The effects of early life pain and juvenile fear conditioning on CRF-receptor expression in the amygdala and hypothalamus of the juvenile rat
Michael A. Burman, Jared T. Zuke

TL;DR
This study explores how early life pain and fear in young rats affect CRF receptor expression in brain regions linked to stress and pain, finding modest sex-dependent changes.
Contribution
The study investigates how early life pain and juvenile fear conditioning affect CRF receptor mRNA expression in specific brain regions of juvenile rats.
Findings
Early life pain and fear conditioning had modest effects on CRF receptor expression in the amygdala and hypothalamus.
Fear conditioning increased crhr2 mRNA in the CeA only in neonatally undisturbed subjects.
Changes in crhr2 mRNA in the BLA and crhr1 mRNA in the PVN were sex-specific.
Abstract
Early life pain and stress have lasting consequences on nervous system development that can interact with later stress or trauma to create a susceptibility to fear, anxiety, depression and chronic pain among other psychological disorders. Recent work has identified changes in corticotropin releasing factor signaling in limbic system structures, such as the amygdala and hypothalamus, as a key mechanism behind these changes – albeit in a sex-dependent manner. CRF has two major receptors, CRFR1 and CRFR2 which have also been shown to play key roles in fear and pain expression. The current work examines the effects of early life pain designed to mimic the neonatal medical trauma that occurs in the Neonatal Intensive Care Unit (NICU), paired with a juvenile trauma in the form of fear conditioning, on expression of crhr1 and crhr2 mRNA in the central nucleus (CeA) and basolateral nucleus…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
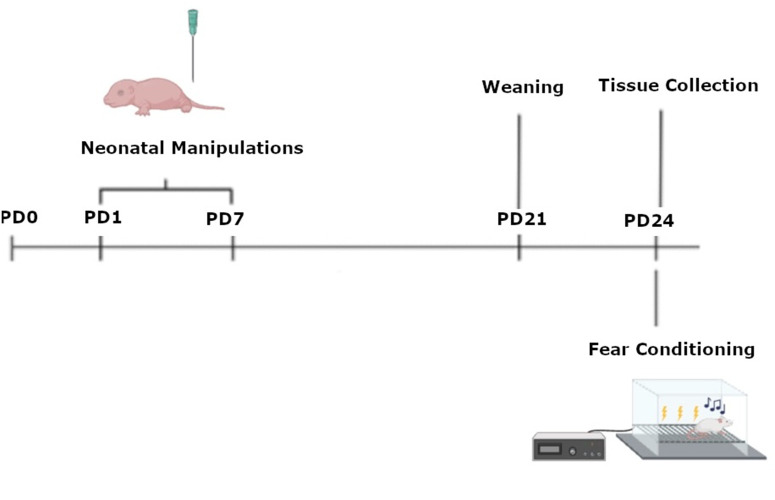 Fig 1
Fig 1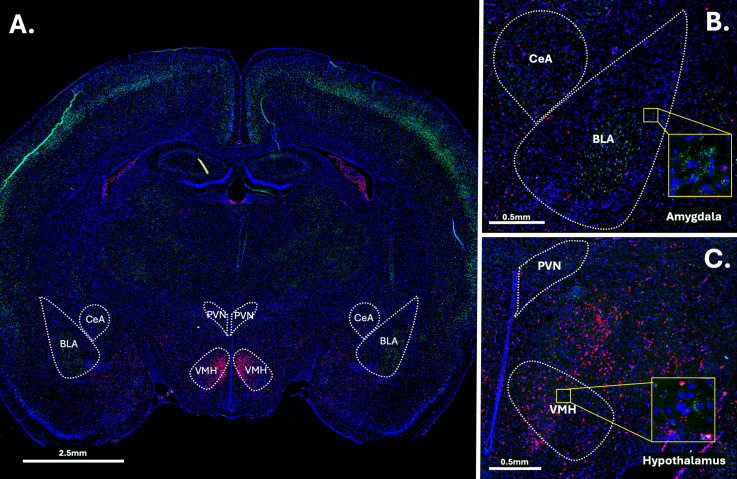 Fig 2
