Two new species of Pseudocosmospora (Hypocreales) revealed through morphological and phylogenetic analyses

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
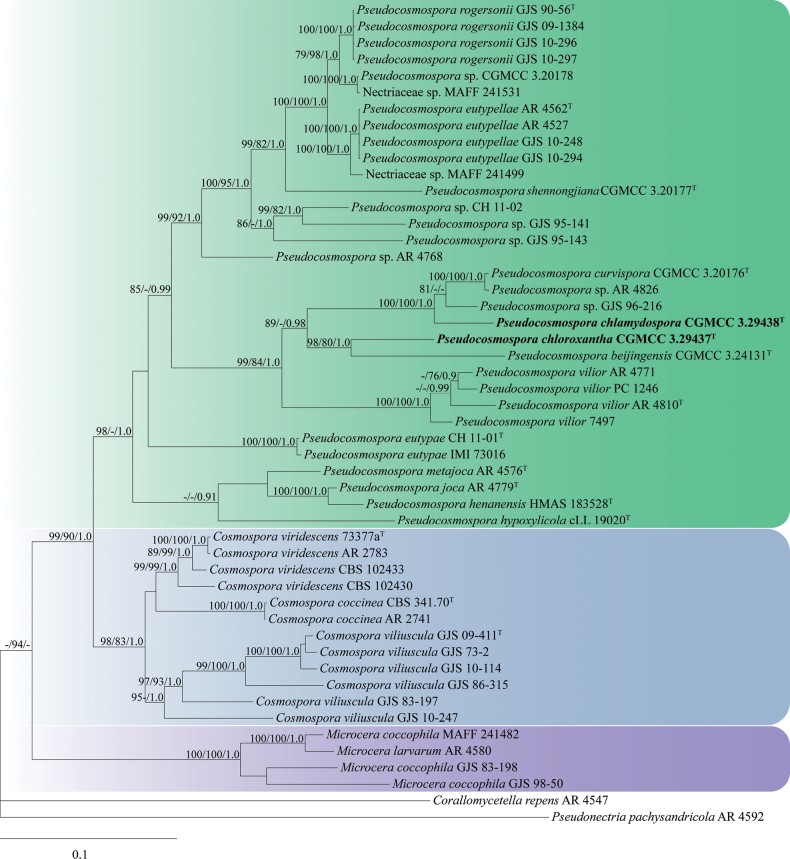 Figure 1
Figure 1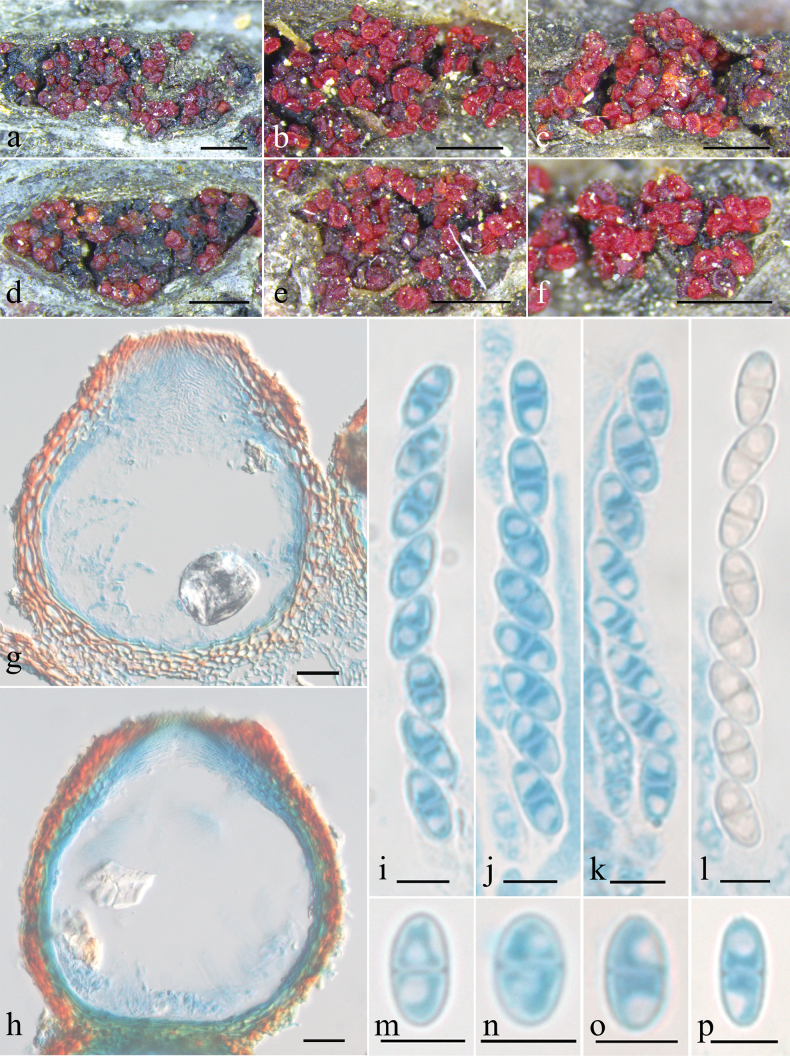 Figure 2
Figure 2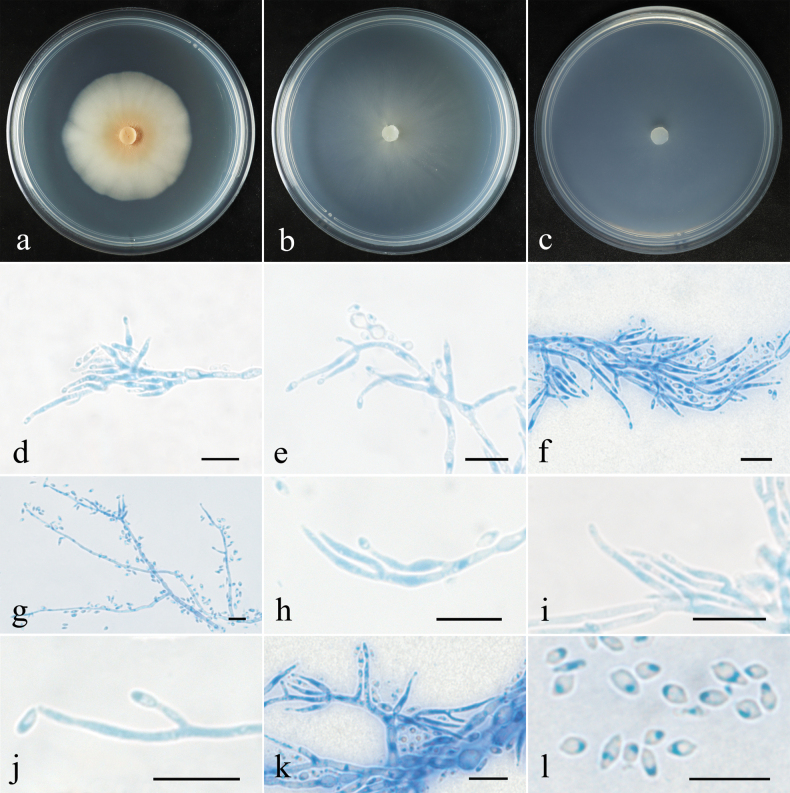 Figure 3
Figure 3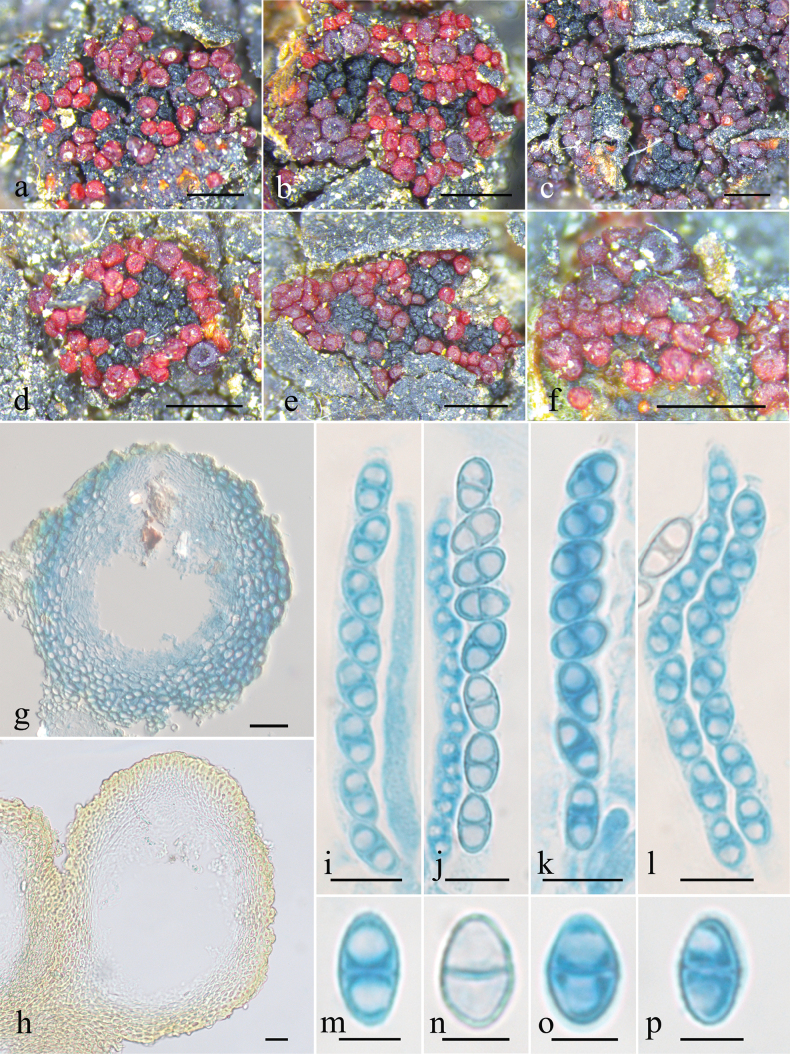 Figure 4
Figure 4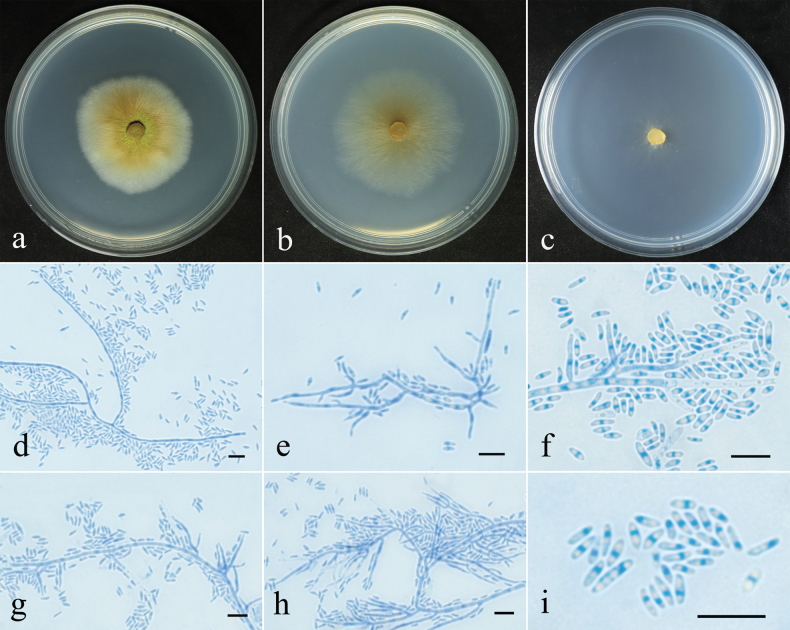 Figure 5
Figure 5| Species | Herbarium/Strain Number | GenBank Accession Numbers | References | ||||
|---|---|---|---|---|---|---|---|
| ITS |
|
|
|
| |||
|
| AR 4547 |
|
|
|
|
|
|
|
| CBS 341.70T |
|
|
|
|
| |
| AR 2741 |
|
|
|
|
| ||
|
| GJS 10-247 |
|
|
|
|
| Schoch |
| GJS 83-197 |
|
|
|
|
| ||
| GJS 86-315 |
|
|
|
|
| ||
| GJS 09-411 T |
|
|
|
|
| ||
| GJS 10-114 |
|
|
|
| – | ||
| GJS 73-2 |
|
|
|
|
| ||
|
| IMI 73377aT |
|
|
|
|
| |
| AR 2783 |
|
|
|
|
| ||
| CBS 102430 |
|
|
|
|
| ||
| CBS 102433 |
|
|
|
|
| ||
|
| MAFF 241482 |
|
|
|
|
|
|
| GJS 83-198 |
|
| – |
|
| ||
| GJS 98-50 |
|
|
|
|
| ||
|
| AR 4580 |
|
|
|
|
|
|
| MAFF 241499 |
|
|
|
|
|
| |
| MAFF 241531 |
|
|
|
|
|
| |
|
|
|
|
| – | – |
| |
|
|
|
|
|
|
| This study | |
|
|
|
|
|
|
| This study | |
|
|
|
|
|
| – |
| |
|
| CH 11-01T |
|
|
|
|
|
|
| IMI 73016 |
|
| – |
|
| ||
|
| AR 4562T |
|
|
|
|
|
|
| AR 4527 |
|
|
|
|
| ||
| GJS 10-248 |
|
|
|
|
| ||
| GJS 10-294 |
|
|
|
|
| ||
|
|
|
|
| – |
| ||
|
| cLL 19020T |
|
| – | – | – |
|
|
| AR 4779T |
|
|
|
| – |
|
|
| AR 4576T |
|
|
|
|
|
|
|
| GJS 90-56T |
|
|
|
|
|
|
| GJS 10-296 |
|
|
|
|
| ||
| GJS 09-1384 |
|
|
|
|
| ||
| GJS 10-297 |
|
|
|
|
| ||
|
|
|
|
|
| – |
| |
|
| AR 4810T |
|
|
|
|
|
|
| AR 4771 |
|
|
|
|
|
| |
| PC 1246 |
|
|
|
|
