The 1 –Cys peroxiredoxin, PRDX-6, suppresses an NHR-49-dependent pro-survival response, including the Flavin monooxygenase, FMO-2, that protects against fungal and bacterial infection
Emma L. Button, Emilia Dwyer, Jake B. Lewis, Madison S. Mortensen, Elise McDonald, Eloise Butler, Fiona Pearson, Alice E. Tang, Jennifer L. Watts, Elizabeth A. Veal

TL;DR
The study shows that the protein PRDX-6 in worms influences lifespan, immunity, and stress resistance by regulating other genes like FMO-2 and NHR-49.
Contribution
The paper reveals a novel protective role of NHR-49-dependent FMO-2 in prdx-6 mutants against infection and stress.
Findings
Loss of PRDX-6 increases lifespan and resistance to arsenite and pathogens in C. elegans.
PRDX-6 loss leads to increased NHR-49 activity and FMO-2 expression, which protects against infection.
NHR-49-driven MUFA synthesis is essential for embryonic development in prdx-6 mutants.
Abstract
Reactive oxygen species (ROS)-induced cell damage contributes to many diseases. However, ROS also contribute to cell signaling and immune defences. As ubiquitous thiol peroxidases, peroxiredoxins (Prdx) play integral roles in balancing ROS functions. High levels of Prdx6 are associated with increased metastasis and resistance to chemotherapy, rendering Prdx6 a therapeutic target for treatment of a broad range of cancers. However, Prdx6 has additional activities, in lipid signaling and selenocysteine metabolism, and it remains unclear how Prdx6's thiol peroxidase activity contributes to disease. Here we have investigated the role/s of Prdx6 in the nematode worm Caenorhabditis elegans. Consistent with a ROS-protective role for PRDX-6, prdx-6 mutant C. elegans exhibit elevated levels of lipid oxidation, more apoptotic corpses in their germline and are more susceptible to the toxicity of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
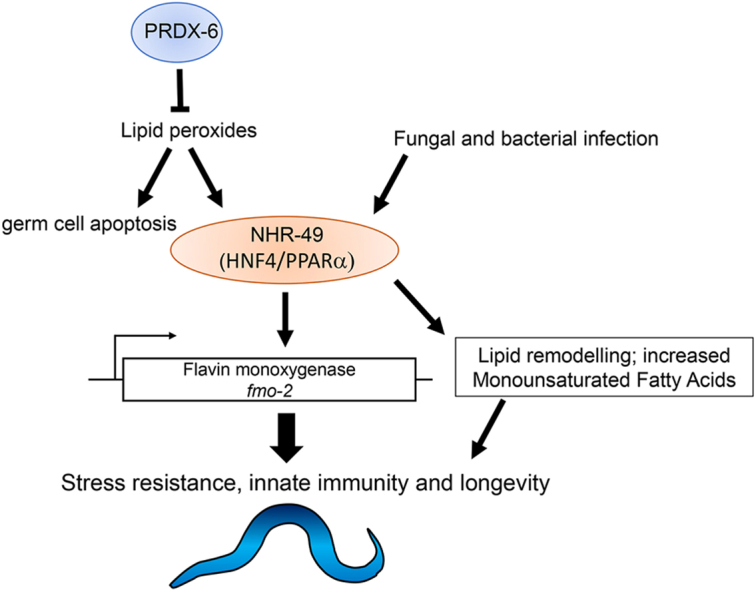 Figure 1
Figure 1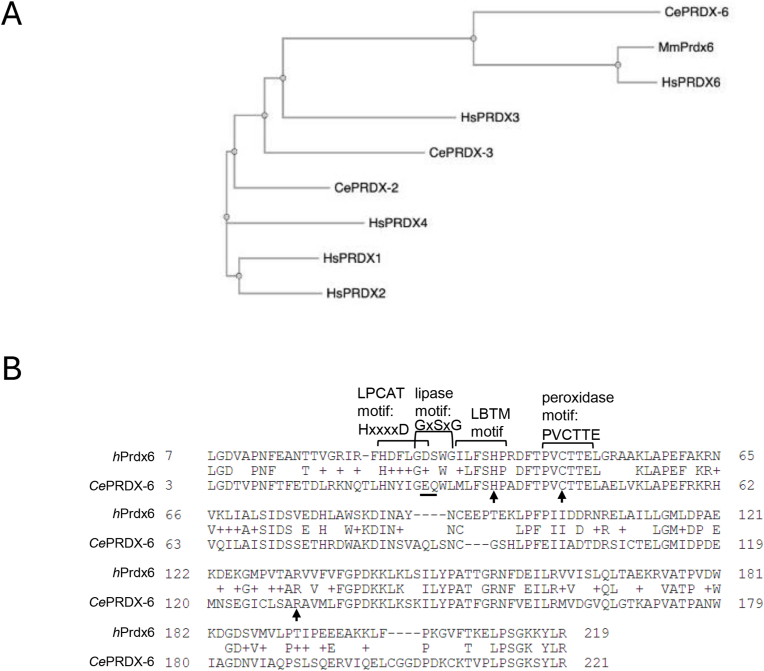 Figure 2
Figure 2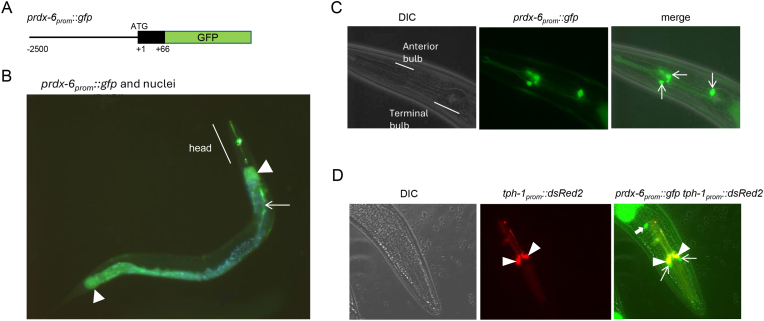 Figure 3
Figure 3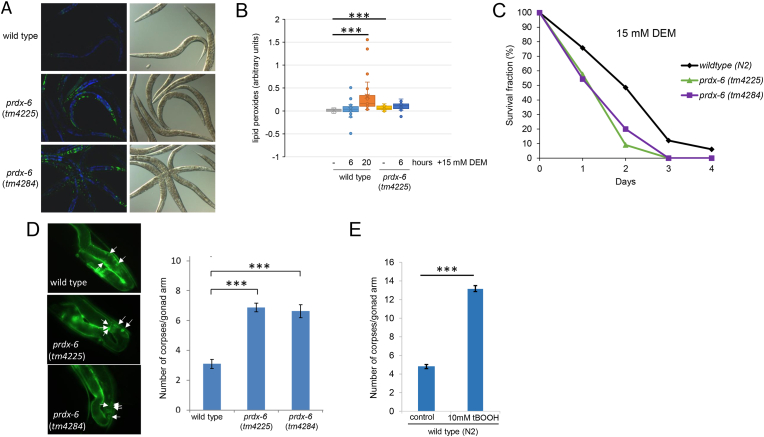 Figure 4
Figure 4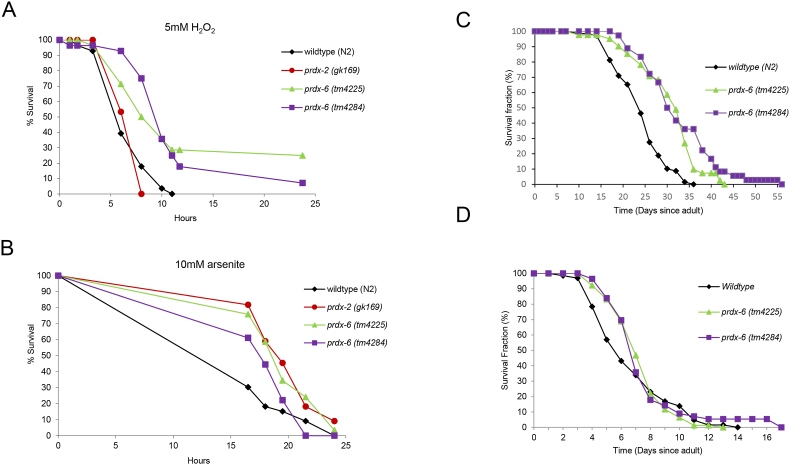 Figure 5
Figure 5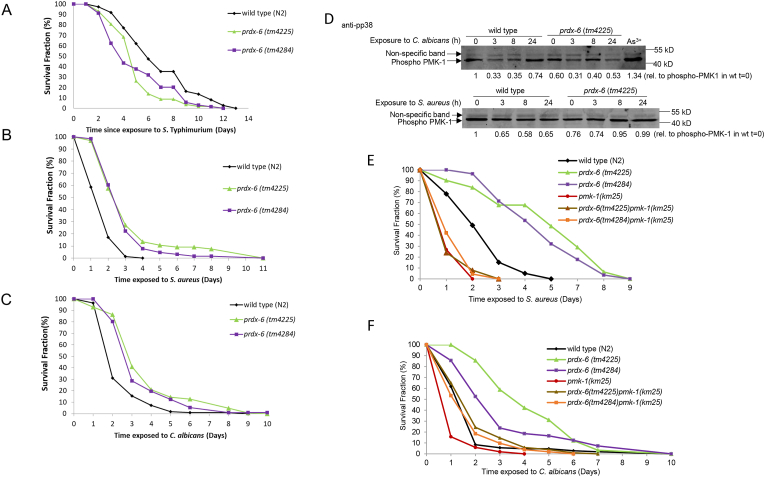 Figure 6
Figure 6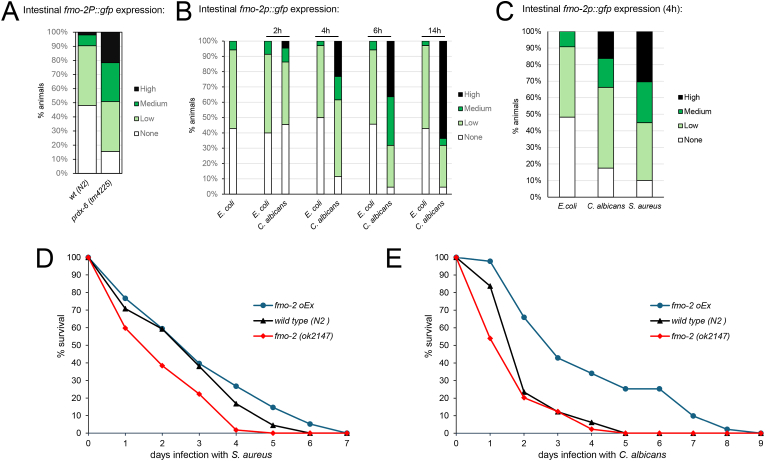 Figure 7
Figure 7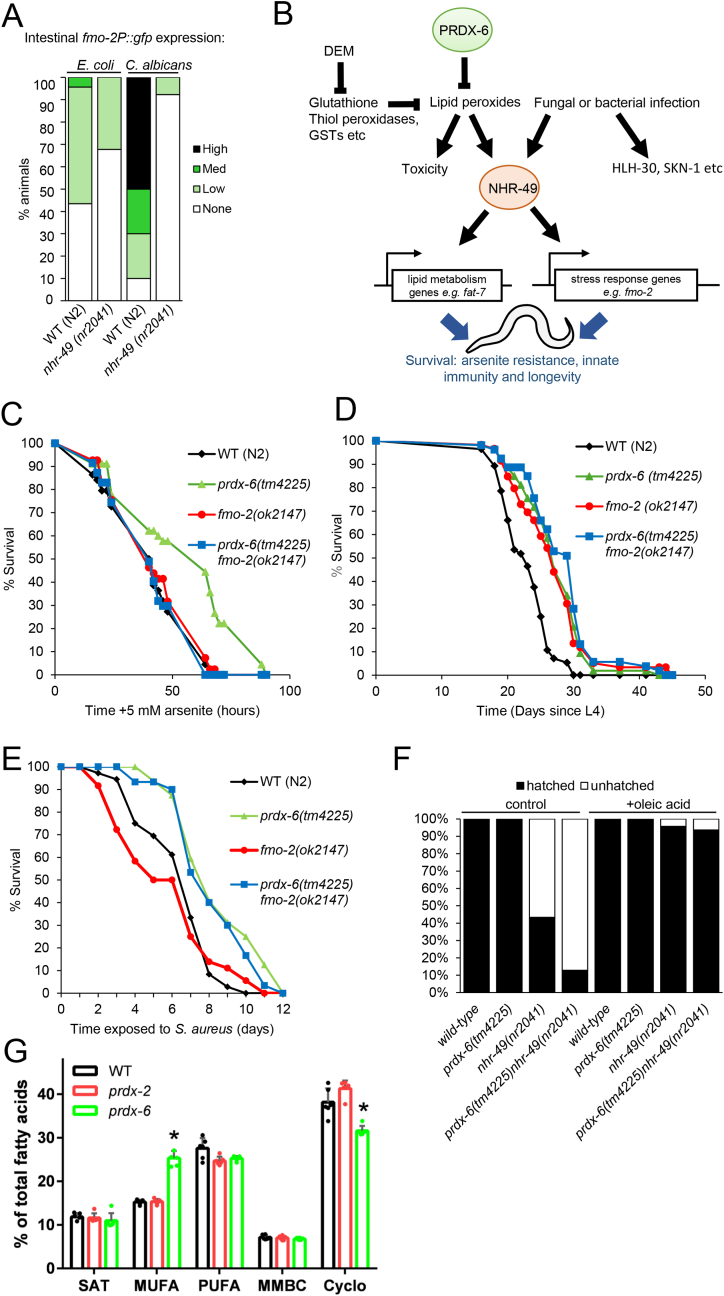 Figure 8
Figure 8Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRedox biology and oxidative stress · Genetics, Aging, and Longevity in Model Organisms · Selenium in Biological Systems