Fig 2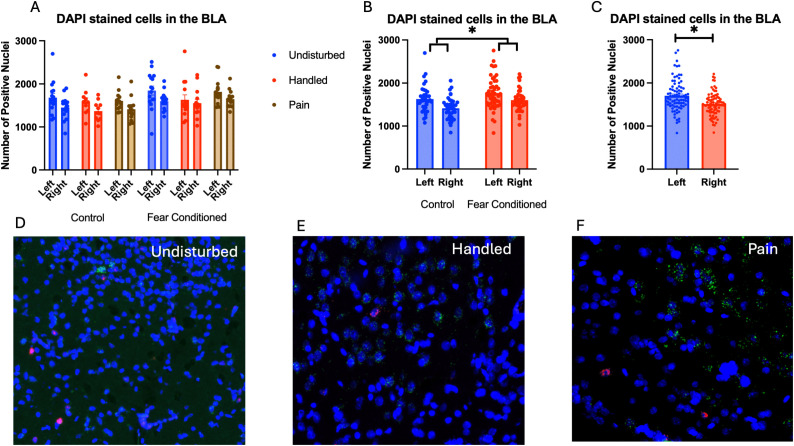 Fig 3
Fig 3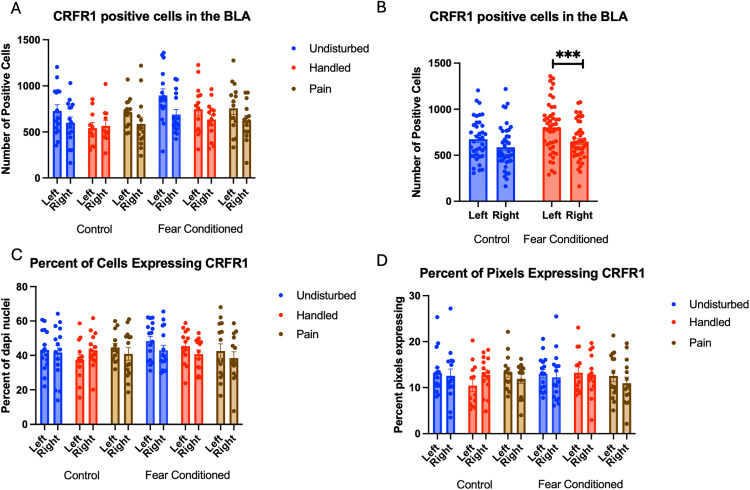 Fig 4
Fig 4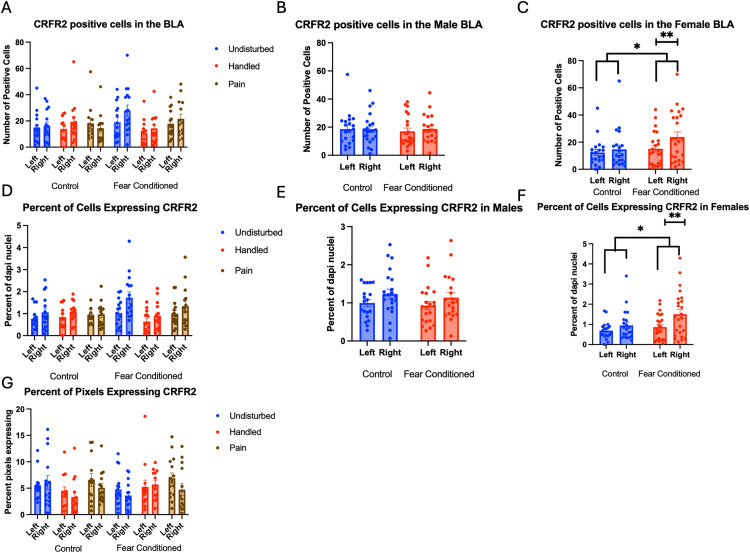 Fig 5
Fig 5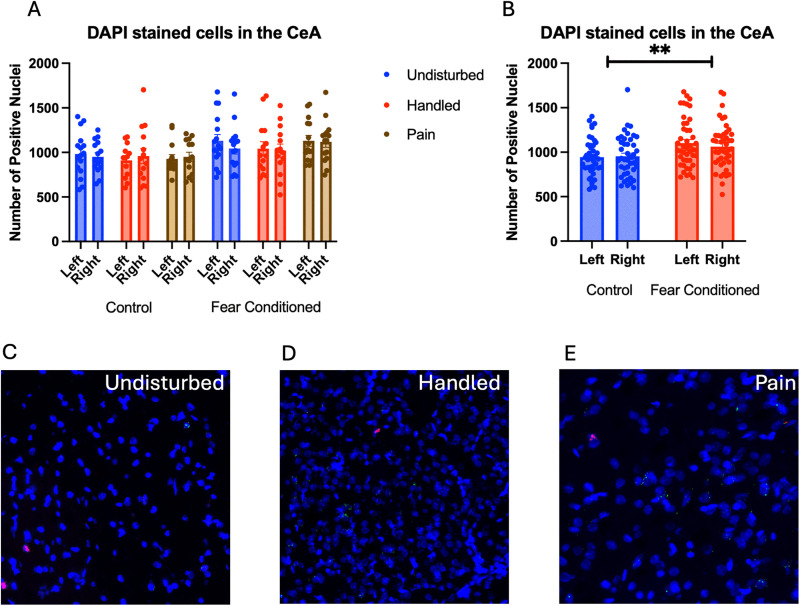 Fig 6
Fig 6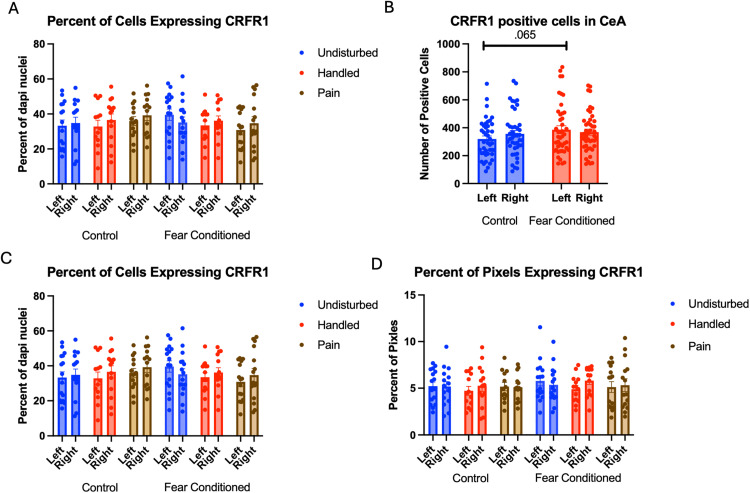 Fig 7
Fig 7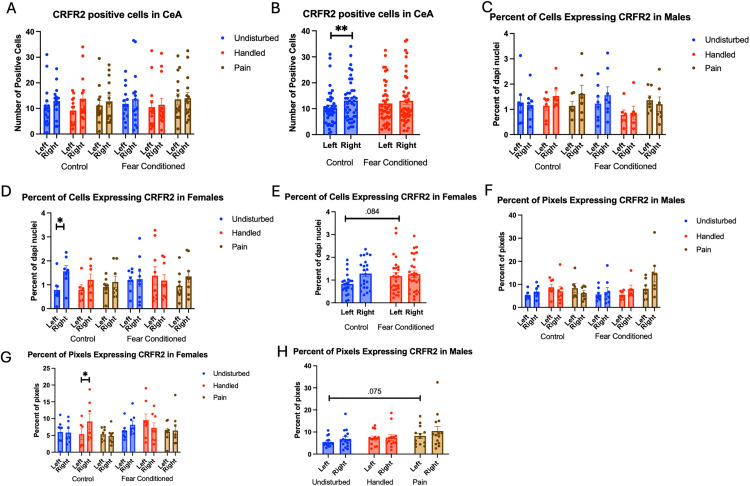 Fig 8
Fig 8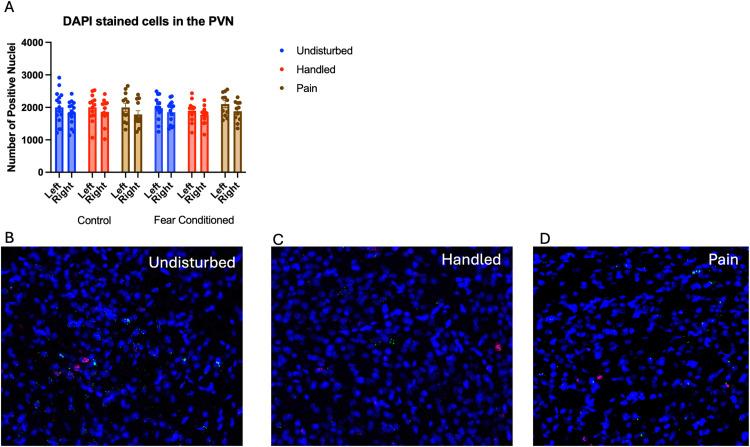 Fig 9
Fig 9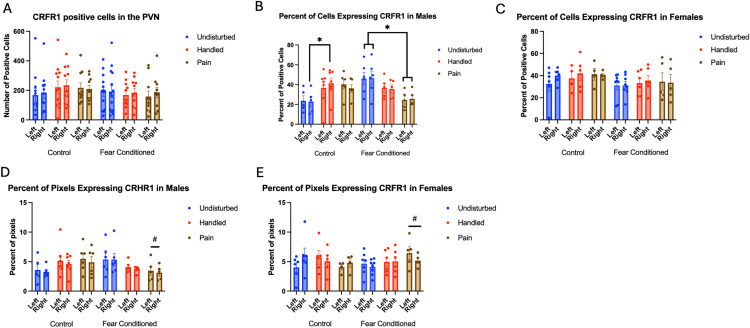 Fig 10
Fig 10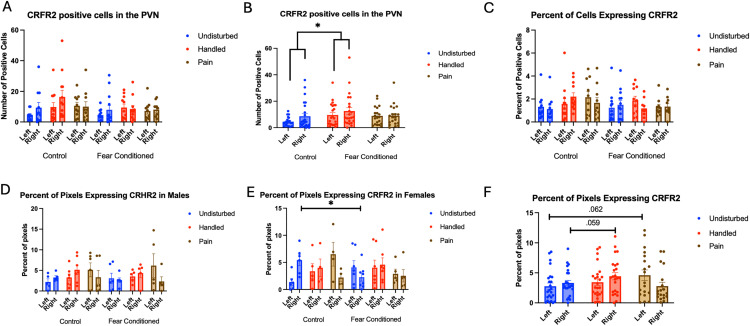 Fig 11
Fig 11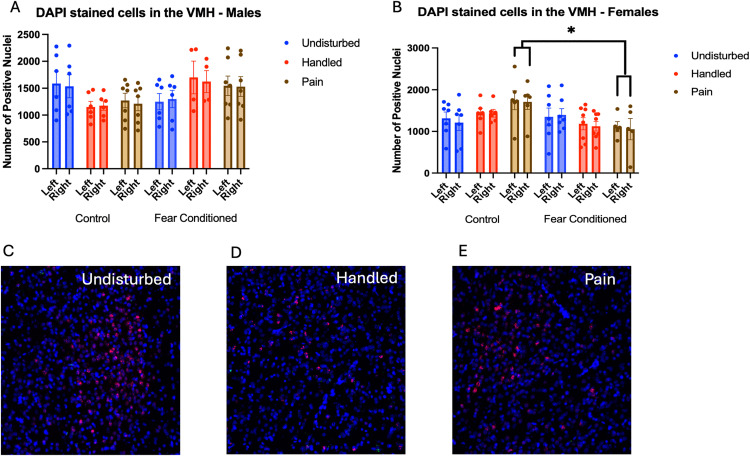 Fig 12
Fig 12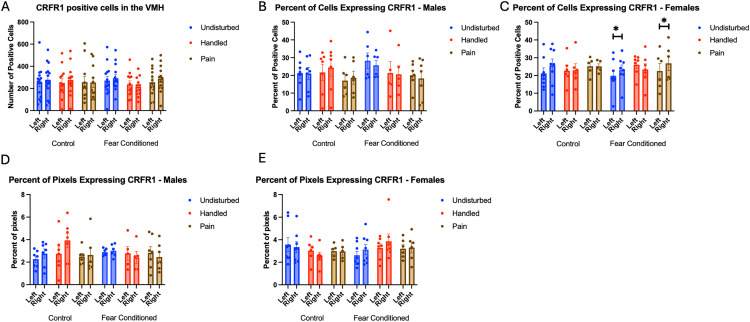 Fig 13
Fig 13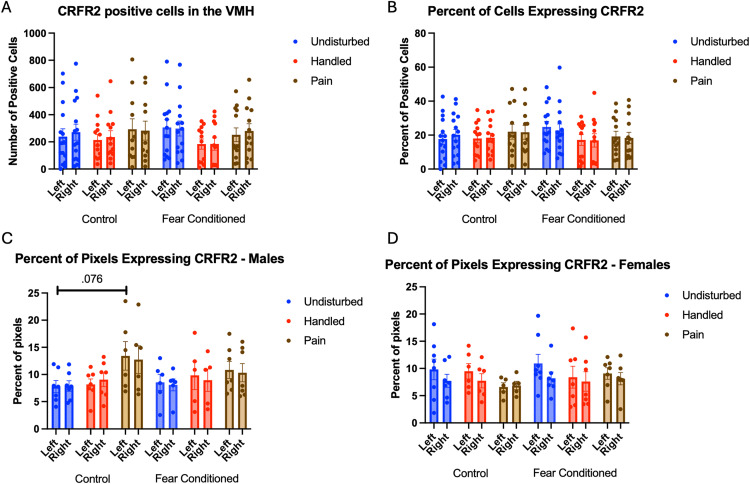 Fig 14
Fig 14- —http://dx.doi.org/10.13039/100000057National Institute of General Medical Sciences
- —http://dx.doi.org/10.13039/100000057National Institute of General Medical Sciences
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsStress Responses and Cortisol · Tryptophan and brain disorders · Pediatric Pain Management Techniques
Introduction
It is now well-established that early life pain and stress have both acute and lasting consequences on brain development and affective behaviors, stress responsiveness, and pain thresholds in both clinical populations and pre-clinical animal models [1–11]. One source of early life adversity is medical trauma that occurs in the neonatal intensive care unit (NICU) in premature or sick neonates. While increases in NICU admissions have coincided with lower levels of infant mortality [12–14], the pain and stress that occur in those settings has been linked with later life dysfunction including mental health vulnerability [6,7,15–19]. However, the mechanisms underlying these changes are still being elucidated, which is a critical impediment to the proper prevention, diagnosis, and treatment of neonates who are exposed to early life medical trauma, such as the repeated heel lancing and other forms of pain and stress that occur in the NICU.
