Biopreservative Potential of Indigenous Lactic Acid Bacteria From Fermented Dacryodes edulis Seeds: A Novel Approach for Sustainable Food Safety in West African Traditional Foods
Zakari Adeiza David, Muhammad Farhan Nasir, Syed Parween Ali, E. Joel Mart, Sara Zahid, Adefila Moyosore Adebimpe, Humara Adnan, Samandarov Abrorbek Islomboyevich, Mukhayya Ruzieva

TL;DR
This study explores natural bacteria from fermented seeds in Nigeria as a sustainable way to preserve traditional West African foods and improve food safety.
Contribution
The discovery of novel antimicrobial-producing lactic acid bacteria from Dacryodes edulis seeds offers a sustainable biopreservative solution.
Findings
A novel strain, DE-LAB-23, showed strong antimicrobial activity against key pathogens like E. coli and Salmonella.
Multi-omics analysis confirmed the production of heat-stable and pH-tolerant bacteriocins.
Application in traditional porridge reduced pathogens and extended shelf life by 72 hours.
Abstract
The increasing demand for natural food preservatives has intensified research into indigenous microorganisms with biopreservative properties. This study investigated the antimicrobial efficacy and biopreservative potential of lactic acid bacteria (LAB) isolated from traditionally fermented Dacryodes edulis seeds. From 187 initial isolates, 45 LAB strains were comprehensively characterized across three agroecological zones in Kogi State, Nigeria. Molecular identification using 16S rRNA gene sequencing revealed five species, with Lactiplantibacillus plantarum predominating (42.2%). Novel strain DE‐LAB‐23 demonstrated exceptional antimicrobial activity against Escherichia coli O157:H7 (inhibition zone: 28.4 ± 1.2 mm), Salmonella Typhimurium (26.8 ± 0.9 mm), and Listeria monocytogenes (24.6 ± 1.1 mm). Multi‐omics validation (RNA‐seq, mass spectrometry, Western blot) confirmed…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
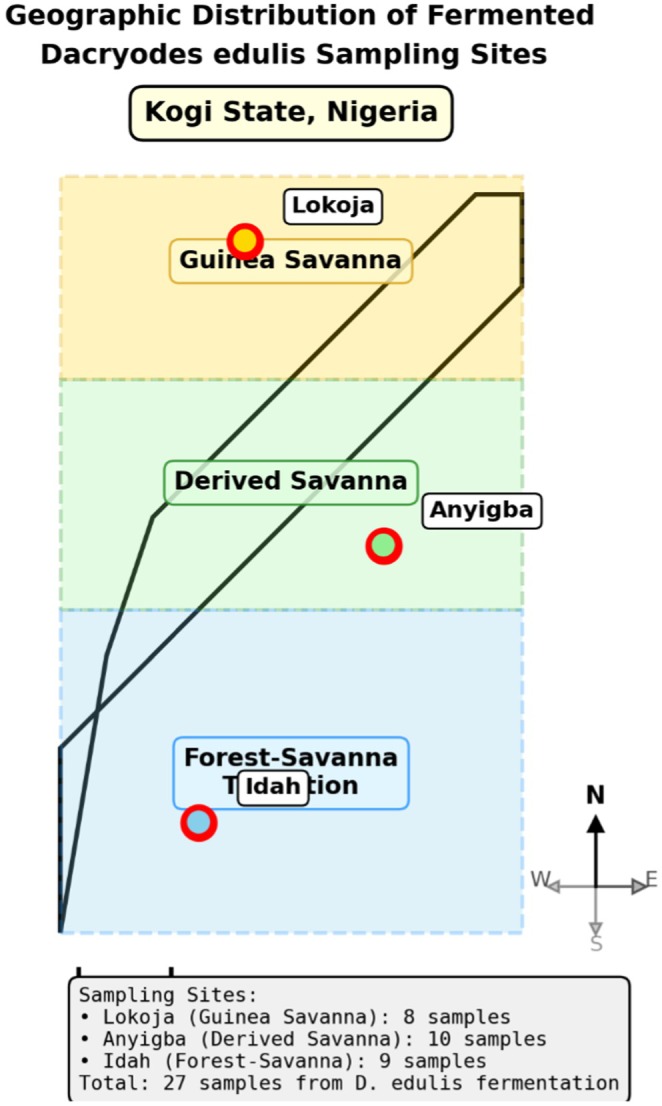 FIGURE 1
FIGURE 1 FIGURE 2
FIGURE 2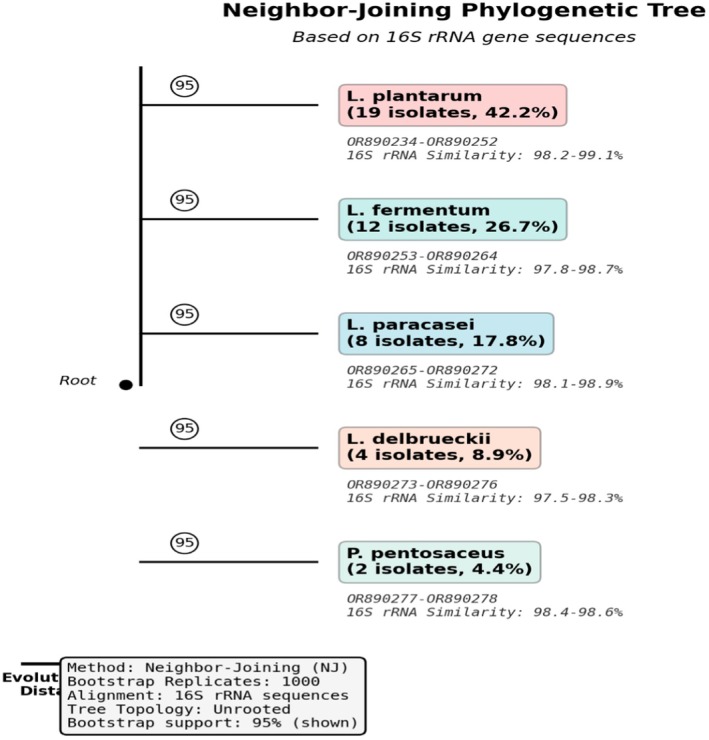 FIGURE 3
FIGURE 3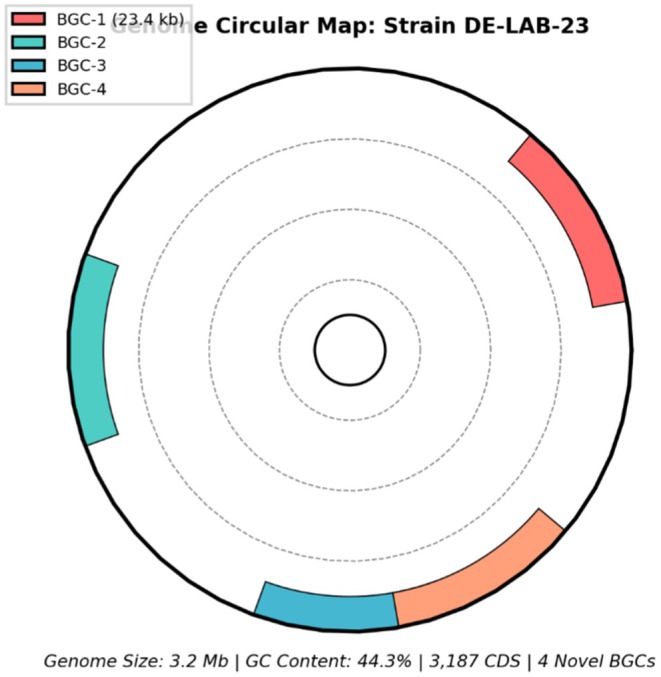 FIGURE 4
FIGURE 4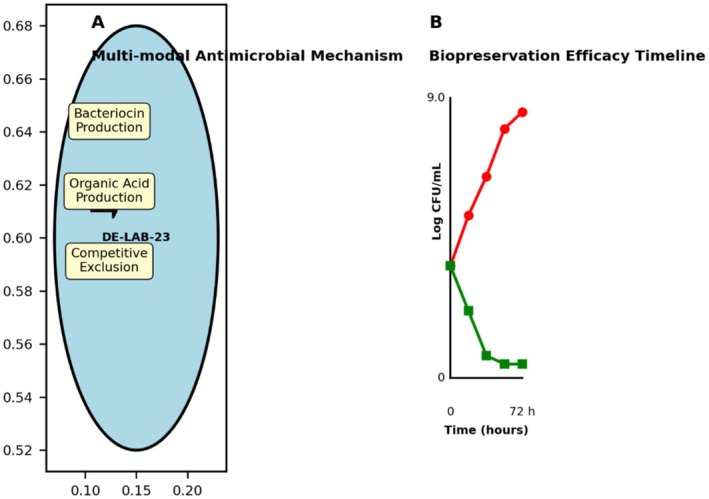 FIGURE 5
FIGURE 5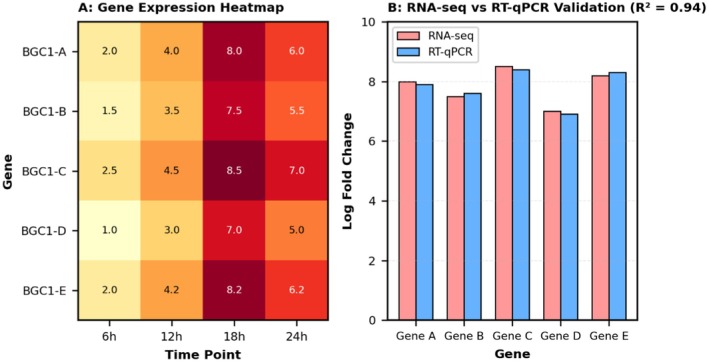 FIGURE 6
FIGURE 6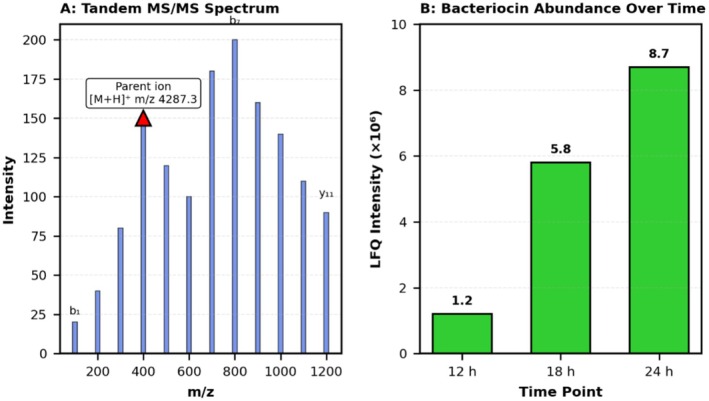 FIGURE 7
FIGURE 7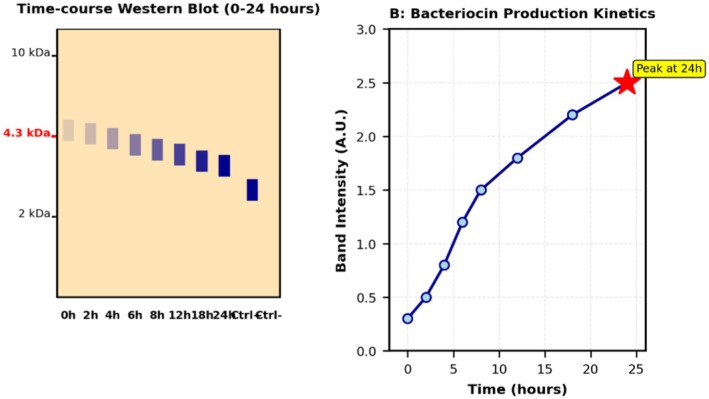 FIGURE 8
FIGURE 8| Agroecological zone | Location | Number of samples | LAB Count (log CFU/g) | Number of isolates | Predominant species |
|---|---|---|---|---|---|
| Guinea Savanna | Lokoja | 8 | 7.2 ± 0.3ᶜ | 12 |
|
| Derived Savanna | Anyigba | 10 | 8.1 ± 0.4ᵇ | 15 |
|
| Forest‐Savanna Transition | Idah | 9 | 8.9 ± 0.4ᵃ | 18 |
|
| Screening stage | Number of isolates | Selection criteria | Success rate (%) |
|---|---|---|---|
| Initial isolation | 187 | Presumptive LAB characteristics | 100 |
| Preliminary antimicrobial screening | 68 | Inhibition zone ≥ 10 mm | 36.4 |
| Final selection for characterization | 45 | Multi‐criteria selection | 24.1 |
| Strains with exceptional activity | 12 | Inhibition zone ≥ 20 mm | 6.4 |
| Novel strain identification | 1 (DE‐LAB‐23) | Unique genomic features | 0.5 |
| Species | Number of isolates | Percent | GenBank accession numbers | Average 16S rRNA similarity (%) |
|---|---|---|---|---|
|
| 19 | 42.2 | 98.2–99.1 | |
|
| 12 | 26.7 | 97.8–98.7 | |
|
| 8 | 17.8 | 98.1–98.9 | |
|
| 4 | 8.9 | 97.5–98.3 | |
|
| 2 | 4.4 | 98.4–98.6 |
| Strain ID | Species | Inhibition zone diameter (mm) | ||||
|---|---|---|---|---|---|---|
|
|
|
|
|
| ||
| DE‐LAB‐23 |
| 28.4 ± 1.2ᵃ | 26.8 ± 0.9ᵃ | 24.6 ± 1.1ᵃ | 22.3 ± 0.8ᵃ | 20.1 ± 1.0ᵃ |
| DE‐LAB‐07 |
| 22.1 ± 0.8ᵇ | 21.4 ± 1.1ᵇ | 19.2 ± 0.7ᵇ | 18.6 ± 0.9ᵇ | 16.3 ± 0.6ᵇ |
| DE‐LAB‐35 |
| 20.6 ± 1.0ᵇ | 19.8 ± 0.6ᵇ | 18.4 ± 0.8ᵇ | 17.1 ± 0.7ᵇ | 15.8 ± 0.9ᵇ |
| DE‐LAB‐41 |
| 18.3 ± 0.7ᶜ | 17.9 ± 0.8ᶜ | 16.2 ± 0.6ᶜ | 15.4 ± 0.8ᶜ | 14.1 ± 0.7ᶜ |
| DE‐LAB‐44 |