Introduction
1
Reactive oxygen species (ROS) are generated as a by-product of aerobic metabolism and can cause profound damage to cellular macromolecules. Indeed, NADPH oxidases (Nox/Duox), ROS-generating systems, have evolved as an innate immune defence, exploiting ROS as powerful antimicrobials (For a review see Ref. [1]). Accordingly, enzymes and systems that reduce ROS levels play important roles in protecting against oxidative damage. Peroxiredoxins (Prdx) are amongst these ROS defences; reducing peroxides via reversible oxidation of a conserved, peroxide-reacting cysteine/s [2]. Alterations in Prdx levels are linked with a number of diseases and ageing. For example, the mammalian 1- Cys peroxiredoxin (Prdx), Prdx6, has important roles in stress resistance [3,4], apoptosis [5,6] and is up-regulated in many cancers [[7], [8], [9]]. Moreover, high levels of Prdx6 expression are associated with increased metastasis and resistance to chemotherapy making them candidate chemotherapeutic targets to treat a range of cancers [[9], [10], [11], [12], [13], [14], [15], [16]].
However, in addition to their peroxidase activity, eukaryotic Prdx have other activities that may contribute to their in vivo function. For example, 2-Cys Prdx directly and indirectly mediate some of the positive signaling functions of low levels of peroxide (For reviews see Ref. [2,17]. Moreover, the mammalian 1-Cys Prdx, Prdx6, has additional signaling activities: a Ca^2+^ independent phospholipase A_2_ (iPLA_2_) activity and a lysophosphatidylcholine acyl transferase (LPCAT) activities (for a review see Ref. [18]. The peroxidase, iPLA_2_ and LPCAT activities have distinct active sites, with the peroxidase activity centered around a catalytic cysteine and the iPLA_2_ activity centered around a catalytic serine and a lipase motif. Studies in mice have suggested that peroxidase, lipase and acylation activities all contribute to Prdx6's physiological functions and roles in disease [11,19,20]. It has been proposed that the lipid-binding capacity of Prdx6 allows it to protect cell membranes against ROS-induced damage by detoxifying phospholipid hydroperoxides [21]. However, Prdx6 was recently shown to protect against lipid peroxides indirectly; by facilitating the incorporation of selenium into selenoproteins, such as the glutathione peroxidase, Gpx4 [[22], [23], [24]].