Early life events have been well established to have lasting impacts on developmental trajectory of the brain and nervous system, despite the existence of infantile amnesia and stress hyporesponsive periods [20–26]. While acute changes in behavior during aversive stimulation are sometimes absent in neonates, as they must continue to stay close to sources of food and protection despite any painful or adverse stimulation [27], it is now clear that plasticity during critical periods of development is necessary to prepare organisms for the particular environmental challenges they will face. Thus, early life adversity can serve to prepare organisms for stressful or painful futures by priming either resilience or vulnerability, which then can manifest during later-life stress or trauma (often called a “two-hit model” when discussing mental health outcomes) [28–31]. Indeed, pre-clinical models have demonstrated several effects of early life pain and stress. For example, brief handling and maternal separation paradigms have also been commonly used to examine the neurobiology of early life stress, which has demonstrated changes in cortisol/corticosterone expression and activation of a variety of stress- and emotion-relevant brain structures [32–40]. In particular, early handling (consisting of repeated short periods of separation and experimenter handling) differs from maternal separation (characterized by long periods of separation of the dam and pups, typically without explicit experimenter handling) in the direction of the effect on the hypothalamus-pituitary-adrenal (HPA)-axis and cortisol expression, with early handling leading to reduced stress responsiveness and maternal separation leading to elevated stress responsiveness, although all of these effects are highly parameter dependent [37,40,41]. Moreover, many forms of early life pain and stress produce a vulnerable phenotype only when the animal is exposed to a later life insult (a “second-hit”) [28,29,31,42–45]
The NICU involves acute and repeated pain, in addition to non-painful stress, which may cause different or additional effects. For example, a NICU-like “pup in a tea-ball” model with paw pricks produces changes in the HPA-axis responsiveness in females [46] and changes to glutamate/GABA signaling in the pre-frontal cortex and hippocampus [47]. Our lab, along with others, has shown sex-dependent changes in corticotropin releasing factor (CRF) expression in the amygdala due to handling [48], cold stress [26,49], and pin-prick pain [50,51]. While CRF (also called CRH) in the amygdala has been increasingly implicated in pain [52,53], it was first identified as a key mechanism of hypothalamic control of the HPA-axis with significant roles in early life stress and sex-specific changes [54,55]. Overall, the effects of early life adversity in the amygdala, as well as the hypothalamus and HPA-axis, may be of critical importance, as these structures are involved in threat, stress, and pain processing, as changes in forebrain CRF levels during development have been linked with long-term changes in affective and sensory function [21].
Importantly, CRF signaling in the brain could be altered not only by changes in expression of the CRF ligand, as have previously been identified, but also by changes in the CRF receptor. CRF has two major receptors, the high affinity CRFR1 and the lower affinity CRFR2. These receptors are often characterized as having generally opposing functions in stress, fear and pain, with CRFR1 activation leading to enhancement of stress, fear, and pain while CRFR2 activation leads to recovery, diminished stress, fear and responding [56–60], although this simplistic view is likely brain region and context dependent [61]. Changes in the relative expression of CRF receptor 1 and receptor 2 have been linked to chronic pain conditions [56,62] and early life stress [38,63,64], including early life inflammatory pain [65].
Thus, this paper follows up on prior work by examining the effects of an early life pain model, designed to capture key features of the NICU, on CRF receptor mRNA expression. Examining the neurobiological consequences of NICU-like experiences is a critical topic of study. Many sources of early life pain and stress are unavoidable and likely lead to similar neurobiological consequences. However, the NICU represents an environment in which there is both high motivation and ability to enact positive changes to improve later outcomes. Our hypothesis involves a two-hit model, in which we believe that early life events prime the nervous system to respond in an altered manner to a later life stressor or trauma. In our case, the initial trauma is designed to model, albeit incompletely, the NICU experience. This repeated removal from the dam, handling, and skin-breaking paw pricks over the first week of life. Then, later in life, a traumatic event in the form of classical fear conditioning activates a susceptible amygdala and hypothalamus, leading to changes in pain thresholds and affective expression, with both vulnerable and resilience phenotypes emerging, depending on the parameters [31,50,66,67]. In our hands, only rats that experienced both the neonatal pain and the later fear conditioning demonstrate a tactile hypersensitivity showing that both events are needed [31]. Moreover, we have previously found sex-dependent changes in the number of CRF-expressing cells in the amygdala (with an acute increase, followed by a lasting decrease) and to a lesser extent, in the hypothalamus [50,51]. The current experiments examine whether our early life pain model also induces changes in the expression of CRF receptors within the amygdala and hypothalamus in post-weaning rats. Given our prior findings regarding changes in CRF expression in the amygdala, our focus was on that structure. However, given the well-established importance of the hypothalamus to early life stress [26,48,68–70], we included this in our analysis also.
Methods
Subjects
Male and female Sprague Dawley rats were bred in-house using a protocol previously described [31]. We used a total of 89 pups (42m and 47F) from 16 (8 NICU-treated; 8 undisturbed) litters. All litters were housed in (W x D x H) 39.5 cm x 34.6 cm x 21.3 cm closed-environment cages (Tecniplast, West Chester PA). On postnatal day (PD) 1, pups were removed from their mother, placed on a heating pad, sexed, marked via crystal violet stain, and culled to no more than 10 rats per litter (5 males and 5 females when possible). Pups were weaned on PD 21 and lived with their same-sex littermates (approximately 5 per cage). No more than one same sex littermate was assigned to each experimental group, with experimental group defined as a combination of sex (male or female), neonatal condition (early pain, handled, undisturbed), and fear conditioning status (fear conditioned or not). All rats were maintained on a 12:12 light/dark cycle with lights on at 07:00. Food and water were available ad libitum, and at the end of experimentation, rats were euthanized via pentobarbital overdose and brains were collected. All rats were treated in accordance with the NIH Guide for the Care and Use of Laboratory Animals (2011) as well as the 3R principles, and all procedures were approved by the University of New England’s Institutional Animal Care and Use Committee (IACUC) protocol 120324−013. See Fig 1 for a timeline of procedures.
A timeline of procedures.Rats were born on postnatal day (PD) 0, were subjected to neonatal manipulations from PD 1 - 7, weaned on PD 21 and underwent fear conditioning on PD 24 prior to tissue collection.
Neonatal pain
The procedure was similar to that previously described (S. M. Davis et al., 2018, 2021). Briefly, for neonatally manipulated litters, on PDs 1–7, rats were removed from the dam, placed on a heating pad and received either a left hindpaw prick using a 24 G needle (early pain; NICU-treated) or a non-painful tactile touch (handled) every two hours, four times per day starting at 09:00 am. The needle prick procedure was limited to breaking the dermal layer of the hind paw and eliciting a drop of blood. Non-painful touch occurred using the tip of the experimenter’s finger in the same location that was pricked in the early pain-treated subjects. For each of the 4 daily pricking events, subjects were separated from the dam for approximately 5–10 min, with 5 min being the minimum separation. During this period, subjects were repeatedly handled by the experimenter while separated from each other. Pups were reunited shortly prior to being returned to the dam. Subjects were marked with crystal violet periodically (typically on PD 1 and 4) to identify individual pups. Control (undisturbed) litters were left undisturbed during these periods.
Apparatus
Fear conditioning was performed in four Startfear chambers (Harvard Apparatus/Panlab model #58722) with two separate contextual cues that differed in shape (square vs. circle), color (black vs. white walls), and scent (70% ethanol vs. 1% ammonia).
Fear conditioning
On PD 24 fear conditioning rats underwent a fear conditioning protocol, similar to that previously described [31,71]. Rats were placed in their preassigned (counterbalanced) fear conditioning chamber and the program was initiated. The percent of time spent freezing during the first 5 mins was recorded (Habituation). Following the habituation period, a 67-dB tone conditioned stimulus (CS) was presented for 10 sec, immediately followed by a 2-s 1.0-mA foot shock serving as the unconditioned stimulus (UCS). There were 10 tone-shock pairings, separated by an average inter-trial interval (ITI) of 2.5 minutes. Subjects were euthanized approximately 30 minutes after this procedure (~1 hr after onset of conditioning) and tissue was collected.