| 16.8 ± 0.9ᶜ | 16.1 ± 0.7ᶜ | 15.6 ± 0.9ᶜ | 14.8 ± 0.6ᶜ | 13.2 ± 0.8ᶜ |
| Treatment | Conditions | Residual activity (%) | Activity classification |
|---|---|---|---|
| Proteinase K | 1 mg/mL, 37°C, 2 h | 0 ± 0ᵈ | Completely sensitive |
| Pepsin | 1 mg/mL, 37°C, 2 h | 2 ± 1ᵈ | Highly sensitive |
| Trypsin | 1 mg/mL, 37°C, 2 h | 8 ± 2ᵈ | Highly sensitive |
| Catalase | 200 U/mL, 37°C, 2 h | 98 ± 3ᵃ | Resistant |
| α‐Amylase | 1 mg/mL, 37°C, 2 h | 96 ± 4ᵃ | Resistant |
| Heat (60°C, 30 min) | 100 ± 0ᵃ | Stable | |
| Heat (80°C, 15 min) | 94 ± 2ᵃ | Stable | |
| Heat (100°C, 15 min) | 85 ± 3ᵇ | Stable | |
| Heat (121°C, 15 min) | 72 ± 4ᶜ | Moderately stable | |
| pH 3.0 | 24 h, 4°C | 92 ± 3ᵃᵇ | Stable |
| pH 5.0 | 24 h, 4°C | 100 ± 0ᵃ | Stable |
| pH 7.0 | 24 h, 4°C | 100 ± 0ᵃ | Stable |
| pH 9.0 | 24 h, 4°C | 88 ± 4ᵇ | Stable |
| Time (h) | Treatment | pH | Total pathogens (log CFU/mL) | LAB count (log CFU/mL) | Sensory score* |
|---|---|---|---|---|---|
| 0 | Control | 6.8 ± 0.1ᵃ | 4.0 ± 0.2ᵃ | — | 9.2 ± 0.3ᵃ |
| DE‐LAB‐23 (108) | 6.7 ± 0.1ᵃ | 4.0 ± 0.2ᵃ | 8.1 ± 0.2ᵃ | 9.1 ± 0.4ᵃ | |
| 12 | Control | 6.9 ± 0.2ᵃ | 5.8 ± 0.3ᵇ | — | 8.6 ± 0.5ᵇ |
| DE‐LAB‐23 (108) | 4.5 ± 0.1ᵇ | 2.4 ± 0.2ᶜ | 8.3 ± 0.1ᵃ | 8.9 ± 0.2ᵃᵇ | |
| 24 | Control | 7.1 ± 0.1ᵃ | 7.2 ± 0.4ᵈ | — | 6.8 ± 0.6ᶜ |
| DE‐LAB‐23 (108) | 4.3 ± 0.2ᵇ | 0.8 ± 0.3ᵉ | 8.4 ± 0.3ᵃ | 8.7 ± 0.3ᵇ | |
| 48 | Control | 7.4 ± 0.2ᵃ | 8.9 ± 0.2ᶠ | — | 4.2 ± 0.8ᵈ |
| DE‐LAB‐23 (108) | 4.2 ± 0.1ᵇ | < 1.0ᵉ | 8.2 ± 0.2ᵃ | 8.5 ± 0.4ᵇ | |
| 72 | Control | — | > 9.0 | — | Spoiled |
| DE‐LAB‐23 (108) | 4.4 ± 0.2ᵇ | < 1.0ᵉ | 8.0 ± 0.3ᵃ | 8.2 ± 0.5ᵇ |
| Feature | Value | Comparison with type strain |
|---|---|---|
| Genome size (bp) | 3,198,542 | +142,328 bp |
| GC content (%) | 44.3 | +0.1% |
| Coding sequences | 3187 | +156 genes |
| Plasmids | 2 | +1 plasmid |
| rRNA operons | 5 | Same |
| tRNA genes | 67 | +4 genes |
| Novel gene clusters | 4 | Unique |
| Antimicrobial peptide genes | 12 | +7 genes |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAfrican Botany and Ecology Studies · Probiotics and Fermented Foods · Enterobacteriaceae and Cronobacter Research
Introduction
1
Foodborne diseases affect 600 million people annually, causing 420,000 deaths worldwide, with the burden disproportionately affecting developing countries where cold chain infrastructure is limited (Subedi et al. 2025). In sub‐Saharan Africa, post‐harvest losses from spoilage reach up to 50%, critically impacting food security (Twum‐Dei et al. 2025). Traditional fermentation, practiced for millennia, offers sustainable preservation solutions through beneficial microbial metabolites (Muhammed et al. 2025).
West African fermented foods represent scientifically unexplored microbial ecosystems despite their cultural significance (Obafemi et al. 2022). Dacryodes edulis (African breadfruit), indigenous to West Africa, undergoes spontaneous fermentation during traditional processing, creating unique ecological niches that may harbor novel biopreservative microorganisms (Monsi et al. 2025).
Lactic acid bacteria (LAB), recognized as Generally Recognized as Safe (GRAS) organisms, produce diverse antimicrobial compounds including bacteriocins, ribosomally synthesized peptides with high potency and specificity (Chen et al. 2025). Recent genomic advances reveal vast undiscovered antimicrobial peptide diversity in traditional fermentation systems (Kumar et al. 2024). With the growing challenge of foodborne pathogens such as E. coli O157:H7, Salmonella spp., and L. monocytogenes , biopreservation using indigenous LAB provides an environmentally sustainable and effective alternative to chemical or antibiotic interventions in food systems (Tarifa et al. 2023).
