Regulation of proteolysis of the sigma factor RpoS by the Gac-Rsm signal transduction system in Azotobacter vinelandii
Juliana Berenice Rojo-Rodríguez, Soledad Moreno, Guadalupe Espin

TL;DR
This paper explains how a signaling system in Azotobacter vinelandii controls the stability of a key protein, RpoS, through a complex regulatory network.
Contribution
The study reveals a novel regulatory mechanism linking the Gac-Rsm system to RpoS proteolysis via RsmA and ClpXP.
Findings
GacA activates RsmZ1 and RsmZ2, which counteract RsmA's repression of target mRNAs.
RsmA acts as a positive regulator of ClpXP expression, affecting RpoS stability.
Inactivation of clpP or clpX in a gacA mutant restores RpoS stability.
Abstract
In Azotobacter vinelandii, the sigma factor RpoS is maintained at low levels in exponentially growing cells due to degradation mediated by the chaperone–protease complex ClpXP, while high levels are observed in the stationary phase. This study showed that degradation of RpoS by ClpXP is under the control of the Gac-Rsm signal transduction system, in which GacA, the transcriptional activator of the two-component system GacS/GacA, activates transcription of the small RNAs RsmZ1 and RsmZ2. These RNAs bind to the translational repressor protein RsmA to counteract its repressor activity on its target mRNAs. We found that in stationary-phase cells, compared with the WT, a gacA mutant exhibited lower RpoS levels due to reduced stability, while levels of the clpP and clpX mRNAs were higher. Furthermore, inactivation of the clpP or clpX genes in the gacA mutant restored the stability of RpoS,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
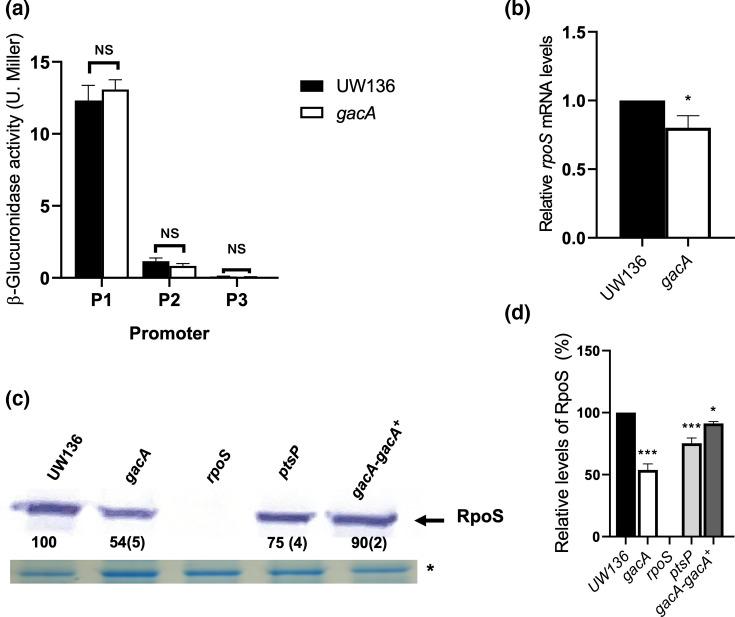 Fig. 1
Fig. 1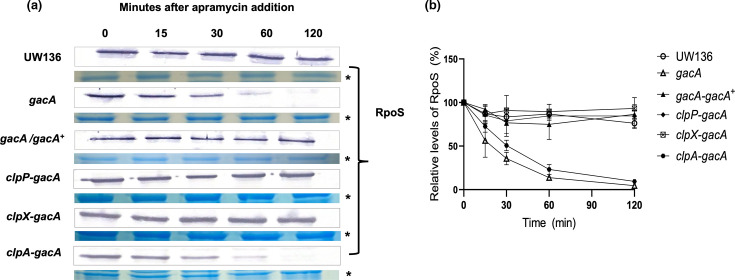 Fig. 2
Fig. 2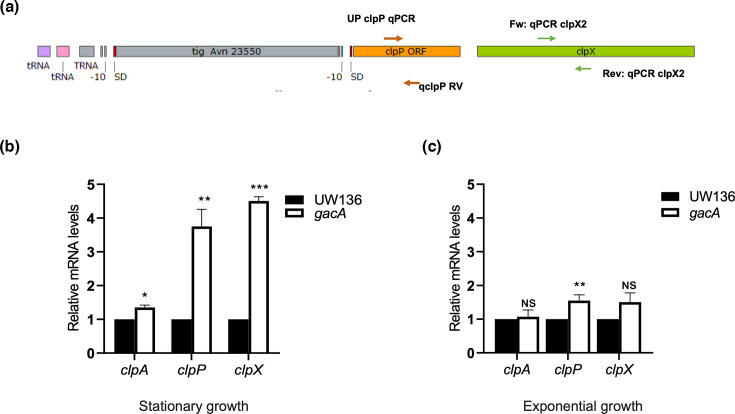 Fig. 3
Fig. 3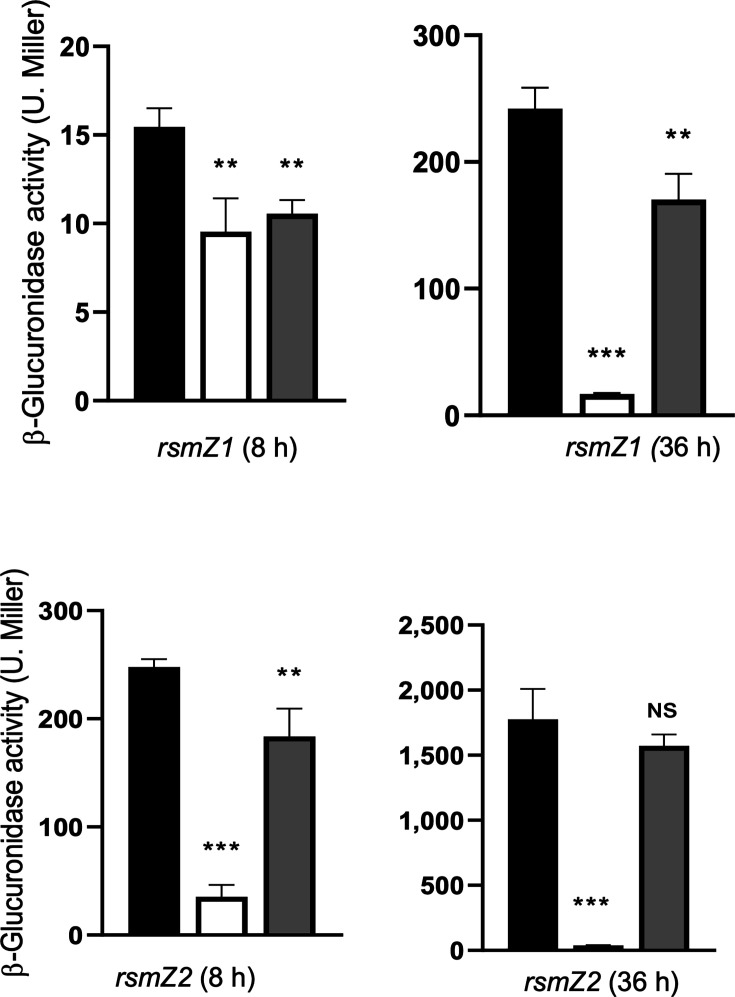 Fig. 4
Fig. 4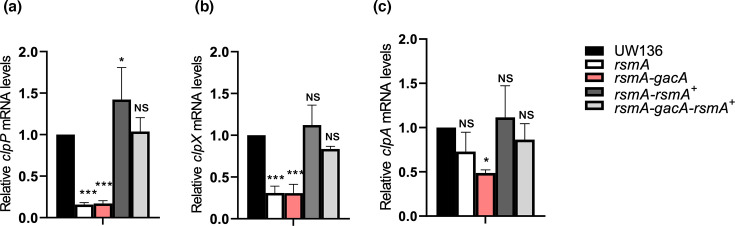 Fig. 5
Fig. 5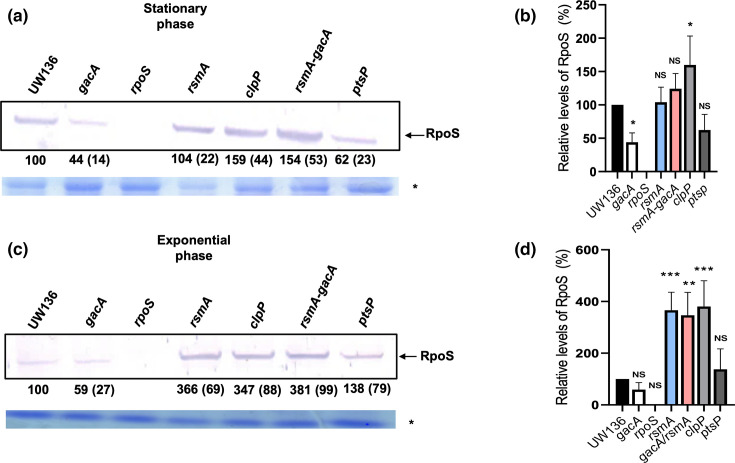 Fig. 6
Fig. 6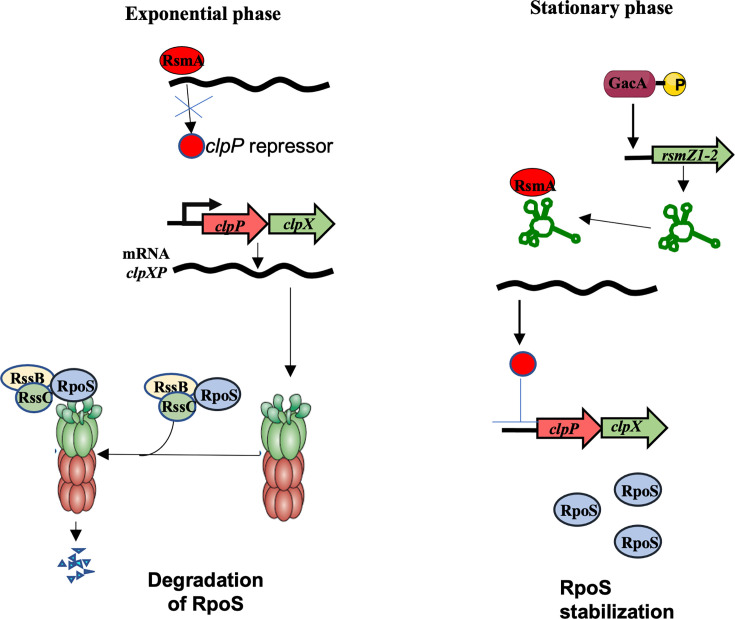 Fig. 7
Fig. 7| Strain | Description | Reference |
|---|---|---|
| TOP10 |
| Thermo Fisher Scientific |
| UW136 | [ | |
| JGWS | UW136 carrying an | [ |
| UW | UW136 derivative carrying an | This study |
| UW | UW136 derivative carrying a | This study |
| Ah | UW136 carrying a | [ |
| AH | AH | This study |
| UW | UW | This study |
| LSW1 | UW136 carrying a | [ |
| LMW33 | UW136 carrying a | [ |
| LMW35 | UW136 carrying a | [ |
| LMW39 | UW136 carrying a | [ |
| UW | LMW33 with the | This study |
| UW | LMW35 with the | This study |
| UW | LMW39 with the | This study |
| UW | UW | This study |
