The Tomato Transcription Factor RAV Affects the Systemic Infection of TYLCV by Interacting With V2
Chenwei Zhang, Xin Jia, Guiyan Fan, Xiaoli Ren, Xing Han, Xiaocong Jiao, Yuan Cheng, Yajuan Cheng, Zihan Liu, Zhehao Yuan, Yongmin Chen

TL;DR
A tomato protein called SlRAV2 interacts with a virus protein to control the spread of tomato yellow leaf curl virus.
Contribution
The study reveals a novel antiviral mechanism involving the interaction between SlRAV2 and the viral V2 protein.
Findings
SlRAV2 suppresses systemic TYLCV accumulation by interacting with the V2 protein.
V2 enhances SlRAV2's binding to siRNA, increasing its RNA silencing suppressor activity.
The interaction leads to localized cell necrosis, preventing viral spread to systemic leaves.
Abstract
Tomato yellow leaf curl virus (TYLCV) is a widely distributed pathogen responsible for tomato yellow leaf curl disease. However, the molecular mechanism underlying TYLCV infection and the plant antivirus pathway remains unclear. This study reports that tomato ( Solanum lycopersicum ) RELATED TO ABSCISIC ACID INSENSITIVE3/VIVIPAROUS2 (SlRAV2), a transcription factor of the APETALA2/ETHYLENE RESPONSE FACTOR (AP2/ERF) superfamily containing an AP2/B3 DNA‐binding domain, suppresses systemic TYLCV accumulation by interacting with the V2 protein, an RNA silencing suppressor (RSS) encoded by TYLCV. Transient overexpression and silencing of SlRAV2 altered TYLCV levels in systemic leaves, prompting us to investigate the underlying mechanism. Molecular analyses revealed that SlRAV2 directly binds to the promoter of the pathogenesis‐related protein 1 (SlPR1) gene, thereby activating its expression…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
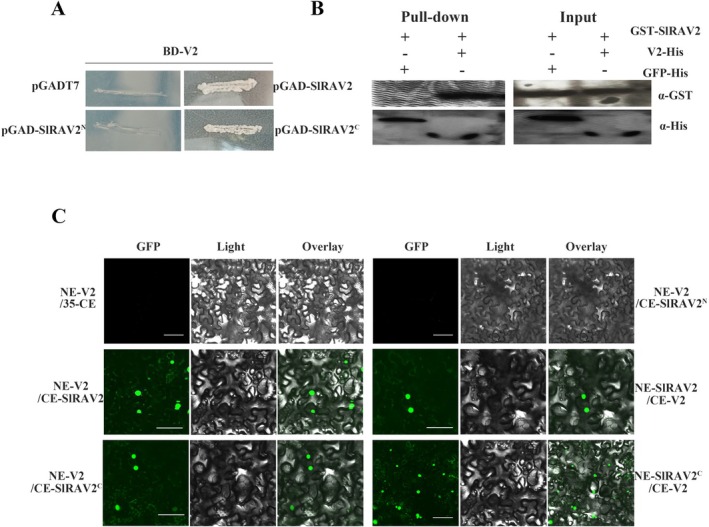 FIGURE 1
FIGURE 1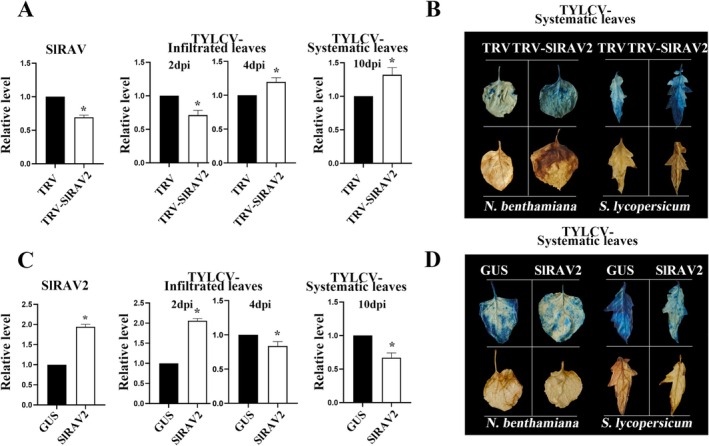 FIGURE 2
FIGURE 2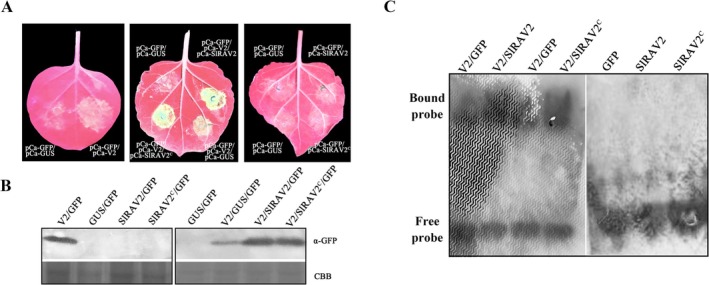 FIGURE 3
FIGURE 3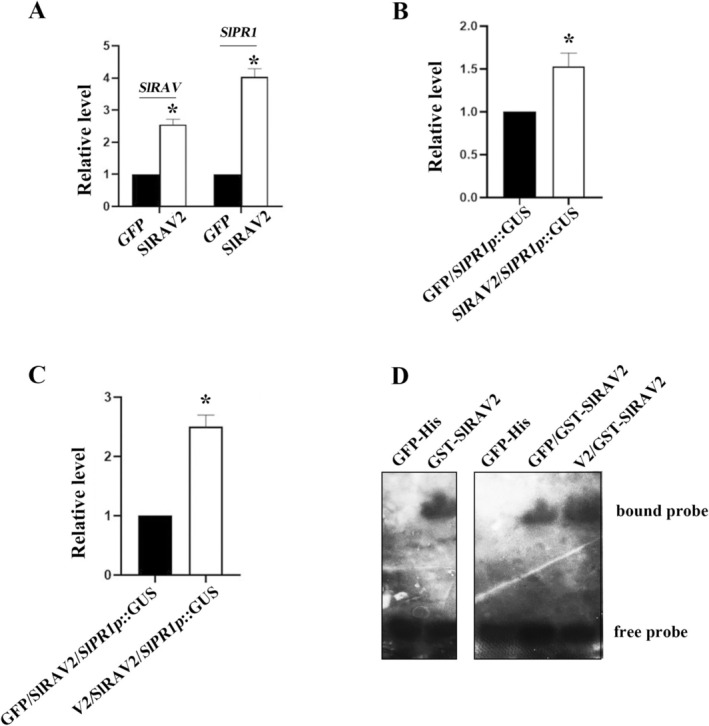 FIGURE 4
FIGURE 4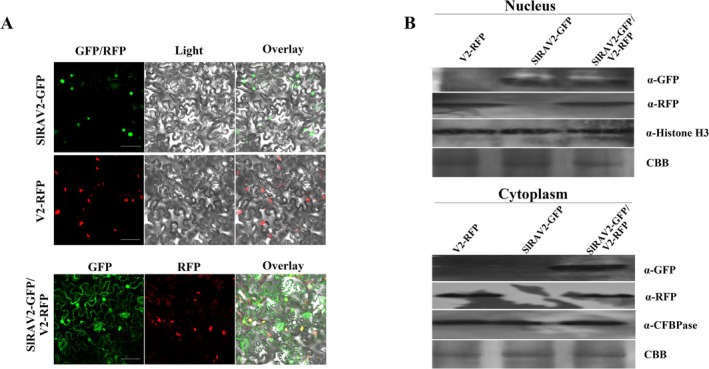 FIGURE 5
FIGURE 5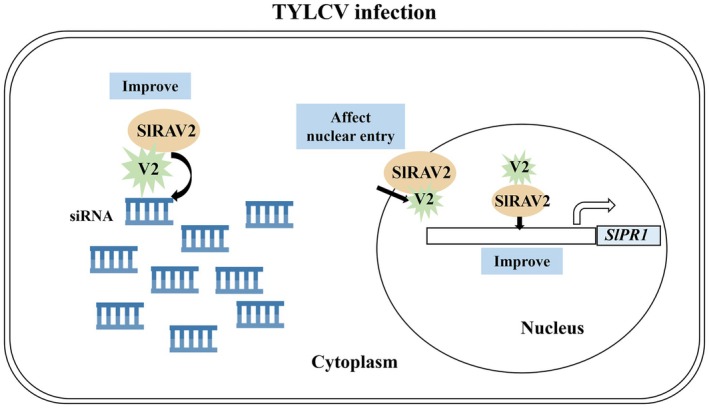 FIGURE 6
FIGURE 6- —National Natural Science Foundation of China10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Virus Research Studies · Plant and Fungal Interactions Research · Plant-Microbe Interactions and Immunity
Introduction
1
Tomato cultivation is plagued by a spectrum of biological stressors, including viral infections, which profoundly impede the growth and severely undermine tomato yield and quality (Wu et al. 2006; Li et al. 2022; Cao et al. 2024). Among these, tomato yellow leaf curl virus (TYLCV)—a member of the Geminiviridae family and Begomovirus genus—is one of the most economically destructive pathogens affecting tomato crops worldwide (Moriones and Navas‐Castillo 2000). TYLCV has a broad host range and exhibits high transmission efficiency, primarily via the whitefly vector Bemisia tabaci (Hemiptera: Aleyrodidae) through a persistent circulative mode of transmission (Goldman and Czosnek 2002). Environmental stressors such as drought and extreme temperatures exacerbate disease severity, further compounding yield losses (Yue et al. 2024). TYLCV possesses a circular single‐stranded DNA genome whose open reading frames (ORFs) encode proteins that contribute to virus replication, movement, and suppression of host defence mechanisms. The viral sense‐strand encodes the coat protein (V1) and V2 protein, while the complementary‐sense strand has six additional ORFs (C1, C2, C3, C4, C5, C6) and a recently identified C7 (Gorovits et al. 2017; Zhao et al. 2018; Liu et al. 2023; Zhao et al. 2023). V2 functions as an RNA silencing suppressor (RSS) protein capable of binding to double‐stranded small interfering RNAs (ds siRNAs) to prevent both transcriptional gene silencing (TGS) and post‐transcriptional gene silencing (PTGS) (Wartig et al. 1997; Rojas et al. 2001; Fukunaga and Doudna 2009; Li et al. 2014; Zrachya et al. 2007; Luna et al. 2012) by disrupting the host RNA interference (RNAi) machinery. Notably, a single amino acid residue of V2 has been shown to mediate its self‐interaction, aggregate formation and pathogenicity of TYLCV (Zhao et al. 2018).