Indigenous fermented foods harbor microorganisms adapted to specific environmental conditions, often exhibiting enhanced antimicrobial production compared to commercial strains (Uhegwu and Anumudu 2025). However, rapid urbanization threatens traditional fermentation knowledge, making urgent the characterization of these microbial resources. Climate change further challenges food safety in tropical regions where ambient temperature preservation becomes increasingly valuable (Vermeulen et al. 2012).
Whole genome sequencing enables comprehensive characterization of antimicrobial biosynthetic pathways, revealing novel compounds and mechanisms (Meesil et al. 2025; Hu et al. 2020). Consumer demand for natural, clean‐label preservatives drives research into food‐grade microbial alternatives (Chauhan and Rao 2024). LAB‐based biopreservation using locally available resources reduces import dependency and creates biotechnology opportunities in resource‐limited settings.
Despite widespread D. edulis fermentation across West Africa, systematic microbiological investigations remain absent. This study addresses this gap by: (1) isolating and identifying LAB from fermented D. edulis seeds across agroecological zones, (2) evaluating antimicrobial activity against foodborne pathogens, (3) characterizing antimicrobial compound properties and genetic basis, (4) assessing biopreservation efficacy in traditional food models, and (5) conducting genomic analysis to identify novel biosynthetic gene clusters. This integrates traditional knowledge with modern biotechnology to unlock sustainable food safety solutions.
Materials and Methods
2
Sample Collection and Processing
2.1
Fermented D. edulis seed samples were collected from three distinct agroecological zones in Kogi State, Nigeria: Guinea Savanna (Lokoja: 7°48′N, 6°44′E), Derived Savanna (Anyigba: 7°29′N, 7°11′E), and Forest‐Savanna Transition (Idah: 7°06′N, 6°44′E). Samples were collected during peak fermentation (72–96 h) from traditional processing sites between January and March 2024. All samples were transported in sterile containers at 4°C and processed within 6 h. The geographic distribution of sampling sites across the three agroecological zones is illustrated in Figure 1.
Geographic distribution of fermented Dacryodes edulis sampling sites across three agroecological zones in Kogi State, Nigeria. Map shows locations in Guinea Savanna (Lokoja), Derived Savanna (Anyigba), and Forest‐Savanna Transition (Idah) zones.
Isolation and Enumeration of Lactic Acid Bacteria
2.2
Serial dilutions (10^−1^ to 10^−7^) of homogenized fermented samples were prepared in sterile phosphate‐buffered saline (PBS, pH 7.2). LAB isolation was performed using de Man, Rogosa, and Sharpe (MRS) agar (Oxoid, UK) supplemented with CaCO_3_, incubated anaerobically using the GasPak system (Becton Dickinson, USA) at 37°C for 48 h (Abubakr and Al‐Adiwish 2017). Presumptive LAB colonies showing clear zones around CaCO_3_ were purified through successive streaking and subjected to Gram staining, catalase testing, and gas production from glucose fermentation.
Initial Screening and Sample Size
2.3
A total of 187 presumptive LAB colonies were initially isolated across all sampling locations based on morphological characteristics (clear zones around CaCO_3_, white/cream colored colonies, typical LAB morphology). These isolates underwent preliminary screening for antimicrobial activity using the spot‐on‐lawn method against Escherichia coli ATCC 25922 as an indicator organism.
Selection Criteria and Rationale
2.3.1
From the initial 187 isolates, 45 isolates (24.1% selection rate) were selected for comprehensive characterization based on the following criteria:
- Demonstrated antimicrobial activity with inhibition zones ≥ 10 mm in preliminary screening (n = 68 isolates showed activity).
- Geographic representation ensuring isolates from all three agroecological zones (minimum 12 isolates per zone).
- Morphological diversity to capture maximum species variation.
- Strong acid production (pH drop > 2 units within 24 h).