| UW | UW | This study |
| UW | UW136 carrying a transcriptional | [ |
| UW | UW136 carrying a transcriptional | [ |
| UW | UW136 carrying a transcriptional | [ |
| UW | UW | This study |
| UW | UW | This study |
| UW | UW | This study |
| SAWZ1 | UW136 with | [ |
| SAWZ2 | UW136 with | [ |
| SAWGZ1 | AHgacA with | [ |
| SAWGZ2 | AHgacA with | [ |
| AHgacA-rsmZ1/gacA+ | SAWGZ1 carrying plasmid pJETgacA-Tc cointegrated into the chromosome | This study |
| AHgacA-rsmZ2/gacA+ | SAWGZ2 carrying plasmid pJETgacA-Tc cointegrated into the chromosome | This study |
|
| ||
| pJET1.2 | pJET1.2 blunt cloning vector | Thermo Scientific |
| pBSL97 | Plasmid with a Km cassette | [ |
| pBSL98 | Plasmid with a Gm cassette | [ |
| pJET | pJET with | This study |
| pJET | pJET with | This study |
| pJET | pJET with | This study |
| pJET | pJET with | This study |
| pJET | pJET | This study |
| pUMAP1rpoS:: | Plasmid carrying a transcriptional rpoS:: | [ |
| pUMAP2rpoS:: | Plasmid carrying a transcriptional rpoS:: | [ |
| pUMAP3rpoS:: | Plasmid carrying a transcriptional rpoS:: | [ |
| pHP45-Ω-Tc | Plasmid with a Tc cassette | [ |
| pK18mob | Mobilizable plasmid with a KmR marker | [ |
| pkgacA | Pk18mob carrying the gacA gene Km | This study |
| pRK2013 | Mobilizing plasmid, Tra+ Km R | [ |
- —DGAPA
- —CONACHyT
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial Genetics and Biotechnology · RNA and protein synthesis mechanisms · Metalloenzymes and iron-sulfur proteins
Introduction
Azotobacter vinelandii is a soil bacterium belonging to the Pseudomonadaceae family that produces the biodegradable thermoplastic polyhydroxybutyrate (PHB) during the stationary phase. In this bacterium, transcription of the phbBAC operon, which encodes the enzymes responsible for the synthesis of this polymer, is dependent on the stationary sigma factor RpoS [12]. In most Gram-negative bacteria, RpoS controls the general stress response [3]. Accordingly, RpoS protein levels increase during the stationary phase, which is a condition of nutrient limitation [46].
The regulatory mechanisms that control RpoS expression have been extensively studied in Escherichia coli, where regulation occurs at multiple levels, including transcription, translation and proteolytic degradation [710]. A proteolytic process mediated by the ClpXP ATP-dependent protease complex occurs in several Gram-negative bacteria [1112], and in E. coli involves several proteins, including the RssB response regulator, which acts as an adaptor protein that binds RpoS and presents it to the ClpXP protease [13].