V2 exerts its effects through several distinct mechanisms. It interacts with the key components of the RNA silencing complex (RISC), including proteins Argonaute 4 (AGO4) and Suppressor of Gene Silencing 3 (SGS3) in the RNA silencing pathway, thereby influencing the assembly of the RISC and compromising the antiviral RNA silencing process (Glick et al. 2018). Additionally, V2 interacts with host histone deacetylase 6 to suppress methylation‐mediated transcriptional gene silencing (Wang et al. 2018). It also modulates the activity of CYP1, a papain‐like cysteine protease implicated in plant immune signalling, thereby influencing downstream defence responses (Bar‐Ziv et al. 2015, 2012). Furthermore, V2 cooperates with another TYLCV‐encoded protein, C4, to enhance its accumulation independently of V2 RNA silencing suppression activity (Wang et al. 2022). Beyond its role in pathogenesis, V2 can also facilitate the nuclear export of V1 (CP) encoded by TYLCV (Zhao et al. 2020). Viral proteins frequently engage in interactions with host proteins to enhance the level of infection and suppress plant immunity. Besides V2, C4 also functions as an RSS that inhibits systemic silencing and has been shown to interact with host proteins involved in RNA silencing, translation, ubiquitination, pathogenesis‐related processes, and plant defence responses (Garnelo Gómez et al. 2019; Kim et al. 2016). Accumulating evidence suggests that many RSSs from different plant viruses often interact or target plant antiviral proteins, especially certain transcription factors (TFs) related to disease resistance pathways (Poque et al. 2018; Demi̇rel et al. 2025; Li et al. 2014; Wu et al. 2019). However, the specific functions, functional diversity, and underlying molecular mechanisms of these TFs in mediating antiviral responses remain largely elusive. Elucidating these interactions represents a critical step toward understanding plant–virus coevolution and developing novel strategies for durable disease resistance.
The APETALA2/ETHYLENE RESPONSE FACTOR (AP2/ERF) superfamily of TFs is classified into three separate subfamilies—AP2, RAV (related to ABI3/VP1), and ERF—based on the number of AP2 DNA‐binding domains and sequence similarity (Licausi et al. 2013). Among them, the RAV protein contains AP2 and B3 domains (Yamasaki 2004). Previous studies have demonstrated that RAVs are involved in various aspects of plant development, including axillary bud outgrowth, floral development regulation, and photoperiod regulation (Kagaya et al. 1999; Zhao, Liu, et al. 2021; Sohn et al. 2006; Li et al. 2011; Pei et al. 2024; Zhu et al. 2025; Woo et al. 2010; Kim et al. 2005; Cheng et al. 2023; Xue et al. 2025). Emerging evidence has highlighted the complex regulatory roles of RAV TFs in plants' biotic stress responses (Pre et al. 2008). More recently, RAVs have also been implicated in salicylic acid (SA)‐mediated defence responses (Zhang et al. 2022; Xie et al. 2024; Zhang, Jia, et al. 2025). For instance, the silencing suppressor HC‐Pro encoded by tobacco etch virus (TEV) and p38 encoded by turnip crinkle virus (TCV) can utilise host AtRAV2 to inhibit the host antiviral RNA silencing pathways (Endres et al. 2010). In grapevine, VvRAV1 interacts with p24 encoded by grapevine leafroll‐associated virus 2 (GLRaV2), and this interaction modulates VvRAV1‐mediated regulation of the defence gene VvPR1 (Zhang et al. 2022). Similarly, in pear, PbRAV1 can enhance resistance to apple stem grooving virus (ASGV) by directly interacting with the RSS (CP) encoded by ASGV (Xie et al. 2024). In rice, OsRAV2 interacts with LHCB5, a protein associated with Magnaporthe oryzae resistance, regulates proline, malondialdehyde, hydrogen peroxide and abscisic acid (ABA)‐signalling under drought stress, and enhances resistance to rice powdery mildew by increasing the expression of genes related to disease resistance (Fu et al. 2025). Furthermore, OsRAV15 can interact with OsMYC2, a key integrator of jasmonic acid (JA) signalling, to activate the JA‐mediated disease defence response pathway (Zhang, Huang, et al. 2025). In addition to RAV, other plant proteins associated with disease resistance have also been shown to target viral proteins. For example, WRKY40 modulates tomato mosaic virus (ToMV) infection by altering viral protein levels (Jiang et al. 2021). SlGRXC6 interacts with V2 of TYLCV to inhibit its nuclear export and affect virus infection of the plant (Zhao, Zhou, et al. 2021). HYD1 in cucumber recognises the P1b protein encoded by cucumber vein yellowing virus (CVYV), ultimately affecting CVYV replication (García et al. 2021). Therefore, interactions between TFs and viral proteins constitute a prevalent mechanism by which plants modulate viral infection.