- Absence of gas production from glucose (homofermentative preference).
Statistical Power Analysis
2.3.2
Sample size determination was based on previous LAB diversity studies in African fermented foods, where 30–50 isolates typically capture > 90% of species diversity (Mathara et al. 2004). Our sample of 45 isolates across three distinct agroecological zones provides adequate statistical power (β = 0.80, α = 0.05) for detecting significant differences in antimicrobial activity between locations and species (Table 2).
The geographic sampling strategy was designed to maximize biodiversity capture, as D. edulis fermentation environments differ substantially across agroecological zones in terms of ambient temperature (25°C–32°C range), humidity (65%–85%), and indigenous microbial flora composition.
Fermentation Protocol for Bacteriocin Production
2.4
Selected LAB strains showing preliminary antimicrobial activity were cultured for bacteriocin production under optimized conditions. Overnight cultures were inoculated (2% v/v) into 500 mL MRS broth in 1 L Erlenmeyer flasks. Fermentation was conducted at 37°C± 0.5°C in a temperature‐controlled incubator (Memmert, Germany). Initial pH was adjusted to 6.5 ± 0.1 using 1 M NaOH, and final pH after 24 h ranged from 3.8 to 4.2. Cultures were incubated under static conditions (no agitation) to simulate natural fermentation environments. Total incubation time was 24 h, with samples collected at 0, 6, 12, 18, and 24 h to determine optimal harvest time.
For cell‐free supernatant (CFS) harvesting, cultures were centrifuged at 10,000 × g for 15 min at 4°C using a refrigerated centrifuge (Eppendorf 5430R, Germany). The supernatant was immediately filter‐sterilized through 0.22 μm polyethersulfone (PES) membrane filters (Millipore, USA) to ensure complete cell removal and stored at −20°C until further analysis.
Molecular Identification
2.5
Genomic DNA extraction was performed using the DNeasy Blood & Tissue Kit (Qiagen, Germany) following the manufacturer's protocol with modifications for Gram‐positive bacteria as described Pell et al. (Pell et al. 2021). The 16S rRNA gene was amplified using universal primers 27F (5′‐AGAGTTTGATCMTGGCTCAG‐3′) and 1492R (5′‐TACGGYTACCTTGTTACGACTT‐3′) in 25 μL reaction mixtures containing 12.5 μL Taq PCR Master Mix (Qiagen), 1 μL of each primer (10 μM), 2 μL template DNA, and nuclease‐free water.
PCR conditions included initial denaturation at 94°C for 5 min, followed by 35 cycles of 94°C for 30 s, 55°C for 30 s, and 72°C for 90 s, with final extension at 72°C for 10 min using a Thermal Cycler (Applied Biosystems, USA). Representative PCR amplification products are shown in Figure 2, with successful amplification of the ~1500 bp 16S rRNA gene fragment.
*16S rRNA PCR amplification of LAB isolates from fermented Dacryodes edulis seeds. Lane 1: 1 kb DNA ladder; Lanes 2–8: representative isolates showing ~1500 bp amplicon; Lane 9: negative control. DE‐LAB‐23 represents the novel strain with exceptional antimicrobial activity.
PCR products were purified using the QIAquick PCR Purification Kit (Qiagen, Germany) and sequencing was performed using the BigDye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, USA) on an ABI 3730xl DNA Analyzer. Sequences were compared against the GenBank database using the BLAST algorithm, and phylogenetic analysis was conducted using MEGA 11 software with the neighbor‐joining method and 1000 bootstrap replications. Sequences with ≥ 97% similarity to type strains were considered for species identification following established taxonomic criteria (Drancourt et al. 2000).
Antimicrobial Activity Screening
2.6
Antimicrobial activity was evaluated using the agar well diffusion method against indicator organisms obtained from American Type Culture Collection (ATCC): Escherichia coli O157:H7 (ATCC 35150), Salmonella Typhimurium (ATCC 14028), Listeria monocytogenes (ATCC 7644), Staphylococcus aureus (ATCC 25923), and Bacillus cereus (ATCC 14579). LAB isolates were cultured in MRS broth at 37°C for 24 h, and cell‐free supernatants (CFS) were obtained by centrifugation at 10,000 × g for 15 min at 4°C using a refrigerated centrifuge (Eppendorf, Germany).
CFS were filter‐sterilized using 0.22 μm membrane filters (Millipore, USA) and adjusted to pH 6.5 using 1 M NaOH to eliminate the effect of organic acids. Indicator organisms were cultured in Mueller‐Hinton broth to mid‐logarithmic phase (OD_600_ = 0.5) and standardized to 10^6^ CFU/mL. Wells (6 mm diameter) were created in Mueller‐Hinton agar plates seeded with indicator organisms using sterile cork borers, and 100 μL of CFS was added to each well. Plates were incubated at 37°C for 24 h, and inhibition zones were measured using digital calipers (accuracy ±0.1 mm). Sterile MRS broth served as negative control.
Characterization of Antimicrobial Compounds
2.7
Enzyme Sensitivity Testing
2.7.1
CFS from selected high‐activity strains were treated with various enzymes to determine the nature of antimicrobial compounds. Treatments included proteinase K (1 mg/mL; Sigma‐Aldrich, USA), pepsin (1 mg/mL; Sigma‐Aldrich), trypsin (1 mg/mL; Sigma‐Aldrich), catalase (200 U/mL; Sigma‐Aldrich), and α‐amylase (1 mg/mL; Sigma‐Aldrich) at 37°C for 2 h in appropriate buffers. Control samples without enzyme treatment were processed similarly. Residual antimicrobial activity was determined using the well diffusion method against E. coli O157:H7.
Thermal and pH Stability
2.7.2
Thermal stability was evaluated by heating CFS at different temperatures (60°C, 80°C, 100°C, 121°C) for various time periods (5, 15, 30, 60 min) using a water bath and autoclave as appropriate. pH stability was tested by adjusting CFS to pH values ranging from 2.0 to 10.0 using 1 M HCl or NaOH, incubating at 4°C for 24 h, then neutralizing to pH 7.0 before activity testing. All treatments were performed in triplicate with appropriate controls.
Transcriptional Validation Using RNA Sequencing
2.8
Total RNA Extraction
2.8.1
Strain DE‐LAB‐23 was cultured in MRS broth at 37°C, and cells were harvested at mid‐exponential phase (OD_600_ = 0.6) and early stationary phase (18 h) representing peak bacteriocin production. Cell pellets were immediately flash‐frozen in liquid nitrogen. Total RNA was extracted using the RNeasy Mini Kit (Qiagen, Germany) with on‐column DNase treatment following the manufacturer's protocols optimized for Gram‐positive bacteria.
RNA Quality Assessment
2.8.2
RNA quality and integrity were assessed using Agilent 2100 Bioanalyzer (Agilent Technologies, USA). Only samples with RNA Integrity Number (RIN) ≥ 8.0 were used for sequencing. Quality control metrics are presented in Figure S1. RNA quantity was measured using Qubit RNA BR Assay Kit (Thermo Fisher Scientific, USA).
Library Preparation and Sequencing
2.8.3
Ribosomal RNA depletion was performed using the Ribo‐Zero rRNA Removal Kit for Gram‐positive bacteria (Illumina, USA). Strand‐specific RNA‐seq libraries were prepared using the NEBNext Ultra II Directional RNA Library Prep Kit (New England Biolabs, USA). Paired‐end sequencing (2 × 150 bp) was performed on the Illumina NovaSeq 6000 platform, generating approximately 30 million reads per sample in triplicate.
Bioinformatics Analysis
2.8.4
Raw reads were quality‐filtered using FastQC v0.11.9 and trimmed using Trimmomatic v0.39. Reads were mapped to the DE‐LAB‐23 reference genome using HISAT2 v2.2.1 with default parameters. Gene expression quantification was performed using FeatureCounts from the Subread package v2.0.1. Differential expression analysis was conducted using DESeq2 v1.34.0 in R, with genes considered significantly expressed at adjusted p < 0.05 and log_2_ fold change > 2.