| ||
| 7497 |
|
|
| – |
| ||
| AR 4826 |
|
|
|
|
|
| |
| GJS 96-216 |
|
|
|
| – |
| |
| AR 4768 |
|
|
|
| – |
| |
| GJS 95-141 |
|
|
|
|
|
| |
| CH 11-02 |
|
|
|
|
|
| |
|
|
|
|
| – |
| ||
| GJS 95-143 |
|
|
|
|
|
| |
|
| AR 4592 |
|
|
|
|
|
|
| Sequence region | No. of seq. | Length of alignment (bp) | No. of variable sites | No. of parsimony-informative sites |
|---|---|---|---|---|
| ITS | 50 | 538 | 147 | 78 |
|
| 50 | 785 | 88 | 52 |
|
| 47 | 602 | 296 | 242 |
|
| 46 | 626 | 298 | 245 |
|
| 41 | 248 | 223 | 196 |
| ITS+ | 50 | 2,799 | 1,052 | 813 |
| Sequences region | ITS (bp) | ||||
|---|---|---|---|---|---|
| Comparing species | |||||
| 8 | 12 | 81 | 102 | 103 | |
| 6 | 4 | 36 | 30 | – | |
| 15 | 4 | 81 | 98 | 101 | |
| 10 | 8 | 70 | 82 | 84 | |
| 33 | 17 | 55 | – | – |
| Species | Perithecia size (µm) | Asci size (µm) | Ascospores size (µm) | Conidia shape | Conidia size (µm) | References |
|---|---|---|---|---|---|---|
|
| 147–196 × 118–176 | 38–58 × 2.5–5 | 8–10 × 2.5–4 | allantoid, curved | 2.6–4.5 × 0.9–1.8 |
|
|
| 144–217 × 101–185 | 42.7–56.7 × 3.4–5.7 | 5.9–8.8 × 3–4.7 | rod-shaped, fusiform, ellipsoidal | 2.5–5.7 × 1.3–2.7 | This study |
|
| 132–230 × 127–210 | 53–76.8 × 4.7–9.2 | 7.1–11.5 × 3.7–6.3 | rod-shaped to cylindrical | 2.6–8.5 × 1.2–2.4 | This study |
|
| 167–235 × 108–167 | 53–68 × 3–5 | 8–10 × 3–5 | allantoid, mainly strongly curved | 3–5 × 0.8–1.2 |
|
|
| 163–245 × 131–180 | 54–69 × 5.7–8.4 | 7.9–12.2 × 3.3–4.9 | oblong to ellipsoidal | 2.9–5.5 × 1.1–2.6 |
|
- —Foundation for Innovative Research Groups of the National Natural Science Foundation of China 501100012659 http://doi.org/10.13039/501100012659
