metal–phenolic network (tannic acid + Fe<sup>3</sup> <sup>+</sup>) + HA‐SH nanoencapsulation</td><td align="center" rowspan="1" colspan="1">Single‐cell nanoencapsulation (metal coordination + HA‐SH coating)</td><td align="center" rowspan="1" colspan="1">Maintains ∼6 log CFU/mL under simulated GI, heat, oxidative stress</td><td align="center" rowspan="1" colspan="1">Enhanced viability under storage and stress relative to free cells</td><td align="center" rowspan="1" colspan="1">Improved survival under simulated GI conditions</td><td align="center" rowspan="1" colspan="1">High tolerance to adverse environments; preserved viability and growth</td><td align="center" rowspan="1" colspan="1">Yang et al. (<xref>2025</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1"> <italic>Limosilactobacillus reuteri</italic> HR7</td><td align="center" rowspan="1" colspan="1">Metal–phenolic network (tannic acid + Fe<sup>3</sup> <sup>+</sup>) + HA‐SH nanoencapsulation |</td><td align="center" rowspan="1" colspan="1">Single‐cell nanoencapsulation (metal coordination with Fe<sup>3</sup> <sup>+</sup> and tannic acid followed by HA‐SH coating)</td><td align="center" rowspan="1" colspan="1">Maintains ∼6 log CFU/mL under simulated gastrointestinal, thermal and oxidative stress conditions</td><td align="center" rowspan="1" colspan="1">Enhanced viability under storage and stress conditions compared to free cells</td><td align="center" rowspan="1" colspan="1">Improved survival under simulated gastrointestinal conditions</td><td align="center" rowspan="1" colspan="1">High tolerance to adverse environments; preserved viability and growth performance</td><td align="center" rowspan="1" colspan="1">Yang et al. (<xref>2025</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1"> <italic>Lactobacillus reuteri</italic> (PTCC‐1655)</td><td align="center" rowspan="1" colspan="1">Sodium alginate (1% w/v) as first layer + chitosan (1% w/v) as second layer on MCC carrier (double‐coated microcapsules)</td><td align="center" rowspan="1" colspan="1">Double coating by Wurster fluidized‐bed process (first layer: sodium alginate 0.5%–1.5% w/v; second layer: chitosan 0.5%–1.5% w/v or arabic gum 1.5%–6% w/v; optimized at alginate 1% + chitosan 1%)</td><td align="center" rowspan="1" colspan="1">Relative survival after simulated gastric conditions (pH 2, 1 h): 11.6% for double‐coated alginate–chitosan microcapsules; significantly higher than monolayer systems</td><td align="center" rowspan="1" colspan="1">Not evaluated during long‐term storage; focus on thermal and acid resistance</td><td align="center" rowspan="1" colspan="1">Improved resistance under simulated gastric conditions and heat treatment (80°C for 15 min: 7.31% survival; 30 min: 0.63%) compared to single‐layer microcapsules</td><td align="center" rowspan="1" colspan="1">Double‐layer architecture enhances protection against low pH and high temperature, demonstrating structure–function relationship between multilayer coating and probiotic survival</td><td align="center" rowspan="1" colspan="1">Zaghari et al. (<xref>2020</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1"> <italic>Lactobacillus reuteri</italic> B2</td><td align="center" rowspan="1" colspan="1">Sodium alginate (Na‐alg) and sodium alginate–starch maleate (Na‐alg + SM) calcium‐crosslinked beads</td><td align="center" rowspan="1" colspan="1">Microencapsulation by extrusion–gelation: Na‐alg or Na‐alg + SM solutions dropped into CaCl<sub>2</sub> (0.1 M) to form beads; optimization by RSM identified 1% Na‐alg + 1.5% SM as optimal formulation</td><td align="center" rowspan="1" colspan="1">Encapsulation yield: 70.2% (Na‐alg) and 88.4% (Na‐alg + SM); postencapsulation viability 7.80 and 7.88 log CFU/mL, respectively</td><td align="center" rowspan="1" colspan="1">Viability remained stable after microencapsulation; long‐term storage stability not specifically evaluated</td><td align="center" rowspan="1" colspan="1">Higher survival in simulated gastric juice (pH 2.5, 120 min): 8.88 log CFU/mL (Na‐alg + SM) vs. 8.76 log CFU/mL (Na‐alg) and 7.72 log CFU/mL for free cells</td><td align="center" rowspan="1" colspan="1">Alginate–starch maleate composite improves protection under acidic gastric conditions, demonstrating a clear structure–function relationship between biopolymer composition and probiotic survival</td><td align="center" rowspan="1" colspan="1">Popović et al. (<xref>2021</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1"> <italic>Limosilactobacillus reuteri</italic> ATCC 23272</td><td align="center" rowspan="1" colspan="1">Polysaccharide‐based bionanocomposite: inulin + pectin + sodium alginate matrix reinforced with MgO nanoparticles</td><td align="center" rowspan="1" colspan="1">Microencapsulation by extrusion–gelation into CaCl<sub>2</sub> (0.1 M); polysaccharide prebiotic solution (1% w/v) containing MgO NPs (5 µg/mL), followed by microwave drying (400 W, 7.5 min)</td><td align="center" rowspan="1" colspan="1">Encapsulation efficiency 97.57%; viability after drying 99.37%; markedly higher survival compared to free cells</td><td align="center" rowspan="1" colspan="1">During 28 days storage, encapsulated cells showed reduced losses compared to free cells (2.56 log CFU/g at 4°C and 3.04 log CFU/g at 25°C prevented by encapsulation)</td><td align="center" rowspan="1" colspan="1">High survival under simulated gastrointestinal conditions (91.74%), whereas free cells lost 2.77 log CFU/g</td><td align="center" rowspan="1" colspan="1">Synergistic effect of prebiotic matrix and MgO nanoparticles enhances buffering capacity, limits acid and bile penetration, and improves probiotic survival and delivery to the colon</td><td align="center" rowspan="1" colspan="1">Mohamadzadeh et al. (<xref>2025</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1"> <italic>Limosilactobacillus reuteri</italic> DPC16</td><td align="center" rowspan="1" colspan="1">Hempseed protein isolate (HPI) coencapsulated with Cyclocarya paliurus leaf extracts (protein–phenolic matrix)</td><td align="center" rowspan="1" colspan="1">Microencapsulation by protein–polyphenol complexation using HPI and C. paliurus leaf extracts at optimized 9:1 (w/w) ratio; drying to obtain stable powder</td><td align="center" rowspan="1" colspan="1">Encapsulation efficiency: 93.06%; post‐GITS viability 7.2 log CFU/g compared to 5.5 log CFU/g for free cells</td><td align="center" rowspan="1" colspan="1">Viability maintained above 6 log CFU/g after 120 days at −20°C, 4°C, and 25°C</td><td align="center" rowspan="1" colspan="1">Significantly higher survival after in vitro gastrointestinal tract simulation compared to free cells</td><td align="center" rowspan="1" colspan="1">Protein–phenolic interactions create compact matrices that reduce oxidation and acid penetration, enhancing probiotic stability and survivability during storage and GI transit</td><td align="center" rowspan="1" colspan="1">Lau et al. (<xref>2025</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1"> <italic>Limosilactobacillus reuteri</italic> (strain not specified)</td><td align="center" rowspan="1" colspan="1">Pea protein microgel (MG)–reinforced low‐methoxyl pectin (LMP) hydrogel beads filled with pectic oligosaccharides (POS)</td><td align="center" rowspan="1" colspan="1">Two‐step encapsulation: preparation of pea protein microgels followed by entrapment into Ca<sup>2</sup> <sup>+</sup>‐crosslinked LMP hydrogel beads containing 0%–0.4% POS (MG/LMP/POS system)</td><td align="center" rowspan="1" colspan="1">Encapsulation efficiency increased with POS concentration; MG/LMP/POS0.4 showed the highest retention of viable cells after processing and simulated digestion</td><td align="center" rowspan="1" colspan="1">MG‐reinforced synbiotic hydrogel beads showed higher thermal and storage stability than MG alone</td><td align="center" rowspan="1" colspan="1">Higher survival throughout simulated oral, gastric and intestinal phases; most cells released under simulated colonic conditions after 48 h</td><td align="center" rowspan="1" colspan="1">Microgel‐reinforced hydrogel architecture and POS incorporation create a compact, dual‐network matrix that reduces cell leakage, improves resistance to upper GI stress, and enables colon‐targeted release</td><td align="center" rowspan="1" colspan="1">Yi et al. (<xref>2025</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1"> <italic>Limosilactobacillus reuteri</italic> DSM 17938</td><td align="center" rowspan="1" colspan="1">Kudzu starch–hemp protein complex coacervates (KS–HP CC)</td><td align="center" rowspan="1" colspan="1">Complex coacervation (KS:HP ratio 1:2 w/w, pH 5.0) followed by spray drying (inlet 160°C, outlet 93°C)</td><td align="center" rowspan="1" colspan="1">High postprocessing viability; KS–HP CC retained ∼89% viability after simulated gastrointestinal digestion, ∼83.2% after 30 days storage at 25°C, and ∼94.2% after thermal treatment (50°C, 10 min)</td><td align="center" rowspan="1" colspan="1">Significantly improved storage stability over free cells and single‐material KS or HP microcapsules during 30 days at room temperature</td><td align="center" rowspan="1" colspan="1">Enhanced survival during simulated gastric and intestinal digestion compared to free cells and single‐wall systems</td><td align="center" rowspan="1" colspan="1">Protein–polysaccharide coacervate structure provides a compact protective matrix, improving resistance to gastrointestinal, thermal, and storage stresses while preserving probiotic functionality</td><td align="center" rowspan="1" colspan="1">Hamdi et al. (<xref>2025</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1"> <italic>Limosilactobacillus reuteri</italic> </td><td align="center" rowspan="1" colspan="1">Low‐acyl gellan gum (LAG) + High‐acyl gellan gum (HAG) + Ca<sup>2</sup> <sup>+</sup> (binary polysaccharide system)</td><td align="center" rowspan="1" colspan="1">Internal ionic gelation (emulsification with Span 80, Ca<sup>2</sup> <sup>+</sup> crosslinking, Box–Behnken optimization)</td><td align="center" rowspan="1" colspan="1">Encapsulation efficiency up