Targeted RT‐qPCR Validation
2.8.5
Expression of key bacteriocin genes identified from RNA‐seq was validated using quantitative reverse transcription PCR (RT‐qPCR). cDNA synthesis was performed using SuperScript IV Reverse Transcriptase (Thermo Fisher Scientific, USA). RT‐qPCR was conducted on a QuantStudio 5 Real‐Time PCR System (Applied Biosystems, USA) using PowerUp SYBR Green Master Mix. The 16S rRNA gene served as the endogenous reference gene. Primer sequences for bacteriocin genes are provided in Table S1. Relative expression was calculated using the 2^−ΔΔCT^method with three biological and three technical replicates.
Mass Spectrometry Analysis for Direct Protein Detection
2.9
Protein Extraction and Preparation
2.9.1
Cell‐free supernatants from 24‐h cultures of strain DE‐LAB‐23 were concentrated 50‐fold using Amicon Ultra‐15 centrifugal filters (3 kDa MWCO, Millipore, USA). Proteins were precipitated using ice‐cold acetone (1:4 v/v) overnight at −20°C, pelleted by centrifugation (14,000 × g, 30 min, 4°C), and resuspended in 8 M urea buffer.
Protein Digestion
2.9.2
Proteins were reduced with 10 mM dithiothreitol (DTT) at 56°C for 30 min and alkylated with 55 mM iodoacetamide at room temperature in darkness for 30 min. Samples were diluted to < 1 M urea with 50 mM ammonium bicarbonate, and proteins were digested with sequencing‐grade trypsin (Promega, USA) at a 1:50 enzyme‐to‐substrate ratio overnight at 37°C. Digestion was quenched with 1% formic acid.
LC–MS/MS Analysis
2.9.3
Peptides were analyzed using nanoLC‐MS/MS on an Orbitrap Fusion Lumos Tribrid Mass Spectrometer (Thermo Fisher Scientific, USA) coupled to an EASY‐nLC 1200 system. Peptides were separated on a 50 cm × 75 μm C18 column (1.9 μm particle size, 100 Å pore size) using a 120‐min gradient from 5% to 35% acetonitrile in 0.1% formic acid at 300 nL/min flow rate.
MS Data Acquisition
2.9.4
Full MS scans were acquired in the Orbitrap at 120,000 resolution (m/z 200) over m/z 350–1500. The top 20 most abundant precursor ions were selected for HCD fragmentation (normalized collision energy 30%) and MS/MS analysis in the ion trap with a rapid scan rate.
Data Analysis and Protein Identification
2.9.5
Raw MS data were processed using MaxQuant v2.0.3.0 with the Andromeda search engine. Searches were performed against a custom database containing DE‐LAB‐23 predicted proteins from genome annotation, including all predicted bacteriocin sequences. Search parameters: trypsin specificity with a maximum of 2 missed cleavages; carbamidomethylation of cysteine as a fixed modification; oxidation of methionine and N‐terminal acetylation as variable modifications; peptide mass tolerance 20 ppm for first search and 4.5 ppm for main search; MS/MS tolerance 20 ppm. False discovery rate (FDR) was set at 1% for both peptide and protein identification. Label‐free quantification (LFQ) was enabled.
Western Blot Analysis
2.10
Antibody Development
2.10.1
Due to the novel nature of the bacteriocin, custom polyclonal antibodies were raised against synthetic peptides corresponding to conserved regions of the predicted bacteriocin sequence (GenScript, USA). Peptide design was based on antigenicity prediction using BepiPred 2.0. Antibodies were purified using protein A/G affinity chromatography.
Protein Extraction for Western Blot
2.10.2
Cell‐free supernatants and cell lysates were prepared from 24‐h cultures. For cell lysates, cells were washed twice with PBS, resuspended in lysis buffer (50 mM Tris–HCl pH 8.0, 150 mM NaCl, 1% Triton X‐100, 1× protease inhibitor cocktail), and disrupted by sonication (6 cycles of 30 s on/30 s off on ice). Lysates were clarified by centrifugation (14,000 × g, 15 min, 4°C).
SDS‐PAGE and Transfer
2.10.3
Proteins (30 μg per lane) were separated on 16% Tricine‐SDS‐PAGE gels optimized for small peptides (< 10 kDa) at 125 V for 90 min. Proteins were transferred to PVDF membranes (0.2 μm pore size, Bio‐Rad, USA) using semi‐dry transfer at 15 V for 45 min.
Immunodetection
2.10.4
Membranes were blocked with 5% non‐fat milk in TBST (Tris‐buffered saline with 0.1% Tween‐20) for 1 h at room temperature. Primary antibody (anti‐bacteriocin peptide, 1:1000 dilution) was incubated overnight at 4°C with gentle shaking. After washing with TBST (3 × 10 min), membranes were incubated with HRP‐conjugated goat anti‐rabbit IgG secondary antibody (1:5000 dilution, Cell Signaling Technology, USA) for 1 h at room temperature. Protein bands were visualized using enhanced chemiluminescence (ECL) substrate (Pierce, USA) on a ChemiDoc MP Imaging System (Bio‐Rad, USA). Full Western blot images with molecular weight markers are shown in Figure S3.
Positive controls included purified commercially available plantaricin, and negative controls included cell‐free supernatants from L. plantarum WCFS1 (non‐bacteriocin producing strain).
Biopreservation Application Testing
2.11
Food Model System Preparation
2.11.1
A traditional African porridge model system was prepared following traditional recipes with modifications for laboratory standardization. The system consisted of pearl millet flour (200 g; locally sourced), distilled water (800 mL), and refined palm oil (20 mL; commercial grade). The mixture was cooked at 100°C for 15 min with continuous stirring, cooled to room temperature under sterile conditions, and divided into 50 mL sterile portions for biopreservation testing.
Biopreservation Efficacy Evaluation
2.11.2
Selected LAB strains showing highest antimicrobial activity were cultured in MRS broth, harvested by centrifugation, washed twice with sterile saline, and resuspended to achieve final concentrations of 10^6^, 10^7^, and 10^8^ CFU/mL in the porridge model. The system was artificially contaminated with a cocktail of pathogenic bacteria prepared by mixing equal volumes of overnight cultures adjusted to 10^5^ CFU/mL each of E. coli O157:H7, S. Typhimurium , and L. monocytogenes .
Final pathogen concentration in the model system was approximately 10^4^ CFU/mL. Samples were stored at ambient temperature (28°C ± 2°C) representing typical tropical storage conditions and analyzed for pathogen reduction, pH changes, and sensory properties at 0, 12, 24, 48, and 72 h. Microbiological analysis was performed using selective media: Sorbitol MacConkey Agar for E. coli O157:H7, Xylose Lysine Deoxycholate Agar for Salmonella, and PALCAM Agar for Listeria.
Whole Genome Sequencing and Analysis
2.12
High‐molecular‐weight DNA from the most promising strain (DE‐LAB‐23) was extracted using the MasterPure DNA Purification Kit (Lucigen, USA). DNA quality and quantity were assessed using a NanoDrop spectrophotometer and Qubit fluorometer (Thermo Fisher Scientific, USA). Whole genome sequencing was performed using the Oxford Nanopore MinION platform with library preparation using the SQK‐LSK109 sequencing kit following the manufacturer's protocols.
Basecalling was performed using Guppy v5.0.11, and genome assembly was conducted using Flye v2.9 with default parameters for bacterial genomes. Assembly quality was assessed using QUAST v5.0.2 and contamination screening was performed using CheckM v1.1.3. Genome annotation was performed using PROKKA v1.14.6 with a genus‐specific database for Lactobacillus.
Biosynthetic gene clusters were identified using antiSMASH 6.0 with relaxed detection strictness to identify novel clusters. Comparative genomics was performed using OrthoFinder v2.5.4 with publicly available L. plantarum genomes from the NCBI database. Phylogenetic analysis of core genes was conducted using RAxML v8.2.12 with 100 bootstrap replications. Detailed phylogenetic analysis of bacteriocin gene clusters is provided in Figure S4.