Indeed, the presence of multiple peroxiredoxins and 30 selenoproteins, including 4 glutathione peroxidases, make the contributions of Prdx6's different activities to health and disease, more challenging to dissect in mammals. In contrast, the genome of the nematode worm Caenorhabditis elegans encodes only 3 peroxiredoxin genes (Fig. 1A) and a single selenoprotein, the thioredoxin reductase, TRXR-1, which is redundant with glutathione reductase [25]. Moreover, C. elegans is also a highly useful system to investigate the roles of ROS in signaling, ageing and infection (for a review see Ref. [26]. For example, the p38 MAPK, PMK-1, and the C. elegans orthologue of the mammalian Nrf2 transcription factor, SKN-1, play important roles in mediating many of the positive effects of ROS (For a review see Ref. [27]). In addition, the NHR-49 transcription factor, which activates similar genes to mammalian PPARα, also responds to ROS, activating a protective transcriptional response to peroxide [28].Fig. 1Sequence comparisons reveal thatC. elegansPRDX-6 is [A] more closely related to mammalian Prdx6 proteins than other 1 and 2-Cys Prdx and [B] contains the peroxidase motif present in Prdx6 family but lacks the phospholipase A2 or LPCAT motifs found in human Prdx6. [A] Phylogenetic tree generated using ClustalW2 software demonstrating the relationships between the C. elegans and mammalian 1-cys peroxiredoxin, CePRDX-6, the human PRDX-6 (HsPRDX-6) and the Mus musculus PRDX-6 (MmPRDX-6), and the 2-cys peroxiredoxins, CePRDX-2 and CePRDX-3 and HsPRDX1, HsPRDX2, HsPRDX3 and HsPRDX4. [B] Alignment of CePRDX-6 and human Prdx6 amino acid sequences indicates that the peroxidase motif, PVCTTE, that is unique to Prdx6 family, and the catalytic triad C47–H39 -R132 (arrows) are both present in human and C. elegans PRDX-6. The LPCAT motif, HxxxxD and the lipase motif, GxSxG, required for acylation of lysophosphatidylcholine and for Ca^2+^ independent phospholipase A2 (iPLA2) activity of mammalian Prdx6 respectively, are absent from C. elegans PRDX-6, with the critical aspartate for LPCAT activity in the LPCAT motif replaced with glutamate and the critical serine for phospholipase activity in the lipase motif replaced with a glutamine (underlined) in C. elegans PRDX-6.Fig. 1
Previous studies have suggested that the mitochondrial 2-Cys Prdx, PRDX-3, does not play an important role in protecting against oxidative damage, and prdx-3 mutant animals have a wild-type lifespan [29,30]. In contrast, the single cytosolic 2-Cys Prdx, PRDX-2, plays several important physiological roles [29,31,32]. This includes tissue-specific roles, with intestinal PRDX-2 protecting against the acute toxicity of hydrogen peroxide [29], whereas PRDX-2 is required in specific neurones for hydrogen peroxide-induced inhibition of feeding [31]. Consistent with a positive role in promoting ROS signal transduction, PRDX-2 is also required for arsenite and metformin-induced activation of the p38 MAPK, PMK-1 [29,33]. However, PRDX-2 limits resistance to some oxidative stress conditions, including arsenite, by promoting insulin secretion that inhibits the activity of SKN-1(Nrf2) and DAF-16(FOXO) [29,32]. Notably, despite reduced insulin signaling and increased SKN-1 and DAF-16 activity, prdx-2 mutant animals are short-lived [29,32]. However, it is unclear whether the C. elegans 1-Cys Prdx, PRDX-6 plays important roles under physiological or stress conditions [34]. The goal of this study was to address this question.
Comparison of the C. elegans PRDX-6 sequence with the human Prdx6 sequence demonstrates that C. elegans PRDX-6 contains the peroxidase domain and histidine important for lipid binding, but the catalytic serine required for iPLA_2_ activity is not conserved (Fig. 1B). Moreover, C. elegans only incorporate selenocysteine into a single protein, the thioredoxin reductase TRXR-1, which is redundant with glutathione reductase [25]. This provided us with the opportunity to investigate the contribution that the thiol peroxidase activity plays to the in vivo biological role/s of a 1-Cys Prdx in a whole animal model. Consistent with PRDX-6's peroxidase activity playing an important role in removing endogenous lipid peroxides we find that loss of prdx-6 causes detectable increases in intracellular levels of ROS, oxidized lipids, germ cell apoptosis and renders worms hypersensitive to the toxicity of DiEthylMaleate (DEM). However, unexpectedly, at lower temperatures prdx-6 mutant animals are long-lived and more resistant to arsenite, hydrogen peroxide, bacterial and fungal infection. Our data suggest that the increased resistance of prdx-6 mutant C. elegans to arsenite and pathogenic infection may reflect increased expression of oxidative stress-induced genes activated by the NHR-49 transcriptional regulator, including the pro-longevity fmo-2 gene that protects against bacterial infection [[35], [36], [37]]. Indeed, we find that fmo-2 also protects C. elegans against the fungal pathogen Candida albicans. Thus, we propose that loss of PRDX-6 activates NHR-49(HNF4/PPARα), remodelling lipid metabolism and priming C. elegans innate immune defences against opportunistic human pathogens.
Results
2
PRDX-6 is expressed in the intestine, I2 and I4 interneurones
2.1
To determine where PRDX-6 was expressed, transgenic animals were generated in which the 2.5 kb promoter region upstream of the prdx-6 orf was used to drive the expression of the first 22 amino acids of PRDX-6 in frame with GFP (prdx-6prom::gfp) (Fig. 2A). Consistent with high throughput single cell mRNA expression analyses [38], prdx-6prom::GFP expression was detected in the intestine and specific neurones within the head in all larval stages and adults and the gonadal sheath of adult hermaphrodites (Fig. 2B). The intestinal expression of prdx-6prom::GFP, was greatest in the most anterior and posterior intestine, consistent with the expression of many stress-induced genes [39,40]. In the head prdx-6prom::GFP was expressed specifically in three neurones. Two of the cell bodies were in the anterior bulb with symmetrical axonal projections, indicating they represented a pair of neurones (Fig. 2C). To determine the identity of these neurones, we compared the expression of prdx-6prom::gfp with that of the tryptophan hydroxylase, TPH-1, that is expressed in serotonergic neurones, including the neurosecretory-motor neurones (NSM) neurones. Co-expression of prdx-6prom::GFP and tph-1::dsRed2, suggested that prdx-6prom::GFP is expressed in the I2L and I2R inter-neurones, that are adjacent to the NSM neurones in which TPH-1 is expressed, as well as a third inter-neuron with a cell body in the terminal bulb and axons projecting to the anterior bulb that is likely to be I4 (Fig. 2C–D).Fig. 2Analysis of transgenic animals expressing a transcriptional reporter reveals that PRDX-6 is expressed in the intestine, gonadal sheath, I2 and I4 neurones. [A] Schematic representation of the prdx-6P::gfp transcriptional reporter [B] Analysis of prdx-6P::gfp expression in a young adult worm, nuclei were imaged by staining with DAPI (blue) and a merged images of green and blue channels is shown. Arrowheads indicate ends of intestine, arrow indicates gonadal sheath. [C] Analysis of prdx-6P::gfp expression in head. GFP-positive neuronal cell bodies are indicated by arrows on merged image of DIC and GFP. [D] Analysis of prdx-6P::gfp expression in head of C. elegans co-expressing tph-1P::dsRed2 in the NSM cell bodies (labelled with triangles). Thin arrows on merged image (right hand panel) indicate prdx-6p::GFP-positive cell bodies adjacent to the NSM (interneurones I2L and I2R). DsRed2 also fluoresces under the GFP excitation wavelength therefore tph-1p::DsRed2-positive NSM neurones appear orange on the merged image. A thick arrow indicates the third, unidentified, prdx-6p::GFP-positive neuronal cell body.Fig. 2
Loss of prdx-6 increases resistance to oxidative stress and lifespan at 15 °C
2.2
To analyse the function of PRDX-6, an RNAi clone was generated targeting PRDX-6. Western blot analysis, using anti-PRDX-6 antibodies, detected a band, consistent with the expected size of PRDX-6 (25.6kD), in lysates from control animals that was absent from prdx-6 RNAi-treated animals, confirming that prdx-6 RNAi was effective at reducing PRDX-6 levels (Fig. S1A). Two prdx-6 mutant alleles were also obtained from the National Bioresource Project (NBRP), prdx-6 (tm4225) and prdx-6 (tm4284) containing a 442bp delete and a 4bp insertion in the second intron and a 356bp deletion in the third exon respectively (Fig. S1B). Analysis of protein lysates from wild type, and outcrossed prdx-6 (tm4225) and prdx-6 (tm4284) mutant worms using anti-PRDX-6 antibodies confirmed the absence of PRDX-6 from either prdx-6 (tm4225) or prdx-6 (tm4284) mutant worms suggesting that both prdx-6 (tm4225) and prdx-6 (tm4284) are null alleles (Fig. S1C).
