The Ies6 subunit is essential for INO80-mediated nucleosome organization
Ashish Kumar Singh, Felix Mueller-Planitz

TL;DR
The Ies6 subunit is crucial for organizing nucleosomes in yeast, and its absence disrupts chromatin structure and interacts with another remodeler.
Contribution
This study reveals the essential role of Ies6 in INO80-mediated nucleosome organization and its functional redundancy with ISW2.
Findings
Ies6 deletion reduces nucleosome spacing by 3 bp and disrupts nucleosome arrays.
IES6 deletion is synthetically lethal with ISW2 deletion, showing functional overlap.
INO80 binding directly correlates with Ies6's role in nucleosome organization.
Abstract
ATP-dependent nucleosome remodelers of the INO80 family regulate chromatin by sliding, spacing and positioning nucleosomes. The INO80 remodeler is organized into structural modules that regulate its remodeling activity. Here, we investigate the role of the Ies6 subunit within the Arp5-Ies6 module towards nucleosome positioning and spacing in Saccharomyces cerevisiae. We show that Ies6 is critical for establishing genome-wide nucleosome organization. Deletion of IES6 reduces nucleosome spacing by 3 bp and disrupts regular nucleosome arrays across most genes. Surprisingly, deletion of IES6 is synthetically lethal with the deletion of ISW2, a remodeler from the ISWI family, indicating functional redundancy in nucleosome