Candida serves as a reservoir associating with facilitating the survival and pathogenesis of Helicobacter pylori
Jianchao Sun, Tingxiu Yang, Qing Luo, Yan Wang, Wei Zhang, Zhenghong Chen

TL;DR
This study shows that Candida can host Helicobacter pylori, helping it survive and cause disease, which could explain its high recurrence rates.
Contribution
The study reveals a novel intracellular association between H. pylori and Candida, suggesting Candida as a survival reservoir for H. pylori.
Findings
H. pylori antigens and 16S rDNA were detected within Candida isolates from multiple clinical sources.
CacoHp strains showed enhanced inhibitory and adhesive effects on GES-1 cells compared to standard Candida.
Fluorescent staining suggests viable H. pylori may reside intracellularly within Candida cells.
Abstract
Helicobacter pylori (H. pylori) is a key pathogen in gastritis and gastric ulcers, noted for its high recurrence rates. H. pylori internalization into Candida vacuoles may enhance its survival and pathogenicity. In this study, we investigated the intrinsic association between H. pylori and Candida by performing H. pylori-related assays on Candida isolates obtained from clinical fecal samples, gastric mucosal tissues, and vaginal secretions. About 399 isolates of Candida were isolated from fecal samples of patients with digestive diseases. H. pylori antigens in feces and 16S rDNA within Candida isolates were detected, showing a significant correlation. After co-culturing H. pylori and Candida in vitro, H. pylori 16S rDNA-positive Candida (CacoHp) were obtained. In vitro, the CacoHp strain showed enhanced inhibitory and adhesive effects on GES-1 cells than standard Candida strain. H.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
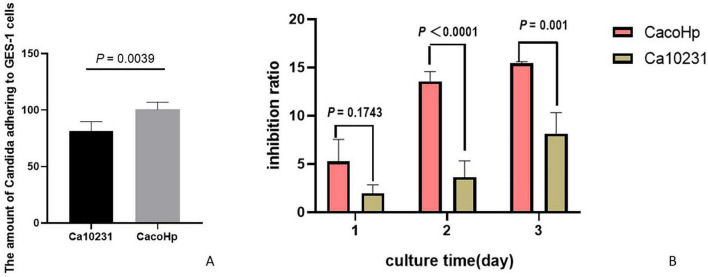 FIGURE 1
FIGURE 1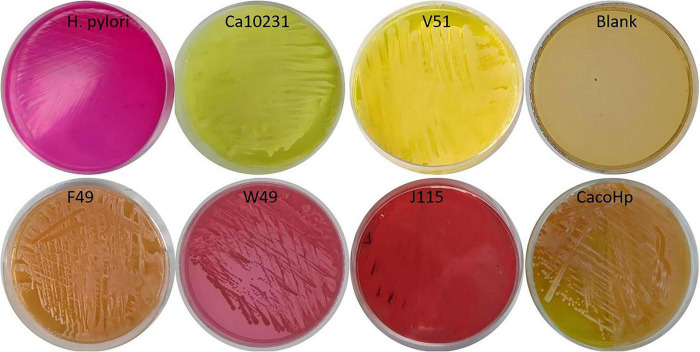 FIGURE 2
FIGURE 2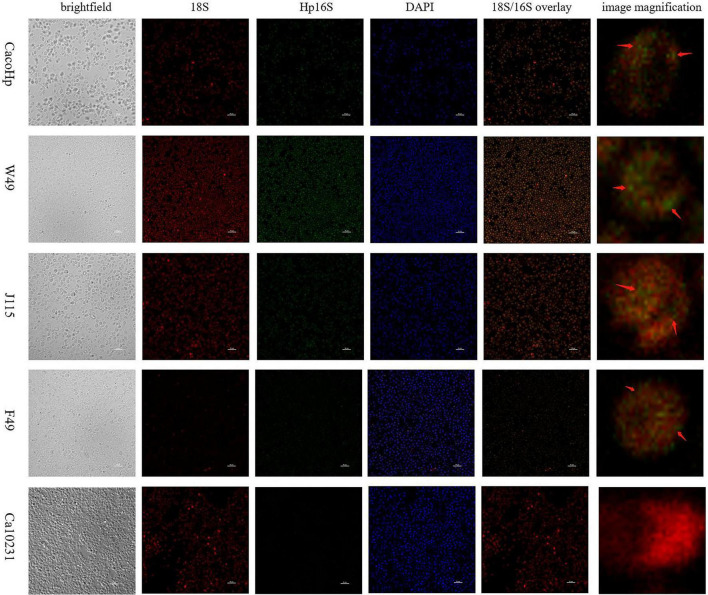 FIGURE 3
FIGURE 3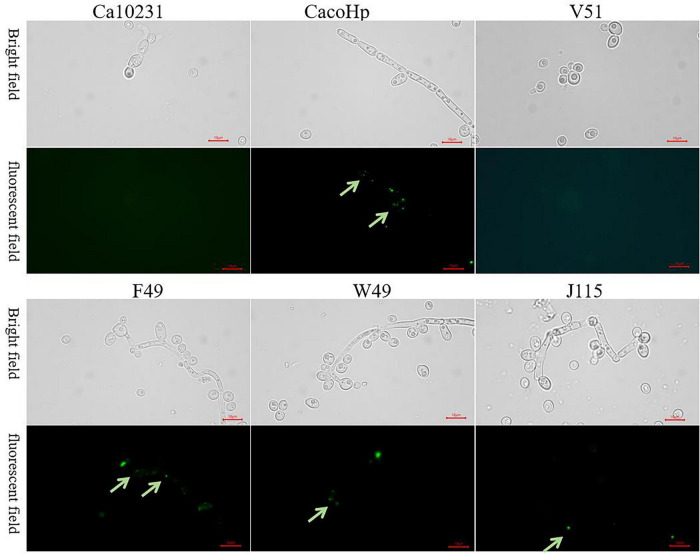 FIGURE 4
FIGURE 4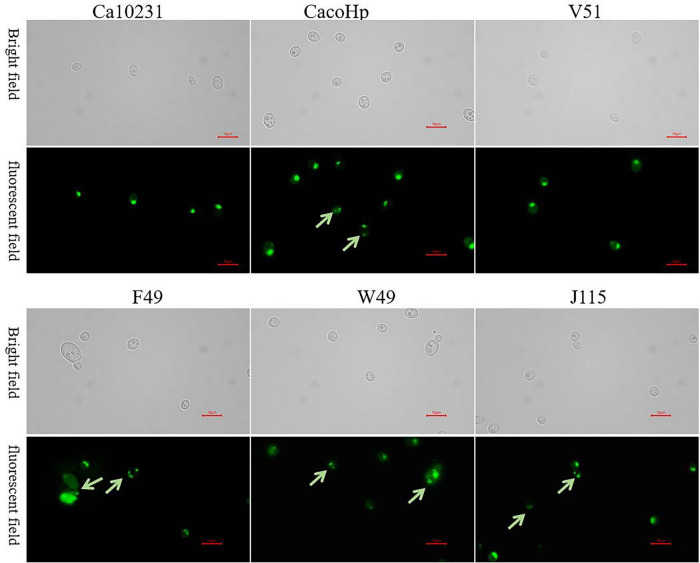 FIGURE 5
FIGURE 5| Gene | PCR type | Stage | Primers | Sequence (5’–3’) | Amplicon (bp) |
|---|---|---|---|---|---|
|
| Nested | First | HeliS | AAGAACCTTACCTAGGCTTGACATTG | 497 |
| HeliN | CCGTGGGCAGTAGCCAATT | ||||
| Second | Hpup | TGAGAGAATCCGCTAGAAATAGTGG | 454 | ||
| Hpdown | TAGCATCCTGACTTAAGGCAAACA |
| Gene | Primer | Sequence (5’–3’) | Amplicon (bp) |
|---|---|---|---|
|
| F | AGCGGTAGCTTTGATTAGTG | 268 |
| R | GCCTTCGTTGATAGTGATGT | ||