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFungal Biology and Applications · Plant Pathogens and Fungal Diseases · Mycorrhizal Fungi and Plant Interactions
Introduction
Historically, the nectroid fungi with small, reddish, KOH+, smooth and thin-walled perithecia were placed in Cosmospora sensu lato (Samuels et al. 1991; Rossman et al. 1999). However, the phylogenetic analysis showed that the genus was heterogeneous (Zhang and Zhuang 2006). Efforts were made continuously towards establishment of a monophyletic Cosmospora Rabenh. as well as its allies (Luo and Zhuang 2008, 2010; Samuels et al. 2009; Gräfenhan et al. 2011; Schroers et al. 2011). As part of this taxonomic revision, Herrera et al. (2013) established the genus Pseudocosmospora C.S. Herrera & P. Chaverri, with P. eutypellae C.S. Herrera & P. Chaverri as the type species, to accommodate C. vilior (Starbäck) Rossman & Samuels and the related taxa occur on Eutypa Tul. & C. Tul., Eutypella (Nitschke) Sacc., and Biscogniauxia Kuntze.
Members of Pseudocosmospora are usually distributed in tropical and temperate regions and occur on fruitbodies of diatrypaceous and xylariaceous fungi. Some species have significant importance in agriculture and biomedicine due to their capability of producing bioactive secondary metabolites (Lee et al. 2011; Hsiao et al. 2014; Shiono et al. 2016; Nakamura et al. 2019). Morphologically, Pseudocosmospora species typically have solitary to gregarious, scarlet, subglobose to obpyriform, blunt-papillate perithecia that collapse laterally when dry and mostly less than 250 μm high; they possess cylindrical to narrowly clavate asci with eight ellipsoidal, 1-septate ascospores and form acremonium- to verticillium-like asexual states with cylindrical phialides