Tissue collection
Subjects were euthanized with a volume of 0.25 ml pentobarbital (390 mg/ml), prior to intra-cardiac perfusion of saline, followed by 4% paraformaldehyde (PFA) on PD 24. Brains were collected and subsequently post fixed for 24 hours in 4% PFA. All animal euthanasia was consistent with the American Veterinary Medical Association procedures. After the 24-hour post-fix, brains were cryoprotected in 30% sucrose. When cryoprotection was finished, brains were embedded in Tissue-Tek O.C.T. Compound (Sakura Finetek) and flash frozen using liquid nitrogen. Brains were then stored at -80C prior to cryosectioning at 15um onto Superfrost plus microscope slides. Emphasis was placed to identify two sections for staining that closely matched pp. 101–103 (4.7–4.4 mm anterior to the interaural point) of a common developmental rat brain atlas [72] (although we also referenced a more modern atlas [73]) and were free of folds and tears in critical regions Using a thaw-mount method, care was taken to preserve laterality of the sections so that hemisphere could be tracked in future analyses. These sections were stored at −80°C until application of our RNAscope® Fluorescence in situ hybridization protocol (approximately 1 month).
Fluorescence in situ hybridization
All fluorescence in situ hybridization (FISH) was performed using a commercially available system [RNAscope; Advanced Cell Diagnostics (ACD)] and utilized probes targeting Crhr1 (product number: 318911) and Crhr2 (product number: 417851-C2). Our protocol was developed using RNAscope Multiplex Fluorescent Reagent Kit v2 user manual (document number: 323100-USM), manufacturer technical note regarding tissue detachment, manufacturer modifications for fixed frozen tissue (ACD), and our previous work [51]. In the concluding steps of our protocol, DAPI was applied to brain sections as a counterstain for cellular and region identification.
Sections were kept at −80°C until staining. For target retrieval, sections were warmed for 1 hour at room temperature, then dried at 60°C for 45 minutes, prior to a post fix in 4% PFA for 1 hour. They were washed in ascending concentrations of ethanol (50%, 70% and 100%), treated with hydrogen peroxide, washed in DI water, and placed in the target retrieval solution for 15 minutes in a steamer at 99°C, prior to a wash in DI water followed by 100% ethanol. Sections were then dried at 60°C for 30 minutes prior to being left overnight. In situ hybridization began with application of the probes and baking at 40°C for 2 hours. Each channel was then hybridized with the appropriate AMP solution for 30 minutes at 40°C. The appropriate HPR was then added at 40°C for 15 min, prior to the application of the dye. Washing using the wash buffer occurred between each step.
RNAscope assays were conducted in batches. We identified two neighboring sections from each subject that appeared to contain the subject regions. Each RNAscope batch contained one section from each condition (sex x neonatal condition x juvenile treatment) from tissue collected on the same day. Additionally, RNAscope was conducted in “batches” which were duplicated such that the two neighboring sections from the same brain were stained in separate batches to control for any inter-batch variability. Lastly, average values for each subject (for each hemisphere), termed “subject averages”, were created from the two sections and these values were used for all subsequent analysis. Thus, each subject contributed only one value (the subject average) per hemisphere to any statistical analysis. Per manufacturer instructions (RNAscope; ACD) to ensure consistency between subsequent FISH batches, each batch consisted of additional sections processed with either positive control probe [product number: 320891; containing Polr2a (channel 1), PPIB (channel 2)] or negative control probe [product number: 320871; containing DapB gene accession EF191515 from the SMY strain of Bacillus subtilis (channels 1 and 2)].
Fluorescence multiplex imaging was done between 2 days and 2 weeks after FISH. All quantified images and channels were taken under the same magnification (20x) and exposure settings (DAPI – 8ms exposure; FITC channel (crhr1) 300ms exposure; CY3 channel (crhr2) – 35ms exposure) on an Olympus VS200 slide scanner (pixel width and height 0.325um). Images were analyzed using the FIJI package of NIH’s open-source image analysis software ImageJ [74,75]. Image analysis and region of interest (ROI) identification occurred using encoded brain numbers keeping the experimenter blind to the identity of the image being analyzed. Images were first subject to channel separation and the DAPI channel was used to identify regions of quantification (Central Nucleus of the Amygdala; CeA, Basolateral Amygdala; BLA, Paraventricular Nucleus; PVN and Ventromedial Hypothalamus; VMH) (See Fig 2). Due to the lack of detailed developmental rat brain atlases, ROIs were created using a combination of developmental brain atlases to ensure we were in the correct area [72,73] and compared to a common adult rat brain atlas [76].
Example image of RNAscope straining.Panel A: Full brain section straining. Panel B: Closeup on the amygdala. Panel C. Closeup on the hypothalamus. Green = crhr1, Red = crhr2. Blue = DAPI.
The DAPI channel was then dilated and used to create a cell mask for these regions. Due to the different levels of background and positive staining, different thresholding algorithms were used on each channel. FIJI’s Triangle Theshold was used for the green channel and the RenyEntropy Threshold for the red channel. The number and percentage of cells containing crhr1 and crhr2 was assessed, to assess phenotypic changes in cell identity and/or changes in cell survival. The percentage of pixels expressing positive staining within positive cells was also assessed, to examine changes in the degree of expression. Additionally, luminance of straining was assessed in these regions via the averaging of grayscale values, as a control. There were no differences in luminance of staining found, suggesting consistent staining and imaging quality across all groups. All images were subject to the same thresholding, processing, and quantification methods.
Experimental design and analysis
Data were analyzed using a 2 (sex) X 3 (Neonatal condition: early pain, handled, undisturbed) X 2 (juvenile treatment: fear conditioning or not) X 2 (hemisphere – a repeated measure) mixed model General Linear Model (GLM) with IBM SPSS version 27. No more than one same sex littermate was assigned to each experimental group. Individual data points were removed as statistical outliers if they were identified as extreme outliers using SPSS’s outlier analysis in their respective group. No more than 1 data point per group (combination of sex, neonatal condition, juvenile condition and hemisphere) met this criterion with 5 total data points in the BLA, 2 in the CeA, 6 in the PVN and 2 in the VHM excluded across all measures. In addition, sections were removed if the region of interest was not present, was damaged, or was otherwise not quantifiable.
3 measures are reported for each target, the number of cells expressing that target, the percentage of DAPI-stained cells that express the target and the overall percentage of pixels in the positive cells that express the target, reflecting increases in the amount of expression in cells that express the target. Examining differences in the number of cells tests the hypothesis that there is a recruitment or decline in the population of cells that express the CRF receptors, whereas examining the percentage of pixels expressing a target is also sensitive to changes in the degree of expression within the existing cell populations. Data are reported as mean ± SEM and a p-value of ≤ 0.05 was considered statistically significant, while p-values between 0.05 and 0.10 were considered trends towards significance. A Tukey’s post hoc analysis was performed when there were more than two levels of a variable to compare. In addition to graphing the overall data, some statistically significant differences were highlighted with additional Figs.
Results
Basolateral Amygdala
After quality control and outlier analysis, between 82–84 brains contributed to each measure. There was a minimum of 5 and a maximum of 8 brains included per group. Fear conditioning, but not neonatal pain, altered crhr2 expression, only in female subjects. NICU-like treatment did not alter expression of CRH receptors.
Number of nuclei stained with DAPI (Fig 3A): These data were first subjected to a 2 (Sex: M,F) X 3 (Neonatal condition: undisturbed, handled, early pain) X 2 (Juvenile treatment: Fear Conditioned or not) X 2 (hemisphere – a repeated measure) mixed model GLM for the number of nuclei stained with DAPI*.* Surprisingly, we found a significant effect of juvenile treatment F(1,70) = 5.058, p < .05 η^2^ = .07 (Fig 3B) with fear conditioned subjects showing more expression and a significant effect of hemisphere F(1,70) = 32.377, p < .01 η^2^ = .316 (Fig 3C) with the left side showing more expression. Further analysis demonstrated that these differences were attributable to incidental differences in ROI volume, which moderately correlated (rs = .40 −.60 for each group) with the number of DAPI-stained nuclei and showed the same pattern of statistical differences. There were no differences in nuclei density. Given that ROIs were created blind to condition, we believe that these differences were spurious, but should be considered when interpreting later results. No other significant differences were found in this measure. For a list of all analyses, see Table 1.
Table 1: BLA.
DAPI stained nuclei in the Basolateral Amygdala (BLA).PD 1-7 male and female rats were subjected to a painful neonatal experience, non-painful removal from the dam and handling, or left undisturbed except for normal animal colony procedures. Subjects were either fear conditioned or not on PD 24. Tissue was collected and subjected to RNAscope in situ hybridization in both hemisphere as shown in Panel A. There were significantly more DAPI-stained nuclei in fear conditioned subjects (Panel B) and in the left hemisphere (Panel C). These differences were attributable to incidental differences in ROI size. Representative images from Undisturbed (Panel D), Handled (Panel E), and Pain (Panel F) fear conditioned subjects are also shown (brightness altered for enhanced visibility).