| 16S rRNA | F | GGGATAGTCAGTCAGGTGTG | 245 |
| R | ACTAGCATCCATCGTTTAGG |
| Total | |||
|---|---|---|---|
| + | - | ||
| SHpAg | + 49 | 63 | 112 |
| - 44 | 243 | 287 | |
| Total | 93 | 306 | 399 |
| 16S rRNA (Cq) | Result | Result | ||
|---|---|---|---|---|
| Ca10231 | / | Negative | / | Negative |
| V51 | / | Negative | / | Negative |
| W49 | 32.29 ± 2.38 | Positive | 33.83 ± 1.84 | Positive |
| J115 | 30.67 ± 2.62 | Positive | 34.20 ± 1.77 | Positive |
| F49 | 32.03 ± 5.23 | Positive | 32.44 ± 1.91 | Positive |
| CacoHp | 29.41 ± 3.68 | Positive | 34.62 ± 2.04 | Positive |
| Blank | / | Negative | / | Negative |
|
| 17.36 ± 1.33 | Positive | 18.78 ± 2.51 | Positive |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHelicobacter pylori-related gastroenterology studies · Esophageal Cancer Research and Treatment · Gastroesophageal reflux and treatments
Introduction
1
Helicobacter pylori (H. pylori), a Gram-negative pathogenic bacterium that colonizes the human stomach, has a global prevalence of approximately 50% (Hooi et al., 2017). This bacterium is the primary causative agent of gastritis and peptic ulcers. Notably, 1–10% of individuals infected with H. pylori may show progression from chronic active gastritis to severe pathological conditions, including gastric mucosal atrophy, intestinal metaplasia, and development of gastric cancer or mucosa-associated lymphoid tissue (MALT) lymphoma (Yamaoka, 2018).
The eradication of H. pylori is essential for the prevention of gastric cancer (Kim et al., 2014). However, between 2000 and 2010, the average eradication rate of H. pylori ranged from 80 to 90%, whereas drug-resistant isolates exhibited a significantly lower eradication rate, ranging from 40 to 60% (Glupczynski et al., 2001; Savoldi et al., 2018; Cortés et al., 2021). Hu et al. (2017) analyzed H. pylori recurrence rates reported between 1988 and 2017, providing evidence for the association between H. pylori recurrence and specific time periods, geographical regions, and socioeconomic statuses. H. pylori recurrence is influenced by multiple factors, including treatment regimens, timing and duration of therapy, mode of administration, presence of oral H. pylori, formation of the coccoid form, and biofilm development (Sun and Zhang, 2019). Ultimately, recrudescence or reinfection leads to recurrence, representing a major challenge in H. pylori management. However, the recurrence of H. pylori may be closely associated with its internalization.