that produce ellipsoidal, ovoid or reniform, aseptate conidia (Herrera et al. 2013; Zeng and Zhuang 2021). Currently, there are 17 globally accepted species in the genus, 12 of them have been reported in China (Herrera et al. 2013; Zeng and Zhuang 2017, 2021, 2022, 2023).
During the examinations of hypocrealean fungi collected from Beijing, China, two interesting fungicolous specimens were encountered. Morphological observations and phylogenetic analyses based on combined nuclear ribosomal DNA ITS1-5.8S-ITS2 (ITS), large subunit of nuclear ribosomal DNA (LSU), β-tubulin (tub2), RNA polymerase II subunit one (rpb1) and translation elongation factor 1-α (tef1-α) sequence data determined their taxonomic position in Pseudocosmospora and are new to the genus. Morphological differences between these novel taxa and their close relatives are compared in detail.
Materials and methods
Sample collection and strains isolation
Fresh specimens were collected from Baili Landscape Gallery in Yanqing District and Miyun Reservoir area in Miyun District. The collection sites were located in mixed forest vegetation. All specimens were preserved in Herbarium Mycologicum Academiae Sinicae (HMAS). Cultures were obtained through single ascospore isolation: fresh perithecia were surface-sterilized by immersion in 75% ethanol for 1 min, followed by three rinses with sterile water, then crushed with tweezers in 200 µL of sterile water to release the ascospores; and finally the spore suspension was spread evenly onto potato dextrose agar (PDA) medium. After 2–7 days of incubation, colonies were subcultured on fresh PDA. All strains were deposited in the China General Microbiological Culture Collection Center (CGMCC).
