Evolutionary History of the Adult β‐Globin Gene in Wild Sheep: First Sequences From the Argali (Ovis ammon) and the Urial (Ovis vignei)
Paolo Mereu, Chiara Multineddu, Daria Sanna, Marco Zedda, Antonio J. Lepedda, Giovanni G. Leoni, Monica Pirastru

TL;DR
This study provides the first complete adult β-globin gene sequences for Argali and Urial sheep, revealing evolutionary patterns and divergence in wild sheep species.
Contribution
The paper presents the first complete adult β-globin gene sequences for Argali and Urial, enabling new phylogenetic and adaptive studies in wild sheep.
Findings
Argali hosts two distinct adult β-globin haplotypes, one with HBBA and the other with HBBB.
Phylogenetic analysis shows sequences cluster by paralog type (HBBA vs. HBBB) in Caprinae and Bovinae.
Molecular dating estimates the HBBA/HBBB duplication event in Caprinae at around 2.55 million years ago.
Abstract
The β‐globin gene cluster is a key model for studying molecular adaptation and gene family evolution in vertebrates, exhibiting dynamic duplication and deletion events in ruminants, particularly within the subfamily Caprinae. However, the complete nucleotide sequences of the adult β‐globin gene remain unknown for several ecologically important wild sheep species, such as the Argali and the Urial, hindering robust phylogenetic and adaptive studies. Genomic DNA was extracted from wild Argali and Urial samples, and the full coding region of the adult β‐globin gene was sequenced and compared against a comprehensive dataset of Bovidae and Cervidae. Sequence comparison confirmed high divergence between paralogous genes (HBBA and HBBB), and critically, revealed that Argali hosts two distinct adult β‐globin haplotypes, one harboring a HBBA gene and the other a HBBB gene. Phylogenetic analysis…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 FIGURE 1
FIGURE 1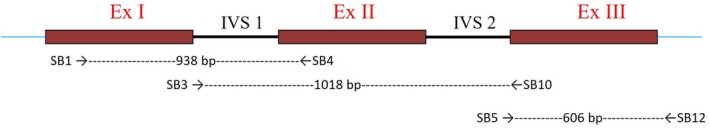 FIGURE 2
FIGURE 2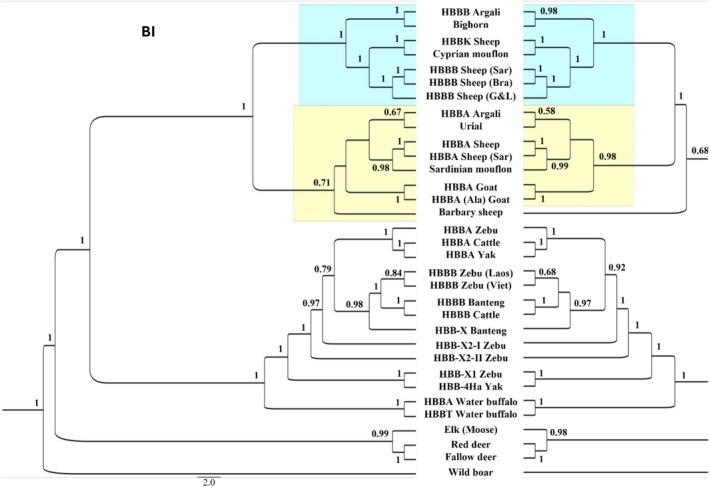 FIGURE 3
FIGURE 3| Species | Locus structure | Genes | Key evolutionary event |
|---|---|---|---|
| Cow | Duplicated | ϵ, ψβ, βA, βF | Ancestral duplication before Caprinae divergence |
| Goat | Triplicated | ϵ, ψβ, βC, βA, βF | Assumed ancestral state for Caprinae |
| A‐Haplotype sheep | Triplicated | ϵ, ψβ, βC, βA, βF | Retention of ancestral Caprinae locus |
| B‐Haplotype sheep | Duplicated | ϵ, ψβ, βB, βF | Recent deletion from a triplicated locus |
| Tibetan antelope | Duplicated | ϵ, ψβ, βC, βF | Secondary deletion from a triplicated locus |
| GenBank # | Species | Common name | Haplotype | Sampling location |
|---|---|---|---|---|
| This study |
| Argali | HBBB | Kazakhstan, Kara‐Tau mountain |
|
| HBBA | Kazakhstan, Kara‐Tau mountain | ||
|
| Bighorn | HBBB | Canada, Rocky mountain | |
|
| Urial | HBBA | Turkmenistan | |
|
| Barbary sheep | HBBA | Egypt | |
|
