TG221: An Experimental Model for Liver Cancer Prevention and Treatment Approaches
Elisa Callegari, Angelo Michilli, Farzaneh Moshiri, Bruno De Siena, Laura Gramantieri, Massimo Negrini, Silvia Sabbioni

TL;DR
The TG221 mouse model helps study liver cancer by mimicking human HCC and testing new treatments like metformin and miRNA-based therapies.
Contribution
TG221 is a novel transgenic mouse model for studying miRNA-driven liver cancer and testing RNA-based therapies.
Findings
Metformin prevents tumor formation and suppresses key signaling pathways in early fibrosis.
miR-199a-3p replacement reduces tumor burden and improves liver integrity.
A miR-199a-3p-responsive oncolytic virus replicates selectively in tumors with low toxicity.
Abstract
Hepatocellular carcinoma (HCC) is a leading cause of cancer mortality. It usually arises in cirrhotic liver, where chronic inflammation and fibrosis create a tumor-permissive microenvironment. Dysregulation of microRNAs (miRNAs), particularly upregulation of the oncomiR miR-221 and loss of the tumor suppressor miR-199a-3p represent key drivers of liver carcinogenesis. The TG221 transgenic mouse, designed to overexpress miR-221 in hepatocytes, provides a relevant in vivo platform for mechanistic studies and for testing preventive and therapeutic approaches. The TG221 model recapitulates miR-221-driven tumorigenesis, including suppression of p27, p57 and Bmf. It is characterized by steatohepatitic injury and accelerated tumor formation after genotoxic challenge. In the cirrhotic CCl4-induced background, TG221 mice develop fibrosis and cirrhosis followed by dysplastic and malignant…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
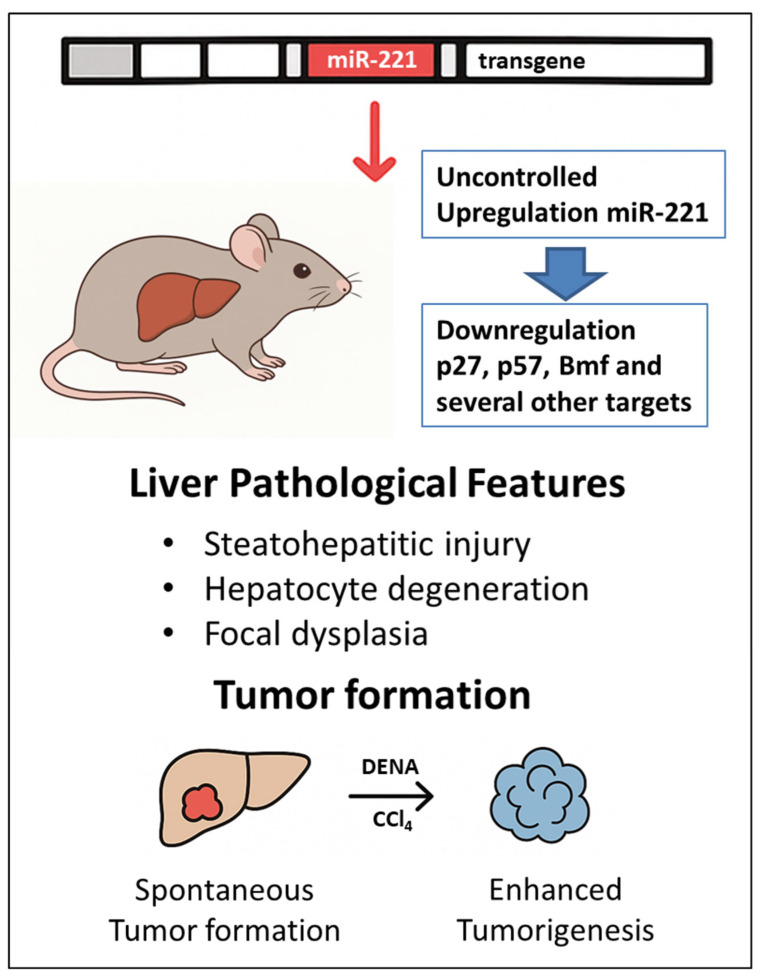 Figure 1
Figure 1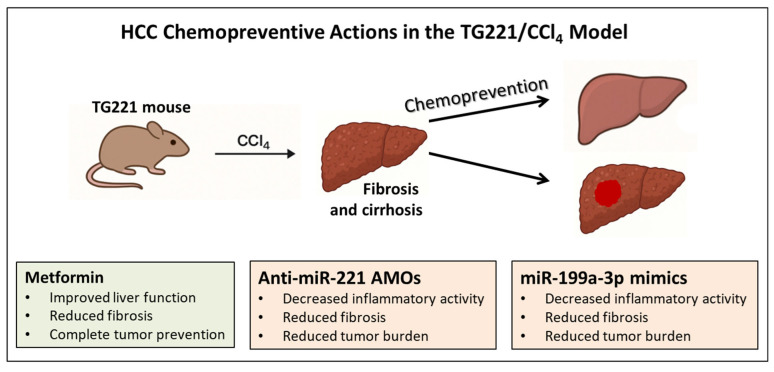 Figure 2
Figure 2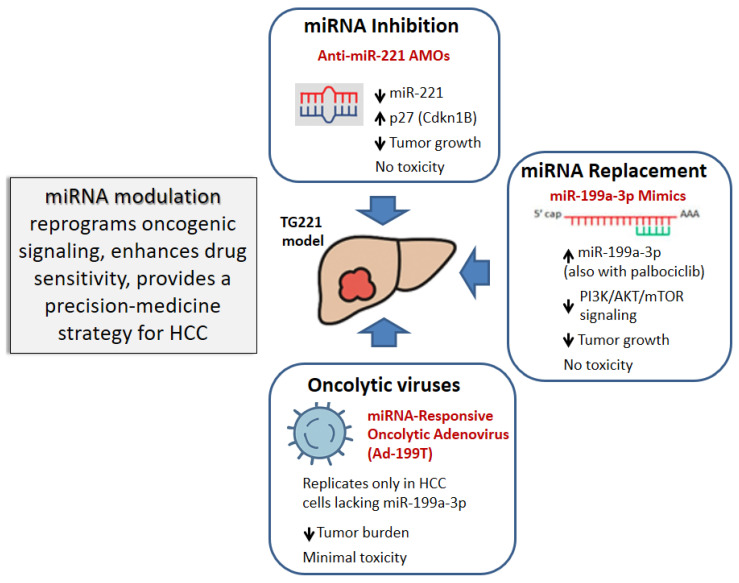 Figure 3
Figure 3- —Italian Ministry of University and Research
- —University of Ferrara
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMetabolism, Diabetes, and Cancer · Cancer-related Molecular Pathways · MicroRNA in disease regulation