Morphological characterization
The methods of Zeng and Zhuang (2022) were generally followed for morphological observations. Strains were cultured on three different media: PDA, cornmeal dextrose agar (CMD), and synthetic low-nutrient agar (SNA), and the plates were incubated under a 12/12 h light/dark photoperiod for 28 d at 25 °C. Colonies were photographed and diameters were measured. Photographs of macroscopic features were taken with a Leica DFC450 digital camera (Wetzlar, Germany) attached to a Leica M125 stereomicroscope (Milton Keynes, UK). Perithecial wall reactions were observed with 3% KOH and 100% lactic acid. Perithecia sections (ca. 6–8 μm thick) were made using a freezing microtome (YD-1508-III, Jinhua, China). The anatomic characteristics of perithecia, asci, ascospores, conidiophores, conidia and chlamydospores were examined, photographed, and measured using a Zeiss AxioCam MRc 5 digital camera (Jena, Germany) attached to a Zeiss Axio Imager A2 microscope (Göttingen, Germany).
DNA extraction, PCR amplification, and sequencing
Genomic DNA was extracted from fresh mycelia of Pseudocosmospora species and the fruitbodies of their hosts using the protocol of Wang and Zhuang (2004). Five loci, namely ITS, LSU, tub2, rpb1 and tef1-α, were amplified and sequenced with the primer pairs ITS5/ITS4 (White et al. 1990), LR0R/LR5 (Vilgalys and Hester 1990), and T1/T22 (O’Donnell and Cigelnik 1997), RPB1-Af/RPB1-Cr (Castlebury et al. 2004) and EF1-728F/EF2 (O’Donnell et al. 1998), respectively.
Phylogenetic analyses
The sequences of Pseudocosmospora species along with other related species were downloaded from GenBank and the accession numbers are listed in Table 1. Sequences were assembled, aligned and trimmed by BioEdit 7.0.5 (Hall 1999), and converted using Clustal X 1.83 (Thompson et al. 1997). Phylogenetic analyses of multiple genes were conducted using the maximum likelihood (ML), Bayesian inference (BI), and maximum parsimony (MP) methods. ML analysis was determined using RAxML with the default GTRCAT model (Samatakis 2006). BI analysis was performed with MrBayes 3.1.2 using a Markov chain Monte Carlo algorithm (Ronquist and Huelsenbeck 2003). For combined sequence dataset, GTR+I+G was selected as the best-fit model using MrModeltest 2.3 (Nylander 2004), and was further used in the BI analysis. Four Markov chains were run simultaneously for 1,000,000 generations with the trees sampled every 100 generations. The first 25% of trees were set as burn-in and discarded. The remaining trees were used to calculate the Bayesian inference posterior probability (BIPP) values. The MP analysis was performed with PAUP 4.0b10, using heuristic searches with 1,000 replicates of random addition of sequences and subsequent TBR (tree bisection and reconnection) branch swapping (Swofford 2002). Topological confidence of resulted trees was tested by maximum parsimony bootstrap proportion (MPBP) with 1,000 replications, each with 10 replicates of random addition of taxa. Trees were visualized by TreeView 1.6.6 (Page 1996). Maximum likelihood bootstrap proportion (MLBP) and maximum parsimony bootstrap proportion (MPBP) ≥ 75%, and Bayesian inference posterior probability (BIPP) ≥ 0.9 are shown at the nodes.
Results
Multi-gene phylogeny
The sequences of ITS, LSU, tub2, rpb1 and tef1-α from 50 strains of 29 species were analyzed. The concatenated sequence matrix was composed of 2,799 characters (ITS: 1–538, LSU: 539–1,323, tub2: 1,324–1,925, rpb1: 1,926–2,551, and tef1-α: 2,552–2,799) (Table 2). Corallomycetella repens (AR 4547) and Pseudonectria pachysandricola (AR 4592) were used as outgroup taxa. The BI tree was shown in Fig. 1. The topologies of the ML and MP trees were similar to that of the BI tree. The isolates CGMCC 3.29437 and 3.29438 were located among species of Pseudocosmospora, and received high statistical support values (MLBP/MPBP/BIPP = 98%/-/1.0). The isolate CGMCC 3.29437 grouped together with P. beijingensis (CGMCC 3.24131) (MLBP/MPBP/BIPP = 98%/80%/1.0), and the isolate CGMCC 3.29438 clustered with P. curvispora (CGMCC 3.20176), Pseudocosmospora sp. GJS 96-216 and AR 4826 (MLBP/MPBP/BIPP = 100%/100%/1.0). These six species formed a well-supported cluster (MLBP/BIPP = 89%/0.98), and were associated with P. vilior (MLBP/MPBP/BIPP = 99%/84%/1.0).