| Goat | HBBA | Italy, Sardinia | |
|
| HBBA(Ala) | n.a. | ||
|
| Domestic sheep | HBBA | Italy, Sardinia | |
|
| HBBB | Italy, Sardinia | ||
|
| HBBB | Brazil | ||
|
| HBBB | Garner & Lingrel (GL) | ||
|
| HBBK | Chios | ||
|
| Sardinian mouflon | HBBA | Italy, Sardinia | |
|
| Cyprian mouflon | HBBB | Cyprus | |
|
| Water buffalo | HBB(A) | Italy, Campania | |
|
| HBB(T) | Italy, Campania | ||
|
| Yak | HBB‐4Ha | Bhutan | |
|
| HBBA | Bhutan | ||
|
| Zebu | HBBA | Vietnam | |
|
| HBBA | Vietnam | ||
|
| HBBB | Vietnam | ||
|
| HBB‐X1 | Vietnam | ||
|
| HBB‐X2‐I | Vietnam | ||
|
| HBB‐X2‐I | Laos | ||
|
| HBB‐X2‐II | Vietnam | ||
|
| Banteng | HBBB | Bali | |
|
| HBBX | Bali | ||
|
| Cattle | HBBA | Japan | |
|
| HBBB | Japan | ||
|
| Elk | HBB | n.a. | |
|
| Red deer | HBB | n.a. | |
|
| Fallow deer | HBB | n.a. | |
|
| Wild boar | HBB | n.a. |
| CP | Divergence (MYA) | Substitution rate | |||
|---|---|---|---|---|---|
| Median | Adjusted time | C.I. | Mean + SD | Gamma | |
|
| 6 | 8.4 | 5.3–9.4 | 0.0011 ± 0.0022 | 0.1214 |
|
| 13.1 | — | 0.8–14.2 | 0.0006 ± 0.0019 | 0.1000 |
|
| 24.6 | 21.6 | 19.1–30.8 | 0.0011 ± 0.0041 | 0.2533 |
|
| 22.8 | 23.7 | 18.5–27.8 | 0.0012 ± 0.0042 | 0.3753 |
| Seq 1 | Seq 2 | SynSub | SynPos |
| NSynSub | NSynPos |
|
|
|---|---|---|---|---|---|---|---|---|
| Argali_HBBB | Argali_HBBA | 0.00 | 106.00 | 0.0000 | 2.00 | 332.00 | 0.0060 | 0 |
| Argali_HBBB | Bighorn | 0.00 | 106.00 | 0.0000 | 1.00 | 332.00 | 0.0030 | 0 |
| Argali_HBBB | Urial | 0.00 | 106.00 | 0.0000 | 2.00 | 332.00 | 0.0060 | 0 |
| Argali_HBBB | HBBB_Sheep | 2.00 | 105.42 | 0.0192 | 4.00 | 332.58 | 0.0121 | 0.63 |
| Argali_HBBB | HBBA_Sheep | 3.00 | 106.50 | 0.0287 | 4.00 | 331.50 | 0.0122 | 0.42 |
| Argali_HBBA | Bighorn | 0.00 | 106.00 | 0.0000 | 1.00 | 332.00 | 0.0030 | 0 |
| Argali_HBBA | Urial | 0.00 | 106.00 | 0.0000 | 0.00 | 332.00 | 0.0000 | 0 |
| Argali_HBBA | HBBB_Sheep | 2.00 | 105.42 | 0.0192 | 6.00 | 332.58 | 0.0183 | 0.95 |
| Argali_HBBA | HBBA_Sheep | 3.00 | 106.50 | 0.0287 | 2.00 | 331.50 | 0.0061 | 0.21 |
| Bighorn | Urial | 0.00 | 106.00 | 0.0000 | 1.00 | 332.00 | 0.0030 | 0 |
| Bighorn | HBBB_Sheep | 2.00 | 105.42 | 0.0192 | 5.00 | 332.58 | 0.0152 | 0.79 |
| Bighorn | HBBA_Sheep | 3.00 | 106.50 | 0.0287 | 3.00 | 331.50 | 0.0091 | 0.32 |
| Urial | HBBB_Sheep | 2.00 | 105.42 | 0.0192 | 6.00 | 332.58 | 0.0183 | 0.95 |
| Urial | HBBA_Sheep | 3.00 | 106.50 | 0.0287 | 2.00 | 331.50 | 0.0061 | 0.21 |
| HBBB_Sheep | HBBA_Sheep | 3.00 | 105.92 | 0.0289 | 8.00 | 332.08 | 0.0245 | 0.85 |
| Node | MYA | Height 95% HPD |
|---|---|---|
| Bovinae/Caprinae HBB | 4.16 | 2.28–6.32 |
| Caprinae HBBA/HBBB | 2.55 | 1.58–3.66 |
| Water buffalo HBBA‐HBBT/other Bovinae | 2.3 | 1.20–3.59 |
| Caprinae HBBA appearance | 1.31 | 0.5–2.22 |
| Caprinae HBBB appearance | 1.16 | 0.41–2.12 |
| Water buffalo HBBA/HBBT | 0.64 | 0.07–1.43 |