1. Introduction
Hepatocellular carcinoma (HCC) represents a major global health burden and is the third leading cause of cancer-related mortality worldwide [1]. Its poor prognosis largely reflects late-stage diagnosis and the limited efficacy of available systemic therapies, which typically provide only transient disease control without achieving durable remission.
HCC arises through a multistep process involving chronic liver injury, inflammation, fibrosis, cirrhosis, and ultimately malignant transformation. The majority of cases develop in the setting of chronic liver disease, commonly associated with hepatitis B or C virus infection, excessive alcohol consumption, or metabolic dysfunction leading to non-alcoholic fatty liver disease (NAFLD) and cirrhosis [2].
Cirrhosis, a preneoplastic condition present in approximately 80% of HCC patients, establishes a tumor-permissive microenvironment characterized by chronic inflammation, oxidative stress, dysregulated regenerative responses and genomic instability. Because hepatocarcinogenesis progresses through well-defined pathological stages over several years, there is a substantial opportunity to identify high-risk individuals who may benefit from targeted surveillance and preventive interventions.
Once established, HCC typically exhibits an aggressive clinical course. Despite therapeutic advances, most notably the introduction of immune checkpoint inhibitor regimens based on atezolizumab plus bevacizumab or durvalumab plus tremelimumab, median survival in patients with advanced, unresectable disease remains unsatisfactory [3]. Historically, multikinase inhibitors such as sorafenib [4] and, more recently, lenvatinib [5] constituted the mainstay of systemic therapy; however, primary and acquired resistance markedly limited their long-term benefit. They still remain important options for patients who are ineligible for or unable to tolerate immunotherapy-based treatments.