organization. Notably, INO80 binding directly predicts the role of Ies6 in INO80-mediated nucleosome organization, whereas changes in gene expression do not correlate with…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
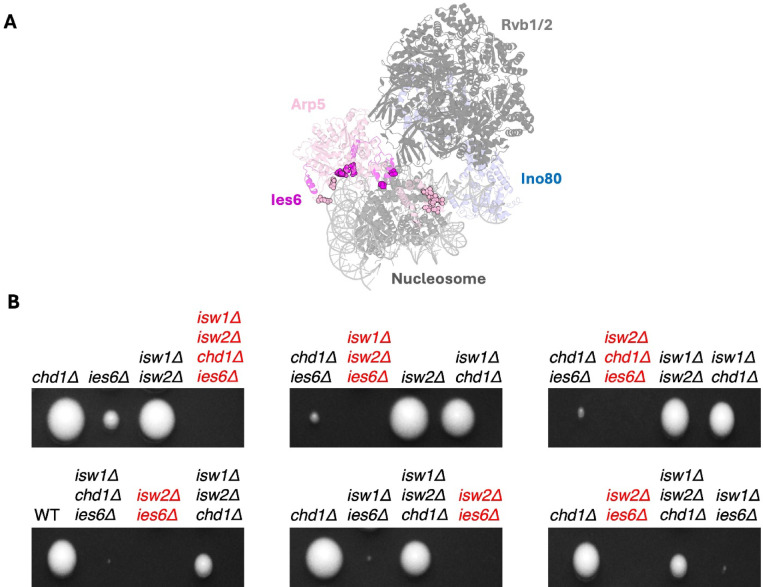 Figure 1
Figure 1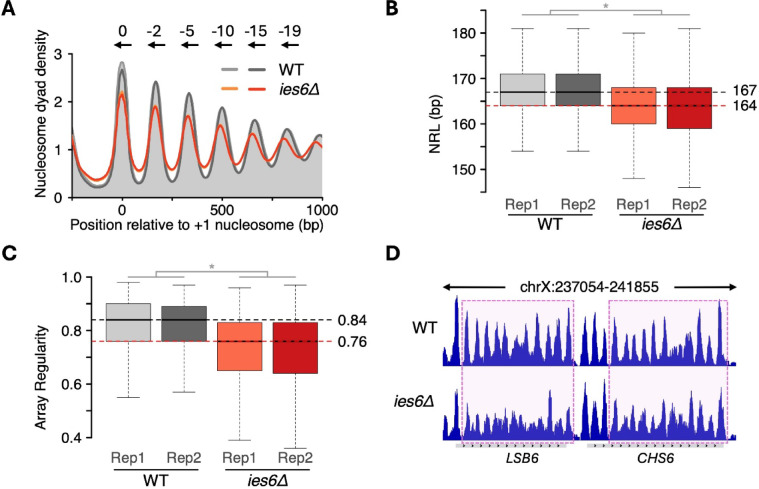 Figure 2
Figure 2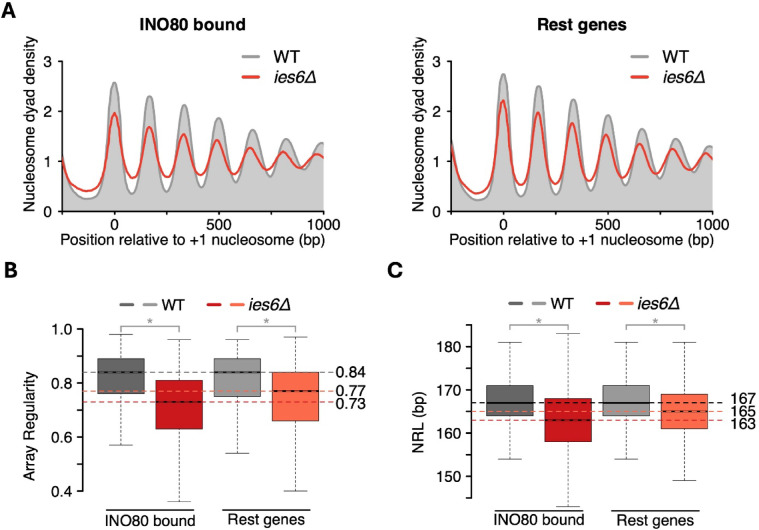 Figure 3
Figure 3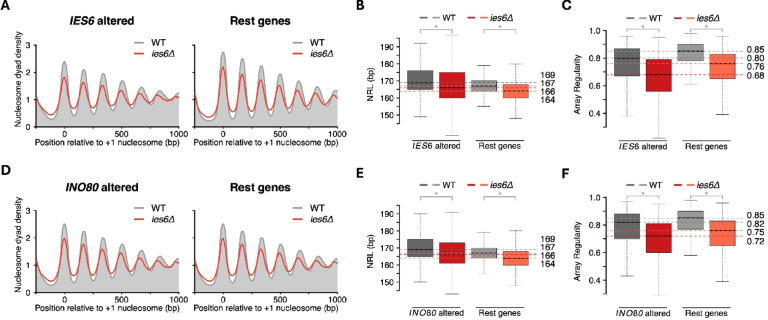 Figure 4
Figure 4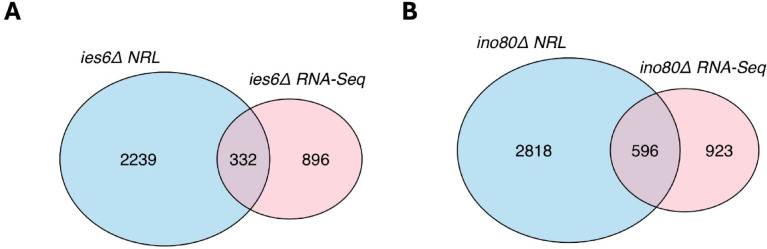 Figure 5
Figure 5- —Technische Universität Dresden (1019)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Chromatin Dynamics · Developmental Biology and Gene Regulation · Chromatin Remodeling and Cancer
Introduction
Genomic DNA is wrapped around nucleosomes, which affects all genomic processes involving DNA transactions^1,2^. Nucleosomes attain a stereotypical architecture downstream of the promoter region where equally spaced nucleosomes, known as regular nucleosome arrays, are aligned next to the transcription start site (TSS). The distance between two adjacent nucleosomes in a regular nucleosome array is referred to as the nucleosome repeat length (NRL)^3,4^. How cells establish and maintain this organization in the face of disruptive transcription and replication processes is an active area of research. ATP-dependent nucleosome remodeling complexes play pivotal role in this process^5^.