H. pylori is an obligate extracellular bacterium that primarily colonizes the surface of gastric mucosal epithelial cells, but also has the ability to survive intracellularly—a feature linked to its immune evasion mechanisms and ability to establish persistent infection (Amieva and El-Omar, 2008; Salama et al., 2013). H. pylori possesses invasive capabilities in multiple cell types, including gastric mucosal epithelial cells (Wang et al., 2017), macrophages (Du et al., 2016), and HEp-2 cells (a human laryngeal epidermoid carcinoma cell line) (Evans et al., 1992). Notably, clinical isolates obtained from patients in whom eradication therapy failed exhibited significantly higher intracellular invasion efficiency of gastric mucosal epithelial cells than isolates from patients with successful eradication outcomes (Wang et al., 2017). Emerging evidence suggests that Candida cells could potentially serve as a reservoir for H. pylori transmission while simultaneously providing protection against hostile environmental conditions (Tavakolian et al., 2018; Heydari et al., 2020; Sánchez-Alonzo et al., 2021).
In vivo, Candida interacts with other members of the human microbiota and encounters co-infecting pathogens. Interactions between Candida and symbiotic bacteria, especially H. pylori, have developed into complex relationships (Chen et al., 2021; Eichelberger et al., 2023). These interactions occur via direct cellular contact, secretion of signaling molecules or toxins, or modification of the environment to favor one or all interacting partners (De Sordi and Mühlschlegel, 2009; Erb Downward et al., 2013). In this study, we aim to investigate the intrinsic association between H. pylori and Candida by performing H. pylori-related assays on Candida isolates obtained from clinical fecal samples, gastric mucosal tissues, and vaginal secretions.
Materials and methods
2
Ethics approval and informed consent
2.1
All protocols were approved by the Human Medical Ethics Committee of Guizhou Medical University (ethics review approval number: 2022-40). All procedures, including the protocol for obtaining informed consent, were performed in accordance with the principles of the Declaration of Helsinki. Informed consent was obtained from all participants or their legal guardians prior to study initiation.
Strains
2.2
The H. pylori strain ATCC 700392 (commonly referred to as H. pylori 26695) was used. For Candida, we used C. albicans ATCC 10231 (Ca10231) as well as clinical Candida isolates V51 and J115 isolated from vaginal secretions, W49 isolated from gastric biopsy specimens, and F49 from fecal samples. All strains were maintained at the Guizhou Key Laboratory of Microbiology and Infectious Disease Prevention and Control.
Samples
2.3
Fecal samples were collected from 399 patients with gastrointestinal disorders at the Department of Gastroenterology of the Affiliated Cancer Hospital, Guizhou Medical University. To diagnose H. pylori infection, the colloidal gold method was employed for the detection of H. pylori antigen in stool (S_HpAg_) (Gisbert et al., 2006). The presence of H. pylori antigen in the feces was detected using a Wondfo H. pylori Antigen Detection Kit (Guangdong, Wondfo). This kit employs the colloidal gold method to detect H. pylori antigen in feces, and is a common method for diagnosing H. pylori infection. Fecal samples were used for the isolation and culture of Candida. A small quantity of fecal sample was taken and added to sterile physiological saline to prepare a suspension. Subsequently, 0.1 mL of the sample should be added to Sabouraud dextrose agar (SDA, Basebio, Hangzhou, China) containing 50 μg/mL chloramphenicol for the isolation and cultivation of Candida. Candida was inoculated onto CHROMagar Candida medium (CHROMagar, Paris, France) at 37°C for 24–48 for 24 France at 37 e) at 37 Candida species were identified based on colony color characteristics: green-blue (C. albicans), metallic blue with a pink halo (C. tropicalis), mauve (C. glabrata), pink and fuzzy (C. krusei), and white (other Candida species). The isolated Candida was stored in 20% glycerol solution at -80°C.
Strain culture
2.4
The H. pylori 26695 strain was cultured in brain heart infusion (BHI) medium (OXOID, Basingstoke, United Kingdom) supplemented with 7% defibrinated sheep blood (Biological Industries, Henan, China) and incubated under microaerobic conditions at 37°C for 48–72 h. Ca10231 and Candida isolates were cultured on SDA supplemented with 50 μg/mL chloramphenicol (Solarbio, Beijing, China). Candida cultures were incubated aerobically at 37°C for 24 h.
Co-culture of H. pylori and Candida
2.5
The H. pylori 26695 strain and Ca10231 strains were cultured separately under specific microaerobic and aerobic conditions, respectively, at 37°C. Prepare suspensions of the H. pylori 26695 strain and Ca10231 strain separately using sterile physiological saline at concentrations corresponding to 5.0 McFarland turbidity and 0.5 McFarland turbidity, respectively. Take 1 mL of H. pylori 26695 bacterial suspension and 0.1 mL of Ca10231 suspension, respectively, and add them to 10 mL of BHI liquid medium containing 10% fetal bovine serum. Cultivate under microaerobic conditions at 37 medium 120 r/min for 24 h. Take 100 μL of the post-culture suspension and inoculate it onto SDA medium supplemented with 50 μg/mL chloramphenicol for subsequent cultivation.