Accordingly, efforts to enhance the efficacy of existing therapies, develop effective preventive strategies and identify robust biomarkers for early detection remain critical priorities to improve patient outcomes and reduce HCC-related mortality.
A wide range of animal models is available for the study of HCC, including genetically engineered mouse models, chemically or diet-induced models (such as NASH-associated HCC), patient-derived xenografts (PDX) and tumoroids, orthotopic or transplantable models and zebrafish models (Table 1) [6,7,8,9,10,11,12]. However, these systems only rarely recapitulate the multistep progression that characterizes human HCC. Rodent models, particularly genetically engineered strains targeting oncogenic drivers, are to be combined with models that reproduce chronic liver disease and its evolution, in order to more accurately mimic the human condition. Such combined approaches can provide powerful tools for investigating HCC pathogenesis and for evaluating novel therapeutic strategies.
This review summarizes studies employing the TG221 transgenic mouse model, which has provided fundamental mechanistic insights into miRNA-driven hepatocarcinogenesis and has emerged as a versatile platform for preclinical testing of preventive and therapeutic interventions targeting molecular drivers of liver cancer. Compared with existing reviews, the present work offers a deeper and distinctive positioning of the TG221 model as a genetically defined, miR-221-driven, immunocompetent system, introducing a miRNA-centric perspective on hepatocarcinogenesis. In addition, it highlights the unique dual applicability of TG221 to both non-cirrhotic and cirrhotic HCC and underscores its direct suitability for evaluating RNA-based chemopreventive and therapeutic strategies, aspects that were not specifically addressed in previous general reviews of HCC models.
Literature Search and Selection Approach
This article is a narrative review focused on the TG221 mouse model. The literature was identified through PubMed searches using the keywords TG221, miR-221, hepatocellular carcinoma mouse models, and miRNA therapeutics, complemented by citation tracking of seminal studies and inclusion of representative independent reviews. Potential selection bias is acknowledged, as a substantial proportion of the cited studies were authored by contributors to the present review.
2. The miR-221 Transgenic Mouse Model TG221
MicroRNAs (miRNAs) are short, noncoding RNAs (20–24 nucleotides) that regulate gene expression post-transcriptionally and are now recognized as key modulators of oncogenesis [30,31]. Dysregulated miRNA expression is a hallmark of HCC, where specific miRNAs critically influence tumor initiation, progression, and therapeutic resistance. Depending on their expression pattern and gene targets, miRNAs may act as oncogenes (oncomiRs) or tumor suppressors.
Among oncomiRs, miR-221 is one of the most consistently upregulated species in human HCC [32,33]. Its overexpression correlates with aggressive tumor features, including poor differentiation, vascular invasion, metastasis, and early recurrence [34,35]. Mechanistically, miR-221 promotes hepatocyte proliferation and survival by repressing key tumor suppressors, Cdkn1b/p27, Cdkn1c/p57, and the pro-apoptotic factor Bmf, as well as several other target genes, thereby driving cell-cycle progression and resistance to apoptosis in cancer cells [32,33,36,37,38,39,40,41].
Given the biological and clinical relevance of miR-221 dysregulation in HCC, the development of in vivo models recapitulating its pathogenic role is essential for elucidating its function and for preclinical testing of RNA-based therapeutic and preventive strategies (Figure 1).
2.1. Generation and Molecular Characterization
To directly assess the oncogenic potential of miR-221 in vivo, a transgenic mouse line (TG221) was generated to constitutively overexpress miR-221 in a hepatocyte-specific manner [42]. The construct, derived from an expression vector including a chimeric regulatory cassette combining the hepatocyte-specific α1-antitrypsin promoter with the hepatitis B virus enhancer II, ensured strong liver-restricted transcription. The murine mmu-miR-221 locus was cloned downstream of this cassette, and the transgene was microinjected into fertilized B6D2F2 oocytes.
Quantitative PCR confirmed sustained hepatic overexpression of miR-221 across development and adulthood, at levels comparable to those observed in human HCC. Western blot analyses demonstrated marked downregulation of canonical miR-221 targets, including p27, p57, and Bmf, in the liver of TG221 mice relative to wild-type (WT) controls.