These nucleosome remodeling complexes can evict, slide and space nucleosomes as well as exchange canonical histones with histone variants. They can be classified into four main families: SWI/SNF, ISWI, CHD and INO80^5,6^. The INO80 remodeler plays essential regulatory roles in multiple processes including transcription, replication, DNA damage response and double-strand break repair^7–9^. Together with the ISWI and CHD families, INO80 has the ability to slide and equally space nucleosomes in vitro^10–13^. In cells, INO80 positions the first nucleosome (+ 1 nucleosome) downstream of the TSS and can generate regular nucleosome arrays^14,15^.
The S. cerevisiae INO80 remodeler is a 15-subunit complex, consisting of the Ino80 ATPase subunit and three major regulatory modules—the Nhp10, Arp8 and Arp5/Ies6 modules^16,17^. The Nhp10 and Arp8 modules can associate with extranucleosomal DNA and are proposed to act as sensors of linker DNA length and regulators of the enzymatic activity of INO80^18–21^. Consistent with such a regulatory role, deletion of ARP8 in S. cerevisiae resulted in a shortened NRL, implicating that the Arp8 module in regulating the nucleosome spacing activity of INO80. Despite its clear involvement in sensing linker length in vitro^20^, deletion of NHP10 however did not alter the NRL, suggesting that this module is dispensable for nucleosome spacing^15^.
In contrast to Arp8 and Nhp10 modules, the Arp5-Ies6 module acts as a central core component of the INO80 remodeler (Fig. 1A). It binds to DNA and histone moieties of the nucleosome core particle. It also contacts the acidic patch on the H2A-H2B dimer^22,23^, which acts as an allosteric activator of INO80 activity. As such, the Arp5-Ies6 module appears to play an important role in regulating ATP hydrolysis and nucleosome sliding by the INO80 complex, as demonstrated in vitro with mono-nucleosome substrates. A mutant INO80 complex lacking Arp5-Ies6 is still able to bind and slide nucleosomes, albeit with decreased efficiency^17,24–27^.
Fig. 1. Deletion of IES6 is synthetically lethal with ISW2 and genetically interacts with ISW1 and CHD1. (A) Structure of the S. cerevisiae INO80 complex bound to the nucleosome. Selected subunits are highlighted. Spheres denote residues from Ies6 (magenta) and Arp5 (light pink) that interact with the nucleosome. PDB code: 9C9G. (B) Representative tetrad dissection of diploids arising from TKO x ies6Δ. Tetrads were dissected on a YPAD plate, colonies grown for five days and replica-plated on minimal media lacking appropriate amino acids or on full media with cloNAT selection marker to identify genes lacking in the colonies. For additional results, see Supp. Fig. S1.
In yeast, deletion of either the Arp5 or Ies6 subunit is suggested to lead to a loss of the entire module from the INO80 complex without affecting the remaining subunits^17,28,29^. These mutants display disrupted yeast metabolic cycle and mitochondrial maintenance that parallel, but do not completely recapitulate the phenotypes observed upon deletion of the Ino80 ATPase subunit^30,31^. Moreover, loss of the Ies6 or of the Ino80 subunits results in increased ploidy due to defective nucleosome organization near centromeres^32^. At the transcriptome level, deletion of Arp5 or Ies6 alters the expression of more than 1,000 genes. Deletion of the Ino80 ATPase subunit produces an even more severe phenotype^25,33^. These findings suggest that an INO80 complex lacking the Arp5–Ies6 module retain partial remodeling activity. Consistent with this, the Arp5–Ies6 module has been proposed to carry out both specialized roles within the INO80 complex and potentially independent functions outside of it in yeast and cancer cells^30,34^.