Detection of H. pylori 16S rDNA in the total DNA extracted from Candida cells
2.6
Candida cells in each experimental group were sub-cultured up to the fifth generation in SDA supplemented with 50 μg/mL chloramphenicol. Subsequently, a specific H. pylori gene within each generation of Candida was examined following a previously established protocol (Šeligová et al., 2020). DNA used as controls in the polymerase chain reaction (PCR) amplification process was extracted from pure cultures of H. pylori 26695 (positive control) and Ca10231 (negative control). Briefly, Candida cells were suspended in distilled water, and the DAAN nucleic acid extraction or purification reagent (magnetic bead method) was used for DNA extraction from the samples. PCR assays were performed using primers targeting H. pylori 16S rDNA to identify H. pylori among Candida. The primer sequences used for PCR are listed in Table 1.
16S rDNA amplification was performed in a thermal cycler for the first reaction with 94°C for 3 min, 37 cycles of the external amplification reaction (94°C, 45 s; 55°C, 1 min; 72°C, 1 min), 72°C for 5 min, and a hold at 14°C. Use the first-round amplification product as the template for the second round of amplification. The temperature cycle for the second reaction was the same; however, the number of cycles was reduced to 25.
To avoid contamination, negative and blank controls were introduced after every sample in the experiments.
Examination of the ability of Candida to adhere to GES-1 cells
2.7
The CacoHp strains were continuously passaged up to two times. 1 × 10^6^ GES-1 cells were grafted into a 6-well plate and cultured in serum-free RPMI 1640 medium for 12 h. After the cells adhered, 1 × 10^5^ CacoHp and Ca10231 cells were added. After 24 h of culture, the Candida cells adhering to GES-1 cells were counted.
Examination of the toxicity of the Candida culture filtrate in GES-1 cells
2.8
The CacoHp strains were continuously passaged up to five times. A total of 1 × 10^6^ CacoHp and Ca10231 strains were inoculated into the YPD liquid medium and incubated for 24 h under microaerobic conditions. Following incubation, the cultures of CacoHp and Ca10231 were centrifuged and filtered through a 0.22-μm bacterial filter to obtain sterile culture filtrates. These filtrates were further diluted fivefold with RPMI 1640 medium and co-cultured with GES-1 cells for 24, 48, and 72 h. The cytotoxic effects of the filtrates on GES-1 cells were assessed using a Cell Counting Kit-8 (CCK-8). The cell inhibition rate was calculated as follows: Inhibition Rate (%) = [(Absorbance of Control Group–Absorbance of Experimental Group)/Absorbance of Control Group] × 100%.
Detection of urease activity of Candida
2.9
In this study, urea-SDA medium was used to detect the urease activity of Candida. Phenol red was added to the SDA medium as an indicator, and urea was added as a substrate for the urease reaction. Under slightly alkaline conditions, phenol red turns red, causing the medium to change color from yellow to red. Urea and phenol red were added to the SDA medium to prepare a urea SDA medium containing 5% urea and 0.01% phenol red. Candida were inoculated in freshly prepared urea SDA medium and incubated under aerobic conditions at 37 °C for 1–3 days. H. pylori 26695 was inoculated onto a urea Columbia serum agar plate (CSA) (5% urea, 10% serum) under microaerobic conditions as a positive control.
Detection of H. pylori 16S rRNA by fluorescence in situ hybridization
2.10
Candida strains cultured for 24 h in SDA supplemented with chloramphenicol were removed and washed twice with Phosphate-buffered saline (PBS) (pH 7.4) to ensure cleanliness. Subsequently, the fixed fluid diethyl pyrocarbonate (DEPC) -treated water was immediately added, followed by incubation for a minimum of 12 h. Candida samples were dehydrated using a gradient alcohol-paraffin embedding method. A DEPC dilution wash was performed afterwards. Depending on the tissue fixation time, the slices were boiled in a retrieval solution for approximately 10–15 min and allowed to cool naturally. Objective Candida cells were marked using a liquid blocker pen based on their characteristics. Proteinase K working solution (20 μg/mL) was applied to cover the objectives and incubated at 37 °C for 15 min. Washing was performed using pure water, followed by three washes with PBS (pH 7.4), each lasting 5 min. Finally, every section was incubated with a pre-hybridization solution at 37 °C for 1 h.
The pre-hybridization solution was removed and replaced with a hybridization solution containing 1 μM of the fungal 18S rRNA probe (5’-CY3-GTGACAAGCATATGACTAC-CY3-3’). The sections were incubated in a humidity chamber at 40°C for overnight hybridization. The hybridization solution was then removed, and the sections were washed in 2 × saline sodium citrate (SSC) for 10 min at 37°C, followed by two washes in 1 × SSC for 5 min each at 37°C, and a final wash in 0.5 × SSC for 10 min at room temperature. Subsequently, the pre-hybridization solution was removed again, and a hybridization solution containing 1 μM of H. pylori 16S-1 (5’-FAM-GGAGTATCTGGTATTAATCATCG-FAM-3’) probes was added. The sections were incubated in a humidity chamber and hybridized overnight at 40°C. The hybridization solution was then removed, and the sections were washed in 2 × SSC for 10 min at 37°C, followed by two washes in 1 × SSC for 5 min each at the same temperature, and a final wash in 0.5 × SSC for 10 min at 25°C.