TG221 tumors also recapitulated miRNA signatures characteristic of human HCC, showing upregulation of miR-221 and miR-21 and downregulation of hepatocyte-specific miRNAs miR-122 and miR-199a-3p, reflecting conserved oncogenic pathways that drive hepatocarcinogenesis.
2.2. Pathological and Molecular Features
Liver of TG221 mice exhibits steatohepatitic injury, hepatocyte degeneration, and focal dysplasia. Approximately 50% of male TG221 mice develop spontaneous hepatic tumor nodules between 9 and 12 months of age. Although these lesions arise in non-cirrhotic livers, the accompanying metabolic alterations and steatosis recapitulate key features of human HCC.
When challenged with the carcinogen diethylnitrosamine (DEN), all male TG221 mice developed multiple liver tumors within four months, demonstrating a strong synergistic effect between miR-221 overexpression and genotoxic injury. Compared with DEN-treated WT controls, TG221 mice exhibited an earlier development and a significantly greater tumor burden, with increased nodule number and size.
In line with human HCC, the TG221 mouse model exhibits a markedly higher susceptibility to liver cancer in males. Female TG221 mice do not develop spontaneous tumors and only a fraction of females develop HCC after 12 months following DEN challenge. Although this sex bias has not been directly investigated in this model, it is plausible that, similarly to human, androgens promote hepatocarcinogenesis in males through activation of androgen receptor (AR) signaling. In contrast, estrogens in females exert a protective effect by suppressing pro-inflammatory cytokine production (e.g., interleukin-6), thereby reducing oxidative stress and fibrogenesis.
3. TG221 Model and HCC Prevention
Hepatocarcinogenesis is a decades-long process that unfolds through chronic inflammation, progressive fibrosis and cirrhosis. This extended latency provides a unique window of opportunities for prophylactic intervention. The TG221 model, given its pronounced predisposition to liver tumorigenesis, represents a powerful system for evaluating chemopreventive strategies.
A major limitation of existing HCC mouse models is that tumors arise in histologically normal livers, failing to reflect the fibrotic/cirrhotic milieu typical of human disease. Moreover, chemically induced cirrhosis in animal models (e.g., by chronic CCl_4_ administration) rarely progresses to cancer [43,44,45,46]. To overcome these limitations, cirrhosis was induced via chronic oral CCl_4_ administration in TG221 mice. Histopathology revealed extensive fibrosis, nodular regeneration and activation of fibrotic and steatosis markers. Importantly, unlike WT controls, CCl_4_-treated TG221 mice developed dysplastic and neoplastic lesions consistent with HCC, thereby providing a preclinical model that faithfully recapitulates the natural history of human HCC [47,48] (Figure 2).
3.1. Chemopreventive Action of Metformin
This combined CCl_4_/TG221 model was used to test chemopreventive interventions. Metformin, an antidiabetic drug epidemiologically linked to reduced HCC incidence in type 2 diabetic patients [49,50,51], was evaluated for preventive action in nondiabetic liver disease. Beginning at fibrosis onset, mice received metformin (300 mg/kg/day, orally) for 24 weeks.
Metformin improved liver function, attenuated fibrosis by suppressing stellate cell activation and collagen deposition (α-SMA, Col1a1, Col3a1, Col4a1), reduced lipid accumulation via LKB1-AMPK activation and inhibitory phosphorylation of acetyl-CoA carboxylase and, most notably, completely prevented macroscopic tumor development in CCl_4_-challenged TG221 mice. Transcriptomic profiling showed suppression of PI3K/AKT/mTOR, KRAS and mTORC1 signaling, accompanied by enhanced apoptosis [47].
These findings indicate that early administration of metformin in fibrotic, non-diabetic livers prevented progression to cirrhosis and blocked hepatocarcinogenesis by reprogramming metabolic and survival pathways, thereby providing the experimental basis for repurposing metformin as a safe, low-cost chemopreventive approach for patients with liver fibrosis, independent of their diabetic status.
The metformin dose used in TG221 mice (300 mg/kg/day) corresponds, after allometric scaling [52,53], to exposures achievable in patients with type 2 diabetes receiving standard therapeutic doses (approximately 1.5–2 g/day), supporting the translational relevance of this chemopreventive approach in metabolically at-risk or advanced fibrosis populations, supporting its consideration for HCC chemoprevention trials in at-risk selected groups.