The Bayesian inference tree inferred from combined ITS, LSU, tub2, rpb1 and tef1-α sequences of Pseudocosmospora species and other related species. MLBP (left) and MPBP (middle) (≥ 75%) and BIPP (right) (≥ 0.9) were shown at the nodes.
Taxonomy
Pseudocosmospora
chlamydospora
Taxon classificationFungiNectriaceae
Xiao Qian Wu, Z.Q. Zeng & W.Y. Zhuang sp. nov.
AAF66A27-4F56-589E-AC6B-DB21B8C1F5AF
Fungal Names: FN 573083
Etymology.
The specific epithet refers to the chlamydospores produced in culture.
Typification.
China • Beijing City, Miyun District, roadside near Miyun Reservoir, on fruitbodies of Paraeutypella karsti growing on rotten bark, 8 September 2022, W.Y. Zhuang, Z.Q. Zeng & H.D. Zheng, 13067 (holotype HMAS 255842, ex-type culture CGMCC 3.29438).
Description.
Mycelium not visible on natural substratum. Perithecia superficial, solitary to gregarious, non-stromatic or with a basal stroma, subglobose to globose with a truncate apex, laterally collapsed upon drying, orange-red to bright red, turning dark red in 3% KOH, becoming light yellow in 100% LA, 144–217 × 101–185 μm. Perithecial walls of two layers, 17–28 μm thick; the outer layer of textura angularis, 10–21 μm thick, cells 2.5–8 × 1.6–4.1 μm, walls 0.9–1.4 μm thick; the inner layer of textura prismatica, 5–10 μm thick, cells 4.7–13.2 × 1.6–3.3 μm, walls 0.8–1.3 μm thick. Asci clavate, with a simple apex, eight-spored, 42.7–56.7 × 3.4–5.7 μm. Ascospores ellipsoidal, 1-septate, hyaline to light yellow-brown, smooth-walled, uniseriate, overlapping obliquely, 5.9–8.8 × 3–4.7 μm.
Macroscopic and microscopic features of Pseudocosmospora chlamydospora (HMAS 255842). a–f. Perithecia on natural substratum; g, h. Longitudinal section through perithecium; i–l. Asci with ascospores; m–p. Ascospore. Scale bars: 500 µm (a–f); 20 µm (g, h); 5 µm (i–p).
Colony characteristics.
Colony on PDA 43 mm diam. after 28 d at 25 °C, crustose, beige, light yellow to salmon-pink, on SNA 44 mm in diam. after 28 d at 25 °C, with sparse whitish aerial mycelium. Conidiophores acremonium- to verticillium-like, septate, of indefinite length, hyaline, with 1–2 whorls and a terminal whorl of 2–4 phialides. Phialides subulate, tapering toward the apex, 5–24 μm long, 1.1–1.9 μm wide at the base, 0.6–1.2 μm wide at the tip. Conidia rod-shaped, fusiform, ellipsoidal, lemon-shaped or subglobose, rare clavate, unicellular, smooth, hyaline, non-guttulate, 2.5–5.7 × 1.3–2.7 μm. Chlamydospores globose to subglobose, rarely oblong, smooth-walled, hyaline, 3.4–8.4 μm in diam.
Colonial and microscopic features of Pseudocosmospora chlamydospora (CGMCC 3.29438). a–c. Colony after 28 d at 25 °C on PDA, CMD, SNA; d–j. Conidiophores and conidia; k. Conidiophores, chlamydospores and conidia; l. Conidia. Scale bars: 10 µm (d–l).
Notes.
Amongst the known species of Pseudocosmospora, P. chlamydospora most resembles P. rogersonii in similar host fungus, subglobose perithecia with a truncate apex, ellipsoidal and smooth ascospores, and pink colony on PDA (Herrera et al. 2013). However, the new species differs in having smaller asci (42.7–56.7 × 3.4–5.7 μm vs. 54–69 × 5.7–8.4 μm), shorter ascospores (5.9–8.8 μm vs. 7.9–12.2 μm), and producing globose to subglobose chlamydospores. Sequence comparisons revealed that there were 8 bp, 12 bp, 81 bp, 102 bp and 103 bp divergences detected for the ITS, LSU, tub2, rpb1 and tef1-α regions between P. chlamydospora and P. rogersonii (Table 3). The multi-gene phylogenetic analyses showed that P. chlamydospora is closely related to P. curvispora and two unnamed species (MLBS/MPBP/BIPP = 100%/100%/1.0). However, P. chlamydospora can be easily distinguished by its shorter asci (42.7–56.7 µm vs. 53–68 µm) and ascospores (5.9–8.8 µm vs. 8–10 µm), wider conidia (1.3–2.7 µm vs. 0.8–1.2 µm), and forming globose to subglobose chlamydospores (Zeng and Zhuang 2021) (Table 4). Thus, both the morphological feature and sequence data indicated that they represent different taxa.
Pseudocosmospora
chloroxantha
Taxon classificationFungiNectriaceae
Xiao Qian Wu, Z.Q. Zeng & W.Y. Zhuang sp. nov.