Previous research results have demonstrated that successful virus infection depends on an interaction between the plant and the viral factors as well as the plant's intrinsic defence response (Tan et al. 2024). However, the mechanisms governing viral invasion and specific host factors involved remain incompletely understood. To gain deeper insights into the host–virus interaction network, a TYLCV infection was employed in our previous study, which revealed that TYLCV infection significantly increases the expression of plant genes associated with disease resistance (Zhang, Jia, et al. 2025). Building upon these findings, the present study employed the viral protein V2 as the bait in a yeast two‐hybrid (Y2H) screen of a tomato cDNA library, leading to the identification of an interacting protein belonging to the AP2/ERF family and RAV subfamily. Here, we demonstrate that TYLCV protein V2 interacts with the tomato antiviral protein SlRAV2 (Solyc10g086270.ITAG4.0, GenBank: NM_001320461.2), with reciprocal impacts on their respective functions. Our results reveal a molecular mechanism underlying the role of SlRAV2 in resisting TYLCV systemic infection: SlRAV2 can inhibit the systemic spread of TYLCV by interacting with V2 and influencing its RSS activity, and SlRAV2 also exploits this interaction to enhance its regulation effect on the gene SlPR1, a hallmark gene of the SA‐mediated defence response. These findings provide novel insights into the host–virus interaction landscape and offer valuable guidance for the development of effective strategies to control the infection of TYLCV.
Results
2
Tomato Transcription Factor SlRAV2 Interacts With TYLCV V2
2.1
Previously, using the V2 (GenBank: PP737594) protein encoded by TYLCV as bait, Y2H screening identified an interacting clone encoding a peptide of the C terminus of SlRAV2 from the tomato cDNA library. The sequence analysis revealed that SlRAV2 encodes a protein of 395 amino acids (aa) containing AP2 and B3 domains. The full sequences of SlRAV2 and V2 were cloned into pGADT7 and pGBKT7 vectors, respectively, and co‐transformed into yeast competent cells. Yeast cells co‐expressing SlRAV2 and V2 grew on the synthetic drop‐out medium lacking Trp, Leu, His and Ade (SD/−Leu/−Trp/−His/−Ade) medium, whereas no growth was observed for the negative control (pGBK‐V2/pGAD‐T7) (Figure 1A). To map the interacting region, SlRAV2 was divided into two segments: the N‐terminal region containing the AP2 domain (SlRAV2^N^, 1–150 aa) and the C‐terminal region containing the B3 domain (SlRAV2^C^, 151–395 aa). Y2H assays revealed that SlRAV2^C^ but not SlRAV2^N^ interacted with V2 (Figure 1A). This interaction was further validated in vitro by pull‐down assays, in which GST‐SlRAV2 interacted with V2‐His (Figure 1B), whereas no interaction was detected between GST‐SlRAV2 and the unrelated protein GFP‐His (Figure 1B, lane without band). A bimolecular fluorescence complementation (BiFC) assay in planta further supported the SlRAV2/V2 and SlRAV2^C^/V2 interactions, yielding results consistent with the Y2H and pull‐down experiments (Figure 1C). These results suggest that following TYLCV infection, V2 physically interacts with the plant transcription factor SlRAV2, raising the possibility that SlRAV2, as a potential antiviral protein, may interact with the protein of the virus to resist viral infection. However, the functional relevance of this interaction remains to be elucidated.
Interaction between V2 and SlRAV2. (A) Interactions between V2 and SlRAV2 in yeast. Yeast strain AH109 co‐transformed with plasmids expressing the fusion proteins were plated on SD/−Leu/−Trp/−His/−Ade. Transformation of pGADT7 and pGBK‐V2 served as negative controls. (B) In vitro pull‐down to determine the interaction between V2 and SlRAV2. The combination GFP‐His/GST‐SlRAV2 served as the negative control. Both the input and pull‐down samples were detected by western blotting with anti‐His and anti‐GST antibodies. (C) In vivo bimolecular fluorescence complementation of V2/SlRAV2 interactions. The proteins were expressed in Nicotiana benthamiana leaves. Co‐expression of V2‐YFPN and 35S‐YFPC served as the negative control. Confocal micrographs were taken 3 days post‐infiltration. Bars = 50 μm. Each assay was repeated three times.
The Effect of SlRAV2 on TYLCV Infection
2.2
Previous experimental findings have shown that SlRAV2 expression undergoes significant alterations following viral infection (Zhang, Jia, et al. 2025). Transient gene silencing and overexpression of SlRAV2 in tomato plants revealed contrasting effects on TYLCV accumulation over time. Silencing SlRAV2 resulted in reduced viral accumulation in the infiltrated leaves at 2 days post‐infiltration (dpi); however, viral accumulation increased by 4 dpi (Figure 2A). By 10 dpi, viral accumulation in the systemic leaves exceeded that of the negative control (Figure 2A). Conversely, pre‐inoculation overexpression of SlRAV2 led to elevated TYLCV DNA accumulation in the infiltrated leaves at 2 dpi, followed by a decline at 4 dpi (Figure 2C). At 10 dpi, viral accumulation in systemic leaves was lower than that observed in the negative control (Figure 2C). Collectively, these experiments suggest that SlRAV2 expression modulates viral accumulation in both infiltrated and systemic leaves. Histochemical analysis using trypan blue and 3,3′‐diaminobenzidine (DAB) staining revealed that silencing SlRAV2 enhanced systemic leaf necrosis and elevated reactive oxygen (ROS) levels relative to the control group (Figure 2B). In contrast, transient overexpression of SlRAV2 attenuated TYLCV‐induced necrosis in systemic leaves and reduced ROS accumulation compared with negative control (Figure 2D). Consistent staining outcomes were observed in Nicotiana benthamiana plants (Figure 2B,D). Staining results and corresponding analyses for tomato and N. benthamiana infiltrated leaves are provided in the Figure S1. Given that SlRAV2 overexpression and silencing alter TYLCV levels in systemic and infiltrated leaves of tomato and N. benthamiana, we propose that SlRAV2 may confer resistance to TYLCV by enhancing localised necrosis responses at the infection site, thereby restricting viral spread to the systemic leaves.
*Modulation of SlRAV2 expression levels influences tomato yellow leaf curl virus (TYLCV) DNA accumulation and symptom development. (A) Silencing SlRAV2 increased TYLCV accumulation in tomato systemic leaves. Tomato plants were subjected to virus inoculation 3 days post‐SlRAV2 silencing. Viral DNA levels in both infiltrated and systemic leaves were quantified by quantitative PCR (qPCR) at 2, 4, 10 days post‐inoculation (dpi). Data are presented as relative expression levels compared to the control. qPCR results are shown as mean ± SD of three independent experiments. SD is denoted by error bars. (B) SlRAV2 silencing enhanced staining intensity in tomato systemic leaves (right panel). Staining of systemic leaves with 3,3′‐diaminobenzidine (DAB) and trypan blue after SlRAV2 silencing was more intense than the negative control. Similar results were observed following SlRAV2 silencing in Nicotiana benthamiana (left panel). (C) Overexpression SlRAV2 in tomato reduced TYLCV accumulation in systemic leaves. Tomato plants were subjected to virus inoculation 3 days post‐SlRAV2 overexpression. Expression levels of SlRAV2 and TYLCV were measured in both infiltrated and systemic leaves by qPCR. Data are presented as relative expression levels. (D) SlRAV2 overexpression reduced the DAB and trypan blue staining intensity in tomato systemic leaves (right panel). Similar results were observed following SlRAV2 overexpression in N. benthamiana (left panel). Significant difference, p < 0.05.