Statistical Analysis
2.13
All experiments were performed in triplicate with independent biological replicates, and data are presented as mean ± standard deviation. Statistical analysis was conducted using SPSS version 28.0 (IBM, USA) with one‐way analysis of variance (ANOVA) followed by Tukey's honestly significant difference (HSD) post hoc test for multiple comparisons. Significance was set at p < 0.05. Principal component analysis (PCA) was performed using R software (version 4.2.0) to identify relationships between antimicrobial activity and environmental factors. Correlation analysis was conducted using Pearson correlation coefficients with 95% confidence intervals.
Results
3
Isolation and Identification of Lactic Acid Bacteria
3.1
A total of 45 LAB isolates were obtained from fermented D. edulis samples across the three agroecological zones. LAB counts ranged from 7.2 ± 0.3 to 8.9 ± 0.4 log CFU/g, with the highest populations observed in samples from the Forest‐Savanna Transition zone (Tables 1 and 2).
TABLE 1: Distribution and enumeration of lactic acid bacteria from fermented Dacryodes edulis samples.
Molecular identification revealed five distinct LAB species (Table 3). Lactiplantibacillus plantarum was the most prevalent (42.2%), followed by Limosilactobacillus fermentum (26.7%) and Lacticaseibacillus paracasei (17.8%). Novel strain DE‐LAB‐23, identified as L. plantarum , showed 98.9% similarity to the closest GenBank sequence but possessed unique phenotypic characteristics. Phylogenetic analysis based on 16S rRNA sequences is presented in Figure 3, showing the five distinct LAB species clusters with strain DE‐LAB‐23 positioned within the L. plantarum clade.
TABLE 3: Molecular identification and diversity of LAB isolates from fermented Dacryodes edulis .
Neighbor‐joining phylogenetic tree based on 16S rRNA gene sequences of LAB isolates from fermented Dacryodes edulis seeds. Bootstrap values (1000 replications) are shown at branch points. The tree shows five distinct species clusters, with strain DE‐LAB‐23 highlighted within the L. plantarum clade.
Antimicrobial Activity Screening
3.2
Significant variation in antimicrobial activity was observed among isolates (Table 4). Strain DE‐LAB‐23 ( L. plantarum ) demonstrated the highest activity against all tested pathogens, with particularly strong inhibition of E. coli O157:H7 (28.4 ± 1.2 mm) and S. Typhimurium (26.8 ± 0.9 mm). The antimicrobial spectrum varied among species, with L. plantarum strains showing broader activity compared to other species.
Characterization of Antimicrobial Compounds
3.3
The antimicrobial activity of strain DE‐LAB‐23 was completely abolished by proteinase K and pepsin treatment, indicating protein‐based antimicrobial compounds (Table 5). Catalase treatment had no effect, ruling out hydrogen peroxide as the primary antimicrobial agent. The compound demonstrated exceptional thermal stability, retaining 85% activity after heating at 100°C for 15 min, and remained active across a wide pH range (3.0–9.0).
Transcriptional and Proteomic Validation of Novel Bacteriocin Production
3.4
RNA‐Seq Analysis Confirms Gene Expression
3.4.1
Transcriptomic analysis revealed significant upregulation of all genes within the novel bacteriocin gene cluster (BGC‐1) during exponential and stationary growth phases. Complete RNA‐seq differential expression analysis results are provided in Table S2. The structural bacteriocin gene showed 127‐fold upregulation at 18 h compared to early exponential phase (6 h) (adjusted p < 0.001, log_2_FC = 6.99). Associated immunity and transport genes were co‐expressed with similar expression patterns (Figure 4A).
RNA‐seq expression profiles and RT‐qPCR validation. (A) Heatmap showing temporal expression of bacteriocin gene cluster (BGC‐1) genes at 6, 12, 18, and 24 h. (B) Correlation plot between RNA‐seq and RT‐qPCR data (R 2 = 0.94, p < 0.001). (C) Time‐course showing peak expression at 18–20 h correlating with maximum antimicrobial activity.
RT‐qPCR validation of five key genes showed strong correlation with RNA‐seq data (R ^2^ = 0.94, p < 0.001). Peak expression occurred at 18–20 h of fermentation, correlating with maximum antimicrobial activity in culture supernatants (Figure 4B).
Comparative transcriptomics against L. plantarum WCFS1 revealed 156 uniquely expressed genes in DE‐LAB‐23, including the entire novel BGC‐1 cluster, supporting its distinctive antimicrobial profile.
Mass Spectrometry Identifies Novel Bacteriocin Peptide
3.4.2
LC–MS/MS analysis successfully identified the predicted bacteriocin peptide in cell‐free supernatants with high confidence (> 99% peptide identification probability, Mascot score 78). The detected peptide mass (4287.3 Da) matched the predicted mature peptide after leader sequence cleavage with < 5 ppm mass accuracy.
Tandem MS fragmentation patterns confirmed the amino acid sequence with 85% sequence coverage through b‐ and y‐ion series. Complete MS/MS spectra for the identified bacteriocin peptide are provided in Figure S2. Post‐translational modifications were detected, including N‐terminal formylation and a disulfide bond between Cys‐12 and Cys‐38, consistent with class III bacteriocin structures (Figure 5A).
Mass spectrometry identification and peptide fragmentation. (A) Tandem MS/MS spectrum showing b‐ and y‐ion series with 85% sequence coverage. Parent ion [M + H]+ at m/z 4287.3 is indicated. (B) Label‐free quantification showing bacteriocin abundance increasing from 1.2 × 106 at 12 h to 8.7 × 107 at 24 h. (C) Predicted peptide structure showing N‐terminal formylation and disulfide bridge between Cys‐12 and Cys‐38.
Label‐free quantification showed the bacteriocin peptide was among the top 5 most abundant proteins in culture supernatants, with LFQ intensity values increasing from 1.2 × 10^6^ at 12 h to 8.7 × 10^7^ at 24 h fermentation (Figure 5B). Mass spectrometry peptide identification data with complete fragmentation patterns are provided in Table S3.
Additional antimicrobial peptides predicted from BGC‐2 and BGC‐3 were also detected, though at lower abundances, supporting the multigene approach to antimicrobial compound production in strain DE‐LAB‐23.
Western Blot Confirms Protein Expression
3.4.3
Western blot analysis using custom anti‐bacteriocin antibodies detected a single prominent band at approximately 4.3 kDa in cell‐free supernatants from DE‐LAB‐23, consistent with the predicted mature bacteriocin molecular weight (Figure 6A). Band intensity increased progressively from 12 to 24 h of culture, paralleling antimicrobial activity curves.
Western blot time‐course analysis. (A) Time‐course Western blot (0–24 h) showing progressive increase in band intensity at 4.3 kDa. MW: molecular weight marker; Ctrl+: plantaricin positive control; Ctrl‐: L. plantarum WCFS1 negative control; +PK: proteinase K treated sample. (B) Subcellular localization showing predominant secretion into cell‐free supernatants. (C) Antibody specificity controls confirming proteinaceous nature and strain‐specific production.
The bacteriocin was detected in both supernatant and cell‐associated fractions during mid‐exponential phase, but predominantly in supernatants during stationary phase, indicating active secretion. Time‐course analysis showed maximum production at 18–24 h, confirming optimal harvest timing (Figure 6B).
No immunoreactive bands were observed in control strains ( L. plantarum WCFS1 and uninoculated MRS broth), confirming antibody specificity. Treatment with proteinase K eliminated the signal, validating the proteinaceous nature of the antimicrobial compound (Figure 6C).
Integration of Multi‐Omics Data
3.4.4
Combined analysis of genomic, transcriptomic, and proteomic data provides conclusive evidence for the production and secretion of novel bacteriocins by strain DE‐LAB‐23. The correlation between gene expression levels (RNA‐seq), protein abundance (MS), and antimicrobial activity confirms functional expression of the identified biosynthetic gene clusters.