to 95.5%; viability after encapsulation up to 97.43% (optimized conditions)</td><td align="center" rowspan="1" colspan="1">Stable viable counts; maintained ≥8.02 log CFU/mL after simulated GI exposure</td><td align="center" rowspan="1" colspan="1">Viability decreased from 9.50 to 8.46 log CFU/mL after SGF (2 h) and to 8.02 log CFU/mL after SIF (2 h); free cells dropped to undetectable levels</td><td align="center" rowspan="1" colspan="1">Enhanced protection against gastric acid and bile salts; suitable for functional food applications</td><td align="center" rowspan="1" colspan="1">González‐Cuello (<xref>2025</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1"> <italic>Limosilactobacillus reuteri</italic> DSM 17938</td><td align="center" rowspan="1" colspan="1">Alginate and chitosan–alginate matrices</td><td align="center" rowspan="1" colspan="1">Vibrating technology (extrusion‐based encapsulation); alginate beads with or without chitosan coating; CaCl<sub>2</sub> gelation; optional freeze‐drying</td><td align="center" rowspan="1" colspan="1">High encapsulation efficiency (∼97%); maintained viability after encapsulation and freeze‐drying (≈100% survival immediately after lyophilization); reduced log losses under thermal, osmotic, and oxidative stress compared to free cells</td><td align="center" rowspan="1" colspan="1">Improved stability during 28 days of storage at 4°C (≈1 log reduction vs. ≈3 log for free cells); better protection in Ringer and NaCl solutions</td><td align="center" rowspan="1" colspan="1">Significantly enhanced survival under simulated gastrointestinal conditions (gastric pH 2.5 with pepsin and intestinal bile salts); chitosan–alginate capsules showed superior protection against bile stress</td><td align="center" rowspan="1" colspan="1">Preserved functional activity, including reuterin production and antimicrobial activity; maintained probiotic functionality while improving technological robustness</td><td align="center" rowspan="1" colspan="1">De Prisco et al. (<xref>2014</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1"> <italic>Limosilactobacillus reuteri</italic> DSM 17938</td><td align="center" rowspan="1" colspan="1">Chia seed mucilage + sodium alginate</td><td align="center" rowspan="1" colspan="1">Electrohydrodynamic spraying (EHDA) and dripping mode; ionic crosslinking with Ca<sup>2</sup> <sup>+</sup>; nonthermal encapsulation</td><td align="center" rowspan="1" colspan="1">Viability ≈ 9.9 log CFU/mL after encapsulation and lyophilization; EE = 99.0 ± 0.01% (EHDA) and 99.0 ± 0.16% (DM)</td><td align="center" rowspan="1" colspan="1">High stability after freeze‐drying; maintained viability during powder formulation</td><td align="center" rowspan="1" colspan="1">Encapsulation matrix protected cells under simulated gastrointestinal conditions, improving survival compared to free cells</td><td align="center" rowspan="1" colspan="1">Successful incorporation into whey‐based functional powders; preserved probiotic viability and functional potential</td><td align="center" rowspan="1" colspan="1">Cid‐Córdoba et al. (<xref>2025</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1"> <italic>Limosilactobacillus reuteri</italic> </td><td align="center" rowspan="1" colspan="1">Alginate beads coated with chitosan (alginate–chitosan microcapsules)</td><td align="center" rowspan="1" colspan="1">Extrusion–gelation technique: cell–alginate suspension extruded into CaCl<sub>2</sub> solution to form beads, followed by chitosan coating via electrostatic interaction</td><td align="center" rowspan="1" colspan="1">Encapsulation efficiency > 90%; encapsulated cells showed significantly higher viability than free cells after encapsulation and thermal treatment</td><td align="center" rowspan="1" colspan="1">Improved stability during storage at 4°C; encapsulated cells showed lower viability loss compared to free cells over storage period</td><td align="center" rowspan="1" colspan="1">Encapsulated cells exhibited significantly higher survival under simulated gastrointestinal conditions (pH 2.0 with pepsin and bile salts) compared to free cells</td><td align="center" rowspan="1" colspan="1">Alginate–chitosan coating provided an effective physical barrier against acid and bile, enhancing probiotic survival and suitability for functional food applications</td><td align="center" rowspan="1" colspan="1">Chitprasert et al. (<xref>2012</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1"> <italic>Limosilactobacillus reuteri</italic> DSM 17938</td><td align="center" rowspan="1" colspan="1">Chia seed mucilage + sodium alginate (binary polysaccharide hydrogel matrix)</td><td align="center" rowspan="1" colspan="1">Electrohydrodynamic spraying (EHDA); ionic crosslinking with CaCl<sub>2</sub>; nonthermal encapsulation followed by freeze‐drying</td><td align="center" rowspan="1" colspan="1">High postprocessing viability; encapsulation efficiency ≈ 98%–99%; viable counts remained > 9 log CFU/mL after encapsulation and lyophilization</td><td align="center" rowspan="1" colspan="1">Good storage stability of dried powders; encapsulated cells maintained viability during storage compared to free cells</td><td align="center" rowspan="1" colspan="1">Encapsulated cells showed significantly higher survival under simulated gastrointestinal conditions (gastric and intestinal phases) than free cells</td><td align="center" rowspan="1" colspan="1">Hydrogel microstructure provided effective protection against acidic and bile environments, enabling application in functional food systems</td><td align="center" rowspan="1" colspan="1">Cerón‐Córdoba et al. (<xref>2025</xref>)</td></tr></tbody></table>This integrative comparison in EE (Table 1) highlights the relationships between structure and function, showing how capsule architecture, polymer composition (wall material), and processing strategy can have a direct influence on L. reuteri viability, EE release behavior, and functional performance across different studies.
Current research on L. reuteri underscores the need to integrate strain physiology, wall material properties, technological outcomes, and functional performance. This review examines encapsulation studies involving L. reuteri, focusing on how strain‐dependent physiological characteristics affect EE, survival during processing and storage, and release under gastrointestinal conditions. It synthesizes evidence from various strains, materials, and encapsulation methods to identify new relationships across different food structures and functions (Figure 1).
Limosilactobacillus reuteri strains in food matrices.
This framework links encapsulation design variables, such as the chemical composition of the coating material, encapsulation size, mechanical properties and process conditions, with strain‐dependent physiological characteristics, including EPS production and tolerance to gastric and intestinal acidity. This relationship clarifies outcome metrics, including the viability of encapsulated microorganisms and their stability during processing, storage and exposure to gastrointestinal stress. Figure 2 illustrates a structure–function approach for interpreting the selection of coating materials, encapsulation methods and processing conditions to maintain performance and functionality in food matrices.
Summarizes this multilayer conceptual framework, illustrating how strain‐dependent physiological traits, encapsulation material properties, and processing strategies interact to determine the functional performance of encapsulated Limosilactobacillus reuteri in food matrices and gastrointestinal environments.
Methodology of the Systematic Analysis
2
For this review, a systematic analysis of the scientific literature was conducted to examine both the functional activity and encapsulation of various L. reuteri strains. The primary objective was to identify the most extensively studied strains, prevalent encapsulation strategies, and the biological and technological mechanisms underlying their applications in food systems. This methodology facilitates the identification of knowledge gaps and technological challenges related to L. reuteri viability during processing, storage, and gastrointestinal transit.
Information Sources and Search Strategy
2.1
Relevant studies were identified through a structured search of three major scientific databases: Scopus, web of Science, and PubMed. The search utilized keywords included L. reuteri, encapsulation, probiotic viability, functional properties, anti‐inflammatory activity, spray drying, electrohydrodynamic spraying, and ionic gelation. Combinations of the search items were also employed. The database search was conducted in July 2025.
Selection and Synthesis
2.2
The initial database search identified approximately 180–220 records. Following title and abstract screening, nonrelevant publications and duplicate entries were excluded, resulting in 60–80 full‐text articles assessed for eligibility. Applications of predefined inclusion and exclusion criteria, which focused on strain‐specific data, food‐grade encapsulation materials, and quantitative performance metrics, yielded a final set of 30–40 studies for quantitative and integrative analysis (Figure 2).
The selected studies were critically analyzed and thematically organized according to three main criteria: (1) L. reuteri strain and associated functional characteristics, (2) encapsulation material architecture and processing technique, and (3) targeted technological or functional outcomes, such as gastrointestinal survival, storage stability, or antimicrobial activity. This thematic classification facilitated cross‐study comparisons and supported the identification of structure–function relationships across different encapsulation systems.
The selection process prioritized the most recent studies (2010–2025), also highlighting fundamental contributions and comparative data between strains and materials. This time frame was chosen because it encompasses the consolidation of probiotic encapsulation research, including the taxonomic reclassification of L. reuteri in 2020.