For nuclear counterstaining, the sections were incubated with Nuclei were counterstained with 4’, 6-diamidino-2-phenylindole (DAPI) in the dark for 8 min before mounting. Microscopic examination and imaging were conducted using a fluorescence microscope. DAPI emitted blue fluorescence under ultraviolet (UV) excitation between 330 and 380 nm, with an emission wavelength of 420 nm; FAM emitted green fluorescence under excitation between 465 and 495 nm and an emission wavelength of 515–555 nm; and CY3 emitted red fluorescence upon excitation between 510 and 560 nm, with an emission wavelength of approximately 590 nm.
Detection of H. pylori gene expression within Candida by quantitative polymerase chain reaction
2.11
Candida strains isolated from feces, vaginal secretions, and gastric mucosa were cultured on SDA medium for five generations before RNA extraction using the phenol-chloroform method. ABScript III RT Master Mix with gDNA Remover was used to generate complementary DNA (cDNA) from 1 μL of total RNA for qPCR analysis with ChamQ qPCR SYBR Green Master Mix (Vazyme Biotech Co., Ltd. Nanjing, China) on a CFX96 Real-time system (Bio-Rad, Hercules, CA, United States). The primer sequences used for qPCR are listed in Table 2 (Raghwan and Chowdhury, 2014).
qPCR Testing process was performed in CFX96 Real-time system with 95°C for 30 s, 40 cycles of the external amplification reaction [95°C, 10 s; 60°C, 30 s (Collect fluorescence signals)]. The test results are described qualitatively.
Detection of fluorescent antibodies against H. pylori within Candida
2.12
Candida was cultured in the BHI medium (OXOID) at 37°C with shaking at 120 r/min for 24 h. The Candida bacterial fluid was centrifuged and washed thrice with saline to obtain a Candida bacterial suspension with a McFarland standard concentration of 2. Fluorescein isothiocyanate (FITC)-labeled anti-H. pylori IgG antibodies (Cat. No. PA1-73161; Thermo Fisher Scientific) were reconstituted at a dilution of 1:100 to ensure thorough mixing, and the mixture was incubated in a light-protected environment at 25°C for 1 h. A total of 10 μL of the suspension was applied to a glass microscope slide, covered with a coverslip, and examined under a fluorescence microscope at 1,000 × magnification. Ca10231 was used as a negative control.
Detection of bacterial activity within Candida
2.13
Candida isolates cultured on SDA supplemented with chloramphenicol for 24 h were used to prepare Candida suspensions with a McFarland standard concentration of 2. A total of 2 μL of LIVE/DEAD^®^ BacLight*™* Bacterial Viability Kits (Thermo Fisher Scientific) was added to 1 mL of Candida bacterial suspension and incubated for 15 min at 25°C away from light. A total of 10 μL of the suspension was applied to a glass microscope slide, covered with a coverslip, and examined under a fluorescence microscope at 1,000 × magnification. Ca10231 was used as a negative control.
Data statistics
2.14
Data analysis was conducted using GraphPad Prism 9.5.0 software. Measured data were presented as the mean ± standard deviation, and one-way analysis of variance (ANOVA) was employed for comparisons between two groups. Correlation analysis was conducted using the Point-Biserial correlation coefficient. When P < 0.05, the difference was deemed statistically significant.
Results
3
Examination of H. pylori 16S rDNA in Candida strains isolated from feces
3.1
Detection of H. pylori antigen in stool samples
3.1.1
Among the 399 stool samples from which Candida was isolated, 112 samples tested positive for S_HpAg_, confirming H. pylori infection in these patients.
Candida isolation and identification
3.1.2
To explore the relationship between Candida and H. pylori, we isolated Candida species from fecal samples of patients with gastrointestinal disorders and performed species identification. In total, 399 Candida isolates were isolated from 997 fecal samples. The distribution of these Candida species was as follows: 319 strains were identified as Candida albicans, 44 as Candida krusei, 14 as Candida glabrata, 7 as Candida tropicalis, and 15 as other Candida species.
Detection of H. pylori 16S rDNA in Candida
3.1.3
Among 399 isolates of Candida isolated from the fecal samples, H. pylori 16S rDNA was detected in 93 isolates. To investigate the correlation between H. pylori infection in patients and H. pylori 16S rDNA-positive Candida, the Point-Biserial correlation coefficient was used to analyze the correlation between fecal H. pylori antigen and Candida isolates with H. pylori 16S rDNA. The findings demonstrated a significant correlation between fecal H. pylori antigen and H. pylori 16S rDNA-positive Candida (P < 0.001), indicating that the presence of H. pylori 16S rDNA in Candida was associated with H. pylori infection (Table 3).
Acquisition and characteristics of H. pylori 16S rDNA-positive Candida strains
3.2
Acquisition of H. pylori 16S rDNA-positive Candida strains
3.2.1
CacoHp strains were sub-cultured consecutively for five passages. Following Candida genomic DNA extraction, PCR testing for H. pylori was performed. The results of the PCR test demonstrated positivity for H. pylori 16S rDNA. Consistent findings were also observed in the isolates obtained from partial gastric mucosa, fecal samples, and vaginal secretions.
CacoHp strain exhibited stronger adhesion capability toward GES-1 cells
3.2.2
To investigate the effect of co-culturing H. pylori with Candida on the adhesion capacity of Candida to GES-1 cells, we selected second-generation CacoHp cells. The number of CacoHp strains that adhered to GES-1 cells was significantly higher than that of the Ca10231 strain (Figure 1A). These results indicate that CacoHp exhibits enhanced adhesive properties toward GES-1 cells, which may be associated with the colonization of H. pylori 16S rDNA-positive Candida strains in the gastrointestinal environment.