| Urial‐HBBA Argali/Sardinian mouflon‐HBBB Sheep | 0.56 | 0.16–1.11 |
| Cattle HBBA/HBBB | 0.54 | 0.19–0.97 |
| HBBB Argali/Bighorn | 0.46 | 0.06–1.04 |
| Cyprian mouflon—HBBK sheep/HBBB Sheep | 0.43 | 0.001–0.32 |
| Urial/HBBA Argali | 0.33 | 0.03–0.73 |
| Sardinian mouflon/HBBA sheep | 0.29 | 0.05–0.65 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetic and phenotypic traits in livestock · Hemoglobinopathies and Related Disorders · Genetic Syndromes and Imprinting
Introduction
1
The β‐globin gene family is known to be an effective model for studying molecular adaptation, gene regulation, and phylogenetic relationship in vertebrates (Storz 2018). Within ruminants (family Bovidae), its evolution has been particularly dynamic, characterized by recurrent, large‐scale duplication and deletion events followed by divergence and specialization that have resulted in striking lineage‐specific genomic architectures. Ruminants show classical hemoglobin (Hb) switching: The multiple copies of β‐globin genes are expressed at different developmental stages—embryonic, fetal, and adult—each with distinct functional properties tailored to the organism's oxygen requirements. While the bovine lineage (subfamily Bovinae) retains the ancestral ruminant pattern of a single adult β‐globin gene and a fused, pan‐specific ε/β‐like gene (MacEachern et al. 2009), the Caprinae subfamily lineage underwent a subsequent duplication event. This gave rise to a distinct genomic arrangement of the β‐globin gene cluster, named A‐haplotype, organized as a triplicated four‐gene set, totaling 12 genes, and is distinguished by the presence of a third β‐globin paralog, the preadult β^C^ (Rangan et al. 2021). Notably, the juvenile/preadult β^C^ and the adult‐expressed β^A^ globins display different oxygen‐affinity properties and can be differentially regulated, providing a refined mechanism for physiological adaptation. Adult sheep and goats exhibit well‐documented switching and some unusual regulatory dynamics, such as the reactivation of the preadult β^C^ gene under certain stress or pathological conditions (Pirastru et al. 2009).
The presence of this additional β^C^ gene block marks a critical point of divergence between Bovinae and Caprinae lineages. This triplicated arrangement has been described in goats ( Capra hircus Linnaeus, 1758), aoudad ( Ammotragus lervia Pallas, 1777), and in two Ovis species including Sardinian mouflon (Ovis gmelini musimon Pallas, 1811) and a subset of domestic sheep ( Ovis aries Linnaeus, 1758), known as the A‐haplotype sheep (Manca et al. 2006; Pirastru et al. 2009). The fact that this identical triplicated architecture is shared among these species provides strong evidence for a common ancestral state. Therefore, this triplicated locus was probably present in a common ancestor of Caprinae after its divergence from Bovinae.