Limosilactobacillus reuteri (LR)
3
L. reuteri is one of the host‐associated bacteria found in different parts of the human body, including the gastrointestinal and urinary tracts, skin, and breast milk (Mu et al. 2018a). It has the ability to secrete bacteriocins, reuterin, and reutericyclin, which contribute to the control of gastrointestinal infections (Paternina et al. 2013). L. reuteri exhibits a wide range of health‐promoting effects, for example, hypocholesterolemic, antimutagenic, antimicrobial, and anti‐inflammatory activities. It also contributes to improved immune response and lactose digestibility, reduces colonization by Helicobacter pylori, and modulates anthropometric indices such as body weight, hip circumference, waist–hip ratio, fat mass, and muscle mass. Additionally, it shows potential for bisphenol degradation, displays moderate hemolytic activity, and helps lowering total cholesterol and triglyceride levels (Figure 2). Likewise, it has inhibitory effects on cytokines secretion, presenting beneficial effects on periodontal diseases (Alok et al. 2017). However, the effectiveness of probiotic microorganisms depends on the number of viable cells that might successfully colonize the intestine, and this viability depends of several factors like gastric acidity, digestive enzymes, and bile salts, all of them with negatively impact under probiotics stability (Juárez‐Tomás et al. 2015). Therefore, during the design of beneficial bacteria‐containing products, ensuring microbial stability throughout production processes and the product shelf life becomes a key technological requirement (Algaithi et al. 2022).
Functional Properties of Limosilactobacillus reuteri and the Role of Encapsulation Technologies
3.1
The health benefits of L. reuteri are well documented in the literature. However, the effectiveness of its antimicrobial, anti‐inflammatory, antioxidant, and antiobesity properties in food applications depends on bacterial survival during processing, storage, and gastrointestinal transit. The following sections outline the main functional activities of L. reuteri and examine its relationship with release systems and encapsulation.
Functional Activity of Limosilactobacillus reuteri in Food Systems
3.1.1
L. reuteri has been incorporated into various food matrices due to its probiotic benefits, including balance of the gastrointestinal microbiota and metabolic regulation (Mu et al. 2018b). Studies have shown that encapsulating L. reuteri in fruit‐based beverages, such as mango pulp with maltodextrin and inulin, results in high survival rates (≈99%) and full cell viability during storage (Gámez et al. 2021). While antioxidant capacity was not directly evaluated, these findings highlight the importance of encapsulation strategies for maintaining probiotic viability in phenolic‐rich foods. Similarly, L. reuteri ATCC 55730 maintained high survival (≈98%) in a bean and oat‐based spread after 28 days at 4°C, supporting its use in plant‐based products (Barboza et al. 2012). Additionally, reuterin produced by L. reuteri has been shown to inhibit Listeria monocytogenes in cold‐smoked salmon (Montiel et al. 2014).
These studies reflect the potential of L. reuteri in food applications. However, its activity is sensitive to stress factors such as oxygen exposure, dehydration, and acidity (Baral et al. 2021). Encapsulation is therefore recommended to maintain the viability and functionality of L. reuteri during processing, storage, and gastrointestinal transit (Teymoori et al. 2024).
Antimicrobial Activity
3.1.2
The antimicrobial activity of LR is primary attributed to the production of organic acids, bacteriocins, and strain‐specific metabolites, particularly reuterin (Banakar et al. 2023; Niamah et al. 2023). Reuterin exhibits broad‐spectrum antimicrobial activity against bacteria, yeasts, and fungi, making it especially relevant for food preservation (Jalali et al. 2024). The production and stability of reuterin are strongly influenced by environmental factors such as oxygen availability and pH. To address these limitations, several encapsulation strategies have been developed to protect both viable cells and antimicrobial metabolites. Juárez Tomás et al. (2015) demonstrated that encapsulating reuterin‐producing L. reuteri DSM 20016 in alginate‐based matrices effectively preserved antimicrobial activity by safeguarding the trapped cells and metabolites.
Jalali et al. (2024) further demonstrated that postbiotics derived from L. reuteri significantly inhibited the growth of Escherichia coli and Staphylococcus aureus on the surface of red meats. Collectively, these studies highlight the importance of encapsulation design in maintaining antimicrobial functionality and support the use of L. reuteri as a natural preservative in food systems. Nevertheless, additional research is still required to demonstrate and confirm its efficacy within food matrices and under processing conditions.
Anti‐Inflammatory Activity
3.1.3
Different LR strains show anti‐inflammatory activity by modulating the immune response, such as downregulation of proinflammatory cytokines and stimulating anti‐inflammatory mediators (Werlinger et al. 2022). These effects are fundamental for maintaining the integrity of the intestinal barrier and immune homeostasis (Alam et al. 2022).
Most evidence for these anti‐inflammatory mechanisms comes from in vivo and in vitro studies using with nonencapsulated cells. While encouraging, practical application in functional foods depends on L. reuteri’s ability to survive gastrointestinal transit and reach the intestinal mucosa in sufficient quantities to exert immunomodulatory effects (Domingo and José 2017).
Therefore, encapsulation systems designed to protect cells against gastric acidity and bile salts are essential to preserve the anti‐inflammatory functionality of L. reuteri when delivered through food matrices.
Antioxidant Activity
3.1.4
Oxidative stress arises from the accumulation of reactive oxygen and nitrogen species, and other free radicals, which damage cellular components through mechanisms such as DNA hydroxylation, lipid peroxidation, and protein denaturation. These processes contribute to the development of chronic degenerative diseases (Allameh et al. 2023; Reddy 2023). Antioxidants, including phenolic compounds, peptides, and probiotics have been extensively studied for their ability to mitigate oxidative damage and reduce disease incidence (J.‐Z. Wang et al. 2017).
Several Lactobacillus species, including L. reuteri, have demonstrated antioxidant effects through the production of bioactive metabolites and enzymatic defense mechanisms. Jalali et al. (2024) assessed the antioxidant activity of postbiotics derived from L. reuteri using the DPPH method and observed significantly higher radical scavenging activity postbiotics‐containing samples compared to controls. This effect was associated with elevated phenolic content, suggesting that metabolites produced by LR contribute to enhanced free radical neutralization. While comparative analyses indicated higher antioxidant activity for postbiotics from L. rhamnosus, the findings confirmed the antioxidant potential of L. reuteri‐derived postbiotics.
Encapsulation strategies have been explored to preserve probiotic viability and antioxidant functionality. Rodklongtan and Chitprasert (2017) investigated the coencapsulation of L. reuteri KUB‐AC5 with ascorbic acid during spray drying, using ascorbic acid as a complementary antioxidant to enhance cellular protection. Their findings showed high retention of ascorbic acid after spray drying and during refrigerated storage, with stable antioxidant activity evaluated by the ABTS method. However, storage at elevated temperatures decreased antioxidant capacity, underscoring the importance of proper storage. These findings demonstrate how encapsulation systems can help maintaining antioxidant functionality by protecting both probiotic cells and antioxidant compounds.
Additional studies have shown that fermentation with L. reuteri can increase antioxidant activity in food matrices by releasing phenolic compounds and bioactive peptides. Tyagi et al. (2021) reported that fermented brown rice had higher antioxidant activity attributed to the release of insoluble phenolic compounds during fermentation. Similarly, Begunova et al. (2021) showed that milk fermentation with LR produces antioxidant peptides through proteolytic activity. However, these studies mainly examined biological activity under controlled conditions and did not assess the stability of antioxidant functionality during food processing or storage.
Although the substantial evidence supporting the antioxidant potential of L. reuteri and its metabolites, few studies have examined how encapsulation contribute to preserving this functionality under food processing and storage.
Antiobesity Potential and Technological Implications
3.1.5
Obesity is a multifactorial metabolic disorder that is strongly associated with gut microbiota dysbiosis, chronic inflammation, and disruptions in glucose and lipid metabolism (Ahmed and Konje 2023). Several studies have shown that different strains of L. reuteri produce antiobesity effects in animal models, such as reductions in body weight gain, adiposity, insulin resistance, and inflammatory markers under high‐fat or high‐fructose dietary conditions (Hsieh et al. 2013; Larsen et al. 2023; Tang et al. 2025).
These beneficial effects have been associated with the modulation of adipogenic and lipogenic gene expression, enhance glucose tolerance, reduction of systemic inflammation, and protection against nonalcoholic fatty liver disease (Di Porzio et al. 2023; Kim et al. 2022; Wu et al. 2025). Notably, most of these studies have been conducted using nonencapsulated cells administered in controlled experimental conditions.
From a technological perspective, the translation of the reported antiobesity effects of L. reuteri into functional food applications remains limited by challenges related to probiotic survival, stability, and targeted intestinal delivery. Encapsulation strategies are therefore critical for maintaining cell viability and ensuring consistent delivery of L. reuteri to the intestine, which is essential to achieve its metabolic benefits in real food systems.
To facilitate comparison between studies, Table 2 compiles the literature analyzed in the section Functional Properties of L. reuteri and the Role of Encapsulation Technologies, indicating whether the functional activity was experimentally evaluated and the presence of encapsulation strategies in food matrices.
TABLE 2: Summary of studies evaluating functional activity and encapsulation strategies of Limosilactobacillus reuteri in food‐related systems.