SlRAV2 Enhances the Ability of V2 to Suppress RNA Silencing
2.3
SlRAV2 interacts directly with V2, and its expression level influences TYLCV accumulation, suggesting that SlRAV2 may modulate TYLCV infection by regulating the function of V2. To investigate this hypothesis, we performed transient co‐expression assays in N. benthamiana using various plasmid combinations. Under ultraviolet (UV) light, co‐expression of pCambia‐V2 and pCambia‐GFP maintained green fluorescence, while the negative control group (pCambia‐GUS/pCambia‐GFP) showed no obvious fluorescence (Figure 3A, left panel). Notably, co‐infiltration of pCambia‐V2/pCambia‐GFP/pCambia‐SlRAV2 or pCambia‐V2/pCambia‐GFP/pCambia‐SlRAV2^C^ led to enhanced fluorescence intensity compared to the negative control group expressing pCambia‐V2/pCambia‐GFP/pCambia‐GUS at 3 dpi (Figure 3A, middle panel). Western blot analysis confirmed that co‐expression of SlRAV2/V2 or SlRAV2^C^/V2 elevated GFP expression levels (Figure 3B, right panel), further supporting the conclusion that SlRAV2 enhanced the ability of V2 to suppress GFP silencing (Figure 3A,B). Additionally, neither pCambia‐SlRAV2/pCambia‐GFP nor pCambia‐SlRAV2^C^/pCambia‐GFP produced detectable fluorescence signals, and western blot analysis confirmed the absence of GFP expression (Figure 3A, right panel; Figure 3B left panel), indicating that SlRAV2 itself does not possess RSS activity.
SlRAV2 enhanced the silencing suppressor activity of V2 by enhancing its ability of binding double‐stranded (ds) small interfering RNAs (siRNAs). (A) SlRAV2 enhanced the RNA silencing suppressor (RSS) activity of V2. The pCambia‐V2/pCambia‐GFP/pCambia‐SlRAV2 was transiently expressed in Nicotiana benthamiana by Agrobacterium‐mediated transient infection. The GFP fluorescence was observed under a UV light and photographed 3 days post‐infiltration. (B) Western blot results. Total protein was extracted from the GFP fluorescent area and subjected to western blot analysis with anti‐GFP antibodies. Coomassie Brilliant Blue (CBB) staining was used for protein detection to verify equal protein loading. (C) Electrophoretic mobility shift assay (EMSA) results. EMSA showed that SlRAV2 enhanced the ability of V2 to bind to 21‐nucleotide (nt) ds siRNA. GST‐SlRAV2 and V2‐His were expressed in prokaryotes and incubated with biotin‐labelled 21‐nt ds siRNA for EMSA. The addition of GST‐SlRAV2 protein enhanced the ability of V2 to bind to 21‐nt ds siRNA. GFP‐His served as a negative control.
Mechanistic investigations have established that most RSSs inhibit RNA silencing by binding to ds siRNA. Consistent with this model, V2 may also exert its RSS function through a similar mechanism. Using recombinant GST‐SlRAV2 and V2‐His fusion proteins (Figure S2) and biotin‐labelled 21‐nucleotide (nt) ds siRNA probes, electrophoretic mobility shift assay (EMSA) demonstrated that V2 binds to 21‐nt ds siRNA, supporting its role as an RSS (Figure 3C). Given the interaction between SlRAV2 and V2, and the observed modulation of V2's silencing suppression activity, we hypothesised that SlRAV2 may influence this activity by affecting V2's ds siRNA binding capacity. Indeed, EMSA results showed the presence of GST‐SlRAV2 or GST‐SlRAV2^C^ significantly enhanced the binding affinity of V2‐His for 21‐nt ds siRNA (Figure 3C). The EMSA results support the observations in Figure 3A,B, and demonstrate that SlRAV2 enhances the RNA silencing activity of V2 via interaction, specifically through modulation of its ds siRNA‐binding capability.
V2 Enhances SlRAV2‐Mediated Transcription Activation of
SlPR1
2.4
The interaction between SlRAV2 and V2 does not solely affect the function of V2. Previous studies have suggested that the members of the RAV TF family may participate in SA‐mediated defence response (Zhang et al. 2022). Consistent with this notion, overexpression of SlRAV2 resulted in a 1.4‐fold increase in its own transcript level and a 2.9‐fold increase of SlPR1 expression (Figure 4A). To further investigate whether SlRAV2 regulates SlPR1 at the transcriptional level, we cloned the 2000‐bp promoter region of the SlPR1 gene into the pBI121 vector, placing it upstream of the GUS reporter gene (designated SlPR1::GUS). Transient expression assays coupled with a promoter transactivation assay (based on GUS activity quantification) revealed that SlRAV2 enhanced GUS expression compared to the negative control expressing unrelated proteins (Figure 4B). This result suggests that SlRAV2 regulates SlPR1 expression, possibly through direct modulation of its promoter (SlPR1p). Notably, co‐expression of V2 with SlRAV2 further elevated GUS activity to 230% relative to the negative control group (Figure 4C), implying that SlRAV2 may employ V2 to enhance transcriptional activation of SlPR1. EMSA results provided mechanistic insight into this regulation: GST‐SlRAV2 directly bound to a DNA probe containing a conserved ‘CACCTG’ motif derived from SlPR1p. Moreover, the presence of V2‐His enhanced the formation of the GST‐SlRAV2 binding to the SlPR1p (Figure 4D). Taken together, these results indicate that SlRAV2 may utilise V2 to augment its binding affinity to the SlPR1p, thereby reinforcing SlPR1 expression. Given that SlPR1 is a key downstream effector of the SA‐mediated defence response against TYLCV, our results provide indirect evidence that SlRAV2 suppresses systemic TYLCV accumulation, potentially by activating the SA signalling pathway, which in turn upregulates SlPR1 expression to mount an effective defence.
V2 affects the regulation of the SlPR1 gene by interaction with SlRAV2. (A) SlRAV2 has a positive regulatory effect on SlPR1. SlRAV2 was overexpressed using the Agrobacterium‐mediated transient infection technology. The expression of SlPR1 was detected using reverse transcription‐quantitative PCR. ‘’ indicates a significant difference, p < 0.05. (B) SlRAV2 activates the SlPR1 promoter. Leaf tissue from each treatment was harvested at 3 days post‐infiltration, and pooled samples were used for protein extraction and subsequent β‐glucuronidase (GUS) activity assays with 4‐methylumbelliferyl β‐D‐glucuronide (MUG) as the substrate. (C) V2 affects SlRAV2's function. Co‐expression of V2 with SlRAV2 enhanced the SlRAV2‐induced activation of the SlPR1 promoter, as evidenced by a significant increase in GUS activity. ‘’ indicates a significant difference, p < 0.05. (D) V2 enhances binding affinity of SlRAV2 for the ‘CACCTG’ motif in the SlPR1 promoter. GST‐SlRAV2 and V2‐His protein were incubated with a DNA probe for the electrophoretic mobility shift assay. GFP‐His served as a control.