In A. vinelandii, as in other bacteria, degradation of RpoS by the ClpXP complex occurs during the exponential growth phase [14]. An operon encoding a RssB-like protein and RssC, a protein annotated as an anti-antisigma factor, was identified in species of the Pseudomonadaceae family [15]. In A. vinelandii and Pseudomonas aeruginosa, inactivation of rssB and/or rssC increases the levels of RpoS during exponential growth and affects RpoS-dependent phenotypes, such as PHB in A. vinelandii and pyocyanin in P. aeruginosa [15]. Moreover, an in vivo interaction between RssB and RpoS was observed only in the presence of RssC. Thus, it was proposed that both RssB and RssC are necessary for RpoS degradation by ClpXP during the exponential phase of growth in bacteria belonging to the Pseudomonadaceae family [15].
RpoS degradation in A. vinelandii was also shown to be under the control of the nitrogen-related phosphotransferase system (PTS^Ntr^) [14], a global regulatory system found in Gram-negative bacteria, composed of the EI^Ntr^, Npr and EIIA^Ntr^ proteins, encoded by the ptsP, ptsO and ptsN genes, respectively [1618]. These proteins participate in a phosphorylation cascade starting from phosphoenolpyruvate, with EIIA^Ntr^ being the final acceptor [19]. In A. vinelandii, degradation of RpoS by the ClpAP chaperone–protease complex occurs during the stationary phase in mutants that carry an unphosphorylated EIIA^Ntr^ [14].
In species of the Pseudomonas genus, the Gac-Rsm signal transduction system, a global regulator of gene expression at the translational level, controls a wide variety of phenotypic traits [2021]. The effector of this system is RsmA, an RNA-binding protein that interacts with its mRNA targets to repress their translation [22]. RsmA activity is controlled by small non-coding RNAs named RsmZ and RsmY, which sequester RsmA to counteract its repressor function [23]. Although RsmA control of translation is mostly negative, it can also be positive and directly activate translation. In P. aeruginosa, RsmA activates translation of phz2, which seems to be carried out by preventing the formation of a secondary structure that sequesters the Shine-Dalgarno SD sequence[24].
Transcription of the rsmZ and rsmY genes is activated by GacA, a response regulator of the GacS/A two-component system [2527]. In A. vinelandii, the Gac-Rsm system regulates the synthesis of alginate, PHB polymers and the phenolic lipids alkylresorcinols. In this bacterium, RsmA binds to the mRNAs encoding proteins involved in the synthesis of these compounds to repress their translation, while the RsmZ1 and RsmZ2 RNAs bind to RsmA to neutralize its activity [2830].