<table><col align="left"/><col align="center"/><col align="center"/><col align="center"/><col align="center"/><col align="center"/><thead><tr><th align="left" rowspan="1" colspan="1"> <italic>L. reuteri</italic> strain</th><th align="center" rowspan="1" colspan="1">Food matrix/system</th><th align="center" rowspan="1" colspan="1">Encapsulation strategy</th><th align="center" rowspan="1" colspan="1">Functional activity or experimentally evaluated</th><th align="center" rowspan="1" colspan="1">Outcome on functional activity</th><th align="center" rowspan="1" colspan="1">References</th></tr></thead><tbody><tr><td align="left" rowspan="1" colspan="1"> <italic>L. reuteri</italic> ATCC 55730</td><td align="center" rowspan="1" colspan="1">Legume‐based spread</td><td align="center" rowspan="1" colspan="1">Not applied</td><td align="center" rowspan="1" colspan="1">Yes</td><td align="center" rowspan="1" colspan="1">High probiotic survival during refrigerated storage, supporting functional performance in plant‐based matrices</td><td align="center" rowspan="1" colspan="1">Barboza Martínez et al. (<xref>2012</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1"> <italic>L. reuteri</italic> (not specified)</td><td align="center" rowspan="1" colspan="1">Cold‐smoked salmon</td><td align="center" rowspan="1" colspan="1">Not applied</td><td align="center" rowspan="1" colspan="1">Yes (antimicrobial)</td><td align="center" rowspan="1" colspan="1">Reuterin production effectively inhibited Listeria monocytogenes</td><td align="center" rowspan="1" colspan="1">Montiel et al. (<xref>2014</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1"> <italic>L. reuteri</italic> DSM 20016</td><td align="center" rowspan="1" colspan="1">Model system</td><td align="center" rowspan="1" colspan="1">Alginate‐based encapsulation</td><td align="center" rowspan="1" colspan="1">Yes (antimicrobial)</td><td align="center" rowspan="1" colspan="1">Encapsulation preserved antimicrobial activity by protecting viable cells and entrapped metabolites</td><td align="center" rowspan="1" colspan="1">Juárez‐Tomás et al. (<xref>2015</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1"> <italic>L. reuteri</italic> KUB‐AC5</td><td align="center" rowspan="1" colspan="1">Powdered food model</td><td align="center" rowspan="1" colspan="1">Spray drying with lactose and ascorbic acid</td><td align="center" rowspan="1" colspan="1">Yes (antioxidant)</td><td align="center" rowspan="1" colspan="1">Coencapsulation preserved antioxidant activity and improved stability during storage</td><td align="center" rowspan="1" colspan="1">Rodklongtan and Chitprasert (<xref>2017</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1"> <italic>L. reuteri</italic> ATCC 53608</td><td align="center" rowspan="1" colspan="1">Mango pulp‐based beverage</td><td align="center" rowspan="1" colspan="1">Spray drying with maltodextrin and inulin</td><td align="center" rowspan="1" colspan="1">No</td><td align="center" rowspan="1" colspan="1">Encapsulation improved probiotic stability, supporting preservation of functional potential</td><td align="center" rowspan="1" colspan="1">Gámez et al. (<xref>2021</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1"> <italic>L. reuteri</italic> (not specified)</td><td align="center" rowspan="1" colspan="1">Meat‐based system</td><td align="center" rowspan="1" colspan="1">Not applied</td><td align="center" rowspan="1" colspan="1">Yes (antioxidant)</td><td align="center" rowspan="1" colspan="1">Postbiotic application enhanced antioxidant capacity of the meat matrix</td><td align="center" rowspan="1" colspan="1">Jalali et al. (<xref>2024</xref>)</td></tr></tbody></table>Strain‐Specific Differences
3.2
Several probiotic strain varieties have been studied in the pharmaceutical and food industries due to their high functional and therapeutic potential, demonstrating substantial genomic diversity and adaptability to different ecological niches Some strain‐dependent characteristics are summarized in Table 3, which compiles the most frequently studied L. reuteri isolates and their functional properties. Among the most extensively studied strain, particularly in pediatric populations, the strains DSM‐17938 have received considerable attention. Several reports demonstrated its effective gastrointestinal tract colonization producing a reduction in diarrhea events related to rotavirus infection (Sun et al. 2023).
TABLE 3: Limosilactobacillus reuteri strains with functional capabilities.
<table><col align="left"/><col align="center"/><col align="center"/><thead><tr><th align="left" rowspan="1" colspan="1"> <italic>Lactobacillus Reuteri</italic> strain</th><th align="center" rowspan="1" colspan="1">Functional activity</th><th align="center" rowspan="1" colspan="1">References</th></tr></thead><tbody><tr><td align="left" rowspan="1" colspan="1">FCQHC8L6 y FYNDL13</td><td align="center" rowspan="1" colspan="1">Anti‐inflammatory and Antimicrobial</td><td align="center" rowspan="1" colspan="1">Lin et al. (<xref>2023</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">BR120</td><td align="center" rowspan="1" colspan="1">Probiotic, anti‐inflammatories, radical scavengers, acid tolerance, bile tolerance and intestinal adhesion</td><td align="center" rowspan="1" colspan="1">Ju et al. (<xref>2023</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">ATCC 53608</td><td align="center" rowspan="1" colspan="1">Antimicrobial activity against viruses, fungi, protozoa and gram‐positive and gram‐negative bacteria</td><td align="center" rowspan="1" colspan="1">Ma et al. (<xref>2023</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">ATCC PTA5289</td><td align="center" rowspan="1" colspan="1">Inhibits the acid tolerance response in oral bacteria</td><td align="center" rowspan="1" colspan="1">Boisen et al. (<xref>2023</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">DSM 17938</td><td align="center" rowspan="1" colspan="1">Colonize the gastrointestinal tract effectively and shorten the duration of watery diarrhea associated with rotavirus infection time significantly</td><td align="center" rowspan="1" colspan="1">Sun et al. (<xref>2023</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">NCIMB 30242</td><td align="center" rowspan="1" colspan="1">Reduced total cholesterol (TC)</td><td align="center" rowspan="1" colspan="1">Liu et al. (<xref>2023</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">DSM 20016 y DSM 17938</td><td align="center" rowspan="1" colspan="1">Antimicrobial characteristics, such as antifungal activity and a broad‐spectrum of activity against Gram‐positive and negative bacteria</td><td align="center" rowspan="1" colspan="1">Rodrigues et al. (<xref>2023</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">DSM17648</td><td align="center" rowspan="1" colspan="1">Reduction in H. pylori colonization</td><td align="center" rowspan="1" colspan="1"/></tr><tr><td align="left" rowspan="1" colspan="1">DSM 17938</td><td align="center" rowspan="1" colspan="1">Antimicrobial activity</td><td align="center" rowspan="1" colspan="1">Zhang et al. (<xref>2022</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">ATCC 53608</td><td align="center" rowspan="1" colspan="1">Antimicrobial activity</td><td align="center" rowspan="1" colspan="1">Soltani Lak et al. (<xref>2021</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">DSM 17648</td><td align="center" rowspan="1" colspan="1">Reduction in H. pylori colonization</td><td align="center" rowspan="1" colspan="1">Yang et al. (<xref>2021</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">DSM 17648</td><td align="center" rowspan="1" colspan="1">Reduction in H. pylori colonization</td><td align="center" rowspan="1" colspan="1">Dargenio et al. (<xref>2021b</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">ATCC 23272</td><td align="center" rowspan="1" colspan="1">Probiotic</td><td align="center" rowspan="1" colspan="1">Karimi et al. (<xref>2021</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">KUB‐AC5</td><td align="center" rowspan="1" colspan="1">Probiotic</td><td align="center" rowspan="1" colspan="1">Diệp Huy Vũ et al. (<xref>2021a</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">B2</td><td align="center" rowspan="1" colspan="1">Antimicrobial activity and Acid resistance and Bile Salt Tolerance</td><td align="center" rowspan="1" colspan="1">Popović et al. (<xref>2021</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">B‐14171.</td><td align="center" rowspan="1" colspan="1">Inhibit the growth of pathogenic bacteria and improve the immune system of the host.</td><td align="center" rowspan="1" colspan="1">Mis Solval et al. (<xref>2019</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">E81</td><td align="center" rowspan="1" colspan="1">Effect against H. pylori infection</td><td align="center" rowspan="1" colspan="1">Ceylan et al. (<xref>2019</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">ATCC 55730</td><td align="center" rowspan="1" colspan="1">Degradation potential of bisphenol</td><td align="center" rowspan="1" colspan="1">Ju et al. (<xref>2019</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">B‐14171</td><td align="center" rowspan="1" colspan="1"> Antimicrobial</td><td align="center" rowspan="1" colspan="1">Mu et al. (<xref>2018</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">ATCC 55730 y ATCC PTA 5282</td><td align="center" rowspan="1" colspan="1">Antimicrobial and anti‐inflammatory properties</td><td align="center" rowspan="1" colspan="1">Kaur et al. (<xref>2018</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">ATCC 55730</td><td align="center" rowspan="1" colspan="1">Treatment of diarrhea, hernia and intestinal backflow</td><td align="center" rowspan="1" colspan="1">Indrio et al. (<xref>2011</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">DSM 20016T; ATCC 23272</td><td align="center" rowspan="1" colspan="1">Biotherapeutic potential</td><td align="center" rowspan="1" colspan="1">Juárez Tomás et al. (<xref>2015</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">DSM17648</td><td align="center" rowspan="1" colspan="1">Reduction in H. pylori colonization</td><td align="center" rowspan="1" colspan="1">Holz et al. (<xref>2015</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">ATCC PTA 6475 y ATCC PTA 5289</td><td align="center" rowspan="1" colspan="1">Antimicrobial activity</td><td align="center" rowspan="1" colspan="1">Jones and Versalovic (<xref>2009</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">NCIMB 30242</td><td align="center" rowspan="1" colspan="1">Reduced total cholesterol (TC)</td><td align="center" rowspan="1" colspan="1">Casas and Dobrogosz (<xref>2000</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">CRL 1098</td><td align="center" rowspan="1" colspan="1">Decreased total cholesterol and triglycerides</td><td align="center" rowspan="1" colspan="1">Taranto et al. (<xref>1998</xref>)</td></tr></tbody></table>Clinical studies in orthodontic patients showed that tablets combining L. reuteri DSM17938 and PTA5289 reduced the growth of Streptococcus mutans, improving indicators such as plaque, gingivitis, and gingival bleeding, although without reaching statistical significance. In another approach, this strain (DSM17938 and PTA5289) has shown to interfere with the adhesion of S. mutans to hydroxyapatite‐coated surfaces demonstrating anticaryotic potential, whereas ATCC 55730 displayed antimicrobial and anti‐inflammatory properties (Kaur et al. 2018). Additional strains, such as ATCC 53608, are widely used as they are considered broad‐spectrum against various pathogenic microorganisms including viruses, fungi, protozoa, and bacteria (Ma et al. 2023; Soltani Lak et al. 2021). On the other hand, CRL 1098 has exhibited a decrease in triglycerides and cholesterol levels, thus improving metabolic health. Similar results were obtained with the NCIMB 30242 strain, which has shown total cholesterol levels reduction in clinical studies (J. Liu et al. 2023).