Although I2 neurones are important for the hydrogen peroxide-induced inhibition of feeding, we found no evidence that PRDX-6 was important for this behavioural response to peroxide (Fig. S2)[31]. However, consistent with the peroxidase activity of PRDX-6 acting as a barrier to lipid hydroperoxides, we observed increased DCFDA staining (Fig. 3A) and significantly more oxidized lipids were detected in prdx-6 mutant animals stained with BODIPY-C11 (Invitrogen™), a well-established reporter of lipid peroxidation (Fig. 3B). Moreover, prdx-6 mutant worms were also highly sensitive to toxic doses of the thiol-reactive diethyl maleate (DEM) that triggers lipid peroxidation [41] (Fig. 3B–C). As the organic peroxide (tBOOH) increases the number of apoptotic corpses detected in C. elegans germlines, the increased number of corpses in prdx-6 mutant germlines, suggested that PRDX-6 also protects against ROS-induced apoptosis (Fig. 3D–E) [42]. In contrast, when we examined the survival of wild type, prdx-6 (tm4225) and prdx-6 (tm4284) mutant worms on plates containing toxic levels of hydrogen peroxide and arsenite, we found that prdx-6 mutant animals were significantly more resistant to both oxidative stress agents (Fig. 4A–B).Fig. 3Loss of PRDX-6 increases levels of lipid peroxides, germ cell death and susceptibility to the toxicity of Diethyl maleate. [A] wild type, prdx-6 (tm4225) and prdx-6 (tm4284) mutant worms were stained with the ROS probe H_2_DCFDA and the fluorescence observed using fluorescence spectroscopy. Over 30 worms were observed in at least three independent experiments and representative DIC and fluorescent images are shown, with DCF fluorescence in green and autofluorescence in blue. [B] lipid peroxidation was measured in wild-type and prdx-6 (tm4225) animals without stress (control), and following timed exposure to 15 mM DEM (n = 20–28 for each group) ∗∗∗P < 0.001 compared with wild-type control. [C] The survival of L4 larval stage wild type, prdx-6 (tm4225) and prdx-6 (tm4284) mutant animals was monitored at the indicated time points on plates containing 15 mM DEM. n = 33–35 in each group. Log rank analysis indicates that the survival of prdx-6 mutant animals is significantly decreased compared with wild-type (p < 0.001). [D] Wild type (N2) ced-1::GFP, prdx-6 (tm4225) ced-1::GFP and prdx-6 (tm4284) ced-1::GFP mutant worms were used to visualise engulfed apoptotic corpses in the germline (indicated by arrows). The number of apoptotic corpses in one gonad arm were counted in young adult wild type (N2) ced-1::GFP, prdx-6 (tm4225) ced-1::GFP and prdx-6 (tm4284) ced-1::GFP mutant animals or [E] before or following 90 min exposure (followed by 60 min recovery) of wild type (N2) ced-1::GFP to 10 mM tert-butyl hydroperoxide (tBOOH) at 24 °C. Mean values are shown from groups of 25–30 animals. P values were determined using Students T-test. ∗∗∗p < 0.001.Fig. 3. Fig. 4Loss of PRDX-6 increases survival under acute oxidative stress conditions and lifespan at 15 °C but not 25 °C. The survival of L4 larval stage wild type, prdx-2 (gk169), prdx-6 (tm4225) and prdx-6 (tm4284) mutant animals was monitored at the indicated time points [A-B] on plates containing either [A] 5.0 mM H_2_O_2_ or [B] 10.0 mM arsenite. These experiments were carried out three times with similar results and a representative experiment is shown. n = 23–35 in each group. Log rank analysis indicates that the survival of prdx-6 mutant animals is significantly increased compared with wild-type (p < 0.002). [C-D] The survival of wild type (N2) (n = 69*), prdx-6* (tm4225) (n = 41) and prdx-6 (tm4284) (n = 36) mutant worms was monitored on live E. coli OP50 at [C] 15 °C or [D] 25 °C. Experiments were repeated 3 times and representative experiments are shown. Mean and median survival of wild type (N2), prdx-6 (tm4225) and prdx-6 (tm4284) mutant worms and log-rank analysis of prdx-6 (tm4225) and prdx-6 (tm4284) mutant worms compared with wild type indicated that differences in survival were statistically significant at 15 °C but not 25 °C (Tables S1–3).Fig. 4
Our discovery that prdx-6 mutant worms have an increased resistance to H_2_O_2_ contrasts sharply with the increased H_2_O_2_ sensitivity of animals lacking PRDX-2 [29]. Moreover, whereas loss of prdx-2 reduces the lifespan of C. elegans at 15 °C [29] prdx-6 mutant animals were long-lived and exhibited a slower decline in mobility, consistent with this extended lifespan reflecting slower ageing (Fig. 4C–Table S1, Fig. S3). The effect of PRDX-2 on ageing is temperature-dependent, with wild-type and prdx-2 mutant worms having similar lifespans at 25 °C [29,34]. Intriguingly, loss of PRDX-6 did not significantly alter the lifespan of animals maintained at 25 °C either, suggesting that like PRDX-2, PRDX-6, has a temperature-dependent role in longevity (Fig. 4D, Tables S2–3). Overexpression of PRDX-2 increases the resistance of wild-type animals to both arsenite and H_2_O_2_ [29]. This raised the possibility that a compensatory increase in PRDX-2 levels might be responsible for the increased lifespan and oxidative stress resistance of prdx-6 mutant C. elegans. However, PRDX-2 levels were similar in wild type, prdx-6 (tm4225) and prdx-6 (tm4284) mutant worms, both before and following exposure to H_2_O_2_ (Fig. S4).