Here, we test the role of Ies6 subunit in regulating nucleosome positioning and spacing in S. cerevisiae by analyzing the phenotypes associated with IES6 deletion. Unexpectedly, we find that ies6Δ is synthetically lethal with isw2Δ, but not with isw1Δ and chd1Δ, suggesting that INO80 and ISW2 may regulate a similar subset of genes. Deletion of IES6 decreased nucleosome positioning, NRL and regular arrays genome-wide. Surprisingly, changes in gene expression detected by RNA-seq upon deletion of IES6 do not predict alterations to the chromatin architecture following the deletion, indicating that transcriptional alterations do not directly account for the chromatin defects observed in ies6Δ cells.
Results
Deletion of IES6 is synthetically lethal with deletion of ISW2 and shows genetic interactions with ISW1 and CHD1
Nucleosome remodelers often act redundantly to position and space nucleosomes. We and others have previously shown that INO80 can contribute to nucleosome spacing within gene bodies of S. cerevisiae, in addition to Isw1 and Chd1 spacing remodelers^14,15,26,35–39^. The role of the Arp5-Ies6 module in nucleosome organization by INO80 remains poorly understood. It forms multiple interactions with the nucleosome (Fig. 1A) and has been suggested to grip on to the nucleosome while the ATPase domain pumps DNA^23^. Since ARP5 deletion is lethal in W303 yeast strains^40^, we focused on IES6 in this study. IES6 could be readily deleted through homologous recombination in W303 wild-type cells indicating that it is not an essential gene (for strain list, see Supp. Table 1). Consistent with previous results, deletion of IES6 gave rise to slow-growing colonies^32^ (Fig. 1B, Supp. Fig. S1).
Since the presence of other spacing remodelers (Isw1, Isw2 and Chd1) obscures INO80 activity in vivo^15^, we attempted to delete IES6 also in a triple knock out (TKO) strain carrying deletions of CHD1 and both ISWI factors (isw1Δ isw2Δ chd1Δ)^41^. Despite multiple attempts, we failed to delete the IES6 gene in TKO cells via homologous recombination. Therefore, we hypothesized that deletion of IES6 may be essential in the TKO but not WT cells, or in other words, that IES6 shows synthetic lethality with one or more of the remodelers deleted in TKO cells.
To test for synthetic lethality of Ies6 with ISW1, ISW2 and Chd1 remodelers, we generated diploid cells by mating haploid strains lacking IES6 (MATα) with the TKO strain (MATa) and performed tetrad dissection. If IES6 becomes essential in absence of any of these remodelers, we would expect to recover no viable spores carrying the corresponding double, triple, or quadruple deletions. Indeed, no viable colonies were recovered with the quadruple deletion (ies6Δ isw1Δ isw2Δ chd1Δ) or with the double deletion of ies6Δ isw2Δ (Fig. 1B, Supp. Fig. S1; lethal combinations in red). In contrast, double deletions of ies6Δ with either isw1Δ or chd1Δ produced viable, albeit small colonies. Similarly, the triple deletion of ies6Δ isw1Δ chd1Δ also resulted in small colonies, whereas combinations such as ies6Δ isw2Δ isw1Δ, and ies6Δ isw2Δ chd1Δ were not recovered (Fig. 1B, Supp. Fig. S1). Taken together, loss of viability in the ies6Δ TKO strain is driven by a synthetic lethal relationship between IES6 and ISW2, with ISW1 and CHD1 playing lesser roles.