V2 Affects the Subcellular Localisation of SlRAV2
2.5
Because SlRAV2 interacts with V2 and reciprocally influences their respective functions, we hypothesised that their subcellular localisation patterns might likewise be interdependent. Confocal microscope showed that V2 predominantly localised to both the nucleus and cytoplasm, exhibiting granular aggregates within the cytoplasm compartment (Figure 5A). In contrast, SlRAV2 was exclusively localised to the nucleus (Figure 5A). Upon co‐expression in N. benthamiana leaves, however, SlRAV2 localisation shifted significantly—from exclusive nuclear localisation to concurrent nuclear and cytoplasmic distribution—while V2 maintained its original localisation pattern (Figure 5A). To confirm these observations from the perspective of proteins, nuclear‐cytoplasmic fractionation assays were conducted. Consistent with confocal imaging, western blot analysis of isolated fractions demonstrated that both V2‐RFP and GFP‐SlRAV2 were present in the nucleus and cytoplasm when co‐expressed (Figure 5B). The above experiments confirmed that V2 alters the subcellular localisation of SlRAV2, probably facilitating its translocation into the cytoplasm or affecting its translocation into the nucleus.
The V2–SlRAV2 interaction alters the subcellular localisation of SlRAV2. (A) The subcellular localisation of V2 and SlRAV2. Confocal micrographs of Nicotiana benthamiana epidermal cells at 3 days post‐infiltration (dpi). SlRAV2‐GFP localises exclusively to the nucleus (top row). V2‐RFP localises both in nucleus and cytoplasmic (middle row). Co‐expression of both proteins leads to the nuclear and cytoplasmic localisation of each. Scale bar = 50 μm. (B) Detection of V2 and SlRAV2 in nuclear and cytoplasmic fractions. Agroinfiltrated tissues of each treatment were sampled 3 dpi and pooled for protein extraction. Histone H3 and cytosolic fructose‐1,6‐bisphosphatase (cFBPase) served as markers for nuclear and cytoplasmic fractions, respectively, validating the purity of the isolation. Coomassie Brilliant Blue (CBB) staining verified equal protein loading. Upper, nuclear fraction; lower, cytoplasmic fraction. Leaves of N. benthamiana plants expressing the indicated protein(s), and confocal micrographs were taken 3 dpi. Bars = 50 μm.
The observed co‐localisation of V2 and SlRAV2 suggested that V2 may interfere with the nuclear import of SlRAV2, potentially through modulation of interaction between SlRAV2 and Importin α—a key nuclear transport receptor responsible for mediating the nuclear import of proteins harbouring nuclear localisation signals (NLSs). To test this hypothesis, we performed a Y2H assay using pGAD‐SlRAV2 and pGBK‐SlIMP (Importin α). The results showed a direct interaction between SlRAV2 and Importin α (Figure S3). Interestingly, in an in vitro pull‐down assay, the addition of V2‐His diminished the interaction between GST‐SlRAV2 and MBP‐SlIMP (Figure S3), suggesting that V2 competes with Importin α for binding to SlRAV2. Collectively, these findings indicate that the interaction between SlRAV2 and V2 influences SlRAV2's subcellular localisation and modulates its functional activity—probably by interfering with its nuclear import machinery.
Discussion
3
TYLCV is a widespread pathogen responsible for tomato yellow leaf curl disease, leading to substantial reductions in tomato yields. Given that plant antiviral proteins can suppress viral infection through direct interactions with the viral‐encoded proteins, we report the functional relationship between the tomato transcription factor SlRAV2 and the V2 protein encoded by TYLCV. This interaction enhanced TYLCV accumulation transiently in the infiltrated leaves, accompanied by an increase in the degree of cell necrosis; subsequently, it decreased local viral levels and inhibited the spread of TYLCV to the systemic leaves.
Previous studies using a TYLCV infectious clone experiments identified marked alterations in disease resistance‐related genes in both tomato and N. benthamiana plants. Among them, one RAV gene (Solyc10g086270.ITAG4.0) was significantly upregulated in TYLCV‐infected tomato plants (Zhang, Jia, et al. 2025). Based on these observations, we hypothesised that modulation of SlRAV2 expression might influence TYLCV accumulation. Indeed, transient overexpression of SlRAV2 in the TYLCV‐infected leaves initially led to a temporary increase in viral accumulation in the infiltrated leaves, followed by a reduction in viral levels in the infiltrated leaves and inhibiting the spread of TYLCV to the systemic leaves (Figure 2B,D). Conversely, silencing SlRAV2 resulted in more extensive systemic infection (Figure 2A,C). Thus, these findings suggest that SlRAV2 may contribute to the early plant response to TYLCV, although its precise role requires further validation using stable transgenic approaches. Overexpressing SlRAV2 also led to elevated ROS content and increased the number of necrotic spots in the infiltrated leaves, thereby inhibiting the systemic TYLCV spread. Similar phenomena have been reported in previous studies, wherein modulation of ROS levels enhance plant resistance to viruses (Hernández et al. 2016; Kozieł et al. 2024; Raza et al. 2021; Su et al. 2021; Hong et al. 2005). Changing the ROS content in plants can lead to a rapid outbreak of viruses within the plants, resulting in inhibition of the systemic infection of viruses (Király et al. 2025; Luo et al. 2023). Consistent with these reports, our finding suggests that SlRAV2 indirectly enhanced the hypersensitive response (HR), resulting in a local increase in the ROS content and initiation of the plant defence response, thereby inhibiting virus spread. Furthermore, DAB and trypan blue staining were less visually striking in infiltrated leaves compared to systemic leaves (Figure 2B,D). At 2 dpi, no significant differences in staining intensity were observed between the experimental groups and the control; by 4 dpi, leaves overexpressing SlRAV2 exhibited slightly intensified DAB and trypan blue staining, whereas SlRAV2‐silenced leaves displayed reduced staining (Figure S1). These results indicate that SlRAV2 may promote a transient acceleration of viral replication in infiltrated leaves, thereby raising the ROS levels in the cells, accelerating cell necrosis, and inhibiting virus spread from the infiltrated leaves to the systemic leaves. This result is similar to the one published by Sun and Folimonova (2019) and Sun et al. (2021), who showed that p33 encoded by the citrus tristeza virus (CTV) can be specifically recognised by the plant defence response, increasing the ROS accumulation, and thereby controlling virus accumulation and spread. These findings show that the strategy for inhibiting systemic viral infection mentioned in this article is not uncommon.