KUB‐AC5 stands out for its application in food matrices as result of its excellent gastrointestinal colonization ability and because its viability is preserved after its spray drying which courage its probiotic applications at industrial level (Diệp Huy Vũ et al. 2021). Strains such as BR120, FCQHRCL6, and FYNDL13 have been identified in internal datasets or preliminary investigations for possessing multifunctional properties including probiotic activity, anti‐inflammatory effects, radical scavenging capacity, tolerance to gastric acidity, and adhesion to intestinal epithelial cells. Among the most relevant strains with health benefits can be find the DSM 17648, which can reduce Helicobacter pylori colonization improving digestive wellness (Dargenio et al. 2021; Yang et al. 2021). Other strains, such as B‐14171 and E81, also exhibit antimicrobial capacity and contribute to the modulation of the immune response, highlighting the breadth of mechanisms by which L. reuteri strains exert their functional roles (Ceylan et al. 2019; Mis Solval et al. 2019).
CRL1324 is a microbial strain designed to enhance vaginal health. It helps maintaining viability in vaginal fluid and inhibiting the proliferation of Streptococcus agalactiae, which can induce the release of cytokines causing inflammation in vaginal tissue and the subsequent decreasing of protective microbiota (Juárez Tomás et al. 2015). In the same study, it was demonstrated that the probiotic potential of L. reuteri differs depending on the strain used.
In brief, the probiotic potential of L. reuteri is not uniform and rather is highly strain‐dependent, with each isolate exhibiting specific applications. Although strain diversity is desirable because this expands the applicability of these bacteria, at the same time it faces a challenge. Mainly because every microbial strain presents different stability and viability under certain conditions making their study or applications into foods or supplements difficult. To overcome these challenges, encapsulation has emerged as strategy to improve preservation of the strain functional activities and to control its release into the diana host.
Encapsulation of Limosilactobacillus reuteri
3.3
Different factors can affect the stability of probiotics in food products including food properties, processing parameters, and microbial characteristics (Algaithi et al. 2022). For probiotic microorganisms to reach the intestinal mucosa, stomach acidity is an almost insurmountable barrier for microorganisms ingested in drinks and foods, thus protecting the intestine against infections. Consequently, the number of viable microorganisms that survive this barrier is often too low to exert a significant beneficial effect.
To address these stability challenges and provide additional protection against environmental stress in the gastrointestinal tract, microencapsulation techniques are considered among the most effective strategies for enhancing probiotic survival during processing and storage, thereby ensuring their activity in the intestine (J. Wang et al. 2017). To overcome this obstacle, encapsulation can be defined as a process in which liquid, solid, or gaseous particles referred to core, active substance, internal phase, or payload are enclosed within a porous polymeric film (Araneda and Valenzuela 2009), forming a capsule, wall material, membrane, carrier, or shell (Gibbs et al. 1999). It can also be regarded as a unique method of packaging, isolating, and storing materials at the micro or nanoscale for subsequent controlled release or to prevent unwanted reactions between components (Astray et al. 2009).
As shown in Figure 3, capsules can also be classified according to structural criteria, such as the core coating (monolayer or multilayer), texture (soft or rigid), morphology (smooth or rough), and core presentation (defined or undefined, with matrix, mixed, or agglomerated). Size is also a determining factor, ranging from macrocapsules (> 100 µm) to microcapsules (10–100 µm) and nanocapsules (1–100 nm). Among these, matrix‐type capsules are the most widely used in food and pharmaceutical systems, as they allow for smaller diameters and a homogeneous distribution of the active compound (Sosnik 2014; J. Wang et al. 2017). This framework supports the interpretation of the diverse encapsulation methods used for L. reuteri, which often result in matrix‐based systems when employing alginate or its blends.
Different classifications of capsules.
This technique allows individual coating of materials to protect them from harmful external factors, creating a microenvironment capable of regulating interactions between the core and its surroundings (Borgogna et al. 2010). The capsule wall can provide protection against heat, light, moisture, oxygen, and pH variations, as well as improve processing resistance, packaging stability, flavor, aroma, nutritional value, and appearance of the final product (Yañez Fernández et al. 2002). Current research continues focusing on developing various macro‐, micro‐, and nanoencapsulation techniques that allow the delivery of probiotics in adequate quantities to confer health benefits, giving place to multifunctional foods (Liu et al. 2016).
The different encapsulation techniques for L. reuteri are divided into physicochemical and physical methods, each with its own advantages and limitations. Chemical methods include interfacial polymerization or molecular inclusion, although the use of nonbiodegradable materials limits their application in foods. Physicochemical techniques, including coacervation and liposome entrapment, provide tailored release and protection of sensitive compounds, although their stability and cost remain challenging.
Physical methods are the most widely applied in probiotics because of their scalability and food‐grade status. Spray drying is the most widely used technique due to its low cost and industrial viability. However, its limitation is caused by thermal stress, leading to a reduction in cell viability unless protective carriers are added. Lyophilization, on the other hand, guarantees high survival rates and long‐term stability, but requires more time and resources. While extrusion with ionotropic gelation, when using alginate or gums, effectively protects L. reuteri from gastrointestinal stress, the capsule's size and mechanical weakness can limit its consumer acceptance. More recently, electrohydrodynamic spraying (electrospraying) has gained attention for producing micro‐ and nanocapsules with controlled morphology and high EE, although scaling the process remains difficult.
Taken together, the method selected depends on striking a balance between probiotic survival, release behavior, compatibility of coating materials, and feasibility for industrial application.
In addition to quantitative outcomes summarized in Table 1, it is relevant to examine the mechanistic basis by which encapsulation materials influence stress tolerance and functional performance of L. reuteri.
In addition to enhancing survival rates, encapsulation materials affect the functional performance of L. reuteri through multiple interconnected mechanisms. Dense or multilayer polymeric matrices, such as alginate‐ chitosan systems or protein polysaccharide coacervated, reduce membranes damage during gastrointestinal transit (Anal and Singh 2007; Cook et al. 2012; de Prisco and Mauriello 2016). Furthermore, polymers with buffering capacity or antioxidant properties can stabilize the local microenvironment, thereby supporting membrane integrity and enzymatic activity. Although encapsulation does not directly enhance adhesion to intestinal epithelium, it preserves related functions after release. Sustained metabolic activity, including the production of antimicrobial metabolites such as reuterin and other postbiotic compounds, may be directly supported by encapsulation strategies that limit oxidative stress and dehydration‐induce cellular damage (Juárez‐Tomás et al. 2015; Rodrigues et al. 2023).
Taken together, the encapsulation methods for L. reuteri vary widely in terms of cost, scalability, and protective capacity. Spray drying and ionic gelation remain the most widely adopted, while emerging methods such as electrospraying provide new opportunities but are still far from industrial application.
Regardless of laboratory‐scale results, global consumption of probiotic‐containing foods has increased substantially in recent years. Market analysts project that the global probiotics sector will continue to expand toward 2030, reflecting growing demand for functional foods and dietary supplements (Grand View Research 2024). This sustained growth underscores the need for formulation strategies that enable industrial‐scale implementation. In this context, advances in encapsulation technologies are critical to ensuring stability, functional performance, and regulatory compliance of commercially viable food products containing L. reuteri strains.
Industrial and Scaling Considerations for Encapsulated Limosilactobacillus reuteri
3.3.1
Although numerous encapsulation strategies have improved viability and function of LR under controlled laboratory settings, their successful application in real‐world food systems depends on maintaining viability and stability under industrial processing conditions. Despite promising results reported at the laboratory scale, the industrial implementation of encapsulated L. reuteri remain challenging, with coating materials as one of the most critical factors. Encapsulation matrices must be food grade, GRAS‐certified, cost‐effective, and compatible with the physicochemical and sensory properties of the food matrix in which they will be incorporated. Polymers such as alginate, certain starch derivatives, proteins, gums, waxes, and some prebiotics meet these requirements and are suitable for large‐scale applications.