Ca10231: Candida standard strain ATCC10231. CacoHp: Laboratory-prepared Candida (ATCC 10231) internalized by H. pylori (ATCC 700392). (A) Adhesion effect of Candida on GES-1 cells. (B) Inhibitory effect of Candida on proliferation toward GES-1 cells.
CacoHp strain exhibited a stronger inhibitory effect on proliferation toward GES-1 cells
3.2.3
To investigate the inhibitory effect of co-culturing H. pylori with Candida on proliferation toward GES-1 cells, fifth-generation CacoHp was selected. The results demonstrated that CacoHp exhibited a significantly greater inhibitory effect on GES-1 cells compared to Ca10231 (Figure 1B). This inhibitory effect on proliferation increased in a time-dependent manner and became progressively more pronounced. These findings suggest that H. pylori 16S rDNA-positive Candida strains exert continuous toxic effects on GES-1 cells.
The adhesion and colonization of bacteria on the cell surface represent critical steps in bacterial pathogenicity (Huang et al., 2016). This study demonstrated that CacoHp cells exhibit significantly stronger adhesion to GES-1 cells. These findings may explain why Candida can be detected in the stomachs of patients with gastric ulcers and gastritis. Furthermore, this study provides insights into the mechanism by which H. pylori 16S rDNA-positive Candida colonizes the stomach.
Detection of H. pylori-related characteristics in H. pylori 16S rDNA-positive Candida isolated from clinical specimens
3.3
Candida exhibited urease activity
3.3.1
After 3 days of incubation in urea-SDA medium, the W49, F49, and J115 Candida strains induced a color change in the medium from yellow to red. This phenomenon may be attributed to the metabolic production of urease by internalized H. pylori, resulting in an elevated pH and subsequent color change (Figure 2).
Ca10231: Candida standard strain. V51: H. pylori 16S rDNA-negative Candida isolated from vaginal secretions. F49, W49, J115: Candida strains isolated from feces, gastric mucosal tissue, and vaginal secretions, respectively. CacoHp: Laboratory prepared Candida internalized by H. pylori. The F49, W49, J115, and CacoHp resulted in a noticeable transition of the medium color from yellow to red, similar to H. pylori. Conversely, no discernible color change was observed in Ca10231, V51 or in the blank control.
As Candida does not contain the ureA gene and cannot express urease, the ability of H. pylori 16S rDNA-positive Candida to induce a color change in urea-SDA medium suggests that urease production originates from H. pylori. As H. pylori can metabolize urea to produce large amounts of urease, Candida strains harboring H. pylori 16S rDNA may secrete urease derived from H. pylori, thereby generating ammonia and increasing the pH of the medium. This pH shift causes phenol red to transition from yellow to red under alkaline conditions. The production of urease by H. pylori 16S rDNA-positive Candida during its metabolic processes further suggests that these strains possess stronger pathogenic potential than other Candida strains.
Detection of H. pylori 16S rRNA in Candida strains
3.3.2
To further investigate whether H. pylori 16S rRNA is present in Candida, we designed an H. pylori-specific 16S rRNA probe and observed a similar signal in CacoHp, F49, W49, and J115 strains, all of which are H. pylori 16S rDNA-positive Candida isolates and green fluorescent spots can be observed within the cells of Candida (Figure 3).
FISH staining with H. pylori-specific probes in Candida strains. Fungal ribosomes were stained with a universal 18S rRNA probe (red), and H. pylori was detected using a specific 16S rRNA probe set (green). Co-localization of 18S and 16S fluorescence (red arrowheads) indicates the presence of H. pylori 16S rRNA in Candida cells. Scale bars = 10 μm.
Detection of H. pylori 16S rRNA and ureA gene expression in Candida strains
3.3.3
Using FISH, we identified H. pylori 16S rRNA in Candida. Results obtained from the urea SDA medium further suggested that the urease gene of H. pylori may be expressed in Candida. Therefore, we employed qPCR to detect both H. pylori 16S rRNA and the ureA gene in Candida. These strains may express H. pylori 16S rRNA and the ureA gene. The positive results indicate that Candida, once internalized by H. pylori, can potentially maintain stable expression of H. pylori genes. The results are summarized in Table 4.
Detection of antigens against H. pylori within Candida strains
3.4
The FISH and PCR results indicated the presence of H. pylori nucleic acids in H. pylori 16S rDNA-positive Candida. To further confirm the presence of H. pylori in Candida, we employed a direct immunofluorescence assay to detect H. pylori antigens within Candida cells. Under a fluorescence microscope, no green fluorescence was observed in the Ca10231 strain. However, H. pylori 16S rDNA-positive Candida exhibited distinct green fluorescence, indicating specific binding of FITC-labeled anti-H. pylori IgG antibodies with H. pylori antigens inside Candida cells (Figure 4). Therefore, the presence of H. pylori antigens was confirmed in H. pylori 16S rDNA-positive Candida cells.
Ca10231: Candida standard strain. V51: H. pylori 16S rDNA-negative Candida isolated from vaginal secretions. F49, W49, J115: Candida strains isolated from feces, gastric mucosal tissue, and vaginal secretions, respectively. CacoHp: Laboratory prepared Candida internalized by H. pylori. No green fluorescence was detected in the Candida standard strain, and no H. pylori antigen was present. F49, W49, J115, and CacoHp exhibited distinct green fluorescence (green arrow), indicating specific binding of FITC-labeled anti-H. pylori IgG antibodies to H. pylori antigen inside Candida cells. Magnification = × 1,000.