Number of cells expressing crhr1: These data were first subjected to a 2 (Sex: M,F) X 3 (Neonatal condition: undisturbed, handled, early pain) X 2 (Juvenile treatment: Fear Conditioned or not) X 2 (hemisphere) mixed model GLM for the number of cells expressing crhr1. There were no significant main effects or interactions with sex, so we combined across sex (Fig 4A). We observed a significant main effect of hemisphere F(1,72) = 16.73, p < .01 η^2^ = .189, and a trend towards a significant hemisphere X juvenile treatment interaction F(1,72) = 3.62, p < .07 η^2^ = .045. A follow-up analysis demonstrated a significant effect of hemisphere only in the fear conditioned subjects F (1,38) = 18.101, p < .01 η^2^ = .323 and not in control subjects F (1,34) = 2.458, n.s. η^2^ = .067 with the left hemisphere demonstrating significantly more expression (Fig 4B).
Crhr1 expression in the BLA.PD 1-7 male and female rats were subjected to a painful neonatal experience, non-painful removal from the dam and handling, or left undisturbed except for normal animal colony procedures. Subjects were either fear conditioned or not on PD 24. Tissue was collected and subjected to RNAscope in situ hybridization in both hemisphere. Panel A: All data from both hemispheres, collapsed across sex due to lack of statistical differences. Panel B: Data collapsed across all variables except juvenile treatment and hemisphere, to highlight the interaction. *** = statistically significant p < .001. Panel C: the percent of DAPI stained nuclei that express crhr1 collapsed across sex due to lack of statistical differences. Panel D: the percent of pixels that express crfr1 collapsed across sex due to lack of statistical differences.
A 2 (Sex: M,F) X 3 (Neonatal condition: undisturbed, handled, early pain) X 2 (Juvenile treatment: Fear Conditioned or not) X 2 (hemisphere) mixed model MANOVA examining percent of cells expressing crhr1 revealed no significant differences (Fig 4C), as did a similar analysis on percent of pixels in positive cells (Fig 4D). As the number, but not the percent of cells or amount of expression differ, the hemispheric difference in the number of cells expressing crfr1 is likely an artifact reflecting differences in the number of DAPI nuclei. All statistics can be found in Table 1.
Number of Cells expressing crhr2: These data were also first subjected to a 2 (Sex: M,F) X 3 (Neonatal condition: undisturbed, handled, early pain) X 2 (Juvenile treatment: Fear Conditioned or not) X 2 (hemisphere) mixed model GLM for the number of cells expressing crhr2 in the BLA (Fig 5A). We observed a significant main effect of hemisphere F(1,70) = 4.567, p < .05 η^2^ = .061, with the right hemisphere demonstrating higher levels of expression and a significant sex X juvenile treatment interaction F(1,70) = 4.06, p < .05 η^2^ = .055, which demonstrated significantly more crhr2 expression in the right hemisphere of fear conditioned subjects only in females, with a moderate effect size (Figs 5B and 5C). Importantly, this cannot be accounted for by the greater number of DAPI-labeled cells in the left hemisphere and suggests that fear conditioning alters crhr2 expression preferentially in females.
Crhr2 expression in the BLA.PD 1-7 male and female rats were subjected to a painful neonatal experience, non-painful removal from the dam and handling, or left undisturbed except for normal animal colony maintenance. Subjects were either fear conditioned or not on PD 24. Tissue was collected and subjected to RNAscope in situ hybridization in both hemispheres. Panel A: The number of cells expressing crhr2 from both hemispheres, collapsed across sex. Panel B: The number of cells expressing crhr2 from male subjects highlighting the lack of effect of fear conditioning. Panel C: The number of cells expressing crhr2 from female subjects highlighting the effect of fear conditioning and hemisphere. Panel D: Percentage of cells expressing crhr2 collapsed across sex. Panel E: Percentage of cells expressing crhr2 from male subjects highlighting the lack of effect of fear conditioning and hemisphere. Panel F: The percentage of cells expressing crhr2 from female subjects highlighting the effect of fear conditioning and hemisphere. Panel G: The percentage of overall ROI expressing positive signal after thresholding, collapsed across sex. * = statistically significant p < .05, ** = statistically significant p < .01.
Percent of DAPI cells that express crhr2: These data were also first subjected to a 2 (Sex: M,F) X 3 (Neonatal condition: undisturbed, handled, early pain) X 2 (Juvenile treatment: Fear Conditioned or not) X 2 (hemisphere) mixed model GLM for the number of cells expressing crhr2 in the BLA (Fig 5D). We again observed a significant main effect of hemisphere F(1,69) = 17.123, p < .01) η^2^ = .199, with the right hemisphere demonstrating higher levels of expression (and a large effect size) as well as a significant sex X juvenile treatment interaction F(1,69) = 4.860, p < .05 η^2^ = .066, which demonstrated significantly more crhr2 expression in the right hemisphere of fear conditioned subjects only in females (Figs 5E and 5F). Because this is a percentage and opposite to the hemisphere with more DAPI, this difference also cannot be accounted for by any difference in the number of cells examined and again confirms that fear conditioning alters crhr2 expression preferentially in females.
A 2 (Sex: M,F) X 3 (Neonatal condition: undisturbed, handled, early pain) X 2 (Juvenile treatment: Fear Conditioned or not) X 2 (hemisphere) mixed model GLM for the percent of pixels expressing crhr2 in the BLA found no significant differences (Fig 5G). All statistics can be found in Table 1.
Central nucleus of the amygdala
A total of 83–87 brains survived quality control and outlier analysis for the CeA, for each measure. There was a minimum of 6 and a maximum of 8 subjects per group. Once again, fear conditioning altered crhr2 expression in female subjects. There was a trend towards an effect of early life pain on crhr2 expression in males.
Number of nuclei stained with DAPI (Fig 6A). These data were again subjected to a 2 (Sex: M,F) X 3 (Neonatal condition: undisturbed, handled, early pain) X 2 (Juvenile treatment: Fear Conditioned or not) X 2 (hemisphere) mixed model GLM for the number of stained nuclei*.* We once again found an unexpected significant effect of juvenile treatment (Fig 6B) F(1,75) = 8.395, p < .01 that correlated with differences in ROI area and reflected more cells in fear conditioned subjects. This is also likely to be spurious, but subsequent analyses will be considered in light of this difference.
DAPI stained nuclei in the Central Nucleus of the Amygdala (CeA).PD 1-7 male and female rats were subjected to a painful neonatal experience, non-painful removal from the dam and handling, or left undisturbed except for normal animal colony procedures. Subjects were either fear conditioned or not on PD 24. Tissue was collected and subjected to RNAscope in situ hybridization in both hemisphere as shown in Panel A. There were significantly more DAPI-stained nuclei in fear conditioned subjects (Panel B). These differences were attributable to incidental differences in ROI size. Representative images from Undisturbed (Panel C), Handled (Panel D), and Pain (Panel E) subjects are also shown (brightness altered for enhanced visibility).
Number of cells expressing crhr1. Similar to above, these data were first subjected to a 2 (Sex: M,F) X 3 (Neonatal condition: undisturbed, handled, early pain) X 2 (Juvenile treatment: Fear Conditioned or not) X 2 (hemisphere) mixed model GLM for the number of cells expressing crhr1. With no sex differences, data are collapsed across this variable (Fig 7A). The initial analysis found a trend toward a significant hemisphere X juvenile treatment interaction F(1, 74) = 3.83, p = .054 η^2^ = .049 (Fig 7B). A follow up analysis showed a trend towards greater expression in left hemisphere of fear conditioned subjects compared to controls (p = .065; η^2^ = .045). This can be accounted for by the greater number of DAPI-stained cells in fear conditioned subjects and likely does not reflect an effect of conditioning. A 2 (Sex: M,F) X 3 (Neonatal condition: undisturbed, handled, early pain) X 2 (Juvenile treatment: Fear Conditioned or not) X 2 (hemisphere) mixed model MANOVA examining percent of cells expressing crhr1 revealed no significant differences (Fig 7C), as did a similar analysis on percent of pixels in positive cells (Fig 7D). Thus, similar to the BLA, the difference in the number of cells expressing crfr1 is likely an artifact reflecting differences in ROI area altering the number of DAPI nuclei. All statistics can be found in Table 2.
Table 2: CeA.
Crhr1 expression in the CeA.PD 1-7 male and female rats were subjected to a painful experience, non-painful removal from the dam and handling, or left undisturbed except for normal animal colony maintenance. Subjects were either fear conditioned or not on PD 24. Tissue was collected and subjected to RNAscope in situ hybridization in both hemispheres. Panel A: Number of cells expressing crhr1 from both hemispheres, collapsed across sex. Panel B: Number of cells expressing crhr1 collapsed across neonatal condition and sex, to highlight the hemisphere by juvenile condition statistical interaction. Panel C: Percentage of cells in overall ROI expressing crhr1, collapsed across sex. Panel D: Percentage of pixels expressing positive signal after thresholding, collapsed across sex. There were no significant differences between groups.