The genus Ovis represents a fascinating group for evolutionary studies, having radiated across a wide array of ecological niches, from lowland deserts to high‐altitude mountain plateaus. This ecological diversity is reflected in their β‐globin gene cluster that, in addition to the triplicate arrangement above described, can also host a duplicated four‐gene set (B‐haplotype) harboring the adult β^B^ gene. This arrangement lacking the β^C^ gene is observed in a different group of domestic sheep (B‐haplotype sheep), Cyprian mouflon (O. gmelini ophion Blyth, 1841), Bighorn ( Ovis canadensis Shaw, 1804), and Snow sheep ( Ovis dalli Nelson, 1884) (Manca et al. 2006). Although the B‐haplotype sheep locus could bear a striking resemblance to the bovine locus, this similarity does not stem from a direct, recent shared ancestry. Indeed, the bovine duplicated locus arose from an ancestral duplication event that occurred before the divergence of cows and goats, estimated to be more than 18–20 million years ago (Jiang et al. 2015). Conversely, the B‐haplotype in sheep is believed to have resulted from a much more recent deletion from a triplicated locus. A detailed resequencing study of the B‐haplotype suggests it is at least 2–3 million years old, still a far more recent event than the deep evolutionary split between Bovinae and Caprinae. This suggests the shared duplicated state in cattle and B‐haplotype sheep as a case of convergent evolution or secondary loss, rather than retention of a shared ancient trait (Jiang et al. 2015). The different evolutionary timelines and mechanisms leading to the duplicated state highlight the distinct evolutionary pathways within these subfamilies. The complex relationships between these gene clusters are summarized in Table 1.
Protein‐coding genes like β‐globin genes, which are under selective pressure and exhibit a workable rate of evolution, are excellent candidates for shedding light on intricate phylogenetic relationships (Hardison 2012; Storz 2016). Their sequences can reveal not only phylogenetic signals but also potential signatures of positive selection related to environmental adaptation.
Despite their utility, the complete nucleotide sequences of the adult β‐globin gene from some wild Ovis species remain entirely unknown. The Argali ( Ovis ammon Linnaeus, 1758) and the Urial ( Ovis vignei Blyth, 1841) are two phylogenetically distinct and ecologically critical wild sheep species (Figure 1). The Argali thrives in the high‐altitude environments of Central Asia, while the Urial occupies a range of intermediate elevations across South and Central Asia. Their evolutionary relationships with each other and to domestic sheep are still refined using primarily mitochondrial DNA and neutral markers (Chen et al. 2021; Dotsev et al. 2023; Lv et al. 2024; Mereu et al. 2024).
Argali and Urial wild sheep. The pictures are freely available on the internet under Creative Commons License.
The absence of β‐globin gene sequences from these species represents a significant gap in our understanding. It prevents a robust phylogenetic analysis of a functional nuclear marker within Ovis and, more importantly, hinders investigations into the molecular basis of potential adaptive evolution.
Therefore, the objectives of this study were to: (i) isolate and sequence the complete coding regions of the adult β‐globin genes from the Argali and the Urial; (ii) characterize these sequences by comparing them to those of domestic sheep and other bovids to identify polymorphisms and potential functionally important substitutions; and (iii) perform a phylogenetic analysis to elucidate the evolutionary history of the β‐globin genes within the Caprinae subfamily. This study provides the first genetic data for this critically important gene family in Argali and Urial, establishing a foundation for future research into the molecular physiology of adaptation and the evolutionary dynamics of multigene families in wild species.
Material and Methods
2
Sample Collection
2.1
The biological samples (hairs) were obtained from one Urial (Turkmenistan), one Bighorn (Canada), and two Argali (Kazakhstan, Kara‐Tau mountain) individuals. This study adhered to ethical standards for animal research, which prioritize animal well‐being. All the animal procedures were performed in strict accordance with the guidelines of the Ethics Committee of Sassari University, Italy, which also approved this study.
DNA Extraction, Amplification, and Sequencing
2.2
Genomic DNA was extracted from hair using the NucleoSpin Tissue XS kit (Macherey‐Nagel, Germany), according to the manufacturer's protocol. DNA quality and concentration were assessed via spectrophotometry using a ND‐8000 (NanoDrop Technologies, Thermo Fisher Scientific Inc., Wilmington, DE, USA).
Oligonucleotide primers for the polymerase chain reaction (PCR) amplification were designed on the basis of sheep HBBA and HBBB sequences available on GenBank under the accession numbers DQ352470‐1 (Manca et al. 2006; Pirastru et al. 2003). A 938 bp fragment, from the promoter region to the second exon, was amplified using SB1 (5′‐AATAATCCATCCACATAGTCTTGAA‐3′, from positions −392 to −368 with respect to the Cap site) and SB4 (5′‐CCTCTTCTCCATTCTAAACTGT‐3′, from +572 to +593) primers. A 1018 bp fragment, from the second to the third exon, was amplified using SB3 (5′‐TGACTTCCTCTGACCTTGT‐3 from +229 to +247) and SB10 (GTCCACGGATTTCTCCAGGC‐3′ from +1273 to +1292) primers. The remaining portion of the gene was obtained using SB5 (5′‐TATTCTTGTGCTTCCCTTGTGG‐3′ from +1144 to +1165) and SB12 (5′‐TACAACCATCTTGTCATTGGG‐3′ from +1772 to +1792) primers (Figure 2). PCR products were purified by electrophoresis on a 1.2% agarose gel using the Montage DNA Gel extraction kit (Millipore, Burlington, MA, USA) and then sequenced by means of automated capillary sequencing using the PCR primers. Nucleotide sequencing was outsourced to Macrogen Europe, with each read performed in duplicate. This redundancy was implemented to maximize the precision of base calling and to mitigate potential sequencing errors. Processed sequences were visualized using FinchTV 1.4.0 (Geospiza Inc.) and assembled into contigs, after identifying overlapping areas on Clustal X 2 (Larkin et al. 2007).