Loss of PRDX-6 increases the resistance of C. elegans to Staphylococcus aureus and C. albicans infection but not Salmonella Typhimurium
2.3
There were a number of possible explanations for the temperature-dependent effect of PRDX-6 on lifespan. For instance, a major lifespan-limiting factor at 25 °C is the increased growth and pathogenicity of the E. coli food source at these higher temperatures [29]. Moreover, while ROS play an important role in protecting C. elegans against some pathogens [43,44] ROS generated during infection have been proposed to be partially responsible for the premature death of C. elegans infected with the gram negative bacteria, S. Typhimurium [45,46]. Indeed, the survival of prdx-6 mutants exposed to S. Typhimurium, which establishes a persistent infection in the intestine that causes the premature death of the worm, was reduced compared with wild-type animals (Fig. 5A, Table S4). However, in contrast, loss of PRDX-6 significantly increased the resistance of C. elegans to infection with two other human pathogens, the gram positive bacteria, Staphylococcus aureus and the dimorphic fungus C. albicans (Fig. 5B–C, Table S5–6). Intriguingly, this suggests that PRDX-6 has pathogen-specific roles in innate immunity. The increased resistance of prdx-6 mutant C. elegans to infection with S. aureus and C. albicans, both of which kill C. elegans more quickly than S. Typhimurium, was consistent with the increased resistance of these animals to the acute toxicity of arsenite and peroxide (Fig. 4A–B). This raised the possibility that loss of PRDX-6 activates a stress-protective response that increases resistance to a variety of acute stresses but does not protect against a more chronic infection with S. Typhimurium.Fig. 5Loss ofprdx-6increases resistance to infection byS. aureusandC. albicansby mechanisms that do not involve increased PMK-1 activity The survival of young adult wild type (N2), prdx-6 (tm4225) and prdx-6 (tm4284) mutant animals transferred on to plates seeded with **[A]**S. Typhimurium (SL1344) **[B]**S. aureus (NCTC 8325) or **[C]**C. albicans (SN148). These experiments were carried out with >50 animals per group and repeated three times with similar results. Representative experiments are shown. The mean survival of wild type, prdx-6 (tm4225), prdx-6 (tm4284) and P-values were determined by log-rank analysis (Table S4–6). [D] Immunoblotting analysis with anti-phospho p38 antibodies of proteins extracted from wild type and prdx-6 (tm4225) mutant worms before and following, 3, 8 and 24 h exposure to C. albicans (upper panel) or S. aureus (lower panel). Relative levels of PMK-1 phosphorylation (normalised for protein loading using non-specific band) are indicated. [E-F] The survival of wild type, prdx-6 (tm4225), prdx-6 (tm4284), pmk-1 (km25), prdx-6 (tm4225) pmk-1 (km25) and prdx-6 (tm4284) pmk-1 (km25) mutant following infection with [E]S. aureus[F]C. albicans. The mean survival of wild type, prdx-6 (tm4225), prdx-6 (tm4284), prdx-6 (tm4225) pmk-1 (km25) and prdx-6 (tm4284) pmk-1 (km25) and P-values were determined by log-rank analysis (Table S7–8).Fig. 5
Loss of PRDX-6 and exposure to C. albicans and S. aureus increase expression of the flavin monooxygenase, FMO-2 which protects against both pathogens
2.4
Next, we investigated the basis for the increased resistance of prdx-6 mutant C. elegans to infection with S. aureus or C. albicans. Arsenite, peroxide and infection are amongst the stresses that have been shown to activate the p38 MAPK, PMK-1, and the C. elegans Cap n Collar transcription factor SKN-1, that mediates protective transcriptional responses. Indeed, the p38 MAPK, PMK-1 is important for oxidative stress resistance and innate immunity. However, levels of phosphorylated PMK-1 were not increased in prdx-6 mutants suggesting that increased PMK-1 activity was unlikely to explain their increased resistance to arsenite or infection (Fig. 5D, Fig. S5). Moreover, while pmk-1 mutants were highly susceptible to both S. aureus and C. albicans, analysis of prdx-6 pmk-1 double mutants indicated that the increased resistance of prdx-6 mutant animals to C. albicans was only partially dependent on PMK-1 (Fig. 5E–F, Tables S7-S8). This suggested that loss of PRDX-6 might increase innate immunity and stress resistance by mechanism/s not involving PMK-1.
In prdx-2 mutants lower levels of insulin signaling increase arsenite resistance by upregulating SKN-1 and the FOXO transcription factor DAF-16 [32]. Indeed, increased SKN-1 and DAF-16 activity can increase resistance to arsenite, protect against pathogens and extend lifespan [27]. However, we could not find any evidence that the increased ROS in animals lacking prdx-6 was sufficient to increase expression of two different SKN-1-dependent reporter genes, expressing GFP from the gcs-1 or gst-4 promoters or a DAF-16-dependent reporter gene, expressing GFP from the sod-3 promoter (Fig. S6).
Notably, one of the most highly peroxide-induced genes, the Flavin monooxygenase, fmo-2, is activated in a skn-1-independent manner by the nuclear hormone receptor NHR-49 [28]. Although wild-type and prdx-6 mutant C. elegans appeared to contain similar nuclear levels of NHR-49 (Fig. S7), it was plausible that loss of PRDX-6 might increase NHR-49 activity by increasing the availability of an oxidized lipid ligand [47]. Hence, we tested whether the increased ROS in prdx-6 mutants might be sufficient to increase the expression of fmo-2. Notably, we found that, although the levels of expression of an fmo-2P::gfp reporter are normally low, intestinal levels of expression were significantly increased in animals lacking prdx-6 (Fig. 6A). Increased FMO-2 expression increases the lifespan of C. elegans maintained on non-proliferating E. coli [35]. Hence, our finding suggested increased expression of fmo-2 might contribute to the increased resistance of prdx-6 mutant C. elegans to infection with S. aureus or C. albicans.Fig. 6FMO-2 expression is induced by loss ofprdx-6and protects against infection withC. albicansandS. aureus The intestinal expression of a fmo-2P::gfp reporter gene was [A] increased in prdx-6 (tm4225) mutant compared with wild-type animals (Chi^2^ test p = 8.89889E-07) [B–C] rapidly increases following transfer of C. elegans to **[B]**C. albicans or **[C]**S. aureus compared with transfer to E. coli control. [D-E] The survival of wild-type (N2), fmo-2 (ok2147) mutant and fmo-2 over-expressing (fmo-2 OEx) C. elegans following exposure to **[D]**S. aureus or **[E]**C. albicans. The experiments shown are representative of at multiple (>3) independent experiments involving approximately 80–150 animals in each group. The mean survival of wild type, fmo-2 (ok2147) and fmo-2oEx and P-values were determined by log-rank analysis (Table S7–8).Fig. 6
C. albicans and S. aureus infection initiate both distinct and overlapping C. elegans transcriptional responses [48]. Consistent with the hypothesis that increased expression of *fmo-*2 might be an important innate response to infection, fmo-2 is very highly induced in response to Staphylococcus aureus [37,49,50]. Moreover, although fmo-2 was not amongst the genes found to be induced in response to C. albicans in previous studies [48], exposure of wild-type C. elegans to C. albicans also caused a rapid increase in the intestinal expression of fmo-2::gfp (Fig. 6B). This increase preceded any visible evidence of C. albicans infection but was not apparent when worms were exposed to heat-killed C. albicans, suggesting that induction was due to a non-denaturable signal emanating from the yeast (Fig. S8). Indeed, we observed a similar increase in intestinal fmo-2p::gfp expression when animals were exposed to either C. albicans or S. aureus indicating that fmo-2 induction is part of a common response to both pathogens (Fig. 6C). As previously reported, the ability of fmo-2 mutant animals to survive infection with S. aureus was significantly compromised (Fig. 6D–Table S9) [37]. There was also a small, but consistent, increase in the susceptibility of fmo-2 mutant animals to C. albicans, particularly in the early stages of infection (Fig. 6E–Table S10). Conversely, the survival of animals expressing multiple copies of the fmo-2 gene infected with C. albicans was significantly prolonged compared with wild-type (p < 0.001) (Fig. 6E) and there was a similar correlation between FMO-2 levels and survival of S. aureus-infected animals (Fig. 6D). This suggests that this flavin monoxygenase is part of a common, protective innate immune response to fungal and bacterial pathogens.