Number of cells expressing crhr2: The 2 (Sex: M,F) X 3 (Neonatal condition: undisturbed, handled, early pain) X 2 (Juvenile treatment: Fear Conditioned or not) X 2 (hemisphere) mixed model GLM (Fig 8A) yielded a significant main effect of hemisphere F(1,71) = 5.325, p < .05 η^2^ = .070 as well as an interaction between hemisphere and juvenile treatment F (1,71) = 4.36, p < .05 η^2^ = .058, with a significant increase in crfr2 expression on the right side only in control subjects (p < .01; η^2^ = .226) (Fig 8B). There were no other significant statistical differences (see Table 2).
Crhr2 expression in the CeA.PD 1-7 male and female rats were subjected to a painful experience, non-painful maternal separation and handling, or left undisturbed except for normal animal colony maintenance. Subjects were either fear conditioned or not on PD 24. Tissue was collected and subjected to RNAscope in situ hybridization in both hemispheres. Panel A: Number of cells expressing crhr2 from both hemispheres, collapsed across sex. Panel B: Number of cells expressing crhr2 collapsed across neonatal condition and sex, to highlight the hemisphere by juvenile condition statistical interaction. Panel C: Percentage of cells in overall ROI expressing crhr1 in males. Panel D: Percentage of cells in overall ROI expressing crhr2 in females. Panel E: Percentage of cells in overall ROI expressing crhr1 in females collapsed across neonatal condition highlighting the hemisphere X juvenile condition interaction. Panel F: Percentage of pixels expressing positive signal after thresholding in males. Panel G: Percentage of pixels expressing positive signal after thresholding in females. Panel H: Percentage of pixels expressing positive signal after thresholding in males collapsed across juvenile condition highlighting the trend towards an effect of sex. * = significant p < .05, ** = p < .01.
The GLM examining the percentage of DAPI-stained cells expressing crhr2 also revealed a significant main effect of hemisphere F(1,73) = 6.59, p < .05 η^2^ = .083, a significant hemisphere X juvenile treatment interaction F(1,73) = 4.13, p < .05 η^2^ = .054 and a significant 4-way interaction among hemisphere X sex X neonatal condition X juvenile treatment F (2, 73) = 4.64, p < .05 η^2^ = .113 (Figs 8C – Males & 8D – Females). Follow up analysis found a trend towards a significant effect of juvenile treatment only on the left side of females (Fig 8E) F(1,38) = 3.155, p = .084 η^2^ = .077, suggesting that the statistical interaction is likely the result of minor hemispheric differences.
The GLM examining the percentage of pixels in positive cells expressing crhr2 found a significant 4-way interaction among hemisphere X juvenile condition X neonatal condition X sex F(2,74) = 4.455, p < .05 η^2^ = .107 (Fig 8F – Males and 8G – Females). Subsequent analyses demonstrate a significant hemisphere X juvenile condition X neonatal condition only in females F (2,38) = 4.978, p < .05 η^2^ = .208 driven by a significant effect of hemisphere only in the neonatal handled, non-fear conditioned subjects F(1,5) = 8.064, p < .05, η^2^ = .617. No other follow-up analyses found significant effects. There was also a trend towards a significant effect of Neonatal Condition only in left hemisphere of males F(2,36) = 2.628, p = .086, η^2^ = .127 (Fig 8H). This moderate-large effect on percent of pixels that express the target likely reflects changes in transcription within existing populations following early life pain.
Paraventricular nucleus of the hypothalamus
A total of 66–68 brains had PVN regions on the slices, survived quality control and outlier analysis for the PVN for each measure. There was a minimum of 4 and a maximum of 8 subjects per group. Early life pain, only in fear conditioned subjects, reduced crhr1 expression. Fear conditioning and early life pain and stress also altered crhr2 expression.
Number of nuclei stained with DAPI (Fig 9). These data were again subjected to a 2 (Sex: M,F) X 3 (Neonatal condition undisturbed, handled, early pain) X 2 (Juvenile treatment: Fear Conditioned or not) X 2 (hemisphere) mixed model GLM for the number of stained nuclei*.* There were no significant or trending main effects or interactions (see Table 3).
Table 3: PVN.
DAPI stained nuclei in the Paraventricular Nucleus of the Hypothalamus (PVN).PD 1-7 male and female rats were subjected to a painful neonatal experience, non-painful removal from dam and handling, or left undisturbed except for normal animal colony procedures. Subjects were either fear conditioned or not on PD 24. Tissue was collected and subjected to RNAscope in situ hybridization in both hemispheres. There were no effects of sex, neonatal treatment, nor juvenile treatment on the number of DAPI cells (Panel A). Representative images from Undisturbed (Panel B), Handled (Panel C), and Pain (Panel D) subjects are also shown (brightness altered for enhanced visibility).
Number of Cells expressing crhr1: As in the amygdala, these data were first subjected to a 2 (Sex: M,F) X 3 (Neonatal condition: undisturbed, handled, early pain) X 2 (Juvenile treatment: Fear Conditioned or not) X 2 (hemisphere) mixed model GLM for the number of cells expressing crhr1 (Fig 10A). There were no significant main effects or interactions (Table 3).
Crhr1 expression in the PVN.PD 1-7 male and female rats were subjected to a painful experience, non-painful maternal separation and handling, or left undisturbed except for normal animal colony maintenance. Subjects were either fear conditioned or not on PD 24. Tissue was collected and subjected to RNAscope in situ hybridization in both hemispheres. Panel A: Number of cells expressing crhr1 from both hemispheres combined across sex. There were no significant main effects or interactions. Panel B: Percent of cells expressing crhr1 from both hemispheres, in males. * = significant differences in labeled groups, p < .05. Panel C: Percent of of cells expressing crhr1 from both hemispheres, in females. There were no significant main effects or interactions. Panel D: Percentage of pixels expressing crhr1 in positive cells in male subjects. # = significant sex differences in marked groups, p < .05. Panel E: Percentage of pixels expressing crhr1 in positive cells in female subjects. # = significant sex differences in marked groups, p < .05.
When examining the percent of DAPI stained cells that express crhr1, we find a significant 2-way interaction between neonatal condition and juvenile condition F(2,55) = 3.704, p < .05 η^2^ = .119 and a significant 3-way interaction among sex, neonatal condition and juvenile condition F(2,55) = 3.478, p < .05, η^2^ = .112 (Fig 10B – Males; 10C – Females). Follow up analyses find a nearly significant hemisphere by neonatal condition interaction F(2,15) = 3.690, p = .05, η^2^ = .330 only in the male rats that were not fear conditioned. This was explained by a significant difference between the handled and undisturbed subjects only in the right hemisphere (Tukey’s p < .05) and a significant effect of neonatal condition only in male rats that were fear conditioned F(2,13) = 4.176, p < .05, η^2^ = .391. Tukey’s post-hoc tests confirmed that only the undisturbed and early pain groups significantly differed (p < .05).
An analysis examining percent of pixels expressing crhr1 (Fig 10D – Males; 10E - Females) demonstrated a significant 3-way interaction of sex X neonatal condition X juvenile condition F(2,55) = 3.768, p < .05, η^2^ = .121. Further analyses suggest this is driven by an significant effect of sex F(1,8) = 6.140, p < .05, η^2^ = .434 only in rats that received neonatal pain and juvenile fear conditioning as well as a trend towards a significant effect of sex F(1,9) = 4.296, p = .068, η^2^ = .323 in undisturbed rats that were not fear conditioned. In both cases, females had greater expression.
Number of Cells expressing crhr2 (Fig 11A): The 4-way analysis found a trend towards significant effect of hemisphere F(1,53) = 3.026, p = .088, η^2^ = .054 with greater expression on the right side (see Table 3). In addition, there was a trend towards a significant effect of neonatal pain condition F(2,53) = 3.000, p = .058, η^2^ = .102 (Fig 11B). Tukey’s post-hoc tests confirm that only the handled and undisturbed subjects differed (p < .05).
crhr2 expression in the PVN.PD 1–7 male and female rats were subjected to a painful experience, non-painful maternal separation and handling, or left undisturbed except for normal animal colony maintenance. Subjects were either fear conditioned or not on PD 24. Tissue was collected and subjected to RNAscope in situ hybridization in both hemispheres. Panel A: Number of cells expressing crhr2 from both hemispheres, collapsed across sex. Panel B: Number of cells expressing crhr2, collapsed across sex and juvenile condition to highlight the effect neonatal condition. * = significant differences in labeled groups, p < .05. Panel C: Percentage of cells in overall ROI expressing crhr2 collapsed across sex. There were no significant main effects or interactions. Panel D: Percentage of pixels in positive cells expressing positive signal in males. Panel E: Percentage of pixels in positive cells expressing positive signal in females. * = significant differences in labeled groups, p < .05. Panel F: Percentage of pixels expressing positive signal collapsed across sex and juvenile condition highlighting the effects of neonatal pain.