On the other hand, maintaining the stability of both the coating material and the microorganism during processing is a significant challenge. Food processing methods, such as spray drying, mixing, pumping, extrusion, and heat treatments, expose probiotic microorganisms to stresses like shear, dehydration, temperature changes, and oxygen, which can compromise their viability. While encapsulation techniques like electrospraying and nanoencapsulation offer strong protection at small scales, their scalability, performance, and cost are limiting. Spray drying and ionic gelation remain to dominate industrial applications due to their robustness and economic effectiveness, but they require careful optimization to preserve microorganism viability and stability.
Based on the above, future industrial implementation should prioritize encapsulation systems that balance EE, stability, and feasibility with production process scalability, regulatory acceptance, and stability in real‐world food matrices. Bridging the gap between laboratory results and industrial viability is essential for the successful commercialization of encapsulated L. reuteri in functional foods.
In recent years, studies on the encapsulation of L. reuteri have shown a steady upward trend (Figure 4). Since the first report in 2004, the field has progressed slowly, with only one or two publications per year until 2015. Subsequently, a gradual increase was observed and by 2020, the number of contributions had increased dramatically, reaching an average of 10 per year for the past 5 years, according to the Scopus database. This increment reflects the growing scientific interest in L. reuteri as a probiotic model, as well as the diversification of encapsulation methods, which include electrospray, ionic gelation, and other techniques using different biopolymers. Notably, the sharp increase after 2020 could also reflect advances in biopolymer science and the urgent demand for stable probiotics. The increase in work related to probiotics encapsulation is the results of demonstration that the encapsulation traps bacteria in a coating matrix, preventing their exposure to harmful environmental conditions and improving their viability. The steady growth of publications also shows that L. reuteri is no longer studied in isolation but rather as a reference strain to test emerging encapsulation technologies. This perspective helps explain its recurrent presence in formulations aimed at improving probiotic stability and functionality.
Articles related to the encapsulation of Lactobacillus reuteri (Scopus database; keywords: “encapsulation” and “Lactobacillus reuteri”; year‐individual: 2004–2025) and limiting it to articles only.
As shown in Table 4, the most commonly used coating materials for L. reuteri encapsulation include biopolymer blends such as sodium alginate, calcium alginate, maltodextrin, and others, mainly applied through ionic gelation techniques. Sodium and calcium alginate predominate due to their low cost, biocompatibility, and GRAS status. However, their combination with proteins (e.g., whey protein or hemp seed protein) or polysaccharides (e.g., inulin or plant mucilage) generally results in higher EE. Accordingly, the physicochemical compatibility between alginate and other complementary biopolymers improves the survival of L. reuteri under gastrointestinal stress, as well as its textural and sensory properties when incorporated into food matrices.
TABLE 4: Coating materials used in the encapsulation of Lactobacillus reuteri.
<table><col align="left"/><col align="center"/><col align="center"/><thead><tr><th align="left" rowspan="1" colspan="1">Core materials</th><th align="center" rowspan="1" colspan="1">Encapsulation efficiencies (EE)</th><th align="center" rowspan="1" colspan="1">References</th></tr></thead><tbody><tr><td align="left" rowspan="1" colspan="1">Protein isolates</td><td align="center" rowspan="1" colspan="1">93.06</td><td align="center" rowspan="1" colspan="1">Lau et al. (<xref>2025</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Beads of alginate hydrogel embedded with gellan fluid gel</td><td align="center" rowspan="1" colspan="1">86.64</td><td align="center" rowspan="1" colspan="1">Najafpour et al. (<xref>2025</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Whey proteins and gum Arabic</td><td align="center" rowspan="1" colspan="1">82.11</td><td align="center" rowspan="1" colspan="1">Atta et al. (<xref>2025</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Mucin and hempseed protein isolate</td><td align="center" rowspan="1" colspan="1">Not reported</td><td align="center" rowspan="1" colspan="1">Lau and Quek (<xref>2025</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Hydrogel formed by hyaluronic acid modified with thiol groups of L‐cysteine</td><td align="center" rowspan="1" colspan="1">38.15</td><td align="center" rowspan="1" colspan="1">Yang et al. (<xref>2025</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Polysaccharide‐based bionanocomposite. Inulin, polydextrose, and pectin were utilized as prebiotics, and magnesium oxide nanoparticles as reinforcing agent in the microgel structure</td><td align="center" rowspan="1" colspan="1">99.37</td><td align="center" rowspan="1" colspan="1">Mohamadzadeh et al. (<xref>2025</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Inulin and maltodextrin</td><td align="center" rowspan="1" colspan="1">43.27</td><td align="center" rowspan="1" colspan="1">Ceron‐Cordoba et al. (<xref>2025</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Whey protein isolate or sodium caseinate (Cas), followed by pectin</td><td align="center" rowspan="1" colspan="1">92</td><td align="center" rowspan="1" colspan="1">Rodríguez et al. (<xref>2025</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Chitosan and Ca‐alginate beads</td><td align="center" rowspan="1" colspan="1">92.5</td><td align="center" rowspan="1" colspan="1">Jeznienė et al. (<xref>2024</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Whey protein concentrate and gum arabic</td><td align="center" rowspan="1" colspan="1">67</td><td align="center" rowspan="1" colspan="1">Teymoori et al. (<xref>2024</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Sodium alginate and tomato seed mucilage</td><td align="center" rowspan="1" colspan="1">91.54</td><td align="center" rowspan="1" colspan="1">Ganje et al. (<xref>2024</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Alginate or pectin followed by polymeric coating (with whey protein concentrate or chitosan)</td><td align="center" rowspan="1" colspan="1">91</td><td align="center" rowspan="1" colspan="1">Mosquera‐Vivas et al. (<xref>2024</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Alginate‐inulin</td><td align="center" rowspan="1" colspan="1">Not reported</td><td align="center" rowspan="1" colspan="1">Yadav et al. (<xref>2024</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Rice protein, pea protein and inulin</td><td align="center" rowspan="1" colspan="1">96.99</td><td align="center" rowspan="1" colspan="1">Mudgil et al. (<xref>2024</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Gelatin and pregelatinized starch, maltodextrin and Cheese whey permeate</td><td align="center" rowspan="1" colspan="1">50.53</td><td align="center" rowspan="1" colspan="1">Zimmermann et al. (<xref>2024</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Binary matrix of inulin and maltodextrin</td><td align="center" rowspan="1" colspan="1">91.35</td><td align="center" rowspan="1" colspan="1">Cerón Córdoba et al. (<xref>2024</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Cruciferin/alginate</td><td align="center" rowspan="1" colspan="1">Not reported</td><td align="center" rowspan="1" colspan="1">Akbari et al. (<xref>2023</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Chitosan‐coated alginate‐inulin matrix</td><td align="center" rowspan="1" colspan="1">95.85</td><td align="center" rowspan="1" colspan="1">Parsana et al. (<xref>2023</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Skim milk and infant formula</td><td align="center" rowspan="1" colspan="1">90</td><td align="center" rowspan="1" colspan="1">Alves Gragnani Vido et al. (<xref>2023</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Binary matrix composed of inulin and maltodextrin</td><td align="center" rowspan="1" colspan="1">89.76</td><td align="center" rowspan="1" colspan="1">Jurado‐Gámez et al. (<xref>2023</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Skim milk, sodium glutamate, polyvinylpyrrolidone, maltodextrin and gelatin</td><td align="center" rowspan="1" colspan="1">Not reported</td><td align="center" rowspan="1" colspan="1">Wang et al. (<xref>2023</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Sodium alginate and sesame seed oil</td><td align="center" rowspan="1" colspan="1">89.22</td><td align="center" rowspan="1" colspan="1">Javed et al. (<xref>2023</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Alginate‐konjac gum</td><td align="center" rowspan="1" colspan="1">Not reported</td><td align="center" rowspan="1" colspan="1">Rodrigues et al. (<xref>2023</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Acacia gum</td><td align="center" rowspan="1" colspan="1">Not reported</td><td align="center" rowspan="1" colspan="1">do Nascimento et al. (<xref>2023</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Alginate</td><td align="center" rowspan="1" colspan="1">97</td><td align="center" rowspan="1" colspan="1">Abuqwider et al. (<xref>2023</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Pequi oil</td><td align="center" rowspan="1" colspan="1">96</td><td align="center" rowspan="1" colspan="1">Cedran et al. (<xref>2022</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">fish skin gelatin</td><td align="center" rowspan="1" colspan="1"> </td><td align="center" rowspan="1" colspan="1">Zhang et al. (<xref>2022</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Sodium hyaluronate</td><td align="center" rowspan="1" colspan="1">79</td><td align="center" rowspan="1" colspan="1">Li et al. (<xref>2022</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Isolated lignin complex from whey protein (WPI) and lignin</td><td align="center" rowspan="1" colspan="1">100</td><td align="center" rowspan="1" colspan="1">Diệp Huy Vũ et al. (<xref>2021b</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Hydrophobic surfactant into the MCT</td><td align="center" rowspan="1" colspan="1">84</td><td align="center" rowspan="1" colspan="1">Marefati et al. (<xref>2021</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">pomegranate (<italic>Punica granatum</italic>)</td><td align="center" rowspan="1" colspan="1">46</td><td align="center" rowspan="1" colspan="1">Bron et al. (<xref>2021</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Sunflower oil, Skim milk powder</td><td align="center" rowspan="1" colspan="1">98.93</td><td align="center" rowspan="1" colspan="1">Kil et al. (<xref>2020</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">novel