F1C42434-0A94-5684-BD48-D3D82856505E
Fungal Names: FN 573084
Etymology.
The specific epithet refers to the greenish yellow pigments on surface of colony.
Typification.
China • Beijing City, Yanqing District, Baili Landscape Gallery, on Paraeutypella karsti growing on rotten bark, 31 August 2022, W.Y. Zhuang, Z.Q. Zeng & H.D. Zheng, 13057 (holotype HMAS 255843, ex-type culture CGMCC 3.29437).
Macroscopic and microscopic features of Pseudocosmospora chloroxantha (HMAS 255843). a–f. Perithecia on natural substratum; g, h. Longitudinal section through perithecium; i–l. Asci with ascospores; m–p. Ascospore. Scale bars: 500 µm (a–f); 20 µm (g, h); 10 µm (i–l); 5 µm (m–p).
Description.
Mycelium not visible on the natural substratum. Perithecia superficial and gregarious, non-stromatic or with a basal stroma, subglobose to globose with a truncate apex, laterally collapsed upon drying, bright red to brownish red, turning dark red in 3% KOH, becoming light yellow in 100% LA, 132–230 × 127–210 μm. Perithecial walls of two-layers, 19–37 μm thick; the outer layer of textura globulosa to t. angularis, 11–24 μm thick, cells 2.8–11.7 × 2–5.5 μm, walls 0.8–1.4 μm thick; the inner layer of textura prismatica, 9–15 μm thick, cells 5.9–14.5 × 1.6–3.1 μm, walls 0.8–1.7 μm thick. Asci cylindrical to clavate, with a simple apex or with an apical ring, eight-spored, 53–76.8 × 4.7–9.2 μm. Ascospores ellipsoidal to broad ellipsoidal, 1-septate, hyaline to light yellow-brown, smooth-walled, uniseriate, overlapping obliquely, 7.1–11.5 × 3.7–6.3 μm.
Colonial and microscopic features of Pseudocosmospora chloroxantha (CGMCC 3.29437). a–c. Colony after 28 d at 25 °C on PDA, CMD, SNA; d–h. Conidiophores and conidia; i. Conidia. Scale bars: 10 µm (d–i).
Colony characteristics.
Colony on PDA 46 mm in diam. after 28 d at 25 °C, crustose, surface greenish yellow and reverse yellowish-brown, on SNA 42 mm in diam. after 28 d at 25 °C, with sparse whitish aerial mycelium. Conidiophores acremonium- to verticillium-like, septate, of indefinite length, and hyaline, with 1–2 whorls and a terminal whorl of 2–4 phialides. Phialides subulate, tapering toward the apex, 9.6–31.1 μm long, 0.9–2 μm wide at the base, 0.8–1.4 μm wide at the tip. Conidia rod-shaped to cylindrical, rarely ellipsoidal, unicellular, smooth-walled, hyaline, non-guttulate, 2.6–8.5 × 1.2–2.4 μm.
Notes.
Among the existing species of the genus, P. chloroxantha is morphologically similar to P. rogersonii and P. chlamydospora in having similar hosts, subglobose and laterally collapsed perithecia with a truncate apex, ellipsoidal and smooth ascospores, and conidia without guttules (Herrera et al. 2013). However, P. chloroxantha differs from P. rogersonii in longer conidia (2.6–8.5 μm vs. 2.9–5.5 μm) and rod-shaped to cylindrical conidia, and differs from P. chlamydospora in larger asci (53–76.8 × 4.7–9.2 μm vs. 42.7–56.7 × 3.4–5.7 μm), longer ascospores (7.1–11.5 μm vs. 5.9–8.8 μm), rod-shaped to cylindrical conidia, and the absence of chlamydospores. Additionally, there were 15 bp, 4 bp, 81 bp, 98 bp and 101 bp nucleotide divergences between the type of P. chloroxantha and P. rogersonii, and 10 bp, 8 bp, 70 bp, 82 bp and 84 bp unmatched loci were found between the type of P. chloroxantha and P. chlamydospora in the ITS, LSU, tub2, rpb1 and tef1-α regions (Table 3). Phylogenetic analyses showed that P. chloroxantha is closely associated with P. beijingensis with strong support (MLBS/MPBP/BIPP = 98%/80%/1.0). However, the P. chloroxantha differs morphologically from the latter in having larger asci (53–76.8 × 4.7–9.2 µm vs. 38–58 × 2.5–5 μm), wider ascospores (3.7–6.3 µm vs. 2.5–4 µm), production of greenish yellow pigments in culture, and longer conidia (2.6–8.5 µm vs. 2.6–4.5 µm) (Table 4). Therefore, both morphological and molecular data support the establishment of P. chloroxantha.