Examining the percent of DAPI-stained cells expressing crhr2 (Fig 11C), we found a significant 3-way interaction of hemisphere, neonatal condition and juvenile condition F(2,52) = 4.143, p < .05 η^2^ = .137, but no other significant effects or interactions. This appears to be driven by a significant interaction of hemisphere and juvenile condition only in handled subjects F(1,18) = 5.720, p < .05 η^2^ = .241, although further analyses found no specific significant differences between groups.
Examining the percent of pixels expressing crhr2 (Fig 11D - males and 11E - females) finds a significant interaction between hemisphere and neonatal condition F(2,54) = 6.093, p < .01 η^2^ = .208, and a 4-way interaction among hemisphere, sex, neonatal and juvenile condition F(2,54) = 3.622, p < .05, η^2^ = .118. Further analyses demonstrate trends toward significant differences between pain and undisturbed subjects on the left (p = .062) and between pain and handled on the right (p = .059). In addition, there was a significant effect of fear conditioning only in female, undisturbed subjects on the right side F(1,12) = 6.598, p < .05, η^2^ = .355. No other differences were significant.
Ventromedial hypothalamus
A total of 75–78 brains survived quality control and outlier analysis for the VMH for each measure. There was a minimum of 5 and a maximum of 8 subjects per group. Detailed statistics can be found in Table 4. There was a trending effect of neonatal pain on crhr2, but there were no other meaningful effects of early life pain or adolescent fear conditioning.
Table 4: VMH.
Number of nuclei stained with DAPI (Fig 12A). These data were again subjected to a 2 (Sex: M,F) X 3 (Neonatal treatment: undisturbed, handled, early pain) X 2 (Juvenile treatment: Fear Conditioned or not) X 2 (hemisphere) mixed model GLM for the number of stained nuclei*.* Results can be found in Table 4. We found a significant 2-way interaction between sex and juvenile condition F(1,63) =5.156, p < .05, η^2^ = .076, as well as a significant 3-way interaction among sex, neonatal condition and juvenile condition F(2,63) = 4.929, p = .01, η^2^ = .135. This appears to be driven by a significant effect of fear conditioning only in the female neonatal pain subjects F(1,9) = 5.460, p < .05, η^2^ = .378, as no other follow-up analyses found significant differences.
DAPI stained nuclei in the Ventromedial Hypothalamus (VMH).PD 1-7 male and female rats were subjected to a painful neonatal experience, non-painful maternal separation and handling, or left undisturbed except for normal animal colony procedures. Subjects were either fear conditioned or not on PD 24. Tissue was collected and subjected to RNAscope in situ hybridization in both hemispheres. Panel A: the number of DAPI strained nuclei in male rats. There were no significant effects or interactions. Panel B: the number of DAPI strained nuclei in females. Representative images from Undisturbed (Panel C), Handled (Panel D), and Pain (Panel E) subjects are also shown (brightness altered for enhanced visibility). = significant differences in labeled groups, p < .05.*
Number of cells expressing crhr1: A 2 (Sex: M, F) X 3 (Neonatal condition: undisturbed, handled, early pain) X 2 (Juvenile treatment: Fear Conditioned or not) X 2 (hemisphere) mixed model GLMs for the number of cells (Fig 13A) yielded no significant effects or interactions.
Crhr1 expression in the VMH.PD 1-7 male and female rats were subjected to a painful experience, non-painful maternal separation and handling, or left undisturbed except for normal animal colony maintenance. Subjects were either fear conditioned or not on PD 24. Tissue was collected and subjected to RNAscope in situ hybridization in both hemispheres. Panel A: Number of cells expressing crhr1 from both hemispheres collapsed across sex. There were no significant effects or interactions = . Panel B: Percentage of DAPI-labeled cells expressing crhr1 from both hemispheres in males. There were no significant effects or interactions. Panel C: Percentage of DAPI-labeled cells expressing crhr1 from both hemispheres in females. * = significant differences in labeled groups, p < .05. Panel D: Percentage of pixels in positive cells expressing crhr1 in males. Panel E: Percentage of pixels in positive cells expressing crhr1 in females. There was a significant effect of sex on the percent of pixels labeled in the right hemisphere (p < .05).
For the percentage of cells expressing crhr1 (Fig 13B and 13C), the ANOVA yielded a significant hemisphere X sex X neonatal condition interaction F(1,66) = 3.437, p < .05 η^2^ = .094. Follow-up analysis found significant effects of hemisphere only in female fear-conditioned subjects that were neonatally undisturbed or exposed to early life pain (ps < .05). No other groups showed significant effects or interactions.
For the percentage of pixels in positive cells expressing crhr1(Fig 13D), the ANOVA yielded a significant hemisphere X sex X juvenile condition interaction F(1,66) = 5.173, p < .05 η^2^ = .073. There was also a trend towards a significant sex X neonatal condition X juvenile condition interaction F(2,66) = 2.684, p = .076, η^2^ = .075. Follow up analyses found a significant effect of sex only in the right hemisphere F(1,64) = 4.244, p < .05, η^2^ = .047, η^2^ = .062. No other significant differences were found.
For the number of cells expressing CRFR2, the GLM yielded no significant main effects or interactions (Fig 14A). Analyses on the percent of DAPI-labeled cells (Fig 14B) expressing crhr2 also yielded no significant main effects or interactions, although there was a trend towards a significant effect of sex F(1,66) = 3.265, p = .075, η^2^ = .047 with males having more expression.
Crhr2 expression in the VMH.PD 1-7 male and female rats were subjected to a painful experience, non-painful removal from dam and handling, or left undisturbed except for normal animal colony maintenance. Subjects were either fear conditioned or not on PD 24. Tissue was collected and subjected to RNAscope in situ hybridization in both hemispheres. Panel A: Number of cells expressing crhr2 from both hemispheres, collapsed across sex. There were no significant main effects or interactions. Panel B: Percentage of cells expressing crhr2 collapsed across sex. There were no significant main effects or interactions. Panel C: Percentage of pixels in positive cells expressing positive signal in males. Panel D: Percentage of pixels in positive cells expressing positive signal in females.
The analysis on percent of pixels expressing crhr2 (Fig 14C – males and 14D – females) yielding a significant main effect of hemisphere F(1,64) = 10.966, p < .01 η^2^ = .146 with more expression on the left. a trend towards an interaction of hemisphere and sex F(1,64) = 3.409, p = .069, η^2^ = .051 and a trend towards a 4-way interaction F(2,64) = 2.690, p = .076, η^2^ = .078. Follow up analyses on the left hemisphere found a significant sex X neonatal condition interaction F(2,66) = 3.657, p < .05 η^2^ = .100. This appears to be driven by a trend towards an effect of neonatal condition only in male rats F(2,32) = 2.964, p = .066, η^2^ = .156, with no significant pairwise differences, but a trend towards a difference in undisturbed and pain rats (p = .073). On the right side, there was a significant effect of sex F(1,64) = 4.244, p < .05, η^2^ = .062, with males having more expression.
Discussion
This paper examines the effects of both neonatal pain and juvenile fear conditioning on the expression of CRF receptor mRNA in critical regions of the amygdala and hypothalamus. The CeA has been heavily implicated in adult pain studies [53,77–79], our prior work has demonstrated effects of our early life pain model on expression of CRF mRNA in the CeA of only male (but not female) rats [50,51]. The BLA is a major input to the CeA and a critical site of plasticity in fear conditioning [80–83]. In contrast, the hypothalamus and HPA-axis have been heavily implicated in early life stress, including maternal separation and handling stress [26,36,38,48,65]. Thus, based on these findings and the literature, we anticipated that the CeA, BLA and PVN would show significant changes in receptor expression. The VMH was largely a region of convenience and serves as a control region showing the specificity of our effects in the critical regions. While we did observe neonatal pain and stress-induced changes in crhr1 expression in the PVN of males, effects in other regions were small and did not reach statistical significance. Of note, because our section selection prioritized the amygdala, we likely captured the primarily the posterior PVN. Effects in the anterior region of the PVN may be different. The lack of effect in the amygdala is particularly surprising given prior data that a single inflammatory injection of carrageenan within a day of birth was found to significantly reduce CRFR1 expression in the BLA (among other changes) [65]. Importantly, the overall direction of change is the same. We have previously shown that carrageenan injections do not produce the same effects as our repeated paw prick model [31,67], likely due to differences in duration and intensity of the pain, as well as the concomitant inflammation that accompanies carrageenan. This may account for some of the difference in outcomes, as might the fact that juveniles were assessed in the current study whereas [65] assessed adults. In our model, the lack of effect of early life pain on CRF receptor expression suggests that the effects of early life pain are largely limited to changes in CRF expression rather than changes in its receptors.