In this study, we provide evidence showing that in a gacA mutant of A. vinelandii derived from strain UW136, RpoS stability during the stationary phase is reduced due to its degradation by ClpXP, but not by ClpAP. Additionally, transcript levels of clpP and clpX are significantly increased. Furthermore, inactivation of rsmA increased RpoS stability during exponential growth and significantly reduced the clpP and clpX transcript levels, indicating that free (unsequestered) RsmA contributes to the reduced RpoS stability and upregulation of ClpXP observed in the gacA mutant.
Taken together, our results reveal that the differences in RpoS stability observed between the exponential and stationary growth phases are controlled by the Gac-Rsm system via a mechanism involving regulation of ClpXP expression.
Methods
Strains, media and culture conditions
The A. vinelandii strain used in this study was UW136 [31]. Bacterial strains and plasmids used or constructed in this study are listed in Table 1. E. coli TOP10 was used for plasmid isolation and maintenance. Media and growth conditions were as follows: A. vinelandii was grown at 30 °C in Peptone Yeast (PY) medium or Burk’s nitrogen-free salts [32] supplemented with 20 g l^−1^ sucrose (BS). Antibiotics were used at the following concentrations (in μg ml^−1^) for A. vinelandii and E. coli, respectively: spectinomycin, 50 and 100; kanamycin, 30 and 3; tetracycline, 40 and 15; gentamicin, 1 and 10; apramycin, 25 (not used for E. coli).
E. coli strains were grown on Luria–Bertani medium at 37 °C.
Construction of mutant strains and plasmids
Oligonucleotides used in this study are listed in Supporting Information Table S1, available in the online Supplementary Material.
To construct the UWrsmA strains, we amplified an 868 bp fragment containing the rsmA gene using RsmAUP and RsmADown oligonucleotides. The amplified fragment was cloned into plasmid pJET1.2 (Thermo Scientific). The resulting plasmid, pJETrsmA, was subjected to reverse PCR using RsmAInvFw and RsmAInvRv primers to introduce a gentamycin-resistant cassette (Gm) from plasmid pBSL98 [33]. The resulting plasmid, pJETrsmA::Gm, was linearized by digestion with ScaI and used to transform the WT A. vinelandii strain UW136, leading to the isolation of the UWrsmA strain.
The UWgacA strain was constructed as follows: UPgacA628bp and DowngacA600bp oligonucleotides were used to amplify the gacA gene, which was subsequently cloned into the pJET1.2 vector, yielding plasmid pJETgacA. To introduce a kanamycin-resistant cassette (Km) from plasmid pBSL97 [33] into the gacA gene, reverse PCR was performed using Fw:gacAinv and Rv:gacAinv oligonucleotides, resulting in a 47 bp deletion in gacA and generation of pJETgacA::Km. This plasmid was used to transform strain UW136, yielding the UWgacA mutant. The presence of the gacA::Km mutation was confirmed by PCR.
The double mutant strain UWrsmA-gacA was isolated by transforming UWrsmA with total DNA from strain UWgacA. Gm- and Km-resistant transformants with a PHB-positive phenotype (Supporting Information Fig. S1) were selected, and the presence of the gacA::Km mutation was confirmed by PCR.
To construct UWclpA-gacA, UWclpP-gacA and UWclpX-gacA double mutant strains*,* LMW33*,* LMW35 and LMW39 strains were transformed with total DNA extracted from AHgacA, which carries a gacA::Gm mutation. Gm- and Km-resistant transformants were isolated and validated via PCR to confirm the *gacA::*Gm mutation.
Transformation of A. vinelandii strains was carried out as described by Page and Sadoff [34].
For the construction of rsmA-complemented strains, a DNA fragment containing the rsmA gene with its native promoter was amplified using primers RsmAUP/RsmADown, respectively. This fragment was cloned into the pJET1.2 vector, generating plasmid pJETrsmA. A tetracycline-resistant cassette (Tc) from plasmid pHP45-Ω-Tc [35] was inserted into the ScaI site of pJET1.2, resulting in plasmid pJETrsmA-Tc.
Because this plasmid does not replicate in A. vinelandii, strains UWrsmA and UWrsmA-gacA were transformed with pJETrsmA-Tc. Tetracycline-resistant transformants, UWrsmA/rsmA^+^ and UWrsmA-gacA/rsmA^+^, were isolated, and PCR analysis confirmed integration by a single recombination event of pJETrsmA-Tc in the corresponding Tc-resistant transformants.
For the construction of gacA-complemented strains, a BglII DNA fragment containing the gacA gene with its native promoter from plasmid pJETgacA was obtained. This fragment was cloned into the BamHI site of the mobilizable Km-resistant plasmid pK18mob [36]. The resultant plasmid, pKgacA, unable to replicate in A. vinelandii, was transferred to strains AHgacA, SAWGZ1 and SAWGZ2 by triparental conjugations mediated by the helper plasmid pRK2013 plasmid [37]. Kanamycin- and gentamycin-resistant transformants showing a PHB^+^ phenotype (Fig. S1), AHgacA/gacA^+^, AHgacA-rsmZ1-gusA/gacA^+^ and AHgacA-rsmZ2-gusA/gacA^+^, were isolated and confirmed by PCR analysis for the presence of the WT gacA gene.