The NHR-49 transcriptional regulator is activated by tBOOH and required for increased fmo-2 expression in response to peroxide, fasting and S. aureus infection [28,37]. Similarly, C. albicans-induced increases in fmo-2p::GFP expression were ablated in nhr-49 mutant C. elegans (Fig. 7A). Accordingly, we propose that the increased ROS present in prdx-6 mutant animals may increase NHR-49 activity driving the expression of fmo-2 and other genes [51,28,37] responsible for their increased survival (Fig. 7B). Indeed, our analysis of prdx-6 fmo-2 mutants indicated that fmo-2 is required for the increased arsenite resistance of prdx-6 mutant animals (Fig. 7C).Fig. 7NHR-49 is required for induction offmo-2in response toC. albicansleading us to propose that NHR-49 mediates the effect of loss ofprdx-6onfmo-2expression an stress resistance and innate immunity [A] The intestinal expression of the fmo-2P::gfp reporter gene was increased in response to 18h C. albicans exposure in wild-type but not nhr-49 (nr2041) mutant C. elegans The experiment shown is representative of multiple independent experiments involving at least 20 animals in each group. There was a significant difference between intestinal fmo-2p::gfp expression in wild-type and nhr-49 mutant C. elegans exposed to C. albicans (Chi^2^ test; p = 4.10 x10^−7^).[B] We propose that loss of PRDX-6's lipid peroxidase activity leads to increased lipid peroxides and NHR-49 activity, activating a transcriptional program that includes the flavin monooxygenase, FMO-2, thus prolonging survival during acute oxidative stress, fungal and bacterial infection. Fungal and bacterial infection also activate other transcriptional regulators (HLH-30, SKN-1 etc) that contribute to innate immunity. [C] FMO-2 is required for the arsenite resistance that results from loss of prdx-6. L4 stage animals were transferred to plates containing 5 mM sodium arsenite and assayed for survival at regular intervals. This experiment was repeated at least three times, and a representative experiment shown. At least 40 animals were assessed per strain. Log Rank analysis, compared to wild-type, found a significant P-value of 0.001 [D] Survival analysis indicated that prdx-6 (tm4225), fmo-2 (ok2147) and prdx-6 (tm4225) fmo-2 (ok2147) mutant animals are all long-lived at 15 °C compared with wild-type (N2). Assays were carried out in the presence of FUDR. Mean lifespan was compared to wild-type by Log Rank (Table S11). [E] The survival of wild-type (N2), prdx-6 (tm4225), fmo-2 (ok2147) and prdx-6 (tm4225) fmo-2 (ok2147) mutant animals infected with S. aureus. Mean survival was assessed by Log Rank (Table S12). [F] Concurrent loss of prdx-6 and nhr-49 results in high levels of embryonic lethality, with 90 % of prdx-6 (tm4225) nhr-49 (nr2041) eggs unhatched after 48 h at 15 °C. This embryonic lethality was rescued when parents were maintained on media supplemented with 0.8 mM oleic acid. Each group contains 55–213 total eggs/progeny from ∼20 parents and experiment was repeated at 15 °C with similar results. [G] Fatty acid composition of young adult wild-type (N2), prdx-2 (gk169), and prdx-6 (tm4225). Major classes of fatty acids (defined in Table S13) were compared, revealing a significant increase in monounsaturated fatty acids and a corresponding decrease in cyclopropyl fatty acids in prdx-6 animals compared to wild-type. Abbreviations: SAT, saturated fatty acids; MUFA, monounsaturated fatty acids; PUFA, polyunsaturated fatty acids; MMBC, monomethyl branched chain fatty acids; Cyclo, cyclopropyl fatty acids.Fig. 7
Although fmo-2 is required to mediate the life-extending effects of hypoxia and dietary restriction at 20 °C, fmo-2 mutant animals were longer lived than wild-type animals at 15 °C. Intriguingly, this suggests that, like PRDX-6, FMO-2's effect on aging is also temperature dependent. Nevertheless, loss of prdx-6 caused no further increase in the lifespan of fmo-2 mutant worms (Fig. 7D–Table S11). This raised the possibility that other nhr-49-upregulated genes could slow ageing in both prdx-6 and fmo-2 mutant animals. Indeed, fmo-2 was not required for the increased resistance of prdx-6 mutants to S. aureus suggesting that other NHR-49-upregulated genes could also be involved in protecting against infection (Fig. 7E–Table S12). To test this hypothesis, we combined prdx-6 and *nhr-*49 mutant alleles. Strikingly, we observed a high level (>90 %) of embryonic lethality in prdx-6 nhr-49 mutant animals (Fig. 7F). This was consistent with increased nhr-49 activity playing an important role in the phenotypes of prdx-6 mutant C. elegans. Moreover, it suggests that in the absence of *prdx-*6, the remodelling of lipid and oxidative metabolism by nhr-49 becomes critically important for embryonic development. The analysis of unhatched eggs indicated no particular stage at which nhr-49 prdx-6 mutant embryos arrested/died, suggesting a stochastic loss of viability. Interestingly, hatched larvae proceeded to reach adulthood, suggesting that dietary factors could be sufficient to support development post-hatching. NHR-49 is an important regulator of lipid metabolism, with lower levels of the C18 monounsaturated fatty acid oleate in nhr-49 mutant animals [52]. Indeed, the essential requirement for NHR-49 in prdx-6 mutant worms could be rescued by supplementation with oleate (Fig. 7F). This raised the possibility that prdx-6 mutant animals might also have a de-regulated lipid metabolism. Indeed, consistent with elevated NHR-49 activity, GC/MS analysis revealed that prdx-6 mutant animals, but not prdx-2 mutants, synthesize increased levels of monounsaturated fatty acids (Fig. 7G–Table S13).
Discussion
3
Here we have uncovered that the single C. elegans 1-Cys Prdx, PRDX-6, plays important and unexpected roles in regulating oxidative stress resistance and innate immunity. Notably, we reveal that loss of this thiol peroxidase not only increases the lifespan of C. elegans but also prolongs survival following exposure to acutely toxic oxidative stress agents and two very different opportunistic human pathogens. To our knowledge, this is the first study to identify a role for a Prdx in suppressing innate immune defences and promoting survival under physiological and acute stress conditions.
Studies with Prdx6 knockout mice have suggested Prdx6 provides a non-redundant barrier to ROS and is important in maintaining normal insulin signaling [53], fertility [54] brain [55,56] and muscle function [57]. Consistent with this we have shown that prdx-6 mutant C. elegans have increased ROS levels. Although, we found no evidence that this resulted in reduced insulin-signaling or affected feeding behaviour, our data are consistent with other work establishing that sub-lethal increases in oxidants can increase stress resistance and longevity [[58], [59], [60], [61], [62], [63], [64]]. In this case, our data suggest that this increased survival under stress conditions may require the transcriptional regulator NHR-49. This adds to the evidence that this HNF4/PPARα-related nuclear hormone receptor's role in regulating lipid metabolism plays an important role in enhancing survival under various stress conditions [47]. Notably, increased MUFA have been shown to increase C. elegans lifespan at 20 °C, raising the possibility that the extended lifespan of prdx-6 mutants at 15 °C could also reflect their increased MUFA content [65,66]. Intriguingly, although fmo-2 is required for the lifespan-extending effects of hypoxia and dietary restriction at 20 °C, we reveal that at 15 °C fmo-2 mutants are themselves long lived. It will be interesting to determine whether this lifespan extension is due to effects on lipid metabolism.
Conserved immune pathways, such as the PMK-1 pathway, are important for the innate immunity of C. elegans to all pathogens [48,[67], [68], [69], [70], [71]]. However, transcriptomic data generated from worms infected with different pathogens show limited overlap in the genes that are up- and down-regulated [36,[48], [49], [50],72,37]. Thus, although there are some core immune pathways, unique responses are also generated in response to different infectious agents. Here we show that the Flavin monoxygenase, FMO-2, identified as part of the NHR-49 up-regulated response to S. aureus, E. faecalis and C. neoformans infection [36,37], also represents a component of the innate immune defence to the fungal pathogen C. albicans. Intriguingly, although overexpression of fmo-2 increased the survival of C. elegans infected with C. albicans, wild type and fmo-2 mutant C. elegans that survived the first day succumbed to infection at similar rates. Similarly, although NHR-49 is important, loss of fmo-2 alone does not increase the susceptibility of C. elegans to the gram negative bacteria Pseudomonas aeruginosa [73]. Thus, a combination of upregulated genes most likely protect prdx-6 mutant C. elegans against C. albicans. Similarly, it is likely that other genes upregulated in prdx-6 mutant animals contribute to their increased stress resistance and extended lifespan.