enzymatically modified resistant starch type III</td><td align="center" rowspan="1" colspan="1">86.73</td><td align="center" rowspan="1" colspan="1">Khan et al. (<xref>2020</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">yellow passion fruit (<italic>Passiflora edulis</italic> Sims f. <italic>flavicarpa</italic> Deg.) and gelatin</td><td align="center" rowspan="1" colspan="1">99</td><td align="center" rowspan="1" colspan="1">Santos Monteiro et al. (<xref>2020</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">PVA</td><td align="center" rowspan="1" colspan="1">78</td><td align="center" rowspan="1" colspan="1">Ceylan et al. (<xref>2019</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Gum arabic</td><td align="center" rowspan="1" colspan="1">63</td><td align="center" rowspan="1" colspan="1">Guergoletto et al. (<xref>2017</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">gelatin</td><td align="center" rowspan="1" colspan="1">24.4</td><td align="center" rowspan="1" colspan="1">Guergoletto et al. (<xref>2017</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Holy basil essential oil</td><td align="center" rowspan="1" colspan="1">47.73 </td><td align="center" rowspan="1" colspan="1">Rodklongtan and Chitprasert (<xref>2017</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Milk powder</td><td align="center" rowspan="1" colspan="1">74.91</td><td align="center" rowspan="1" colspan="1">Liu et al. (<xref>2016</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">xanthan–gellan gum</td><td align="center" rowspan="1" colspan="1">72,5 </td><td align="center" rowspan="1" colspan="1">Juárez Tomás et al. (<xref>2015</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Aluminum carboxymethyl cellulose–rice bran</td><td align="center" rowspan="1" colspan="1">99.71 </td><td align="center" rowspan="1" colspan="1">Chitprasert et al. (<xref>2012</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Palm oil was of food grade</td><td align="center" rowspan="1" colspan="1">99</td><td align="center" rowspan="1" colspan="1">Chitprasert et al. (<xref>2012</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Mixtures of sodium alginate with other biopolymers</td><td align="center" rowspan="1" colspan="1">83.08</td><td align="center" rowspan="1" colspan="1">Akbari et al. (<xref>2023</xref>), Javed et al. (<xref>2023</xref>), Parsana et al. (<xref>2023</xref>), Rodrigues et al. (<xref>2023</xref>), Algaithi et al. (<xref>2022</xref>), Rodrigues et al. (<xref>2022</xref>), Popović et al. (<xref>2021</xref>), Karimi et al. (<xref>2021</xref>), Huang et al. (<xref>2021</xref>), Nakkarach and Withayagiat (<xref>2018</xref>), Atia et al. (<xref>2016</xref>), De Prisco et al. (<xref>2015</xref>), Rodklongtan and Chitprasert (<xref>2017</xref>), Petraitytė and Šipailienė (<xref>2019</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Sodium alginate</td><td align="center" rowspan="1" colspan="1">70</td><td align="center" rowspan="1" colspan="1">Ebrahimi Monfared et al. (<xref>2022</xref>), Qaziyani et al. (<xref>2019</xref>), Muthukumarasamy et al. (<xref>2006</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Maltodextrin</td><td align="center" rowspan="1" colspan="1">57</td><td align="center" rowspan="1" colspan="1">Guergoletto et al. (<xref>2017</xref>), Schell and Beermann (<xref>2014</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Blends of maltodextrin with other biopolymers</td><td align="center" rowspan="1" colspan="1">89.75</td><td align="center" rowspan="1" colspan="1">Gámez et al. (<xref>2021</xref>), Breitenbach et al. (<xref>2016</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Whey protein isolate, sucrose</td><td align="center" rowspan="1" colspan="1">68.4</td><td align="center" rowspan="1" colspan="1">Sompach et al. (<xref>2022</xref>)</td></tr><tr><td align="left" rowspan="1" colspan="1">Whey</td><td align="center" rowspan="1" colspan="1">0.54</td><td align="center" rowspan="1" colspan="1">Jantzen et al. (<xref>2013</xref>)</td></tr></tbody></table>These findings emphasize that the choice of wall material is as crucial as the encapsulation technique by itself. Consistent with the trends summarized in Table 4, several studies have shown that combining two or more coating materials further enhances encapsulation performance and probiotic viability. For example, Lau et al. (2025) developed a novel microencapsulation system using hemp seed protein isolate (HPI) and Cyclocarya paliurus leaf extracts (CP) for the targeted delivery of L. reuteri DPC16 to the human intestine. The optimized HPI/CP (9:1, w/w) formulation achieved the highest EE and maintained 93.06% survival in an in vitro gastrointestinal simulation (GITS) during 120 days of storage at −20°C, 4°C, and 25°C. After in vitro GITS, encapsulated L. reuteri DPC16 cells maintained mean cell counts of 7.2 log CFU/g, higher than those observed in unbound cells (5.5 log CFU/g; p < 0.05). In line with this evidence, several studies have shown that combining two or more coating materials further enhances encapsulation performance and probiotic viability. Confocal microscopy confirmed its robust survivability in the HPI/CP (9:1) matrices, with viability remaining above 10^6^ CFU/g after 120 days. These findings highlight the potential of HPI and CP formulations for developing functional foods and nutraceuticals with enhanced probiotic stability and survivability.
Besides, the alginate remains one of the most widely used coating materials due to its biocompatibility, low toxicity, cost‐effectiveness, and minimal impact on sensory attributes (Tekin et al. 2023). A key feature of alginate is its ability to form stable gels in the presence of divalent cations such as Ca^2^ ^+^, which is the fundamental mechanism exploited in ionotropic gelation techniques. Despite these advantages, when Alginate is used alone, encapsulation efficacies may be moderate, or can be relatively low (70%) and this makes it necessary to combine it with other biopolymers such as whey protein, chitosan, inulin, or plant mucilages (see Table 4) to increase efficiencies until averaged values of around 83% (Pereira Silveira et al. 2023).
On the other hand, regarding the encapsulation methods, they are selected based on the desired capsule size and type, the physical and chemical properties of the materials, and the expected release mechanism (Culas et al. 2024). Encapsulation methods can be classified as physical (e.g., spray drying, lyophilization, fluidized‐bed coating, extrusion, coextrusion and melt extrusion), physicochemical (e.g., molecular inclusion and liposomal encapsulation), or chemical (e.g., coacervation, ionic gelation and interfacial polymerization).
It is important to mention that every encapsulation technique has specific advantages and limitations. For example, spray drying remains the most scalable method; however, its high‐temperature exposure can reduce viability in sensitive strains. In contrast, EHDA including electrospray, avoids intense temperature variations and allows high particle size and morphology control, enhancing microbial integrity preservation; however, it presents limitations in terms of industrial scalability and cost‐effectiveness. Despite all the above, it has been observed that spray drying is emerging as one of the most widely used techniques for encapsulating microorganisms, followed by extrusion and ionic gelation (Figure 5). Particularly, ionic gelation has been extensively used due to its biocompatibility and the production of capsules with a broad size distribution; even so, it exhibits lower mechanical resilience, which could affect viability during storage or when incorporated into liquid food matrices.
Trend in the number of publications related to Lactobacillus reuteri encapsulation methods (Scopus database; keywords: “encapsulation” and “Lactobacillus reuteri,” year‐individual: 2004–2025) and limiting it to articles only.
Recently, coencapsulation strategies have also been explored to improve probiotic survival. These approaches usually combine probiotics with proteins, polysaccharides, or prebiotics through established methods like spray drying or ionic gelation. Promising results have been reported, for example with rice protein or pea–inulin mixtures, which achieved survival rates above 90% for L. reuteri DSM 17938 after spray drying. In addition to protecting the cells during GITS, these hybrid matrices seem to provide extra resistance to thermal and osmotic stress compared to single‐material systems.
Regarding Lactobacillus reuteri strains reported in encapsulation procedures, the analysis of the articles consulted for this review, demonstrated that the most studied strain is DSM 20016 due to its antimicrobial characteristics, like antifungal activity and effect against Gram positive and negative bacteria (Juárez‐Tomás et al. 2015; Rodrigues et al. 2023) and which has been used with different encapsulation matrices (Figure 6). In addition to the above, in a related study, L. reuteri DSM 20016 produced reuterin in situ (46.67 mmol/L) in alginate–konjac gum film‐forming solution through anaerobic glycerol bioconversion. After film production by extrusion, both L. reuteri cells and reuterin were successfully entrapped (98% and 67.6%, respectively). However, the metabolites responsible for these inhibitory effects need to be studied (Juárez‐Tomás et al. 2015). Moreover, it has been shown that the DPC16 strain colonizes human intestine due to the increased expression of proteins in the gastrointestinal mucosa, which supports its application in therapeutic supplements (Lau et al. 2025). Another very important studied strain is KUB‐AC5 for its probiotic viability (Diệp Huy Vũ et al. 2021a).
Articles related to encapsulated strains of Lactobacillus reuteri (Scopus database; keywords: “encapsulation” and “Lactobacillus reuteri,” year‐individual: 2004–2025) and limiting it to articles only.
In addition to these strains, encapsulation of L. reuteri CRL 1324 by ionic gelation extrusion with xanthan and gellan as coating materials, followed by lyophilization with lactose or skimmed milk as cryoprotectants, has also been investigated. This approach improved bacterial survival during prolonged storage at 4°C and increased viability in simulated vaginal fluid. The encapsulated cells also retained their functional properties by inhibiting the growth of Streptococcus agalactiae. Moreover, reuterin‐producing strains (DSM 20016 and DSM 17938) incorporated into alginate–konjac gum edible films were reported to inhibit foodborne pathogens (B. cereus, C. perfringens, P. aeruginosa) and spoilage fungi (F. oxysporum, C. gloeosporioides, A. alternata, P. digitatum), indicating broader antimicrobial potential. Overall, this review highlights the strains with the most significant number of studies in various polymer matrices and encapsulation methods, aiming to better understand current trends in encapsulation and the applications of this probiotic microorganism.