As an antiviral TF, RAV may be subject to viral attack or may defend against viral infection. For instance, PbRAV1 from pear interacts with the RSS of ASGV, thereby enhancing antiviral defence (Xie et al. 2024). Similarly, VvRAV1 from grapevine activates the SA‐mediated antivirus signalling (Zhang et al. 2022), while OsRAV15 in rice activates the JA‐mediated antiviral immunity (Zhang, Huang, et al. 2025). In this study, we found an interaction between V2 and SlRAV2 through Y2H screening of a tomato cDNA library, and subsequent validation using Y2H, pull‐down, and BiFC experiments confirmed this interaction (Figure 1A–C). Further truncation analysis showed that the B3 domain (204‐306 aa) of SlRAV2 was sufficient for the interaction with V2 in yeast (Figure S4), it revealed that B3 is the key functional domain for the SlRAV2/V2 interaction site. Interestingly, SlRAV2 and V2 seem to interact mainly within the nucleus. Confocal microscopy revealed distinct nuclear colocalisation of the two proteins, with only weak punctate green fluorescence visible in the cytoplasm (Figure 1C). Therefore, based on their interaction, we hypothesised that the subcellular localisation of both proteins might be affected—a prediction subsequently validated by our experimental results (Figure 5A). Consistent with previous reports in rice (Guo et al. 2024), SlRAV2 localised exclusively to the nucleus in plants, while V2 exhibited both nuclear and cytoplasmic distribution, with forming small aggregates in the endoplasmic reticulum—also aligning with earlier studies (Zhang et al. 2012; Zhao et al. 2020; Zhao, Zhou, et al. 2021; Zhao et al. 2023; Hak et al. 2015; Moshe et al. 2015). The modulation of SlRAV2's subcellular localisation by V2 represents a key regulatory node in their interaction. While V2 did not alter its own nuclear‐cytoplasmic distribution, it significantly restricted SlRAV2's nuclear accumulation, leading to their co‐presence in both compartments (Figure 5B). This effect is mechanistically linked to V2's interference with SlRAV2's interaction with Importin α (Figure S3)—a critical nuclear import mediator (Xiao et al. 2025)—thereby partially sequestering SlRAV2 in the cytoplasm. This finding contrasts with previous reports that V2 facilitates the nuclear entry of certain proteins (Zhao et al. 2020, 2023), highlighting the functional role of V2 in modulating nuclear transport mechanisms. Therefore, V2 affects SlRAV2 localisation, and the functional interaction between V2 and SlRAV2 might be due to the influence of their subcellular localisation.
Plants use Argonaute (AGO) proteins as the main components of RISC, which together with Dicer‐like enzymes, process exogenous viral RNA into siRNAs (Alazem and Lin 2015; Duan et al. 2012). In the presence of RNA‐dependent RNA polymerase, the RNA silencing signal is continuously amplified and transmitted throughout the plant to resist viral infection. Consequently, disruption of siRNA biogenesis or mobility compromises RNA silencing signals. V2 acts as an RSS and binds to ds siRNAs, inhibiting RISC assembly and preventing systemic silencing (Zhang et al. 2012). Here we demonstrated that V2 interacts with the host protein SlRAV2, altering the subcellular localisation of SlRAV2. This interaction represents a mutual regulatory mechanism impacting both viral and host protein functions. Endres et al. (2010) reported that potyvirus HC‐Pro and carmovirus p38 require the ethylene‐inducible host transcription factor RAV2 to block RNA silencing. Xie et al. (2024) and Zhang et al. (2022) also showed that in pear or in grapevine RAV1 enhances the ability of ASGV and GLRaV‐2 RSS to bind to 21‐nt ds siRNAs. Our results are similar to a previous experiment; prokaryotically expressed SlRAV2 enhanced the ability of V2 to bind to 21‐nt ds siRNAs (Figure 3C). The finding indicates that SlRAV2 enhances the silencing suppression activity of V2 by influencing its ability to bind to 21‐nt ds siRNA. Moreover, the full sequence and C‐terminal region of SlRAV2 interacted with V2, thereby influencing its RSS function, but neither SlRAV2 nor SlRAV2^C^ bound to 21‐nt ds siRNA (Figure 3C). These results reveal that SlRAV2's regulatory effect on V2 is mediated through protein–protein interaction rather than direct RNA binding itself. Mechanistically, V2 influenced SlRAV2's nuclear‐cytoplasmic partitioning, partially retaining SlRAV2 in the cytoplasm where V2 resides, thereby increasing local ds siRNA binding and enhancing V2's localised RSS activity. Agrobacterium‐mediated transient infection experiments also confirmed that SlRAV2 indeed enhanced the RSS activity of V2 as a result of its subcellular redistribution (Figure 3A). Collectively, integrating the results from Figures 1, 2, 3, SlRAV2 augments V2's RSS efficacy by protein interaction and strengthening its ds siRNA‐binding capacity, leading to a transient increase in TYLCV accumulation and ROS content in infiltrated tissues. Based on our analysis, we propose that the mode of action of SlRAV2 may involve amplifying viral disease symptoms, thereby prematurely and rapidly inducing localised leaf necrosis, which in turn restricts systemic viral spread. We speculate that this mechanism represents an antiviral strategy mediated by RAV proteins.
V2 interacts with SlRAV2 and alters its nuclear localisation, which potentially impacts SlRAV2's transcriptional regulatory function. RAV family proteins are known to act as TFs; for instance, VvRAV1 can regulate VvPR1 and enhance the SA‐mediated defence response in grapevine (Zhang et al. 2022). Thus, SlRAV2 probably has a regulatory effect on the SlPR1 gene, a pathogenesis‐related gene activated upon pathogen attack and instrumental in SA‐dependent antiviral defence (Shi et al. 2025; Essa et al. 2025; Zhou et al. 2025). Some reports indicate that plant virus infection elevates PR genes expression (Demi̇rel et al. 2025; Zanini et al. 2025; Zhang, Jia, et al. 2025), and we suspect this may be mediated by SlRAV2. Indeed, overexpression of SlRAV2 resulted in an increase in SlPR1 expression (Figure 4A); thus, SlRAV2 positively regulates SlPR1. Transient expression assays revealed that SlRAV2 directly activated the SlPR1 promoter, as co‐expression with SlRAV2 significantly elevated GUS activity (Figure 4B). This effect was further enhanced by V2 (Figure 4C), indicating that V2 enhances SlRAV2's regulatory capacity. EMSA confirmed SlRAV2's specific binding to a conserved ‘CACCTG’ motif recognised by the B3 domain (Yamasaki 2004; Kagaya et al. 1999), and V2 strengthened this binding (Figure 4D). This V2‐mediated enhancement contrasts with a report in grapevine, where a viral protein attenuated the DNA‐binding ability of VvRAV1 (Zhang et al. 2022), highlighting species‐specific regulatory outcomes. Together, SlRAV2 not only enhances V2's RSS activity—facilitating transient TYLCV accumulation in infiltrated leaves, but also synergises with V2 to amplify SA‐mediated defence via SlPR1 activation. This dual effect may trigger elevated ROS production and localised necrotic lesions, thereby restricting systemic viral spread. These two findings provide a relatively complete account of the antiviral role of SlRAV2.
There is an arms race between viruses and plants. TYLCV avoids excessive cell death to preserve a viable host environment necessary for long‐term infection, actively suppressing plant cell necrotic response (Gorovits et al. 2022). This study demonstrates that SlRAV2 counters this stealth strategy by interacting with RSS V2. Based on our findings, we propose a working model illustrating how SlRAV2 inhibits the systemic infection of TYLCV (Figure 6). In this model, V2 employs SlRAV2 to bind 21‐nt ds siRNA by changing the subcellular localisation of SlRAV2 and effecting the interaction between SlRAV2 and Importin α, thereby augmenting V2's RSS activity. Concurrently, nuclear‐localised V2 potentiates SlRAV2‐mediated transcriptional activation of the SlPR1p. This dual modulation transiently elevates local TYLCV accumulation and ROS levels, triggering localised necrosis that restricts viral spread. From a holistic perspective, while it may appear that V2 is exploiting SlRAV2, in reality it is SlRAV2 that is leveraging V2 to mount an antiviral defence. It is noteworthy that although SlRAV2 was demonstrated to restrict systemic viral infection, necrotic symptoms were still observed in systemic tissues, indicating potential limitations of this defence strategy. While V2 modulates SlRAV2 nuclear import and potentially reduces its regulation of SlPR1, its cooperative enhancement of SlRAV2 activity predominates, resulting in overall increased regulation of SlPR1. We speculate that V2 exerts dual effects on RAV function: it may interfere with certain aspects while partially enhancing others. Consistent with this, V2 does promote a transient rise in viral accumulation; however, this increase is short‐lived, probably due to the counteracting activity of SlRAV2. Eventually, SlRAV2 utilises V2 to achieve the effect of resisting viral infection. The final outcome probably reflects a dynamic balance between viral infection and host defence, which may be influenced by plant developmental stage and environmental conditions. Furthermore, the antiviral function of SlRAV2, particularly its stability, requires further investigation.