To construct UWgacA derivatives carrying rpoS::gusA transcriptional fusions, plasmids pUMAP1rpoS::gusA, pUMAP2rpoS::gusA and pUMAP3rpoS::gusA [38], linearized by ScaI digestion, were used to transform the UWgacA strain. This resulted in UWgacA-rpoS::gusAP1, UWgacA-rpoS::gusAP2 and UWgacA-rpoS::gusAP3 strains. Integration of rpoS::gusA gene fusions by double homologous recombination was confirmed by PCR.
PCR and qRT-PCR
Total RNA extraction was performed using the TRIzol^™^ Max^™^ Bacterial RNA Isolation Kit (Thermo Scientific) according to the manufacturer’s protocol. To eliminate genomic DNA contamination, RNA was treated with DNase (DNA-free^TM^, Ambion), and RNA concentration was measured by absorbance at 260 nm. RNA purity was assessed using the 260/280 nm absorbance.
cDNA synthesis was performed using the RevertAid ^TM^ First Strand cDNA Synthesis Kit (Fermentas Inc.) and gene-specific primers. Primer sequences for cDNA synthesis and real-time PCR (RT-PCR) are listed in Table S1. The cDNA was used as a template for RT-PCR. Quantification of fluorescence was performed on a LightCycler 480 System (Roche Diagnostics) using SYBR Green dye.
To determine transcript levels of rpoS, clpP, clpX and clpA, the following primers were used: rpoS: qRT-rpoSUp/qRTrpoSDown; clpP: FWqPCRclpP/RVqPCRclpP; clpX: FWqPCR-clpX2/RVqPCR-clpX2; clpA: qRT-clpA Up/qRT-clpA Down.
Western blot assays
RpoS protein was detected by Western blot using polyclonal RpoS-6His antiserum, as described previously [14]. Total protein was extracted from cultures grown in PY after 8 h (exponential phase) or 36 h (stationary phase). To assess in vivo RpoS stability, apramycin (25 µg ml^−1^) was added to cultures to block protein synthesis. Protein concentrations were determined using the Lowry method [39].
Results
The effect of a gacA mutation on RpoS expression
Previously, a gacA mutation in an A. vinelandii strain ATCC9046 (non-isogenic to UW136) was found to exert a negative effect on the level of rpoS transcript, assessed by Northern blot analysis [40]. We investigated whether the expression of RpoS in A. vinelandii strain UW136 is under the control of the Gac/Rsm system by analysing the effect of a gacA mutation on rpoS transcription and on RpoS protein levels. In UW136, rpoS transcription was previously shown to initiate from three promoters (P1, P2 and P3), with P1 accounting for ~95% of rpoS transcriptional activity [38].
Strains UW136 and the UWgacA mutant carrying rpoS::gusA transcriptional fusions containing each of the three promoters were used to determine β-glucuronidase activity in stationary-phase cultures (36 h of growth). Unexpectedly, as shown in Fig. 1a, and in contrast to the ATCC9046 gacA mutant, under these conditions, the UWgacA mutant displayed rpoS transcription levels similar to those of the WT.
A mutation in gacA reduces RpoS protein level. (a) β-Glucuronidase activity in the UWgacA mutant and UW136 WT strains carrying rpoS::gusA transcriptional fusions to the P1, P2 and P3 rpoS promoters. (b) Relative expression of the rpoS gene, as determined by RT-qPCR in UWgacA and UW136 WT strains. (c) Detection of RpoS by Western blot in strains UW136, and in gacA, rpoS, ptsP and gacA/gacA+ mutants. Twenty micrograms of total protein was loaded per sample. RpoS levels in UW136 strain were considered 100%. Values represent the mean of three independent experiments, with sd shown in parentheses. The asterisk () indicates a protein loading control in a Coomassie-stained gel. For panels (a) and (b), data represent the mean of three independent experiments. Error bars indicate sd. Statistical significance was determined using an unpaired Student’s t-test (P<0.05). All experiments were performed with samples harvested after 36 h of growth in PY liquid medium at 30 °C. (d) Mean±sd of three independent biological replicates. Asterisks indicate statistically significant differences compared with WT [unpaired Student’s t-test for panels (a) and (b); one-way ANOVA with Dunnett’s post-hoc test for panel (d); P<0.05].*
The level of rpoS mRNA was also measured by qRT-PCR in the gacA mutant and UW136 strains (Fig. 1b). A reduction of ~20% in rpoS transcript levels was observed in the gacA mutant compared with the WT.
The level of the RpoS protein was determined by Western blot analysis in stationary-phase cultures of UW136 and the gacA strains. We found that RpoS levels in the gacA mutant were reduced to ~54% of those observed in the parental WT UW136 strain; a reduction was also observed in strain LSW1, a ptsP mutant that carries an unphosphorylated EIIA^Ntr^ (Fig. 1c). To confirm that this effect was due to the absence of the GacA protein, we generated the complemented strain UWgacA/gacA^+^, in which the RpoS level was restored to 90% of the WT (Fig. 1c, d).
Taken together, these results suggest that the absence of GacA does not significantly affect rpoS transcription but causes a reduction in RpoS protein levels.
RpoS instability in the gacA mutant is due to its degradation by ClpXP, not ClpAP
To determine whether the reduction in RpoS levels was due to protein instability, we assessed RpoS stability in vivo by adding apramycin to stationary-phase cultures to block protein synthesis. As shown in Fig. 2, RpoS levels remained stable in UW136 and UWgacA/gacA^+^ strains after apramycin addition, whereas in the gacA mutant, RpoS levels significantly declined after 60 min.