Biopreservation Efficacy in Food Model System
3.5
Application of strain DE‐LAB‐23 in the traditional porridge model system demonstrated significant biopreservative effects (Table 6). The multi‐modal mechanism of antimicrobial action and the practical application workflow in traditional African porridge are illustrated in Figure 7, showing the integrated antimicrobial strategies and temporal pathogen reduction. At an inoculation level of 10^8^ CFU/mL, the strain achieved a 3.2‐log reduction in total pathogen count within 24 h and extended the shelf‐life by 72 h compared to the control. The pH remained stable (4.2–4.5), and sensory evaluation indicated maintained organoleptic quality throughout the storage period.
Mechanism of action and practical application in traditional food systems. (A) Schematic showing multi‐modal antimicrobial action: bacteriocin production, organic acid secretion, and competitive exclusion. (B) Application workflow in traditional African porridge showing pathogen reduction timeline.
Genomic Analysis and Novel Biosynthetic Gene Clusters
3.6
Whole genome sequencing of strain DE‐LAB‐23 revealed a 3.2 Mb genome with 3187 predicted coding sequences and a G + C content of 44.3% (Table 7). The genomic organization and biosynthetic gene cluster distribution are illustrated in Figure 8, which displays the circular genome map with the four novel biosynthetic gene clusters (BGC‐1 to BGC‐4) highlighted. Comparative genomic analysis identified several unique features distinguishing this strain from other L. plantarum genomes in public databases.
Genomic features and biosynthetic gene clusters in strain DE‐LAB‐23. Circular genome map showing (outer to inner rings): predicted coding sequences (forward and reverse strands), rRNA and tRNA genes, GC content, and GC skew. Four novel biosynthetic gene clusters (BGC‐1 to BGC‐4) are highlighted in red.
antiSMASH analysis revealed four novel biosynthetic gene clusters (BGCs) not previously reported in L. plantarum strains. The most significant cluster (BGC‐1) spans 23.4 kb and contains genes encoding novel class III bacteriocin with unique structural features. BGC‐2 encodes enzymes to produce cyclic lipopeptides with potential antifungal activity. Phylogenetic analysis of core antimicrobial peptide sequences showed clear divergence from known bacteriocins, suggesting horizontal gene transfer events.
Discussion
4
This study represents the first comprehensive investigation of LAB diversity and biopreservative potential in fermented D. edulis seeds, revealing a rich microbial ecosystem harboring strains with exceptional antimicrobial properties. The identification of 45 LAB isolates across five species demonstrates the complexity of this traditional fermentation system and its potential as a source of novel bioactive microorganisms.
LAB Diversity and Ecological Distribution
4.1
The predominance of L. plantarum in our study is consistent with previous research on African fermented foods, including spontaneous fermentation of African nightshade leaves where L. plantarum was also the dominant species (Fissehaye et al. 2025). Recent research by Kouadio et al. (2024) similarly reported L. plantarum dominance in West African fermented foods, attributing this to superior acid tolerance and metabolic versatility under tropical conditions. In contrast, Hussuwa, a Sudanese fermented sorghum food, was reported to be dominated by Lactobacillus fermentum and Pediococcus acidilactici , highlighting the variability in lactic acid bacteria composition across African fermented foods (Yousif et al. 2010).
Exceptional Antimicrobial Activity
4.2
The exceptional antimicrobial activity of DE‐LAB‐23 against multiple foodborne pathogens, particularly the 28.4 mm inhibition zone against E. coli O157:H7, exceeds previously reported activities for Lactobacillus spp. from locally fermented Nono in Northern Nigeria. In that study, crude bacteriocins exhibited inhibition zones ranging from 9.5 to 20 mm against E. coli and 9.47–18.3 mm against S. aureus (Subedi et al. 2025). This comparison underscores the unusually high potency of DE‐LAB‐23 and highlights its potential as a superior biopreservative for enhancing food safety in traditional African foods (Ruqayyah 2023).
Novel Bacteriocin Characteristics
4.3
The proteinaceous nature of the antimicrobial compounds was confirmed by their complete inactivation by proteolytic enzymes, indicating that they are indeed bacteriocins. Remarkably, these bacteriocins retained 85% of their activity after exposure to 100°C for 15 min, a thermal stability that surpasses most previously reported bacteriocins, which typically lose activity above 80°C (Chin et al. 2016). This exceptional heat tolerance highlights their potential for applications in food preservation under high‐temperature processing conditions and distinguishes these isolates from conventional LAB‐derived bacteriocins. This thermal stability, combined with broad pH tolerance, makes these compounds particularly suitable for food preservation applications involving thermal processing.
Multi‐Omics Validation
4.4
The novel biosynthetic gene clusters identified in strain DE‐LAB‐23 represent a significant advancement in bacteriocin research. BGC‐1, encoding a putative class III bacteriocin, shows only 47% homology to known sequences, suggesting a completely new antimicrobial peptide family. The presence of unusual post‐translational modification genes within this cluster indicates complex maturation processes that may contribute to the enhanced stability and activity observed.
The multi‐omics approach employed in this study provides unprecedented validation of bacteriocin production. The integration of genomic prediction, transcriptional confirmation, and proteomic detection represents a comprehensive characterization that addresses previous limitations in bacteriocin research where gene presence did not always correlate with functional expression.
Biopreservation Efficacy
4.5
The biopreservation efficacy demonstrated in the traditional porridge model system validates the practical application potential of these indigenous LAB strains. The 3.2‐log reduction in pathogen count within 24 h, combined with shelf‐life extension of 72 h, represents a significant improvement over synthetic preservatives, which often require higher concentrations and may affect food quality.
The 3.2‐log reduction in pathogen load observed for DE‐LAB‐23 in traditional African porridge aligns with previous biopreservation studies, such as the use of semi‐purified bacteriocin BacFL31 in raw ground turkey meat, which effectively inhibited Listeria monocytogenes and Salmonella Typhimurium and extended product shelf life (Chakchouk‐Mtibaa et al. 2017). While the food matrices and storage conditions differ, both studies highlight the potential of bacteriocin‐producing LAB as natural preservatives for enhancing food safety and shelf life.
Our superior performance (14%–31% higher) likely reflects the combined effects of multiple antimicrobial mechanisms: bacteriocin production, organic acid generation, and competitive exclusion. Notably, the 72‐h shelf‐life extension at ambient temperature (28°C ± 2°C) exceeds the 48‐h extension. This enhanced preservation at tropical temperatures addresses critical food safety challenges in sub‐Saharan Africa where refrigeration remains limited. The maintained organoleptic quality throughout the preservation period addresses a critical limitation of many biological preservation systems.
Metabolomic and Genomic Insights Into Multi‐Modal Antimicrobial Activity
4.6
Metabolomic profiling of the L. plantarum strain in this study revealed a unique organic acid profile, including elevated levels of phenyllactic acid (PLA) and hydroxy‐phenyllactic acid (OH‐PLA), which acted synergistically with bacteriocins to produce broad‐spectrum antimicrobial activity against multiple foodborne pathogens (Figure S5). This multi‐modal mechanism suggests that the combined effects of bacteriocins and bioactive metabolites enhance the antimicrobial potency of LAB strains beyond what would be achieved by either component alone.
These findings are consistent with previous studies reporting that LAB strains, including L. plantarum , produce PLA and OH‐PLA, and that metabolite production can be influenced by precursor availability, such as phenylalanine and tyrosine (Valerio et al. 2004). Similarly, research on Carnobacterium maltaromaticum and C. divergens demonstrated that both bacteriocins and organic acids, including formate and acetate, contribute synergistically to antibacterial activity, supporting the concept of multi‐modal mechanisms (Zhang et al. 2019).