Viable bacteria may exist within Candida strains
3.5
The LIVE/DEAD^®^ BacLight Bacterial Viability Kit was used to assess bacterial cell membrane integrity. Bacteria with intact membranes fluoresced green, indicating metabolic activity, while those with compromised membranes, suggestive of cell death, fluoresced red. Green fluorescent signals were observed within H. pylori 16S rDNA-positive Candida, indicating the presence of H. pylori with intact membranes and the potential viability of H. pylori within Candida (Figure 5).
Ca10231: Candida standard strain. V51: H. pylori 16S rDNA negative Candida isolated from vaginal secretions. F49, W49, J115: Candida strains from feces, gastric mucosal tissue, and vaginal secretions, respectively. CacoHp: Laboratory-prepared Candida internalized by H. pylori. No green fluorescence was observed in the Candida standard strain, F49, W49, J115, and CacoHp exhibited faint green fluorescence (green arrow). The results suggest that H. pylori 16S rDNA-positive Candida may contain viable H. pylori bacteria.
Discussion
4
H. pylori is a gram-negative pathogen that colonizes the human stomach in specific gaseous environments and nutritional requirements. H. pylori exhibit abundant urease production, along with oxidase and catalase activities, as well as metabolism and generation of multiple virulence factors crucial for colonization and pathogenicity (Clyne and Ó Cróinín, 2025). H. pylori have been classified as a class I carcinogen due to its clear pathogenic effect.
Infection with H. pylori is strongly associated with an increased risk of gastric cancer. H. pylori infection can cause chronic gastritis and peptic ulcers. Chronic inflammation induces sustained damage to the gastric mucosal cells and establishes a microenvironment that facilitates gene mutations, potentially promoting tumorigenesis. The H. pylori Kyoto Global Consensus (2015) (Sugano et al., 2015) and the H. pylori Maastricht V/Florence Consensus (2016) (Malfertheiner et al., 2017) both assert that H. pylori infection is closely associated with the development of gastric cancer, and eradication of H. pylori is an effective preventive measure against gastric cancer. However, the therapeutic outcomes of H. pylori treatment remain unsatisfactory. The primary factor contributing to the ineffectiveness of H. pylori eradication is the growing resistance to commonly prescribed antibiotics (McNicholl et al., 2012; Gong and Yuan, 2018; Chey et al., 2024). Additionally, the invasion of H. pylori into the vacuoles of Candida strains may be associated with recurrence (Yang et al., 2024).
It is now well-established that diverse symbiotic fungal communities exist both internally and externally on mammalian surfaces (Alexiev et al., 2023; Limon et al., 2017). The emerging concept of a fungal bacteriome refers to bacteria residing within and closely associated with fungal host cells (Robinson et al., 2021). Evidence suggests co-colonization of the upper gastrointestinal tract by both H. pylori and Candida, suggesting a significant association between Candida and H. pylori in the ulcerative lesions of gastric ulcers, non-ulcer dyspepsia, and giant gastric ulcers (Ramaswamy et al., 2007). Candida isolates obtained from clinical fecal samples showed H. pylori 16S rDNA with a positivity rate of 28.1%. A correlation was observed between the detection of H. pylori 16S rDNA Candida and fecal H. pylori antigen test results in patients. Clinical sample test results suggest a potential interaction between Candida and H. pylori, which may exacerbate disease severity or contribute to chronic infections.
To verify the adhesion and toxicity of H. pylori 16S rDNA-positive Candida toward GES-1 cells, we co-cultured Candida with H. pylori in vitro to obtain H. pylori 16S rDNA-positive Candida. In vitro cell experiments demonstrated that the CacoHp strain filtrate exhibited significantly stronger adherence and inhibitory effects in GES-1 cells. The enhanced toxicity of CacoHp may be associated with the presence of H. pylori in Candida cells. This further explains why Candida can be detected in patients with gastritis or gastric ulcers.
While Candida usually does not colonize the stomach, we found that H. pylori 16S rDNA positive Candida exhibited a significantly stronger adhesion capability toward GES-1 in vitro. Furthermore, the detection of H. pylori 16S rDNA in Candida isolates suggests the internalization of H. pylori into Candida. H. pylori can enter Candida vesicles, where it produces toxic metabolites (Heydari et al., 2020). In this study, the isolated Candida exhibited urease activity, which is not a common biological characteristic of Candida but represents a prominent feature and pathogenic mechanism of H. pylori. The presence of H. pylori 16S rDNA-positive Candida in the stomach may be associated with positive urea activity test results.
Although recent research has started elucidating the mechanisms underlying bacterial-fungal interactions in opportunistic infections (MacAlpine and Lionakis, 2024; d’Enfert, 2009), our understanding of how these interactions contribute to pathogenesis remains largely incomplete. Notably, we observed the internalization of H. pylori into Candida cells present in the gastric mucosa, as well as intestinal and vaginal isolates of Candida. This suggests a potential mechanism for the transmission of pathogenic H. pylori via internalization into Candida cells (Yang et al., 2023). Therefore, the interaction between H. pylori and Candida may represent a novel pathogenic mechanism associated with H. pylori infection.