Ies6 is essential for INO80 nucleosome positioning and spacing activity
To investigate the role of Ies6 in nucleosome organization, we performed micrococcal nuclease sequencing (MNase-Seq) on WT and two independent ies6Δ colonies. Deletion of IES6 led to a genome-wide decrease in nucleosome positioning. Compared to WT cells, the + 1-nucleosome and the downstream nucleosomes in the gene body were less well positioned as seen in gene-averaged composite plots. Furthermore, gene body nucleosomes were shifted upstream towards the TSS, suggesting that ies6Δ cells possess on average a shorter NRL (Fig. 2A). Fig. 2. Deletion of IES6 leads to genome-wide decrease in nucleosome positioning and spacing in S. cerevisiae. (A) Average nucleosome organization for all 5015 genes in the indicated yeast strains. Arrows indicate upstream shift in bp for each nucleosome in ies6Δ cells relative to WT. Two replicates are shown for WT and ies6Δ cells. (B) Boxplots showing NRL distribution in all yeast genes in WT and ies6Δ cells for two independent replicates, Rep1 and Rep2. (C) Same as (B), but for array regularity. Horizontal dotted lines indicate the average median values in WT (gray) and ies6Δ cells (red). (D) Genome browser snapshot over two representative genes in WT and ies6Δ cells. Nucleosomes showing major change in nucleosome organization relative to WT are highlighted. To quantify changes to the structure of nucleosome arrays over each gene, we applied a previously published method for calculating NRL and array regularity^15,37^. This method cross-correlates MNase-Seq profiles around the TSS of each gene with a series of idealized Gaussian patterns of varying NRLs (Methods). Regularly spaced arrays that are aligned (phased) to TSSs are detected this way. The NRL for each gene is obtained from the best fitting Gaussian pattern and the correlation coefficient of the fit is used as an indicator of regularity of the phased arrays (Supp. Fig. S2A). We found that the NRL decreased by 3 bp upon IES6 deletion relative to WT (Fig. 2B). In addition, array regularity also declined, consistent with the reduced nucleosome positioning observed in composite plots (Fig. 2C). These genome-wide trends were also evident at individual genes (Fig. 2D). Collectively, these results demonstrate that the Ies6 subunit is critical for establishing nucleosome organization in vivo.
We next asked to what extent IES6 deletion phenocopies loss of the INO80 ATPase subunit. We previously showed that acute depletion of INO80 in W303 cells decreases the NRL by 3 bp^15^, mirroring the decrease in NRL upon IES6 deletion (Fig. 2B). Likewise, array regularity also showed a significant decrease upon depletion of the INO80 subunit (Supp. Fig. S2B).
To bolster these results that were obtained in the W303 yeast background, we inspected nucleosome array structure in BY4741 cells lacking the Ino80 ATPase using a published dataset^26^. Consistent with results in the W303 background, deletion of Ino80 ATPase in BY4741 cells reduced NRL by 3 bp and decreased array regularity (Supp. Fig. S2C, D). Importantly, we observed a substantial overlap between genes showing decreased NRL in ies6Δ and in cells lacking the INO80 complex (Supp. Fig. S2E, F). Overall, these findings indicate that the INO80 remodeler regulates nucleosome organization in S. cerevisiae and that the Ies6 subunit plays an essential role within the INO80 complex, closely phenocopying the loss of the entire INO80 complex.
Ino80 binding, rather than expression changes, predicts altered nucleosome organization following IES6 deletion
To assess the causal basis of the effects observed upon IES6 deletion, we compared INO80-bound genes with all other genes^14^. If Ies6 is important for INO80 activity, IES6 deletion should produce a stronger disruption at INO80-bound genes than across the rest of the genome. Consistent with this expectation, INO80-bound genes showed a greater reduction in array regularity relative to non-bound genes following IES6 deletion (Fig. 3A, B). INO80-bound genes also experienced a larger drop in the NRL (4 bp) compared to the remaining genes (2 bp; Fig. 3C).
Fig. 3INO80-bound genes exhibit a greater reduction in nucleosome positioning and spacing than other genes following IES6 deletion. (A) Average nucleosome organization for all 1646 INO80 bound (left) and rest (right) of the genes in the indicated yeast strains. (B) Boxplots showing array regularity distribution in the same set of genes as in (A). (C) Boxplots showing NRL distribution in the same set of genes as in (A). Horizontal dotted line indicates the median NRL or array regularity in WT (gray) and ies6Δ (red) cells.