PCR amplification strategy based on the use of three primer pairs to isolate the adult β‐globin gene sequences.
Homologous Sequences Retrieving and Analyses
2.3
BLAST queries with both the Argali and the Urial sequences were carried out against the GenBank database, and the dataset was complemented by adding 26 homologous sequences from Bovidae and three from Cervidae species. A further sequence from wild boar was downloaded and used as outgroup (Table 2).
Sequences were aligned using the Muscle algorithm implemented in MEGA v. 11 (Tamura et al. 2021). To check for selective pressure on the HBB gene, we estimated the numbers of synonymous and nonsynonymous nucleotide substitutions per site and the K a/K s ratios using the counting method by Nei and Gojobori (1986) implemented in DnaSP v.6 (Rozas et al. 2017).
Both the Bayesian Inference (BI) and Maximum Likelihood (ML) trees were inferred on the nucleotide sequences, using the software MrBayes v. 3.2.4 (Ronquist et al. 2012) and RAxML v. 8 (Stamatakis 2014), respectively, with 10,000 bootstrap replicates. The choice of the best model of nucleotide substitution, the GTR + G + I, was performed with MEGA v.11 on the basis of the BIC and AICc criterion values.
Molecular dating of the main splitting events within Bovidae was inferred using a Bayesian approach as implemented in BEAST v.1.10.4 (Suchard et al. 2018). Overall, a total of four calibration points (CPs) based on fossil records were used: three providing time estimates for nodes within Bovidae and one marking the divergence between Cervidae and Bovidae (Kumar et al. 2017) (Table 3).
Sequences were analyzed under the GTR + G + I model of sequence evolution, assuming 4 gamma categories, a Yule process speciation rate, and empirical base frequencies. We used a lognormal relaxed clock with uncorrelated rates (ULRC) approach. Two independent Markov chain Monte Carlo runs were carried out with 200,000,000 iterations each and sampled every 10,000 steps after a 10% burn‐in. The runs were combined after checking for convergence > 100. A maximum clade credibility (MCC) tree was generated using TreeAnnotator 1.10.4 and visualized with FigTree 1.4.4 (http://tree.bio.ed.ac.uk/).
Results
3
Four adult β‐globin gene sequences from three wild Ovis species—Argali, Urial and Bighorn –were produced in the present study and deposited in GenBank under the accession number PX441602‐5. The four sequences were quite similar in length, ranging from about 1820 to 1920 bp, and included the entire coding region plus partial 5′ and 3′ region ends. The two Argali sequences were 1828 bp and 1860 bp, respectively, the Urial 1917 bp and the Bighorn 1821 bp. Slight differences were detected at the level of intronic region I (IVS‐1), which in the Bighorn was 126 bp while in the other species 128 bp. More marked were those detected in intron II (IVS‐2), which was 902 bp long in the Urial and one of the two Argali, 900 bp in the Bighorn and 891 bp in the other Argali. The average nucleotide composition was 29.6 T, 21.3 C, 24.7 A and 24.3 G. The dataset was completed by downloading 30 additional homologous sequences from GenBank, for a total of 33 sequences.
Sequence Comparison, Genetic Distances Estimations and Selective Pressure
3.1
To quantify the genetic divergence between the newly sequenced β‐globin genes, we calculated pairwise genetic distances within Ovis under the Tamura 3‐parameter (T92) model of nucleotide evolution, assuming a rate variation among sites modeled by a gamma distribution (G = 1.02) (Table S1).
The greatest genetic distances were observed between the two paralogous genes, HBBA and HBBB, with values ranging from 0.01334 to 0.02410. The highest divergence was detected between the Sardinian mouflon/A‐haplotype sheep and the B‐haplotype sheep (0.02410 ± 0.00415). Within the same species, the two Argali sequences exhibited a distance of 0.01334 ± 0.00315, with one sequence found more similar to the HBBA sheep gene (0.0627 ± 0.00210) and the other to B‐haplotype sheep. Such evidence allowed us to infer that Argali species host two distinct arrangements of the β‐globin gene cluster, as previously described for domestic sheep.