The reduction of RpoS level in the gacA mutant is caused by instability due to its degradation by ClpXP. (a) Determination of RpoS stability in the gacA/gacA+ complemented strain and the gacA mutant and its derivatives carrying clpA, clpX and clpP mutations. RpoS levels were determined as described in Fig. 1. The asterisk () in panel (a) denotes a protein loading control in a Coomassie-stained gel. (b) Densitometric analysis of the experiment shown in panel (a). The mean of three independent experiments is presented. RpoS level at time 0 was considered 100%. Error bars indicate sd.*
In A. vinelandii mutants derived from strain UW136 that carry unphosphorylated EIIA^Ntr^, such as ptsP mutants LSW1, RpoS stability is reduced due to its degradation by the ClpAP complex [14].
Additionally, it was previously shown that in a gacA mutant derived from A. vinelandii AEIV (a strain non-isogenic to UW136), EIIA^Ntr^ is found predominantly in an unphosphorylated state [41]. Therefore, we hypothesized that the reduction in RpoS stability observed in the gacA mutant derived from the UW136 strain could also be due to ClpAP-mediated degradation.
To test this, we constructed a gacA-clpA double mutant and evaluated RpoS stability in vivo. As shown in Fig. 2, inactivation of clpA in the gacA mutant did not restore RpoS stability, indicating that RpoS degradation in the gacA mutant is not mediated by ClpAP.
To investigate whether the ClpXP complex was responsible for the observed instability, we analysed RpoS stability in gacA-clpP and gacA-clpX double mutants. As shown in Fig. 2, inactivation of either clpP or clpX restored RpoS stability in the gacA mutant.
These results strongly suggest that ClpXP is responsible for RpoS degradation in the absence of GacA.
GacA regulates the expression of clpP and clpX
In A. vinelandii, as in other bacteria, RpoS proteolysis by ClpXP occurs predominantly during the exponential growth phase [14]. As shown above, the reduced stability of RpoS in stationary-phase gacA mutant appears to be caused by ClpXP-mediated degradation. This raised the question of whether ClpXP expression is regulated by GacA.
We evaluated clpA, clpX and clpP transcript levels by RT-qPCR in WT UW136 and the gacA mutant. A schematic representation of the genetic context of clpP and clpX and the oligonucleotides used is shown in Fig. 3a. As shown in Fig. 3b, a ~fourfold increase in clpP and clpX transcript levels was observed in stationary-phase gacA mutant cultures compared with the WT. In contrast, clpA transcript level was only slightly elevated.
The gacA mutation increases clpP and clpX transcript levels during the stationary phase. (a) Schematic representation of the positions of oligonucleotides used in qRT-PCR to determine clpP and clpX transcript levels. (b) Relative expression of clpP and clpX genes measured by RT-qPCR in the gacA mutant during the stationary growth phase (36 h). (c) Relative expression of clpA, clpP and clpX genes measured by RT-qPCR in the gacA mutant during the exponential growth phase (8 h). Cells were grown in PY liquid medium at 30 °C. The mean of three independent experiments is presented. Error bars indicate sd. Asterisks indicate statistical significance (unpaired Student’s t-test) when comparing each transcript in the gacA mutant versus the WT strain (P<0.05).
Transcript levels were also measured in exponential-phase cultures. A moderate (~50%) increase in clpP expression was observed in the gacA mutant compared with the WT (Fig. 3c). These results indicate that the gacA mutation strongly upregulates ClpXP expression during stationary phase but has only a modest effect during exponential growth. As expected, ClpA expression was not significantly affected.
Effect of an rsmA mutation on clpX and clpP mRNA levels
The translational regulatory activity of RsmA is controlled through its interaction with the small non-coding RNAs RsmZ, which counteract its function. In A. vinelandii, transcription of the RsmZ1 and RsmZ2 RNAs, as determined using rsmZ-gusA transcriptional fusions, occurs during stationary phase and is dependent on GacA [29] (Fig. 4). To confirm the positive regulation of rsmZ transcription by GacA, we constructed the complemented strains AHgacA-rsmZ1-gusA/gacA^+^ and AHgacA-rsmZ2-gusA/gacA^+^. As expected, transcription of rsmZ1 and rsmZ2 during the stationary phase was restored to WT levels in both strains (Fig. 4).
Transcription of the non-coding RNAs RsmZ1 and RsmZ2 is activated during the exponential phase and is dependent on GacA. β-Glucuronidase activity in strains UW136 (black bars) and AHgacA (white bars), carrying rsmZ1-gusA and rsmZ2-gusA transcriptional fusions, and complemented derivatives gacA-rsmZ1::gusA/gacA+ and gacA-rsmZ2:gusA/gacA+ (grey bars). β-Glucuronidase activity was determined in exponential (8 h) and stationary (36 h) phase cultures. Values represent the mean of three independent experiments; error bars, sd.
Accordingly, in exponential-phase cultures of the WT, RsmA is expected to be active and able to bind its target mRNAs. In contrast, during the stationary phase, RsmA becomes inactive due to sequestration by RsmZ1 and RsmZ2. In the gacA mutant, however, transcription of these RNAs is significantly reduced during stationary phase (Fig. 4), allowing RsmA to remain active (unsequestered) throughout both growth phases. Consequently, and consistent with the positive effect of the gacA mutation on clpX and clpP transcript levels described above, inactivation of rsmA was expected to affect the expression of these genes.
As shown in Fig. 5, inactivation of rsmA caused a strong reduction in clpP and clpX transcript levels during stationary phase. This effect is opposite to that observed in the gacA mutant, in which RsmA remains in its active, unsequestered form. We also constructed an rsmA-gacA double mutant. As expected, in this strain, clpP and clpX transcript levels were similar to those observed in the rsmA single mutant (Fig. 5). To confirm that the reduction in clpP and clpX transcripts was due to the absence of RsmA, we generated the complemented strains UWrsmA/rsmA^+^ and UWgacA-rsmA/rsmA^+^. In both cases, clpP and clpX transcript levels were restored to WT levels, or even higher in the case of clpP in the UWrsmA/rsmA^+^ strain (Fig. 5).