Analogous results were obtained when we tested the effect of the loss of the INO80 subunit: INO80 bound genes displayed a 4 bp decrease in NRL and a greater loss of array regularity compared to the rest of the genes (Supp. Fig. S3A, B, C)^15^. These findings support a model in which INO80 induces regular nucleosome spacing across the gene bodies it binds, with Ies6 serving as a pivotal subunit in this activity.
Motivated by these results, we next tested for a potential relationship between gene expression changes and changes in nucleosome organization over gene bodies upon Ies6 or Ino80 ablation. To test for such a relationship, we used published RNA-Seq datasets for ies6Δ and ino80Δ cells^25,33^ and compared nucleosome features between genes with altered expression and the rest of the genome. Surprisingly, no preferential effects were observed. Following deletion of IES6, both differentially expressed and non-differentially expressed genes showed a similar decrease in NRL and array regularity (Fig. 4A-C, Supp. Fig. S4A, B). Similarly, genes with altered expression in ino80Δ cells did not exhibit preferential changes in nucleosome organization upon IES6 deletion (Fig. 4D-F, S4C, D). Lastly, we examined nucleosome organization in ino80Δ BY4741 cells and again observed no preferential differences between genes with altered expression upon either IES6 or INO80 deletion (Supp. Fig. S5A-F). These results suggest that gene expression changes are not reliable indicators of altered nucleosome organization over gene bodies in ies6Δ or ino80Δ cells. Or, put differently, that an altered array structure over gene bodies in the deletion mutants is not predictive of changes in gene expression levels.
Fig. 4. Gene expression changes upon IES6 deletion do not correlate with changes in nucleosome architecture. (A) Average nucleosome organization in WT and ies6Δ cells for all 1304 genes with at least 1.5-fold change in gene expression in ies6Δ cells (left) and remaining genes (right). List of differentially expressed genes was obtained from^33^. (B) Boxplots showing the NRL distribution in the same set of genes as in (A). (C) Boxplots showing array regularity distribution in the same set of genes as in (A). (D) Average nucleosome organization in WT and ies6Δ cells for all 1519 genes with at least 1.5-fold change in gene expression in ino80Δ cells (left) and for the remaining genes (right). List of differentially expressed genes was obtained from^33^. (E) Boxplots showing the NRL distribution in the same set of genes as in (D). (F) Boxplots showing array regularity distribution in the same set of genes as in (D). Horizontal dotted line indicates the median NRL or array regularity in WT (gray shades) and ies6Δ (red shades) cells.
To further examine the relationship between the structure of nucleosome arrays and gene expression, we next compared genes with altered NRL to those with altered gene expression in ies6Δ and ino80Δ cells. Notably, we observed only a small overlap between genes showing NRL changes and those with differential expression as measured by RNA-Seq for both mutants (Fig. 5A, B). Even a stringent cutoff (> 5 bp change in NRL) did not improve the overlap between the two gene sets (Supp. Fig. S6A, B). Taken together, these findings suggest that the limited overlap between NRL changes and RNA-seq data explains the lack of a direct effect of IES6 or INO80 deletion on nucleosome organization.
Fig. 5. Genes with altered expression and NRL in IES6 and INO80 lacking cells show a partial overlap. (A) Venn diagram showing overlap of genes with altered RNA levels and NRL in ies6Δ cells. (B) Venn diagram showing overlap of genes with altered RNA levels and NRL in ino80Δ cells.
Discussion
Our study uncovers an essential role for the Ies6 subunit of the INO80 remodeler in establishing nucleosome architecture across the S. cerevisiae genome. We show that the loss of Ies6 decreases nucleosome spacing and disrupts regular nucleosome arrays to a similar extent as deletion of the Ino80 ATPase. Our results strongly implicate Ies6 as a critical determinant of INO80-mediated chromatin organization in vivo.