The mechanism by which FMO-2 protects against infection is unclear. Although increased fmo-2 extends the lifespan of C. elegans maintained on dead bacteria [35], our data are consistent with other work suggesting that levels of this Flavin monooxygenase also affects interactions with the microbiome [36,37]. Consistent with the increased oxidative stress resistance of prdx-6 mutant animals, high levels of FMO-2 protect against oxidative stress, potentially via increased stress-induced activation of the JNK pathway that plays important roles in stress resistance, apoptosis and innate immunity [74,75] or via effects on one carbon metabolism [76].
NHR-49 levels are regulated by transcriptional and post-transcriptional mechanisms [28,77]. Consistent with previous studies, we observed no increase in NHR-49 levels in response to infection [37] or in prdx-6 mutant C. elegans. However, the increased levels of monounsaturated fatty acids present in prdx-6 mutants is evidence of increased NHR-49 activity. It has been proposed that NHR-49's natural ligand might be an oxidized lipid [47]. Our data suggest that the phospholipid peroxidase activity of Prdx6 provides an important barrier to lipid peroxidation. Hence, it is possible that increased NHR-49 activity/fmo-2 expression in prdx-6 mutant C. elegans could reflect an increase in NHR-49's natural ligand.
Recent work has suggested that the main function of the redox-active cysteine in mammalian Prdx6 could be to mediate incorporation of selenium into selenoproteins, such as glutathione peroxidases [22]. The C. elegans genome encodes a single, selenoprotein, thioredoxin reductase, loss of which does not increase lifespan or innate immunity [25]. From this we conclude that the pro-survival phenotypes we describe for the prdx-6 mutant are unlikely to reflect any effect on selenocysteine metabolism. Instead, our study provides evidence that the thiol peroxidase activity of a 1-Cys Prdx can provide an important barrier to lipid hydroperoxides rendering prdx-6 mutants more sensitive to the acute toxicity of agents that increase lipid peroxidation. However, we reveal that Prdx6 also limits expression of fmo-2 and potentially other NHR-49-activated genes associated with increased lifespan and innate immunity. Significantly, this highlights the important positive role that low levels of specific ROS species can play in maintaining health.
Materials and methods
4
C. elegans strains, strain construction and maintenance
4.1
The following strains were used in this study: wild-type N2; VE2 prdx-2 (gk169) II; VC1668 fmo-2 (ok2147) IV; VE40 N2 eavEx20[fmo-2p::gfp + rol-6(su1006)]; VE43 nhr-49 (nr2041) eavEx20[fmo-2p::gfp + rol-6(su1006)]; LX960 lin-15B&lin-15A(n765) X;vsIs97 [tph-1p::DsRed2 + lin-15(+)]. C. elegans bearing prdx-6 (tm4225) IV and prdx-6 (tm4284) IV were obtained from the Mitani lab and outcrossed 6x with our wild-type N2 strain to generate VE44 prdx-6 (tm4225) IV and VE45 prdx-6 (tm4284) IV. VE40 and VE44 were outcrossed to generate VE46 prdx-6 (tm4225) eavEx20[(fmo-2p::gfp + rol-6(su1006)]. VE44 was crossing with VC1668 to generate VE47 prdx-6 (tm4225) IV fmo-2 (ok2147) IV. VE48 N2 eavEx21[prdx-6p::gfp + rol-6(su1006)] was generated by microinjection of wildtype (N2) with pPD95.67+prdx-6P::GFP (containing the 2.5 kb immediately upstream of the N-terminus of prdx-6 plus the first 22 amino acids of prdx-6 cloned in frame with GFP) together with the dominant co-injection marker rol-6 (su1006). pPD95_67 was a gift from Andrew Fire (Addgene plasmid # 1490; http://n2t.net/addgene:1490; RRID:Addgene_1490). VE49 N2 eavEx21[prdx-6p::gfp + rol-6(su1006)] vsIs97 [tph-1p::DsRed2 + lin-15(+)] was generated by crossing VE48 and LX960. VE40, VE43, VE46, VE48 and VE49 were maintained by selection of worms with the roller phenotype. MD701 bcIs39 [lim-7p:ced-1:GFP + lin-15 (+)] V was crossed with VE44 to generate VE50 prdx-6 (tm4225) IV bcIs39 [lim-7p:ced-1:GFP + lin-15 (+)] V or with VE45 to generate VE51 prdx-6 (tm4284) IV bcIs39 [lim-7p:ced-1:GFP + lin-15 (+)] V. VE44 or VE45 were crossed with KU25 pmk-1(km25) to generate VE52 prdx-6 (tm4225) pmk-1 (km25) and VE53 prdx-6 (tm4284) pmk-1 (km25) respectively. VE44 was crossed with CL2166 dvIs19[pAF15(gst-4::GFP::NLS)] to generate VE54 *prdx-*6 (tm4225) dvIs19[pAF15(gst-4::GFP::NLS)] and CF1553 muIs84 [(pAD76) sod-3p::GFP + rol-6] to generate VE55 prdx-6 (tm4225) muIs84 [(pAD76) sod-3p::GFP + rol-6] respectively. VE40, VE43, VE46, VE48 and VE49 were maintained by selection of worms with the roller phenotype. VE44 was crossed with STE68 nhr-49 (nr2041) I to generate VE56 prdx-6 (tm4225) IV nhr-49 (nr2041) I. C. elegans were maintained on NGM-L plates, unless otherwise indicated.
Imaging and analysis of animals expressing fluorescent protein-encoding transgenes by fluorescent microscopy
4.2
Animals were transferred into M9 buffer containing 0.06 % levamisole (Sigma L9756) for immobilization on 2 %–2.5 % (w/v) agarose pads. Fluorescence was observed using an epifluorescent Zeiss Axioskop 2 compound microscope and imaged under appropriate filters using AxioVision software (version 3.1.2.1). Intestinal expression of fmo-2p::gfp and sod-3p::gfp was assessed based on the number of GFP-positive intestinal nuclei as high (>12), medium (6-11), low (1–6) or none (no expression) as described previously [78]. CED-1:GFP engulfed corpses within the most visible arm of the gonad were counted.
Detection of reactive oxygen species by H2DCFDA staining
4.3
Approximately 500 synchronised young adult C. elegans were washed off a NGM-L plate in 1 ml of M9 buffer and transferred to a microfuge tube. Animals were allowed to settle for approximately 3 min to form a pellet before the supernatant was removed. Animals were washed with 1 ml of M9 buffer for 5 min two further times to remove any bacteria then incubated in 250 μl of 25 μM H_2_DCFDA diluted in M9 buffer, on a gently shaking platform in the dark for 30 min. Worms were allowed to settle and the supernatant removed. Worms were washed for 5 min with M9 buffer, the supernatant removed, and then a 30 min wash in M9 buffer followed by a final 5 min wash in M9 buffer. The supernatant was then removed and worms were gently resuspended in 20 μl of 0.06 % levamisole in M9 before mounting on agarose pads and imaging using a Zeiss Axioskop 2 fluorescent microscope.
Analysis of lifespan
4.4
L1 larval stage worms were allowed to develop to L4 larval stage at the described temperature before being transferred on to four separate plates. Worms maintained at 25 °C were moved to fresh plates daily, or on alternate days if at 15 °C or 20 °C, until progeny had finished being laid except in Fig. 7D (Table S11), where animals were transferred as young adults to NGM-L media supplemented with 25 μM 5-Fluoro-2′-deoxyuridine (FUDR) (Sigma) to prevent development of progeny instead. Survival was observed on the indicated days with worms not responding to prodding with a platinum wire scored as dead and removed from the plate.
Analysis of oxidative stress resistance
4.5
A minimum of 30 L4 larval stage worms were transferred to freshly prepared NGM-L plates containing 5 mM hydrogen peroxide (Sigma), 5–10 mM sodium arsenite or 15 mM diethyl maleate (Sigma) along with a small amount of E.coli (OP50). Survival was observed at the indicated time points with worms not responding to prodding with a platinum wire scored as dead and removed from the plate.