3.2. Targeting miRNA Dysregulation for Chemoprevention
miRNAs serve as master regulators of hepatocarcinogenesis, suggesting that early correction of alterations in pathogenic miRNAs could represent a promising preventive approach. In the TG221/CCl_4_ model, inhibition of the oncogenic miR-221 and replacement of the tumor-suppressive miR-199a-3p were evaluated.
Systemic delivery of anti-miR-221 oligonucleotides or miR-199a-3p mimics, facilitated by the use of lipid nanoparticles as vehicles [54], effectively blocked the malignant progression of dysplastic nodules, significantly reducing tumor number and size without detectable toxicity [48]. Compared with controls, treated animals exhibited a marked reduction in both number and size of hepatic nodules, along with a shift toward a loss of dysplastic phenotype. Moreover, treated mice showed decreased inflammation, reduced fibrosis and reactivation of tumor-suppressive pathways, underscoring the central role of these miRNAs in the earliest steps of liver cell transformation [48]. Together, these results demonstrate that targeted miRNA modulation can reprogram the tumor-permissive microenvironment, leading to a stop of hepatocarcinogenesis at a precancerous stage. To this end, the TG221/CCl_4_ model provided a robust platform for preclinical evaluation of RNA-based chemopreventive strategies.
4. TG221 Model and miRNA-Based Therapeutic Strategies
Despite recent advances, systemic therapy for advanced HCC remains suboptimal. Sorafenib, the first agent to confer a survival benefit, is limited by toxicity and inevitable resistance. Next-generation TKIs offer modest benefits. Immunotherapy regimens improve survival in about one-third of patients.
The TG221 model offers a physiologically relevant platform for evaluating miRNA-based therapeutics and combination strategies (Figure 3).
4.1. MiRNA Inhibition
The study by Callegari et al. [42], which led to the development of the TG221 transgenic mouse, established the role of miR-221 as a tumor driver as well as the therapeutic feasibility and safety of miR-221 silencing as an antitumor approach. Synthetic anti-miR oligonucleotides (AMOs), have proven effective for silencing miRNA activity in vivo [55,56]. In the TG221 model, intravenous delivery of synthetic 2′-O-methyl modified anti-miR-221 oligonucleotides (anti-miR-221 AMOs) successfully silenced miR-221 expression in the liver and the circulatory system, which was functionally confirmed by a concurrent increase in Cdkn1b/p27 protein expression, thus restoring a mechanism for cell cycle control. Crucially, systemic administration of AMOs to DEN-treated TG mice resulted in a significant reduction in both the number and the size of liver tumor nodules compared to untreated controls, demonstrating that miR-221 can be effectively targeted to reduce tumor burden. As reported by Krützfeldt et al., in vivo silencing of microRNAs using antagomirs is relatively long-lasting; in their seminal study, miR-122 remained undetectable for up to 23 days following a single systemic administration [56]. Consistent with these findings, in the TG221 model, mice received repeated intravenous injections of anti-miR-221 antagomirs every 15 days (10 mg/kg in saline at 60, 75, and 90 days of age). Throughout the treatment period, hepatic miR-221 levels were significantly and persistently reduced [42]. This successful in vivo inhibition, achieved without appreciable toxicity, confirms the therapeutic potential of anti-miRNA approaches for liver cancer [42].
4.2. MiRNA Replacement
MicroRNAs (miRNAs) have emerged as experimental therapeutic molecules against cancer, offering potential strategies such as the restoration of tumor suppressor miRNAs [30,57,58,59]. MiR-199a-3p is highly expressed in normal liver but downregulated in virtually all HCCs, a decrement correlated with poor prognosis. Given the downregulation of miR-199a-3p also in TG221 tumors, the therapeutic efficacy of a miRNA replacement strategy, based on miR-199a-3p mimic molecules, was investigated in the TG221 mouse model. MiR-199a-3p replacement was achieved using unmodified single-stranded RNA oligonucleotides, which are characterized by low intrinsic stability and whose biological effects typically fade within three days. Accordingly, in vivo administration was performed intraperitoneally at a dose of 5 mg/kg, three times per week for three consecutive weeks, using a lipid-nanoparticle-based formulation to enhance oligonucleotide stability, bioavailability and cellular uptake [60].