Ies6 forms a structural module with Arp5. Together, these subunits establish multiple contacts with both nucleosomal DNA and histones (Fig. 1A), potentially gripping the nucleosome while the ATPase subunit translocates DNA^23^. Loss of Ies6 likely destabilizes Arp5’s engagement with the nucleosome, reducing the efficiency of nucleosome sliding. In addition, Ies6 deficiency may impair Arp5’s ability to recognize the acidic patch of the H2A–H2B dimer, a critical allosteric activator of INO80 activity^21^.
An unexpected outcome of our genetic analysis is the synthetic lethality observed between ies6Δ and isw2Δ. Whereas INO80 broadly shapes nucleosome architecture across the genome, Isw2 functions as a highly specialized remodeler that influences only a few hundred genes^14,15,36,42^. Both remodelers are known to reposition + 1 nucleosomes upstream into the nucleosome‑depleted region, and we speculate that this shared activity underlies the synthetic lethality when both remodelers are absent. Consistent with this interpretation, growth defects have been reported upon simultaneous depletion of the Isw2 and Ino80 ATPases^14^. In addition, both ISW2 and INO80 play critical roles in nucleosome organization near replication origins and within ribosomal DNA^43,44^. Thus, the synthetic lethality between ies6Δ and isw2Δ may arise, at least in part, from combined defects in these genomic regions.
In addition to the lethal genetic interaction with ISW2, we also observed growth defects when ies6Δ was combined with isw1Δ or chd1Δ. Although these double and triple mutants remained viable, they displayed markedly slower growth than ies6Δ alone, suggesting partial redundancy between INO80 and spacing remodelers of ISWI and Chd1 families in maintaining nucleosome organization. These results are also consistent with the model that multiple remodeler families collaborate to generate and preserve regular nucleosome arrays, yet certain remodelers play non-redundant roles at specific chromatin sites.
Another unexpected outcome of our study is the weak correlation between alterations in nucleosome organization and steady-state gene expression following deletion of either Ies6 or the Ino80 ATPase. This disconnect suggests that disruptions in nucleosome organization within gene bodies do not necessarily translate into measurable changes in steady-state transcription. Instead, more subtle architectural features, such as altered chromatin accessibility near TSS, shifts in the positioning of the + 1 nucleosome or reorganization within the nucleosome-depleted region upstream of it, may underlie the transcriptional alterations observed upon loss of Ino80 function^45^. Furthermore, indirect or secondary consequences of Ino80 ablation, including perturbations in transcription factor recruitment, altered nucleosome dynamics, histone variant exchange, or an activated stress-response, could also contribute to the observed transcriptional outcomes^46,47^. If not directly regulating gene expression, we speculate that the nucleosome-spacing activity of Ino80 serves to suppress cryptic transcription within intergenic regions and gene bodies, just like other spacing remodelers from the ISWI and Chd1 families^15,33,48–50^.
Methods
Yeast strain generation
All yeast strains used in this study (Supp. Table 1) were derived from the W303 background. To delete the IES6 gene via homologous recombination, oligonucleotides oFMP1110 (gaaggttaaaattgtcatcatcatcagcgtgagaaagtcgaaacagatccccgggttaattaagg) and oFMP1111 (gaaaggttgtctacaagctaaaatacatacatacatatacaatgcgaattcgagctcgtttaaactgg) were used to amplify natMX6 marker from plasmid pFA6a-natMX6 (yFMP524). PCR products were purified from agarose gel and transformed into WT yeast strain, generating yFMP627 and yFMP628 lacking IES6. Gene deletion was confirmed by PCR using oligos oFMP1112 (cgatgacgacgactacct) and oFMP1113 (caaagtggagacgatgctg).
Sporulation and tetrad dissection
The haploid strain lacking IES6 (yFMP628) was mated with TKO (YTT227). Resulting diploids (yFMP468, yFMP469) were grown in YPA + 4% glucose media. Sporulation was induced in media lacking adenine, leucine and trpytophan (1% KOAc supplemented with uracil 5 mg/L and histidine 5 mg/L) at 23 °C for 4–5 days. Tetrads were dissected using a Singer MSM 400 dissection microscope onto YPAD plates and incubated at 30 °C for 5 days. Tetrads were replica-plated onto minimal media lacking appropriate amino acids or on YPAD plates with cloNAT selection and grown for an additional 3 days.