Expression of clpP and clpX is downregulated in the absence of RsmA. Transcript levels of clpP (a), clpX (b) and clpA (c), measured by RT-qPCR in rsmA and rsmA-gacA mutants, and in complemented strains rsmA-rsmA+and rsmA-gacA-rsmA+. All experiments were performed using samples collected after 36 h of growth in PY liquid medium at 30 °C. Data represent the mean of three independent experiments. Error bars indicate sd. Statistical significance was determined using an unpaired Student’s t-test (P<0.05).
Together, these findings indicate that, in the absence of RsmA, clpP and clpX expression is significantly downregulated, supporting the conclusion that RsmA acts as a positive regulator of ClpXP expression.
Inactivation of rsmA restores RpoS expression in the gacA mutant
As shown above, ClpXP expression is downregulated in strains carrying an rsmA mutation. Therefore, we hypothesized that RpoS levels would be positively affected in these strains.
We measured RpoS levels in stationary-phase cultures of rsmA and rsmA-gacA strains, where ClpXP-mediated proteolysis reduces RpoS levels in WT cells. Consistent with our hypothesis, in both rsmA and rsmA-gacA mutants, RpoS levels increased to values similar to those observed in the clpP mutant (Fig. 6 A and B).
Inactivation of rsmA increases RpoS levels during the exponential phase and restores RpoS expression in the gacA mutant. Detection of RpoS by Western blot in strains UW136, gacA, rpoS, rsmA, clpP, rsmA-gacA and ptsP in (a) stationary phase (36 h) and (c) exponential phase (8 h) was carried out as described in Fig. 1c. Asterisks () in panels (a) and (c) indicate loading controls from Coomassie-stained gels. (b and d) Densitometric quantification of RpoS protein levels from panels (a) and (c), respectively. Data represent the mean±sd of three independent experiments. Asterisks in panels (c) and (d) indicate statistically significant differences compared with WT (one-way ANOVA with Dunnett’s post-hoc test; P<0.05).*
We also analysed RpoS levels in exponential-phase cultures, as shown in Fig. 6 C and D.
RpoS levels in the rsmA-gacA double mutant were even higher than in the WT, while in the rsmA single mutant, they were similar to those of the WT.
These findings further support the conclusion that GacA-dependent regulation of RpoS proteolysis involves RsmA as a positive effector of clpXP expression.
Discussion
The degradation of RpoS is tightly regulated at different stages of bacterial growth. In E. coli, during the exponential phase, the rapid degradation of RpoS by the ClpXP complex is mediated by the response regulator RssB [42], whereas in A. vinelandii and P. aeruginosa, both RssB and RssC participate in this process [15]. In contrast, the stationary-phase accumulation of RpoS mainly results from increased protein stability [44344]. However, the mechanism responsible for RpoS stabilization during this phase remains poorly understood.
In the present study, we describe a previously uncharacterized regulatory mechanism for the control of RpoS stability in A. vinelandii, mediated by the post-transcriptional regulatory system Gac-Rsm. We show that in a gacA mutant derived from UW136, RpoS protein levels decreased during the stationary phase. Although a negative effect of a gacA mutation on rpoS transcript levels, as determined by Southern blot analysis, was previously reported in A. vinelandii strain ATCC9046 [40], no significant effect on rpoS transcription was observed in strain UW136. Therefore, the reduction in RpoS levels in the UW136 gacA mutant is not due to transcriptional downregulation, but rather to decreased RpoS protein stability. The discrepancy in the rpoS transcript levels between ATCC9046 and UW136 gacA mutants may be explained by the fact that these strains are not isogenic.
In previous studies, we reported that the global regulatory PTS^Ntr^ system, composed of EI^Ntr^, Npr and EIIA^Ntr^ proteins and conserved in Gram-negative bacteria, controls RpoS stability. When unphosphorylated, EIIA^Ntr^ promotes ClpAP-dependent degradation of RpoS during the stationary phase [14]. Since in a gacA mutant derived from AEIV (a strain non-isogenic to UW136), EIIA^Ntr^ is predominantly unphosphorylated [41], we initially hypothesized that the reduced RpoS stability observed in the UW136 gacA mutant could result from ClpAP-mediated proteolysis. However, inactivation of clpA in the gacA mutant did not restore RpoS stability (Fig. 2), indicating that this pathway is not responsible. In contrast, inactivation of clpP or clpX genes in the gacA mutant restored RpoS stability (Fig. 2), implying that the instability of RpoS in the gacA mutant was due to ClpXP-dependent proteolysis rather than ClpAP activity.
In Pseudomonas species phylogenetically closely related to A. vinelandii [45], the post-transcriptional Gac-Rsm regulatory system controls diverse cellular processes. The response regulator GacA activates transcription of one or more small non-coding RNAs termed RsmZ, RsmY and RsmX. These RNAs sequester RsmA, a translational regulatory protein, thereby counteracting its repressor activity on target mRNAs [23,4649].
In A. vinelandii, the Gac-Rsm system regulates the synthesis of PHB, alginate and alkylresorcinols by controlling translation of algD, phbR and arpR mRNAs through RsmA binding [2830]. Expression of eight small RNAs (RsmZ1–7 and RsmY) is low during the exponential phase but strongly induced during the stationary phase in the WT UW136 strain, whereas this induction is absent in a gacA mutant. In addition, RsmA was shown to bind RsmZ1 and RsmZ2 [29]. Consequently, in the gacA mutant, RsmA remains active (unsequestered) in both growth phases, while in the WT, it is active only during the exponential phase.