The comparison between sequences of the same paralog (orthologs) showed a higher level of similarity across species. The HBBB gene showed low sequence divergence ranging from 0.00000 (HBBB Sheep Sar vs. HBBB Sheep Bra) to 0.00417 (HBBB Argali vs. Bighorn). Similarly, the HBBA gene showed a divergence ranging from 0.00277 (HBBA Argali vs. Urial) to 0.00487 (HBBA Argali vs. Sardinian mouflon).
The counting of synonymous and nonsynonymous substitutions is reported in Table 4. The ω = K a/K s ratios were < 1 in all the pairwise sequence comparisons, ranging between 0 and 0.95.
Phylogenetic Analysis
3.2
The BI and the ML trees showed the same topology, with the only exception of the Barbary sheep sequence, that was basal to all the Caprinae in the ML tree while clustering within HBBA clade in the Bayesian tree (Figure 3). The two phylogenetic trees were both completely resolved and with high statistical support to most of the nodes retrieved (PP for Bayesian tree and bootstrap values for ML tree), the only uncertainty being represented by the position of the Barbary sheep sequence. Within Caprinae group, sequences were split into two clades according to the β‐globin gene cluster haplotypes. One clade, highlighted in blue, included all the sequences from HBBB and HBBK sheep along with three wild species: the Cyprian mouflon, the Bighorn and the HBBB Argali (Figure 3). The first split in this clade separated the Bighorn and HBBB Argali from the other sequences, followed by the divergence of HBBK sheep and Cyprian mouflon from domestic sheep. The second clade (yellow) was composed of all sequences from goat and HBBA domestic sheep, along with three wild species: the Sardinian mouflon, the Urial and the HBBB Argali. The first split separated goat from the Ovis species that were grouped as follows: Argali and Urial on one side, and Sardinian mouflon and domestic sheep on the other. In the BI‐tree reconstruction, the Barbary sheep sequence was included within this second clade.
Evolution of the adult β‐globin gene from ruminant species based on the Bayesian Inference (BI; top left) and Maximum Likelihood (ML; top right) methods of tree reconstruction under the GTR model of nucleotide substitution. Nonuniformity of evolutionary rates among sites was modeled by using a discrete Gamma distribution (+G) with 5 rate categories and by assuming that a certain fraction of sites were evolutionarily invariable (+I). Within Caprinae, HBBA and HBBB clades were highlighted in yellow and blue, respectively.
The same pattern of sequences grouping was observed in the Bovinae subfamily where sequences were split into two major clades on the basis of the type of the β‐globin adult gene harbored by the species (Figure 3). Consequently, for example, the HBBA Cattle sequence clustered with HBBA Zebu and Yak rather than with HBBB Cattle. The Water buffalo HBBA and HBBT genes were found basal to the bovine HBBA/HBBB split, followed by the HBB‐X genes.
Molecular Dating of Beta Globin Gene Divergence
3.3
To elucidate the evolutionary timeline of the β‐globin genes within Bovidae, we performed a Bayesian molecular dating analysis on the complete nucleotide sequences of the adult β‐globin gene (HBBA and HBBB). The analysis yielded well‐supported divergence time estimates for key nodes within the phylogeny, with results summarized in Table 5.
The deepest divergence within our dataset, that between the bovine and caprine lineages (Bovinae/Caprinae), was estimated to have occurred in the mid‐Miocene, at 4.16 million years ago (MYA) (95% HPD: 2.28–6.32 MYA). Subsequently, the duplication event giving rise to the HBBA and HBBB paralogs within the Caprinae lineage was dated to the late Pliocene, at 2.55 MYA (95% HPD: 1.58–3.66 MYA).
The divergence between the major caprine genera, Capra (goat) and Ovis (sheep), was estimated at 1.31 MYA (95% HPD: 0.5–2.22 MYA), placing this event in the early Pleistocene.