CRF receptors are G-protein coupled receptors that play differential roles in fear, anxiety and pain [56,84]. [31,65,67] CRF receptors 1 and 2 may play opposite, or at least differential, roles in emotional outcome, such as anxiety, fear, pain, and threat responding [84–87]. In addition to the effects on CRF receptor 1 in the PVN, we also observed trends towards increases in CRF receptor 2 expression following early life pain in the PVN, VMH, and CeA. That the effect in the CeA was limited to males is consistent with our prior work, demonstrating effects on CRF-expressing cells in our model in the CeA only in male subjects [50,51], thus male-only effects on the CeA may be expected. Regarding the specificity of the effect to CRF receptor 2, there are known differences in CRF receptor function. CRF receptor 1 activation in the amygdala tends to escalate active forms of anxiety, as well as pain. In contrast, CRF receptor 2 appears to be involved in pain and stress resolution, or passive forms of anxiety and threat responding. Increases in crhr2 is typically associated with reductions in anxiety [86]. In our hands, this was an increase in expression as measured by the percentage of pixels expressing signal in positive cells, but not in increase in the number of cells showing expression. This suggests an upregulation in crfr2 expression in the population of cells already expressing the receptor. Given that our prior work has demonstrated an anxiolytic effect of our model – decreasing conditioned freezing and increased time in open arms on the elevated plus maze [31,51], these changes in receptor expression appear to be consistent with the observed changes in behavior. Similar results have also been associated with chronic pain conditions [79]. Thus, when compared with our prior work and the literature, it appears that the NICU-like experience produces and lasting changes in CRF-expressing cells [50], as well as changes in CRF-receptor 2 expression. Of course, the current data are not informative regarding any acute effects of the neonatal trauma or pain on CRF receptor 2 expression.
We also saw an increase in crfr2 expression in the BLA following fear conditioning, albeit only in females. Given the known role for this region in fear conditioning, the effect on fear conditioning is not surprising. Previous work has also shown changes in CRF receptor signaling and function, including sex differences, during fear conditioning [84,88–93], although studies on crfr2 are less consistent [94], and this work should be interpreted as broadly consistent with those findings.
The current data also demonstrate differences in CRF receptor expression in both the amygdala and hypothalamus as a function of hemisphere. While some of this was attributable to differences in ROI creation, there were several meaningful differences that this does not account for. For example, there were increases in CRF receptor 2 expression only in the right hemisphere of the BLA and CeA following early life pain and fear conditioning. This type of lateralization is likely reflective of our growing understanding that affective function is lateralized. In both pain and emotion processing, the left and right amygdala appear to have different functions with the right side being especially important for pain and fear responding [80,95–100]. Lateralization was less profound in the hypothalamus, although we did observe changes in crhr1 expression only in the right PVN of males. While hemispheric specialization is less well understood in the hypothalamus, there is emerging evidence of lateralization there as well [101].
We also observed significant sex differences in the effects of pain and fear conditioning on CRF receptor expression. This was most notable with greater crhr1 in the male PVN. We also saw trends towards increased crhr2 in the male VMH, which is consistent with the known androgen-mediation of crhr2 expression in the hypothalamus [102]. We also observed female-specific changes in crhr2 expression especially in the BLA following fear conditioning. The small effects of pain on the CeA were limited to males – consistent with our prior work [50,51]. Indeed, there are robust sex differences published in CRF signaling [103,104], stress [105–107], pain responding [108,109], HPA-axis activation [46], and the effects of early life stress and pain [108,110–115]. These effects have been associated with sex differences in CRF, adrenocorticotropin hormone (ACTH) and cortisol/corticosterone (CORT) expression. Thus, we anticipated sex differences in CRF receptor expression also.
There are some important limitations to this study. While we observed fear conditioning-induced differences in CRF receptor expression in the amygdala, suggesting that our timepoint (1 hour after onset of conditioning) was sensitive to this manipulation, many other studies have suggested that peak stress-induced expression is 1.5–4 hours after the stressor [116–118], although others have found increases earlier [119]. Thus, it’s possible that this study underestimates the effects of the fear conditioning manipulation on receptor expression. Of course, this limitation does not apply to any lasting changes that could have been induced by the neonatal manipulation, which was conducted weeks earlier. Second, RNAscope is designed to produce discrete puncta, with each dot representing a single transcript. Our imaging yielded a clear signal, but the degree of overlap and larger “blobs” did not leave us confident in our ability to assess individual puncta and complicated quantifying the degree of expression using individual dots as measures. This is common in studies using similar targets [120–122]. We dealt with this in two ways. First, we simply scored cells as having expression above threshold, or not. This does not depend on the resolution of discrete puncta. Second, we examined the percentage of pixels that expressed signal in positive cells. This allows us to assess the degree to which clustering of multiple puncta occurred and likely reflects the degree of expression within cells already expressing the transcript. Our use of consistent imaging and thresholding parameters was designed to offset any potential confounds in this interpretation. Finally, the assessment of changes in mRNA expression may, or may not, reflect changes in protein expression levels.
Overall, when combined with the literature, these data are consistent with the hypothesis that early life pain and juvenile stress alter CRF signaling in the amygdala and hypothalamus. While there are lasting effects of our NICU-model on CRF receptor expression, these appeared limited to CRF receptor 2 and predominately in the PVN. Our prior work demonstrating changes in expression of the CRF peptide itself, including the number of cells expressing CRF and their amount of expression, suggests that changes to the CRF peptide are likely more important than long term changes in receptor distribution and expression, at least within in the amygdala [50,51]. The current paper demonstrates, similar to our prior work [50,51], that our “handled” group produces neurobiological changes that are less severe than the pain condition. The current data also demonstrate that CRF receptor expression is affected by juvenile fear conditioning in the amygdala, especially in females. Thus, we believe that early life pain and stress during a critical developmental period in the perinatal window alters the development of CRF signaling creating an underlying and enduring vulnerability which alters later life responses to stress. It is likely the combination of these effects (early life trauma creating “organizational” changes and later life trauma “activating” those altered circuits) that likely creates the unique outcomes from both “hits” resulting in the tactile hypersensitivity we have observed in our previous work.
Conclusions
Neonatal pain, in our hands, only subtly altered CRF receptor expression in a sex-dependent manner. We observed significant effects on early life pain and stress on CRF receptor 1 in the PVN of male rats and non-significant, but trending, effects on CRF receptor 2 mRNA in the hypothalamus and amygdala. Fear conditioning primarily altered CRF receptor 2 in the amygdala of females. This has implications on the mechanisms by which early life trauma leads to a lasting vulnerability to a later life stressor or trauma.
Supporting information
S1 FileBurman and Zuke “Data for submission_CRFReceptor_RNAscope.xlsx”: This file contains our raw data prior to analysis.(XLSX)
S2 FileBurman and Zuke Macros.zip.This file contains the FIJI macros used in our analyses.(ZIP)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cong X, Wu J, Vittner D, Xu W, Hussain N, Galvin S, et al. The impact of cumulative pain/stress on neurobehavioral development of preterm infants in the NICU. Early Hum Dev. 2017;108:9–16. doi: 10.1016/j.earlhumdev.2017.03.003 28343092 PMC 5444300 · doi ↗ · pubmed ↗
- 2Grunau RE, Holsti L, Peters JWB. Long-term consequences of pain in human neonates. Semin Fetal Neonatal Med. 2006;11(4):268–75. doi: 10.1016/j.siny.2006.02.007 16632415 · doi ↗ · pubmed ↗
- 3Grunau RE, Weinberg J, Whitfield MF. Neonatal procedural pain and preterm infant cortisol response to novelty at 8 months. Pediatrics. 2004;114(1):e 77-84. doi: 10.1542/peds.114.1.e 77 15231977 PMC 1351380 · doi ↗ · pubmed ↗
- 4Porter FL, Grunau RE, Anand KJ. Long-term effects of pain in infants. J Dev Behav Pediatr. 1999;20(4):253–61. doi: 10.1097/00004703-199908000-00008 10475600 · doi ↗ · pubmed ↗
- 5Grunau RE, Whitfield MF, Fay T, Holsti L, Oberlander T, Rogers ML. Biobehavioural reactivity to pain in preterm infants: a marker of neuromotor development. Dev Med Child Neurol. 2006;48(6):471–6. doi: 10.1017/S 0012162206001010 16700939 · doi ↗ · pubmed ↗
- 6Brummelte S, Chau CMY, Cepeda IL, Degenhardt A, Weinberg J, Synnes AR, et al. Cortisol levels in former preterm children at school age are predicted by neonatal procedural pain-related stress. Psychoneuroendocrinology. 2015;51:151–63. doi: 10.1016/j.psyneuen.2014.09.018 25313535 PMC 4268136 · doi ↗ · pubmed ↗
- 7Doesburg SM, Chau CM, Cheung TPL, Moiseev A, Ribary U, Herdman AT, et al. Neonatal pain-related stress, functional cortical activity and visual-perceptual abilities in school-age children born at extremely low gestational age. Pain. 2013;154(10):1946–52. doi: 10.1016/j.pain.2013.04.009 23711638 PMC 3778166 · doi ↗ · pubmed ↗
- 8Chaloner A, Greenwood-Van Meerveld B. Early life adversity as a risk factor for visceral pain in later life: importance of sex differences. Front Neurosci. 2013;7:13. doi: 10.3389/fnins.2013.00013 23407595 PMC 3570767 · doi ↗ · pubmed ↗