MNase-seq
Yeast nuclei preparation, MNase digestion and DNA isolation was performed as described previously^15,51^. Briefly, cells were grown to OD_600_ 0.8 in 500 ml YPAD media. Spheroplasts were prepared by treating cells with 2 mg zymolyase at 30 °C for 20 min, followed by washing with ice-cold 1 M sorbitol. Nuclei were resuspended in MNase digestion buffer (15 mM Tris–Cl pH 7.5, 50 mM NaCl, 1.4 mM CaCl_2_, 0.2 mM EGTA pH 8.0, 0.2 mM EDTA pH 8.0, 5 mM β-mercaptoethanol) and digested with increasing amounts of MNase for 20 min at 37 °C. DNA was purified via Proteinase K treatment, followed by Phenol: Chloroform: Isoamyl alcohol extraction and RNase digestion. Samples with ~ 70% mononucleosome content were selected for library preparation. Sequencing libraries were prepared using the NEBNext Ultra II DNA Library Prep kit for Illumina and sequenced in 50 bp paired-end mode, yielding five to ten million reads per sample.
Data analyses
MNase-Seq data were analyzed as described previously^15^. Briefly, fastq files were mapped using Bowtie2 v2.2.9 to R64-1-1 sacCer3 genome with settings -X 500, -no-discordant, -no-mixed options^52^. Fragments of 140–160 bp were selected and used for further. To generate nucleosome maps, dyad positions were inferred from fragment midpoints and extended by 50 bp to depict a corresponding nucleosome. NRL and array regularity were calculated after subsampling all samples to 5 million reads using scripts (https://github.com/musikutiv/tsTools) which adapts previously published method^37^. To generate the nucleosome dyad density plots, we first calculated a signal matrix aligned to the + 1-nucleosome using the coverageWindowsCenteredStranded function in tsTools v0.1.2. Mean signal values were then calculated for each base pair across all 5,015 genes, and the resulting profiles were normalized to the mean signal within the specified window. MNase-Seq data of cells lacking Ino80 ATPase in BY4741 background^26^ were processed similarly as in this study. Genes with altered expression in ies6Δ and ino80Δ cells (fold change > 1.5) were downloaded from GSE52000 and GSE77257^25,33^. Boxplots represent the median, interquartile range and 1.5X of the interquartile range. Statistical significance comes from a two-tailed Welch’s t-test performed on the median NRL values of two biological replicates. A single asterisk (*) denotes p < 0.05, a double asterisk (**) denotes p < 0.01 and n.s. indicates for p ≥ 0.05. Pymol 3.0 was for structural visualizations. Residues interacting with the nucleosome were identified by selecting atoms from relevant subunits located within 4.0 Å of the nucleosome. The corresponding residues were then mapped and displayed as spheres to highlight potential contact sites.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary Material 1
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Xu, Z., Prajapati, H. K., Eriksson, P. R. & Clark, D. J. The ISW 1 and CHD 1 chromatin remodelers suppress global nucleosome dynamics in living yeast cells. Science Adv Ance S (2025).10.1126/sciadv.adw 7108 PMC 1231596540749069 · doi ↗ · pubmed ↗
- 2Grava, S., Dumoulin, P., Madania, A., Tarassov, I. & Winsor, B. Functional analysis of six genes from chromosomes XIV and XV of Saccharomyces cerevisiae reveals YOR 145c as an essential gene and YNL 059c/ARP 5 as a strain-dependent essential gene encoding nuclear proteins. 9 (2000).10.1002/1097-0061(200008)16:11<1025::AID-YEA 602>3.0.CO;2-110923024 · doi ↗ · pubmed ↗