Within the genus Ovis, our analysis revealed recent and rapid radiation. The split between the Sardinian mouflon and domestic sheep was the most recent, estimated at 0.29 MYA (95% HPD: 0.05–0.65 MYA). The divergence between the Urial and the Argali was also very recent, occurring at 0.33 MYA (95% HPD: 0.03–0.73 MYA), while the split between the Argali and the Bighorn sheep was dated to 0.46 MYA (95% HPD: 0.06–1.04 MYA). The node uniting the Argali‐Bighorn clade with the Cyprian mouflon‐domestic sheep clade was estimated at 1.16 MYA (95% HPD: 0.41–2.12 MYA).
Within the Bovinae, the duplication event leading to the HBBA and HBBB genes in cattle ( Bos taurus ) was estimated to have occurred 0.54 MYA (95% HPD: 0.19–0.97 MYA). Similarly, the divergence between the HBBA and HBBT paralogs in the water buffalo was dated to 0.64 MYA (95% HPD: 0.07–1.43 MYA). The splitting event between cattle and water buffalo at the HBBA locus was estimated at 2.3 MYA (95% HPD: 1.20–3.59 MYA).
Discussion and Conclusions
4
In this study, we sequenced the entire adult β‐globin gene from three wild Ovis species—Argali, Urial, and Bighorn—with the first two never being sequenced before, and performed a comprehensive comparative phylogenetic analysis to elucidate the evolutionary history of this gene family within Bovidae. Our findings provide robust insights into the phylogenetic relationships and divergence timeline of the β‐globin genes in caprines and bovines.
The analysis of synonymous and non‐synonymous substitutions in the coding portion of the β‐globin gene has highlighted the presence of a purifying selection removing deleterious substitutions and maintaining the sequence highly conserved between domestic and wild sheep. Indeed, the ratio K a/K s quantifying the rate of evolution by stating efficient evolutionary changes in relation to silent background evolutionary changes, and also reflecting the selection pressure on the evolution of organisms, was found to be lower than 1, indicating a negative selection with a reduced rate of fixation of amino acid changes.
The sequence comparison revealed that the greatest divergence was between the paralogous genes HBBA and HBBB, as expected. More notably, the analysis within the Argali species uncovered the presence of two distinct β‐globin sequences, one clustering with the HBBA haplotype and the other with the HBBB haplotype of domestic sheep. This finding strongly suggests that the Argali, like domestic sheep, possesses two distinct arrangements of the β‐globin gene cluster. This implies that the genetic polymorphism underlying the two haplotypes is an ancestral trait within the genus Ovis, predating the domestication of sheep.
The phylogenetic reconstructions using both BI and ML methods yielded highly congruent and well‐supported trees, with a single discrepancy concerning the placement of the Barbary sheep sequence. The consistent topology confirms a deep evolutionary split within Caprinae into two major clades defined by the type of adult β‐globin gene. The first clade (blue, Figure 3) comprises species carrying the HBBB/HBBK genes, including the Bighorn and the HBBB Argali sequence, while the second clade (yellow, Figure 3) includes species with the HBBA gene, such as the Urial, the other Argali sequence, and the Sardinian mouflon. This pattern of clustering by gene type rather than strictly by species taxonomy was mirrored within the Bovinae subfamily, where sequences grouped according to their paralog (HBBA, HBBB, or HBBT). This consistent phylogenetic signal across two subfamilies underscores the independent evolutionary trajectories of these paralogous genes following duplication, a phenomenon likely driven by concerted evolution or gene conversion within each lineage.
The molecular dating analysis provided a strong temporal framework for these evolutionary events. The estimated divergence between the Bovinae and Caprinae β‐globin gene lineages in the mid‐Miocene (~4.16 MYA; height 95% HPD: 2.28–6.32) does not align with established fossil records for the diversification of the Bovidae family that date back to ~20 MYA (Bibi 2013). This would indicate that such diversification occurred after the separation of the two groups, independently in each lineage. A key finding is the timing of the duplication event that gave rise to the HBBA and HBBB paralogs within the Caprinae lineage, which we dated to the late Pliocene (~2.55 MYA; height 95% HPD: 1.58–3.66). This estimate is consistent with the result inferred by the analysis of mitochondrial and nuclear markers on the Ovis appearance estimated at 2–3 MYA, previously reported (Bibi et al. 2012; Rezaei et al. 2010). However, more recent analyses based on the whole mitogenome sequences dated back the Ovis early radiation to 4.6 MYA (Mereu et al. 2025). Such a discrepancy could be explained by assuming that the radiation of the genus Ovis event significantly predates the more recent and rapid diversification of the β‐globin gene cluster during the Pleistocene.