Discussion
In this study, P. chlamydospora sp. nov. and P. chloroxantha sp. nov. are described based on morphological characteristics and DNA sequence analyses of combined ITS, LSU, rpb1, tub2 and tef1-α regions. Morphological observations revealed that they share key sexual- and asexual- features with other members of this genus, while a few differences existed that could set them apart. The result of phylogenetic analyses showed that with the addition of the two new species, the tree topology remains mainly consistent with the previous studies (Herrera et al. 2013; Zeng and Zhuang 2021). Pseudocosmospora chlamydospora and P. chloroxantha are closely related to, but clearly distinct from, the existing members of Pseudocosmospora (Fig. 1). Both morphological characteristics and muti-locus phylogenetic analyses support their placements in Pseudocosmospora and are new to science.
Both the morphology and DNA sequence analyses indicate the two new taxa have diatrypaceous hosts (Zhang et al. 2023), which are consistent with the other Pseudocosmospora species (Herrera et al. 2013). By parasitizing the fruitbodies of the hosts, and eventually suppressing or killing them, Pseudocosmospora species may be able to regulate the distribution of diatrypaceous and xylariaceous fungi in nature, thus maintaining the microbial community diversity and balance in forest ecosystems. Some Diatrypaceae species are reported as plant pathogens (Moyo et al. 2018), such as Eutypa lata and Eutypella leprosa are agents of diseases of grapevines (Munkvold et al. 1994; Díaz et al. 2011). Given the parasitic features, Pseudocosmospora species may have potential for biological control applications (Shiono et al. 2016). Moreover, some strains of Pseudocosmospora can produce metabolites with antimicrobial activity and cytotoxicity, which can inhibit plant pathogenic fungi and cancer cells, giving them great potential in agriculture and biomedicine (Hsiao et al. 2014; Shiono et al. 2016).
Since Pseudocosmospora was established by Herrera et al. (2013) who accepted ten species in the genus, several taxa were successively described (Zeng and Zhuang 2017, 2021, 2022; Lechat and Fournier 2020). Study of Pseudocosmospora in China was initiated by Teng (1934) who described Nectria nummulariae Teng [≡P. nummulariae (Teng) Z.Q. Zeng & W.Y. Zhuang] from Hainan Province. Additional ones were consecutively reported from different regions of the country (Zeng and Zhuang 2017, 2021, 2022). Until now, 14 Pseudocosmospora species are known in China, including the two newly described ones in this study. However, since the species diversity of the genus has not been fully revealed, further surveys in the unexplored regions with diverse vegetations are expected in the future.
Supplementary Material
XML Treatment for Pseudocosmospora chlamydospora
XML Treatment for Pseudocosmospora chloroxantha
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Castlebury LA, Rossman AY, Sung GH, Hyten AS, Spatafora JW (2004) Multigene phylogeny reveals new lineage for Stachybotrys chartarum, the indoor air fungus. Mycological Research 108(Pt 8): 864–872. 10.1017/S 095375620400060715449591 · doi ↗ · pubmed ↗
- 2Díaz GA, Prehn D, Latorre BA (2011) First report of Cryptovalsa ampelina and Eutypella leprosa associated with grapevine trunk diseases in Chile. Plant Disease 95(4): 490. 10.1094/PDIS-12-10-091930743357 · doi ↗ · pubmed ↗
- 3Gräfenhan T, Schroers HJ, Nirenberg HI, Seifert KA (2011) An overview of the taxonomy, phylogeny and typification of nectriaceous fungi in Cosmospora, Acremonium, Fusarium, Stilbella and Volutella. Studies in Mycology 68: 79–113. 10.3114/sim.2011.68.04PMC 306598621523190 · doi ↗ · pubmed ↗
- 4Herrera CS, Rossman AY, Samuels GJ, Chaverri P (2013) Pseudocosmospora, a new genus to accommodate Cosmospora vilior and related species. Mycologia 105: 1287–1305. 10.3852/12-39523921243 · doi ↗ · pubmed ↗
- 5Herrera CS, Rossman AY, Samuels GJ, Pereira OL, Chaverri P (2015) Systematics of the Cosmospora viliuscula species complex. Mycologia 107(3): 532–57. 10.3852/14-12225800252 · doi ↗ · pubmed ↗
- 6Hirooka Y, Rossman AY, Samuels GJ, Lechat C, Chaverri P (2012) A monograph of Allantonectria, Nectria, and Pleonectria (Nectriaceae, Hypocreales, Ascomycota) and their pycnidial, sporodochial, and synnematous anamorphs. Studies in Mycology 71(1): 1–210. 10.3114/sim 0001 PMC 331023622685364 · doi ↗ · pubmed ↗
- 7Hsiao CJ, Hsiao G, Chen WL, Wang SW, Chiang CP, Liu LY, Guh JH, Lee TH, Chung CL (2014) Cephalochromin induces G 0/G 1 cell cycle arrest and apoptosis in A 549 human non-small-cell lung cancer cells by inflicting mitochondrial disruption. Journal of Natural Products 77: 758–65. 10.1021/np 400517 g 24588135 · doi ↗ · pubmed ↗
- 8Lechat C, Fournier J (2020) Pseudocosmospora hypoxylicola (Nectriaceae), a new species from the French Alps. Ascomycete.org 12: 6–10. 10.25664/art-0290 · doi ↗