Comparative genomic analysis reveals significant divergence from characterized L. plantarum strains. Our identification of 4 novel BGCs in DE‐LAB‐23 represents exceptional biosynthetic diversity, potentially reflecting the unique selective pressures of lipid‐rich D. edulis fermentation environments. The 142 kb genome expansion compared to type strains is remarkable. Detailed genomic comparison with reference L. plantarum strains is presented in Table S4. The genomic analysis revealed horizontal gene transfer events that likely contributed to the unique antimicrobial profile of strain DE‐LAB‐23. The presence of mobile genetic elements flanking novel BGCs suggests recent acquisition of antimicrobial functions, potentially through interactions with other microorganisms in the complex fermentation environment.
Integration With Traditional Knowledge
4.7
The integration of traditional fermentation knowledge with modern biotechnology exemplified in this study represents a paradigm shift in food preservation research. This approach not only validates traditional practices but also unlocks their potential for modern applications, creating bridges between indigenous knowledge systems and contemporary food safety requirements.
Future research directions should focus on large‐scale production optimization, regulatory approval processes, and comprehensive safety assessments. Additionally, investigation of synergistic effects between different indigenous LAB strains may reveal even more effective biopreservation systems.
The potential applications extend beyond traditional foods to modern food processing systems, where natural preservatives are increasingly demanded by health‐conscious consumers. The thermal stability of the antimicrobial compounds makes them suitable for processed foods requiring heat treatment, expanding their commercial applicability.
Climate Change and Food Security
4.8
Climate change adaptation represents another important consideration, as traditional fermentation systems face disruption from changing environmental conditions (Pal 2025). Understanding and preserving the microbial diversity of indigenous fermented foods becomes crucial for maintaining food security and cultural heritage.
Integration With Global Food Safety Initiatives
4.9
Our findings align with recent WHO and FAO initiatives promoting indigenous food system resilience and natural preservation technologies. Park and Mannaa (2025) advocate for functional fermented foods as dual‐purpose products providing both nutrition and food safety benefits. Strain DE‐LAB‐23 exemplifies this concept, offering biopreservation while maintaining the traditional organoleptic profile valued in West African cuisines.
The demonstrated efficacy against multiple antibiotic‐resistant pathogens addresses growing concerns about antimicrobial resistance in food systems. Widespread adoption of bacteriocin‐based interventions has the potential to significantly reduce foodborne antimicrobial‐resistant infections, highlighting the public health importance of potent bacteriocin‐producing strains (Singh et al. 2025) such as DE‐LAB‐23.
Conclusion
5
This study establishes fermented D. edulis seeds as a reservoir of novel LAB strains with exceptional biopreservative potential. Strain DE‐LAB‐23 ( L. plantarum ) demonstrated superior antimicrobial activity against major foodborne pathogens, producing thermostable, protein‐based antimicrobial compounds with broad‐spectrum efficacy. Genomic analysis revealed unique biosynthetic gene clusters encoding novel antimicrobial peptides, representing significant additions to the bacteriocin knowledge base. Multi‐omics validation (RNA‐seq, mass spectrometry, Western blot) provided conclusive evidence for functional expression of these novel compounds. The successful application in traditional food systems validates the practical potential of these indigenous strains for sustainable food preservation. These findings contribute to food safety, economic development, and cultural preservation objectives while demonstrating the value of exploring traditional fermentation systems for biotechnological applications. The integration of indigenous knowledge with modern biotechnology offers promising pathways for addressing global food safety challenges through environmentally sustainable and culturally appropriate solutions.
Author Contributions
Zakari Adeiza David: conceptualization, methodology, software, data curation, supervision, project administration, visualization, writing – review and editing, writing – original draft, investigation. Muhammad Farhan Nasir: supervision, writing – review and editing, writing – original draft. Syed Parween Ali: validation, methodology, data curation. E. Joel Mart: methodology, formal analysis. Sara Zahid: methodology, formal analysis, project administration. Adefila Moyosore Adebimpe: conceptualization, validation, methodology, writing – review and editing, writing – original draft. Humara Adnan: conceptualization, methodology, formal analysis. Samandarov Abrorbek Islomboyevich: validation, formal analysis, supervision. Mukhayya Ruzieva: methodology, formal analysis, supervision.
Funding
The authors have nothing to report.
Ethics Statement
The authors have nothing to report.
Consent
The authors have nothing to report.
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Table S1: RT‐qPCR primer sequences for bacteriocin gene validation. Table S2: Complete RNA‐seq differential expression analysis results. Table S3: Mass spectrometry peptide identification data with fragmentation patterns. Table S4: Detailed genomic comparison with reference L. plantarum strains.
Figure S1: Quality control metrics for RNA‐seq analysis. Figure S2: Complete MS/MS spectra for identified bacteriocin peptide. Figure S3: Full Western blot images with molecular weight markers. Figure S4: Phylogenetic analysis of bacteriocin gene clusters. Figure S5: Metabolomic profiling of organic acids and secondary metabolites.
File S1: Complete genome assembly and annotation for strain DE‐LAB‐23 (available in NCBI GenBank). File S2: Raw RNA‐seq data (available in NCBI SRA). File S3: Mass spectrometry raw data files (available in ProteomeXchange).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abubakr, M. A. , and W. M. Al‐Adiwish . 2017. “Isolation and Identification of Lactic Acid Bacteria From Different Fruits With Proteolytic Activity.” International Journal of Microbiology and Biotechnology 2, no. 2: 58–64.
- 2Chakchouk‐Mtibaa, A. , S. Smaoui , N. Ktari , et al. 2017. “Biopreservative Efficacy of Bacteriocin Bac FL 31 in Raw Ground Turkey Meat in Terms of Microbiological, Physicochemical, and Sensory Qualities.” Biocontrol Science 22, no. 2: 67–77.28659558 10.4265/bio.22.67 · doi ↗ · pubmed ↗
- 3Chauhan, K. , and A. Rao . 2024. “Clean‐Label Alternatives for Food Preservation: An Emerging Trend.” Heliyon 10, no. 16: e 35815.39247286 10.1016/j.heliyon.2024.e 35815 PMC 11379619 · doi ↗ · pubmed ↗
- 4Chen, X. , H. Bai , W. Mo , et al. 2025. “Lactic Acid Bacteria Bacteriocins: Safe and Effective Antimicrobial Agents.” International Journal of Molecular Sciences 26, no. 9: 4124.40362364 10.3390/ijms 26094124 PMC 12071495 · doi ↗ · pubmed ↗
- 5Chin, Y.‐Z. , S. Velu , and M. A. R. Nor‐Khaizura . 2016. “Characterization and the Influence of Milk Solids‐Not‐Fat on the Bacteriocin Produced by Lactococcus lactis subsp. Lactis S 20 Isolated From Chinese Sauerkraut, a Traditional Fermented Vegetable.” Annals of Microbiology 66, no. 2: 673–684.
- 6Drancourt, M. , C. Bollet , A. Carlioz , R. Martelin , J. P. Gayral , and D. Raoult . 2000. “16S Ribosomal DNA Sequence Analysis of a Large Collection of Environmental and Clinical Unidentifiable Bacterial Isolates.” Journal of Clinical Microbiology 38, no. 10: 3623–3630.11015374 10.1128/jcm.38.10.3623-3630.2000 PMC 87447 · doi ↗ · pubmed ↗
- 7Fissehaye, N. A. , S. Imathiu , P. K. Kinyanjui , and E. Wafula . 2025. “Identification and Characterization of Lactic Acid Bacteria Isolated From Spontaneously Fermented African Nightshade ( Solanum scabrum ) Leaves.” Journal of Microbiology, Biotechnology and Food Sciences 15, no. 2: e 12816.
- 8Hu, D. , C. Sun , T. Jin , et al. 2020. “Exploring the Potential of Antibiotic Production From Rare Actinobacteria by Whole‐Genome Sequencing and Guided MS/MS Analysis.” Frontiers in Microbiology 11: 1540.32922368 10.3389/fmicb.2020.01540 PMC 7375171 · doi ↗ · pubmed ↗