Infection experiments
4.6
Synchronised L4/Young adult animals were transferred on to an unseeded NGM-L plate for 1–1.5h before being transferred on to plates seeded with the pathogen. Survival was then monitored daily at 25 °C. For S. Typhimurium experiments cdc25.1 RNAi-treated L4/young adult animals were used. Staphylococcus aureus-seeded plates were prepared essentially as described previously [70]. A single colony of S. aureus (NCTC 8325) was inoculated in 10 ml of Tryptic Soy liquid media (Fluka) supplemented with 5ug/ml Nalidixic acid (Sigma-Aldrich) and grown overnight at 37 °C on a shaking platform. 10 μl of the S. aureus (NCTC8325) culture was spotted onto TS supplemented with 5ug/ml Nalidixic acid agar plates and plates were dried overnight at 37 °C. Candida albicans-infection experiments were carried out essentially as described by Ref. [48]. A single colony of C. albicans (SN148) was inoculated in 10 ml of Yeast Peptone Dextrose (YPD) liquid media, and grown overnight at 30 °C on a rotating wheel then 10 μl of this culture was seeded onto BHI media (BBL (6.0 g/L Brain Heart, Infusion from (solids), 6.0 g/L Peptic digest of Animal Tissue, 5.0 g/L Sodium Chloride. 3.0 g/L Dextrose, 14.5 g/L Pancreatic Digest of Gelatin, 2.5 g/L Disodium Phosphate) and 10 g/L Bacto Agar (BD) supplemented with 45ug/ml kanamycin (Sigma-Aldrich). For survival assays, S. aureus-seeded TS plates and C. albicans-seeded BHI plates were spotted with 8–16 μl of 40 mM 5′Fluoro-2′-deoxyuridine (FUDR) immediately prior to addition of C. elegans. Survival was observed at the indicated time points with worms not responding to prodding with a platinum wire scored as dead and removed from the plate.
Measuring lipid peroxidation
4.7
A mixed population of animals at 15 °C was washed with M9 buffer and incubated on plain NGM-L plates or plates supplemented with 15 mM diethyl maleate (Sigma) for the indicated length of time. Animals were washed and stained with 10 μM BODIPY 581/591 C11 (Invitrogen™) for 1 h, then washed three times in M9 buffer. Animals were then suspended in 0.06 % levamisole (Sigma), mounted on 3 % (w/v) agarose pads and imaged under 40x lens on a Zeiss Axioskop 2 fluorescent microscope using Axiovision software. Non-oxidized BODIPY-C11 was detected as emission at 607 nm following excitation at 538–558 nm and oxidized BODIPY was detected as emission at 525 nm following excitation at 450–490 nm and images collected with identical exposure time. Images were exported as TIFFs and analyzed in Image J where an anterior intestinal cell in each of ∼30 L4 stage animals per group was selected for analysis. Mean fluorescence was determined for green (oxidized) and red (non-oxidized) channels from the Pixel density for an identical area from which mean background values (determined from 10 to 12 unstained wild-type or prdx-6 mutant animals) were subtracted. Lipid peroxidation in each animal was then determined as green fluorescence/red fluorescence. This experiment was repeated with 20–30 animals analyzed per group in each experiment with similar results.
Oleic acid supplementation
4.8
NGM plates were prepared to include 0.1% (v/v) IGEPAL CA-630 and 0.8mM sodium oleate (Sigma).
Fatty acid analysis
4.9
Fatty acid composition was analyzed by gas chromatography/mass spectrometry (GC/MS) using the fatty acid methyl ester (FAME) method [79]. For each sample, roughly 400 worms were collected in water, allowed to settle, and most of the water removed before freezing the worm pellet. 2.5 % sulfuric acid in methanol was added to frozen pellets and then incubated at 70ᴼC for 1 h in a glass tube. FAMEs were extracted in hexane and separated using an Agilent 7890 GC/5975C MS in scanning ion mode equipped with a SP-1380 column. Relative amounts of fatty acid methyl esters are reported.
Statistical analysis
4.10
Experimental data shown in each figure panel are representative of at least three independent biological repeats. Log Rank analysis was used to identify any statistically significant differences in survival data. Chi^2^ tests were used to determine the significance of differences between the intestinal expression profiles for transcriptional reporters e.g. fmo-2::gfp under different conditions or in different genetic backgrounds. Student's T tests were used to analyse differences between the number of ced-1:GFP engulfed nuclei in different groups or differences between the level of lipid peroxides in different samples. For analysis of fatty acid composition, the relative composition of six independent populations of nematodes were analyzed using multiple T tests (Prism Graph), and statistical significance was determined using the Holm-Sidak method, with alpha = 5.000 %.
CRediT authorship contribution statement
Emma L. Button: Conceptualization, Formal analysis, Investigation, Methodology, Writing – original draft. Emilia Dwyer: Conceptualization, Formal analysis, Investigation, Methodology, Validation, Visualization, Writing – review & editing. Jake B. Lewis: Formal analysis, Investigation, Validation. Madison S. Mortensen: Formal analysis, Investigation. Elise McDonald: Investigation, Validation. Eloise Butler: Investigation. Fiona Pearson: Investigation. Alice E. Tang: Investigation, Methodology. Jennifer L. Watts: Conceptualization, Funding acquisition, Supervision, Visualization. Elizabeth A. Veal: Conceptualization, Data curation, Formal analysis, Funding acquisition, Project administration, Supervision, Visualization, Writing – original draft, Writing – review & editing.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Sarr D.Toth E.Gingerich A.Rada B.Antimicrobial actions of dual oxidases and lactoperoxidase J. Microbiol.5620183733862985882510.1007/s 12275-018-7545-1PMC 7336354 · doi ↗ · pubmed ↗
- 2Bolduc J.Koruza K.Luo T.Malo Pueyo J.Vo T.N.Ezerina D.Messens J.Peroxiredoxins wear many hats: factors that fashion their peroxide sensing personalities Redox Biol.42202110195910.1016/j.redox.2021.101959 PMC 811303733895094 · doi ↗ · pubmed ↗
- 3Eismann T.Huber N.Shin T.Kuboki S.Galloway E.Wyder M.Edwards M.J.Greis K.D.Shertzer H.G.Fisher A.B.Peroxiredoxin-6 protects against mitochondrial dysfunction and liver injury during ischemia-reperfusion in mice Am. J. Physiol. Gastrointest. Liver Physiol.2962009 G 266G 2741903353210.1152/ajpgi.90583.2008 PMC 2643922 · doi ↗ · pubmed ↗
- 4Wang X.Phelan S.A.Forsman-Semb K.Taylor E.F.Petros C.Brown A.Lerner C.P.Paigen B.Mice with targeted mutation of peroxiredoxin 6 develop normally but are susceptible to oxidative stress J. Biol. Chem.278200325179251901273262710.1074/jbc.M 302706200 · doi ↗ · pubmed ↗
- 5Choi H.Chang J.W.Jung Y.K.Peroxiredoxin 6 interferes with TRAIL-induced death-inducing signaling complex formation by binding to death effector domain caspase Cell Death Differ.1820114054142082988410.1038/cdd.2010.113PMC 3132003 · doi ↗ · pubmed ↗
- 6Kim S.Y.Chun E.Lee K.Y.Phospholipase A(2) of peroxiredoxin 6 has a critical role in tumor necrosis factor-induced apoptosis Cell Death Differ.182011157315832141586010.1038/cdd.2011.21PMC 3172113 · doi ↗ · pubmed ↗
- 7Karihtala P.Mantyniemi A.Kang S.W.Kinnula V.L.Soini Y.Peroxiredoxins in breast carcinoma Clin. Cancer Res.920033418342412960131 · pubmed ↗
- 8Lehtonen S.T.Svensk A.M.Soini Y.Paakko P.Hirvikoski P.Kang S.W.Saily M.Kinnula V.L.Peroxiredoxins, a novel protein family in lung cancer Int. J. Cancer 11120045145211523912810.1002/ijc.20294 · doi ↗ · pubmed ↗