Taking together, the results of the direct immunofluorescence, fluorescence in situ hybridization, and bacterial activity detection, it is possible that H. pylori may exist as a complete bacterial entity within Candida cells, maintaining a relatively low level of metabolic activity. H. pylori may reside in Candida cells in the form of coccoid bodies. Consequently, during in vitro isolation and culture processes, H. pylori within Candida was not isolated. However, under the influence of environmental factors, nutritional conditions, and in vivo cytokines, live H. pylori are released from Candida cells.
Recent studies have demonstrated the role of fungi in maintaining bacterial microbiota balance and influencing overall intestinal health (Krüger et al., 2019). However, the mechanism by which Candida affects bacterial colonization remains unclear. In our study, the internalization of H. pylori by Candida conferred the ability to break down urea, a property not observed in standard Candida strains. qPCR successfully identified gene expression of H. pylori. These findings provide compelling evidence for the interplay between H. pylori and Candida, suggesting that internalized H. pylori can maintain gene expression within Candida.
Furthermore, the internalized state of H. pylori within Candida may enable its persistence as an active entity while benefiting from the conducive environment provided by Candida under specific conditions. Consequently, this internalized form of Candida harboring H. pylori has the potential to establish chronic infections or trigger recurrence when external conditions change. Understanding the mechanisms underlying the interplay between H. pylori and Candida opens new avenues for research on chronic infections and potential treatment strategies. By targeting this internalized form of Candida harboring H. pylori, it may be possible to develop more effective therapies to disrupt symbiotic relationships and prevent recurrent infections.
These findings provide insights into a potential novel mechanism of pathogenesis involving Candida or H. pylori alone or in combination. However, we were unable to isolate cultivable H. pylori from Candida. Further investigations are necessary to elucidate the precise processes by which internalization occurs. Our research has not yet conclusively demonstrated that H. pylori can persist for an extended period of time within Candida cells while maintaining its structural integrity and metabolic activity. Whether H. pylori can remain as an independent, structurally complete, and viable bacterium within Candida cells remains to be further investigated. Moreover, it would be valuable to explore whether viable forms of H. pylori could be released from internalized Candida cells into the surrounding environment. Understanding this aspect may have significant implications for disease transmission and treatment strategies. Future research should adopt a multidisciplinary approach, integrating molecular biology techniques with genomics, chemical, and microbial ecology methodologies, biophysical investigations, and ecological modeling to comprehensively examine the interactions between Candida and H. pylori.
Conclusion
5
Overall, our study provides important insights into the complex relationship between Candida and H. pylori. Candida may provide a protective microenvironment against bactericidal agents, which could, which could, in part, explain cases of treatment failure and recurrence. Moreover, after internalization by Candida, H. pylori may retain its metabolic activity and with the potential continue to produce toxic metabolites, leading to chronic damage to the gastric mucosa. These insights may help guide the exploration of new preventive or therapeutic approaches.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alexiev A. Melie T. Martindale R. Delacey C. Quandt C. A. Mc Kenzie V. J. (2023). Mr. Toad’s wild fungi: Fungal isolate diversity on colorado boreal toads and their capacity for pathogen inhibition. Fungal Ecol. 66:101297. 10.1016/j.funeco.2023.101297 38487623 PMC 10938945 · doi ↗ · pubmed ↗
- 2Amieva M. R. El-Omar E. M. (2008). Host-bacterial interactions in Helicobacter pylori infection. Gastroenterology 134 306–323. 10.1053/j.gastro.2007.11.009 18166359 · doi ↗ · pubmed ↗
- 3Chen X. Zhou X. Liao B. Zhou Y. Cheng L. Ren B. (2021). The cross-kingdom interaction between Helicobacter pylori and Candida albicans. P Lo S Pathogens 17:e 1009515. 10.1371/journal.ppat.1009515 33956895 PMC 8101746 · doi ↗ · pubmed ↗
- 4Chey W. D. Howden C. W. Moss S. F. Morgan D. R. Greer K. B. Grover S. (2024). ACG clinical guideline: Treatment of Helicobacter pylori infection. Am. J. Gastroenterol. 119 1730–1753. 10.14309/ajg.0000000000002968 39626064 · doi ↗ · pubmed ↗
- 5Clyne M. Ó Cróinín T. (2025). Pathogenicity and virulence of Helicobacter pylori: A paradigm of chronic infection. Virulence 16:2438735. 10.1080/21505594.2024.2438735 39725863 PMC 12915409 · doi ↗ · pubmed ↗
- 6Cortés P. Nelson A. D. Bi Y. Stancampiano F. F. Murray L. P. Pujalte G. G. A. (2021). Treatment approach of refractory Helicobacter pylori infection: A comprehensive review. J. Prim. Care Commun. Health 12:21501327211014087. 10.1177/21501327211014087 33949229 PMC 8114244 · doi ↗ · pubmed ↗
- 7d’Enfert C. (2009). Hidden killers: Persistence of opportunistic fungal pathogens in the human host. Curr. Opin. Microbiol. 12 358–364. 10.1016/j.mib.2009.05.008 19541532 · doi ↗ · pubmed ↗
- 8De Sordi L. Mühlschlegel F. A. (2009). Quorum sensing and fungal-bacterial interactions in Candida albicans: A communicative network regulating microbial coexistence and virulence. FEMS Yeast Res. 9 990–999. 10.1111/j.1567-1364.2009.00573.x 19845041 · doi ↗ · pubmed ↗