Working model for the SlRAV2/V2 interaction. The interaction between SlRAV2/V2 changes the subcellular localisation of SlRAV2, some SlRAV2 protein enters the cytoplasm and enhances the ability of V2 to bind 21‐nucleotide double‐stranded small interfering RNA (siRNA), causing the accumulation level of tomato yellow leaf curl virus (TYLCV) to surge in the infiltrated leaves. In the nucleus, SlRAV2 utilises V2 to enhance the effect on the SlPR1 promoter and finally inhibit TYLCV systemic infection.
Experimental Procedures
4
Materials and Methods
4.1
Wild‐type N. benthamiana plants and tomato seedlings ( Solanum lycopersicum ‘Jinpeng No. 1’) with non‐detectable TYLCV were used in this study. The seedlings were grown in individual pots containing commercial soil at 23°C–25°C with a 16‐h light regime, and 4‐week‐old plants were used.
Plasmid Construction
4.2
Total RNA was extracted from tomato leaves, followed by reverse transcription to generate complementary DNA (cDNA), which served as the template for PCR amplification of SlRAV2. The V2 gene was amplified from an infectious clone of TYLCV (this plasmid was constructed and stored by our laboratory). SlRAV2 and V2 were inserted into the pMD19‐T intermediate vector.
Sequences of SlRAV2, SlRAV2 ^C^, SlRAV2 ^N^, SlRAV2 ^B3^ and V2 were PCR amplified with primer pairs F1/R1, F2/R2, F3/R3, F4/R4 and F5/R5, respectively. PCR products were digested with BamHI/XhoI (for SlRAV2, SlRAV2 ^C^, SlRAV2 ^N^, SlRAV2 ^B3^) or EcoRI/SalI (for V2) and cloned into pGAD‐T7 and pGBK‐T7.
Then, sequences of SlRAV2, SlRAV2 ^C^, SlRAV2 ^N^, V2, GFP and GUS were PCR amplified using F6/R6, F7/R7, F8/R8, F9/R9, F10/R10 and F11/R11 primer pairs, respectively. PCR products were digested with XhoI/BamHI (for SlRAV2, SlRAV2 ^C^, and SlRAV2 ^N^), XhoI/SalI (for V2) and XhoI/BamHI (for GFP and GUS) and cloned into pCambia1300. V2 was PCR amplified with primer pair F12/R12, and the PCR products were digested with XhoI/SalI and cloned into pCambia1300‐RFP. SlRAV2 was PCR amplified with primer pair F13/R13, and PCR products were digested with XhoI/BamHI and cloned into pCambia1300‐GFP. V2 and GFP sequences were PCR amplified with F14/R14 and F15/R15 primer pairs, respectively. PCR products were digested with SalI/XhoI (for V2) or BamHI/XhoI (for GFP) and cloned into pET28a. SlRAV2 sequences were amplified with primer pair F16/R16, and the PCR products were digested with BamHI/XhoI and inserted into pGEX‐4T‐1 to generate GST‐fusion constructs.
V2, SlRAV2, SlRAV2 ^C^ and SlRAV2 ^N^ sequences were PCR amplified with F17/R17, F18/R18, F19/R19, and F20/R20 primer pairs, respectively. PCR products were digested with BamHI/XhoI (for SlRAV2, SlRAV2 ^C^ and SlRAV2 ^N^) or SpeI/XhoI (for V2) and cloned into BiFC vectors pSPYNE‐35S and pSPYCE‐35S. SlRAV2 sequences were PCR amplified with primer pair F21/R21. The PCR products were digested with EcoRI/BamHI and cloned into TRV vectors for gene silencing.
Finally, the SlPR1 promoter region was PCR amplified from tomato leaves using primer pair F22/R22. The PCR products were digested with PstI/SmaI and inserted into pBI121 to produce pBI121‐SlPR1p::GUS. All other constructs used in this study were described previously. The primers mentioned above are listed in Table S1.
Y2H Experiment
4.3
Co‐transformations were carried out with pairs of plasmids introduced into the yeast strain AH109 via the small‐scale yeast transformation protocol provided by the manufacturer (Coolaber). AH109 was streaked on solid YPDA medium and grown at 30°C until single colonies appeared. One colony was selected and inoculated into broth until the OD_600_ reached 0.3. Yeast cells were centrifuged at 700g, washed and resuspended in 1.1 × Tris‐EDTA/lithium acetate buffer. For each reaction, 100 μL competent cells were mixed with 20 μL carrier DNA, 300 μL 50% polyethylene glycol, and the plasmid pair, then incubated at 30°C for 30 min, followed by the addition of 20 μL dimethyl sulppoxide and heat shocked at 42°C for 20 min. After recovery in 1 mL YPD‐Plus at 30°C, cells were resuspended in 0.9% NaCl and plated on the yeast double‐dropout medium (SD/−Trp−Leu) and quadruple‐dropout medium (SD/−Trp−His−Leu−Ade). Then, the yeast strain was cultivated at 30°C for about 3–4 days.
Prokaryotic Expression and Protein Purification
4.4
The fusion protein expression vector was transformed into Escherichia coli BL21 and cultivated at 37°C. Next, IPTG was added to induce protein expression, followed by using the magnetic beads containing GST‐ or His‐tags, following the manufacturer's instructions (Novagen) to purify the GST‐ or His‐tagged fusion protein (Zhang et al. 2022).
Extraction of Total Protein From Plants and Western Blot Experiment
4.5
The protein extraction and western blot experiments were performed using the Fast Western Detection kit (Seven) following the protocol provided in the kit. The film was then photographed to obtain the image.
Protein extraction and western blot analyses were performed as described previously (Li et al. 2018). Dilutions of antibodies or antiserum (noncommercial antibody) were 1:5000 (anti‐GFP, anti‐RFP, anti‐GST, anti‐His and goat anti‐mouse) or 1:2000 (anti‐histone H3, anti‐cFBPase and anti‐MBP). The hybridisation signals were detected using an enhanced chemiluminescence system (eECL Western Blot Kit; ComWin).
Agrobacterium‐Mediated Transient Infection
4.6
Plasmids pCambia1300‐V2, pCambia1300‐SlRAV2 and pCambia1300‐GFP were transformed into Agrobacterium tumefaciens competent cells. Single colonies were selected and cultured at 28°C for 72 h. Bacterial broth cultures were adjusted to OD_600_ of 0.8, harvested by centrifugation at 5000 rpm for 5 min and resuspended in infiltration buffer containing 10 mM MgCl_2_, 10 mM MES and 100 μM acetosyringone. Infiltration was performed by drawing approximately 200–500 μL of the bacterial suspension into a 1 mL syringe and injecting it into the abaxial side of leaves of N. benthamiana plants and tomato seedlings.