Our results show that inactivation of rsmA increased RpoS levels during the exponential phase by ~3.5-fold, an effect comparable to that observed in a clpP mutant (Fig. 6). These findings indicate that RsmA promotes RpoS degradation by increasing clpP and clpX expression (Fig. 6).
Consistent with this interpretation, clpP and clpX transcript levels were elevated in the gacA mutant, where RsmA remains unsequestered and active, but were reduced in the rsmA mutant, correlating with the observed RpoS stability patterns (Figs1c, 56bundefined). Although growth phase-dependent regulation of clpP and clpX transcription has been reported in other bacteria [5051], regulation of clpP and clpX by the Gac-Rsm system has not previously been reported.
While RsmA in A. vinelandii generally functions as a translational repressor [2830], the reduction of clpP and clpX transcripts in the rsmA mutant suggests a positive regulatory role on these genes at the level of transcription rather than translation. If this is the case, RsmA may regulate clpP and clpX expression indirectly, for example, by inhibiting the translation of a negative regulator. The possibility that RsmA might directly bind clpP and clpX transcripts to promote their translation, as reported for the E. coli flhDC or P. aeruginosa phz2 transcripts [245253], is not ruled out. However, analysis of the Shine-Dalgarno regions and upstream clpP and clpX regions revealed no canonical RsmA-binding motifs corresponding to the SELEX-derived consensus sequence (CAMGGAYG) identified for P. aeruginosa RsmA targets [54].
Taken together, our results support the model proposed in Fig. 7. During exponential growth, low RsmZ expression allows RsmA to remain unsequestered, enabling it to inhibit the expression of a putative negative regulator of ClpXP, leading to increased clpP and clpX transcription and, consequently, enhanced ClpXP-dependent degradation of RpoS. In the stationary phase, GacA induces rsmZ transcription, resulting in RsmA sequestration, repression of clpP and clpX and stabilization of RpoS. This regulatory strategy is consistent with the observation that transcription of A. vinelandii genes involved in alginate, PHB and alkylresorcinols synthesis initiates from RpoS-dependent promoters. Thus, in addition to post-transcriptional regulation of these genes by the Gac-Rsm system, this pathway indirectly promotes their optimal transcription by stabilizing RpoS.
Proposed model for the regulation of RpoS levels during exponential and stationary phases. During the exponential growth phase, transcription of the rsmZ RNAs is significantly reduced. As a result, unsequestered RsmA protein might inhibit the translation of a putative repressor of clpP and clpX transcription, resulting in ClpXP-dependent degradation of RpoS. During the stationary phase, GacA is phosphorylated by GacS and activates transcription of the rsmZ genes. The accumulation of rsmZ RNAs results in sequestering RsmA, inhibiting its activity, thereby downregulating clpXP repression and allowing RpoS accumulation.
Finally, an interplay between Gac-Rsm and RpoS regulons has also been reported in Pseudomonas species. However, in these bacteria, Gac-Rsm typically regulates RpoS at the transcriptional or translational level, rather than by controlling proteolysis, as shown in Pseudomonas fluorescens, Pseudomonas chlororaphis, Pseudomonas putida and P. aeruginosa [5559].
Supplementary material
10.1099/mic.0.001672Uncited Supplementary Material 1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Peralta-Gil M Segura D Guzmán J Servín-González L Espín G Expression of the Azotobacter vinelandii poly-beta-hydroxybutyrate biosynthetic phb BAC operon is driven by two overlapping promoters and is dependent on the transcriptional activator Phb RJ Bacteriol 20021845672567710.1128/JB.184.20.5672-5677.200212270825 PMC 139623 · doi ↗ · pubmed ↗
- 2Hernandez-Eligio A Castellanos M Moreno S Espín G Transcriptional activation of the Azotobacter vinelandii polyhydroxybutyrate biosynthetic genes phb BAC by Phb R and Rpo S Microbiology 20111573014302310.1099/mic.0.051649-021778206 · doi ↗ · pubmed ↗
- 3Bouillet S Bauer TS Gottesman S Rpo S and the bacterial general stress response Microbiol Mol Biol Rev 202488 e 001512210.1128/mmbr.00151-2238411096 PMC 10966952 · doi ↗ · pubmed ↗
- 4Lange R Hengge-Aronis R The cellular concentration of the sigma S subunit of RNA polymerase in Escherichia coli is controlled at the levels of transcription, translation, and protein stability Genes Dev 199481600161210.1101/gad.8.13.16007525405 · doi ↗ · pubmed ↗
- 5Mandel MJ Silhavy TJ Starvation for different nutrients in Escherichia coli results in differential modulation of Rpo S levels and stability J Bacteriol 200518743444210.1128/JB.187.2.434-442.200515629914 PMC 543567 · doi ↗ · pubmed ↗
- 6Hengge R Proteolysis of σS (Rpo S) and the general stress response in Escherichia coli Res Microbiol 200916066767610.1016/j.resmic.2009.08.01419765651 · doi ↗ · pubmed ↗
- 7Hengge-Aronis R Signal transduction and regulatory mechanisms involved in control of the sigma(S) (Rpo S) subunit of RNA polymerase Microbiol Mol Biol Rev 20026637339510.1128/MMBR.66.3.373-395.200212208995 PMC 120795 · doi ↗ · pubmed ↗
- 8Mika F Hengge R A two-component phosphotransfer network involving Arc B, Arc A, and Rss B coordinates synthesis and proteolysis of sigma S (Rpo S) in E. coli Genes Dev 2005192770278110.1101/gad.35370516291649 PMC 1283968 · doi ↗ · pubmed ↗