Similarly, within Bovinae, the duplication events in Cattle (~0.54 MYA; height 95% HPD: 0.19–0.97) and Water buffalo (~0.64 MYA; height 95% HPD: 0.07–1.43) are also recent, occurring long after the split of the bovine ancestral β‐globin gene (~2.3 MYA). This indicates that the HBBA/HBBB paralogs in Cattle and the HBBA/HBBT paralogs in Water buffalo are the result of independent, lineage‐specific duplication events, further highlighting the dynamic evolution of the β‐globin cluster in ruminants (Dutta et al. 2020).
In conclusion, even taking into account the constrained sampling, our integrated analysis of sequence variation, phylogeny, and molecular dating demonstrates that the evolution of the β‐globin gene family in Bovidae is characterized by ancient paralogous diversification followed by recent conspecific radiation. The confirmation of two β‐globin haplotypes in the wild Argali provides important context for understanding the genetic basis of Hb variation in domestic sheep. The well‐resolved phylogeny and specific divergence time estimates presented here offer a valuable resource for future studies on mammalian molecular evolution and the evolutionary history of the genus Ovis.
Author Contributions
Paolo Mereu: conceptualization (lead), formal analysis (equal), investigation (equal), writing – original draft (lead), writing – review and editing (equal). Chiara Multineddu: conceptualization (equal), investigation (lead), writing – review and editing (equal). Daria Sanna: formal analysis (equal), investigation (equal), writing – review and editing (equal). Marco Zedda: formal analysis (equal), supervision (equal), writing – review and editing (equal). Antonio J. Lepedda: formal analysis (equal), writing – review and editing (equal). Giovanni G. Leoni: formal analysis (equal), supervision (lead), writing – review and editing (equal). Monica Pirastru: formal analysis (equal), investigation (equal), writing – review and editing (equal).
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Table S1: Estimates of evolutionary divergence between Ovis species sequences.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bibi, F. 2013. “A Multi‐Calibrated Mitochondrial Phylogeny of Extant Bovidae (Artiodactyla, Ruminantia) and the Importance of the Fossil Record to Systematics.” BMC Evolutionary Biology 13: 166. 10.1186/1471-2148-13-166.23927069 PMC 3751017 · doi ↗ · pubmed ↗
- 2Bibi, F. , E. Vrba , and F. Fack . 2012. “A New African Fossil Caprin and a Combined Molecular and Morphological Bayesian Phylogenetic Analysis of Caprini (Mammalia: Bovidae).” Journal of Evolutionary Biology 25: 1843–1854. 10.1111/j.1420-9101.2012.02572.x.22816969 · doi ↗ · pubmed ↗
- 3Chen, Z.‐H. , Y.‐X. Xu , X.‐L. Xie , et al. 2021. “Whole‐Genome Sequence Analysis Unveils Different Origins of European and Asiatic Mouflon and Domestication‐Related Genes in Sheep.” Communications Biology 4: 1307. 10.1038/s 42003-021-02817-4.34795381 PMC 8602413 · doi ↗ · pubmed ↗
- 4Dotsev, A. , O. Koshkina , V. Kharzinova , et al. 2023. “Genome‐Wide Insights Into Intraspecific Taxonomy and Genetic Diversity of Argali (Ovis ammon).” Diversity 15: 627. 10.3390/d 15050627. · doi ↗
- 5Dutta, P. , A. Talenti , R. Young , et al. 2020. “Whole Genome Analysis of Water Buffalo and Global Cattle Breeds Highlights Convergent Signatures of Domestication.” Nature Communications 11: 4739. 10.1038/s 41467-020-18550-1.PMC 750598232958756 · doi ↗ · pubmed ↗
- 6Hardison, R. C. 2012. “Evolution of Hemoglobin and Its Genes.” Cold Spring Harbor Perspectives in Medicine 2: a 011627. 10.1101/cshperspect.a 011627.23209182 PMC 3543078 · doi ↗ · pubmed ↗
- 7Jiang, Y. , X. Wang , J. W. Kijas , and B. P. Dalrymple . 2015. “Beta‐Globin Gene Evolution in the Ruminants: Evidence for an Ancient Origin of Sheep Haplotype B.” Animal Genetics 46: 506–514. 10.1111/age.12318.26096044 · doi ↗ · pubmed ↗
- 8Kumar, S. , G. Stecher , M. Suleski , and S. B. Hedges . 2017. “Time Tree: A Resource for Timelines, Timetrees, and Divergence Times.” Molecular Biology and Evolution 34: 1812–1819. 10.1093/molbev/msx 116.28387841 · doi ↗ · pubmed ↗