In‐Vitro Pull‐Down Assay
4.7
Fusion proteins were incubated in binding buffer (50 mM Tris–HCl pH 7.5, 100 mM NaCl, 0.25% w/v Triton X‐100, 35‐mM β‐mercaptoethanol) for 2 h at 4°C. Then, 25 μL of His‐Trap (Novagen) was added and the mixture was incubated for an additional 1 h before detecting the pulled‐down proteins using the anti‐GST antibody.
EMSA
4.8
The EMSA experiment was conducted using the chemiluminescent EMSA kit (Beyotime). Two biotin‐labelled single‐stranded RNA (sRNA1 and sRNA2) oligonucleotides synthesised by Sangon Biotech were annealed to form 21‐nt ds siRNA probes (sRNA1: 5′‐GUCACUAUGGGUUUAUGG‐3′ and sRNA2: 5′‐CAUAACCAUAGUAGUUGACUG‐3′). Equal amounts (1 ng) of biotin‐labelled DNA or ds siRNA probe were incubated with 2 μg of GST or His‐labelled fusion protein or a mixture of the two proteins. Electrophoresis was performed using a 6% polyacrylamide gel in a buffer (20 mM EDTA, 890 mM Tris‐H_3_BO_3_, pH 8.3). The biotin‐labelled probe was detected using chemiluminescence, and each experiment was repeated three times.
qPCR and RT‐qPCR
4.9
qPCR was performed as previously described (Zhang et al. 2022), using the SYBR PrimeScript real‐time PCR Kit (Vazyme), and the cDNA transcribed from RNA as the template. TYLCV genomic DNA was extracted directly using an EZ‐10 Spin Column Plant Genomic DNA Purification Kit (Sangon, Shanghai, China). The following primers, F23/R23, F24/R24, F25/R25 and F26/R26, were used for analysing the relative levels of mRNA (SlRAV2, SlPR1 and Actin) and viral DNA. The primers mentioned above are listed in Table S1.
Confocal Microscopy
4.10
Fluorescence signals were observed and recorded under a FluoView 3000 confocal microscope (Olympus) equipped with FluoView FV10‐ASW 4.0 Viewer software (Olympus). GFP fluorescence signals were excited at 488 nm, and the signal was captured at 500–540 nm. RFP was excited at 543 nm, and the signal was captured at 570–670 nm. Images were captured using 5% of the maximum light intensity value and a gain of 500–600.
Promoter Transactivation Assay
4.11
Recombinant plasmids were introduced into N. benthamiana plants by agroinfiltration. The quantitative GUS activity assay was performed as described by Blázquez (2007) using 4‐methylumbelliferyl β‐D‐glucuronide as the substrate.
Nuclear‐Cytoplasmic Fractionation
4.12
Infiltrated leaf patches were sampled 3 dpi and pooled. Nuclear‐cytoplasmic fractionation was performed using the Nuclear and Cytoplasmic Protein Extraction Kit (Beyotime) following the manufacturers' protocols.
Statistical and Phylogenetic Analyses
4.13
The statistical analyses were performed using SPSS software (SPSS Inc.), and significant differences between treatments were evaluated with Tukey's multiple range test.
Author Contributions
Chenwei Zhang: conceptualization, software, investigation, methodology, data curation, validation, formal analysis, supervision, visualisation, project administration, resource, writing – original draft, writing – review and editing, funding acquisition. Xin Jia: conceptualization, software, writing – review and editing, investigation, software, data curation. Guiyan Fan: methodology, data curation, software, investigation. Xiaoli Ren: methodology, software, data curation, visualisation. Xing Han: methodology, data curation, software. Xiaocong Jiao: investigation, methodology, validation. Yuan Cheng: methodology, software, data curation. Yajuan Cheng: data curation, validation, investigation. Zihan Liu: data curation, validation, investigation. Zhehao Yuan: data curation, validation, investigation. Yongmin Chen: conceptualization, resources, project administration, supervision, writing – review and editing.
Funding
This work was supported by National Natural Science Foundation of China, 32302527.
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Figure S1: The result of DAB and trypan blue staining in infiltration leaves.
Figure S2: Purification of the related prokaryotic expressed protein.
Figure S3: V2 affects the interaction between SlRAV2 and SlIMP.
Figure S4: The interaction between SlRAV2^B3^ and V2.
Table S1: Primer sequences.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alazem, M. , and N. S. Lin . 2015. “Roles of Plant Hormones in the Regulation of Host Virus Interactions.” Molecular Plant Pathology 16: 529–540.25220680 10.1111/mpp.12204 PMC 6638471 · doi ↗ · pubmed ↗
- 2Bar‐Ziv, A. , Y. Levy , V. Citovsky , and Y. Gafni . 2015. “The Tomato Yellow Leaf Curl Virus (TYLCV) V 2 Protein Inhibits Enzymatic Activity of the Host Papain‐Like Cysteine Protease CYP 1.” Biochemical and Biophysical Research Communications 460, no. 3: 525–529.25797621 10.1016/j.bbrc.2015.03.063PMC 4439401 · doi ↗ · pubmed ↗
- 3Bar‐Ziv, A. , Y. Levy , H. Hak , et al. 2012. “The Tomato Yellow Leaf Curl Virus (TYLCV) V 2 Protein Interacts With the Host Papain‐Like Cysteine Protease CYP 1.” Plant Signaling & Behavior 7, no. 8: 983–989.22827939 10.4161/psb.20935 PMC 3474700 · doi ↗ · pubmed ↗
- 4Blázquez, M. 2007. “Quantitative GUS Activity Assay of Plant Extracts.” CSH Protocols 2007: pdb.prot 4690.21357025 10.1101/pdb.prot 4690 · doi ↗ · pubmed ↗
- 5Cao, X. , M. Huang , S. Wang , T. Li , and Y. Huang . 2024. “Tomato Yellow Leaf Curl Virus: Characteristics, Influence, and Regulation Mechanism.” Plant Physiology and Biochemistry 213: 108812.38875781 10.1016/j.plaphy.2024.108812 · doi ↗ · pubmed ↗
- 6Cheng, H. , Q. Wang , Z. Zhang , et al. 2023. “The RAV Transcription Factor TEMPRANILLO 1 Involved in Ethylene‐Mediated Delay of Chrysanthemum Flowering.” Plant Journal 116, no. 6: 1652–1666.10.1111/tpj.1645337696505 · doi ↗ · pubmed ↗
- 7Demi̇rel, S. , M. Usta , G. Korkmaz , et al. 2025. “Recombinant Tritin Protein Exhibits Antiviral Activity Against Zucchini Yellow Mosaic Virus.” BMC Plant Biology 25, no. 1: 978.40731260 10.1186/s 12870-025-07080-x PMC 12305900 · doi ↗ · pubmed ↗
- 8Duan, C. G. , Y. Y. Fang , B. J. Zhou , et al. 2012. “Suppression of Arabidopsis ARGONAUTE 1‐Mediated Slicing, Transgene‐Induced RNA Silencing, and DNA Methylation by Distinct Domains of the Cucumber Mosaic Virus 2b Protein.” Plant Cell 24, no. 1: 259–274.22247253 10.1105/tpc.111.092718 PMC 3289565 · doi ↗ · pubmed ↗