Regulatory
3.4
Beyond the technical progress in encapsulation, the relevance of L. reuteri depends largely on whether these advances can be translated into products that are safe, compliant with regulation, and attractive for the market.
Regulatory Overview
3.4.1
Regarding the regulations governing the use and application of probiotic microorganisms, the European Food Safety Authority (EFSA) applies the concept of Qualified Presumption of Safety (QPS) as a tool to simplify the safety assessment of microorganisms. If a species appears on the QPS list, as is the case with L. reuteri, it is considered safe unless there is evidence to the contrary (EFSA 2023). However, to date, there are regulatory barriers to health claims. In this regard, both the Health Claims Regulation (EC 1924/2006) and the Novel Foods Regulation (EU 2015/2283) require solid clinical evidence of their functionality, and the encapsulation of L. reuteri still requires additional studies to demonstrate stability, viability, bioavailability, safety, and functional benefits (EFSA 2024).
In addition to the above, the Food and Drug Administration in the United States, classifies probiotic microorganisms as dietary supplements or functional foods, depending on their formulation and intended use. Concerning their labeling, it is prohibited to suggest therapeutic effects unless supported by scientific evidence. This has prompted researchers and industry to adopt more rigorous evaluation methods. Furthermore, the US Pharmacopeia has recently highlighted that conventional toxicological approaches are not always suitable for evaluating live microorganisms (Merenstein et al. 2023; Roe et al. 2022).
Security, Regulations and Emerging Technological Trends (Limosilactobacillus reuteri Encapsulated)
3.4.2
From a safety perspective, the regulatory assessment of probiotic microorganisms, including L. reuteri, is primarily strain‐specific due to differences in genomic backgrounds and functional characteristics, which impact antibiotic resistance profiles. In this regard, the European Union's QPS provides a basis for microbial preassessment and several EFSA opinions detail the evaluation of various L. reuteri strains. However, labeling specific functions for each strain still requires solid scientific/clinical justification in accordance with the Regulation on Health Claims, particularly for claims related to gastrointestinal or immunomodulatory benefits (EFSA BIOHAZ Panel 2023).
In addition to the above, in the United States, regulations depend specifically on the intended use, that is, food versus supplements. Notifications have been issued for foods that do not pose a risk when used correctly. For example, the United States has recently issued specific notifications for the DSM 17938 strain, supporting its use in various food categories under defined conditions (FDA 2008). However, despite existing regulations, encapsulating these strains introduces an additional layer of regulatory and safety considerations beyond the microorganism itself. Specifically, food quality can be affected by wall materials, as incorporating polymeric matrices based on alginate, chitosan, milk proteins, or plant polymers such as pectins can alter labeling requirements, stability during processing, sensory properties, and consumer acceptance (Cook et al. 2013).
Current technological research focuses on multilayer capsules and designs that enhance cell protection during processing and gastrointestinal stress. Encapsulation strategies that combine L reuteri with prebiotic substrates are also being explored to improve viability and functional performance after release.
Conclusion
4
The evidence compiled in this review highlights L. reuteri as a versatile and highly promising probiotic, whose functional properties extend from antimicrobial and anti‐inflammatory activity to metabolic regulation, antioxidant effects, and applications in gastrointestinal, oral, and systemic health. Until now, the ability of this microorganism to exert benefits has been strongly conditioned by its survival processing, storage and gastrointestinal transit. Encapsulation has emerged as one of the most effective strategies to address these challenges, with ionic gelation, extrusion, spray drying, and some other approaches. According to the literature consulted in this review, alginate remains as the most widely used biopolymer used in encapsulation due to its high biocompatibility and low cost. However, recent studies have demonstrated that some other materials including proteins and polysaccharides improve EE and controlled release. Despite all the progress in this matter, critical gaps remain in translating the laboratory research to industry scale. Most current studies are limited to in vitro models, with fewer investigations exploring performance in food matrices or under real industry‐scale conditions. Moreover, there is a limited knowledge of how encapsulation affects consumer acceptance and regulatory approval pathways. To move forward, future research must prioritize integrated approaches that combine material science, microbiology and clinical validation. Also, multidisciplinary efforts will allow the development of encapsulation platforms with capacity to maintain probiotic viability and ensure functional efficacy in vivo.
Despite the wide variety of functional properties exhibited by L. reuteri strains ranging from antimicrobial activity to therapeutic potential studies on their encapsulation remain limited. This limitation makes it difficult to predict their technological behavior and response to various processes. Hence, the importance of expanding research, especially under conditions that replicate real‐life food matrices and industrial processing scenarios.
In the next stage of research, work on encapsulation should move past basic comparisons of insufficient materials and methods. It is essential to consider how these strategies impact release kinetics and sensory response in both food and pharmaceutical applications. Such an approach will support the creation of delivery systems specifically designed to maximize the functional benefits of L. reuteri and to demonstrate consistent performance in real‐world applications.
Nomenclature
ABTS2,2‐azino‐bis‐3‐ethylbenzothiazoline‐6‐sulfonic acidCFUcolony forming unitsCAGRcompound annual growth rate CAGRDPCdairy products collectionCP Cyclocarya paliurus leaf extractsDPPH2,2‐diphenyl‐1‐picrylhydrazylDSMDeutsche Sammlung von MikroorganismenEHDAelectrohydrodynamic atomizationEFSAEuropean Food Safety AuthorityFAOFood and Agriculture OrganizationGITSgastrointestinal simulationGRASgenerally recognized as safeHPIhemp seed protein isolateLABlactic acid bacteriaLR Limosilactobacillus reuteri pHpotential of hydrogenQPSQualified Presumption of SafetySEMscanning electron microscopyWHOWorld Health Organization
Author Contributions
León‐Espinosa Erika Berenice: conceptualization, methodology, writing – review and editing, investigation, writing – original draft. Barrios‐Francisco Rigoberto: methodology, writing – review and editing. Colin‐Molina Abraham: investigation, resources, writing – review and editing. Martínez‐Palma Nikte Yoliztli: investigation, writing – review and editing. Rentería‐Ortega Minerva: conceptualization, methodology, resources, writing – original draft, writing – review and editing, supervision, project administration.
Conflicts of Interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abuqwider, J. , A. Di Porzio , V. Barrella , et al. 2023. “Limosilactobacillus Reuteri DSM 17938 Reverses Gut Metabolic Dysfunction Induced by Western Diet in Adult Rats.” Frontiers in Nutrition 10: 1236417. 10.3389/fnut.2023.1236417.37908302 PMC 10613642 · doi ↗ · pubmed ↗
- 2Ahmed, B. , and J. C. Konje . 2023. “The Epidemiology of Obesity in Reproduction.” Best Practice & Research Clinical Obstetrics & Gynaecology 89: 102342. 10.1016/j.bpobgyn.2023.102342.37276817 · doi ↗ · pubmed ↗
- 3Akbari, A. , M. G. Gänzle , and J. Wu . 2023. “Cruciferin Improves Stress Resistance and Simulated Gastrointestinal Survival of Probiotic Limosilactobacillus Reuteri in the Model Encapsulation System.” Food Hydrocolloids for Health 3: 100118. 10.1016/j.fhfh.2023.100118. · doi ↗
- 4Alam, M. J. , M. M. Z. Islam , M. A. Tayab , et al. 2022. “Role of Probiotic Lactobacillus reuteri in Improving Gut Health and Immunity in Infants and Toddlers: A Review.” International Journal of Nutrition Sciences 7, no. 2: 75–80. 10.30476/ijns.2022.94849.1182. · doi ↗
- 5Algaithi, M. , P. Mudgil , M. Hamdi , et al. 2022. “ Lactobacillus reuteri‐Fortified Camel Milk Infant Formula: Effects of Encapsulation, In Vitro Digestion, and Storage Conditions on Probiotic Cell Viability and Physicochemical Characteristics of Infant Formula.” Journal of Dairy Science 105, no. 11: 8621–8637. 10.3168/jds.2022-22008.36207183 · doi ↗ · pubmed ↗
- 6Allameh, A. , R. Niayesh‐Mehr , A. Aliarab , G. Sebastiani , and K. Pantopoulos . 2023. “Oxidative Stress in Liver Pathophysiology and Disease.” Antioxidants 12, no. 9: 1653. 10.3390/antiox 12091653.37759956 PMC 10525124 · doi ↗ · pubmed ↗
- 7Alok, A. , I. D. Singh , S. Singh , M. Kishore , P. C. Jha , and M. A. Iqubal . 2017. “Probiotics: A New Era of Biotherapy.” Advanced Biomedical Research 6: 31. 10.4103/2277-9175.192625.28401078 PMC 5360003 · doi ↗ · pubmed ↗
- 8Alves Gragnani Vido, M. , I. Dutra Alvim , G. Vinderola , et al. 2023. “Microencapsulation of Limosilactobacillus Reuteri (DSM 23878) for Application in Infant Formula: Heat Resistance and Bacterial Viability During Long‐time Storage.” Food Research International 173: 113378. 10.1016/j.foodres.2023.113378.37803716 · doi ↗ · pubmed ↗