The administration of miR-199a-3p mimics in TG221 mice harboring liver cancer led to a significant reduction in both the number and the size of tumor nodules compared to control animals. Mechanistically, this anti-tumor activity results from the enforced expression of miR-199a-3p, which effectively downregulates in vivo its direct targets MTOR (mechanistic target of rapamycin) and PAK4 (p21 activated kinase 4), thereby suppressing PI3K/AKT/mTOR signaling This modulation ultimately led to the repression of the oncogenic transcription factor FOXM1, which is otherwise overexpressed in several solid tumors, including HCC and is associated with poor prognosis [61,62]. Remarkably, the anti-tumor activity achieved by miR-199a-3p mimics in the TG221 model was comparable to that obtained using sorafenib, in tumor-bearing TG221 mice with comparable disease stage, using matched treatment windows and tumor burden endpoints (nodule number and size). These results suggest that miR-199a-3p may be considered a promising HCC therapeutic option.
4.3. Combination Strategies and Sorafenib Resistance
The above findings established the basis for designing miRNA-based combination strategies. The miR-199a-3p mimics were found to act synergistically with the CDK4/6 inhibitor palbociclib. In fact, while palbociclib stops cell-cycle progression, it also triggers compensatory AKT activation, a response that miR-199a-3p can effectively suppress, thus providing the rationale for combining the two agents.
In TG221 mice bearing established liver tumors, the palbociclib plus miR-199a-3p mimics regimen produced the greatest reduction in tumor nodule volume compared with single-agent palbociclib, miR-199a-3p mimics, sorafenib, or control treatments. Importantly, body weight remained stable, indicating that the combination did not cause overt toxicity. Mechanistically, the dual regimen simultaneously inhibited the mTOR and CDK/RB1 pathways while preventing the compensatory AKT activation, resulting in enhanced antitumor activity with good tolerability [63].
Because the palbociclib–miR-199a-3p combination regimen does not include sorafenib, its activity was assessed in sorafenib-resistant cells, where it was indeed found to be effective [63]. In xenograft models generated from sorafenib-resistant HCC cells, enforced miR-199a-3p expression significantly enhanced the antitumor response to palbociclib. In a paired implantation design, AAV-miR-199a-3p transduced cells were injected into one flank, while AAV-control cells were implanted into the opposite contralateral flank, enabling direct intra-animal comparison of treatment effects. As anticipated and consistent with the resistant phenotype, sorafenib exerted minimal activity in both xenograft types. In contrast, across all animals, xenografts derived from miR-199a-3p-expressing cells, but not control-transduced cells, exhibited a markedly reduced growth in response to palbociclib. These results demonstrated that the combination of palbociclib- and miR-199a-3p-based strategies represents a potential approach to overcome sorafenib resistance in HCC [63]. Collectively, these findings highlight miR-199a-3p replacement as a potent sensitizer of CDK4/6 inhibition and support combined miRNA-targeted and cell-cycle-directed strategies as a promising therapeutic approach for HCC.
MiR-221 has also been implicated in sorafenib resistance through suppression of Caspase-3-mediated apoptosis [64], suggesting that the TG221 model could be employed for testing the use of anti-miR-221 in overcoming sorafenib resistance.
4.4. Oncolytic Viruses
Differential miRNA expression between normal hepatocytes and tumor cells can potentially be exploited to enhance the selectivity of oncolytic virotherapy. Oncolytic viruses are genetically engineered viruses that are designed to selectively infect, replicate within and kill cancer cells while sparing the normal ones. They represent an emerging class of cancer therapeutics that leverage both direct tumor cell lysis and stimulation of anti-tumor immunity. Ongoing research and clinical trials suggest that oncolytic viruses will play an increasingly important role in multimodal cancer treatment strategies. Multiple clinical trials are ongoing in solid tumors, often in combination with checkpoint inhibitors or radiation therapy [65,66,67,68]. The first FDA-approved oncolytic virus therapy was Talimogene laherparepvec (T-VEC), an HSV-1-based oncolytic virus engineered to express GM-CSF approved for metastatic melanoma [69].
To enhance tumor selectivity, a miRNA-responsive oncolytic adenovirus (Ad-199T) was designed to exploit the differential expression of miR-199a-3p, abundant in normal liver but absent in HCC. In TG221 mice, intravenous administration of Ad-199T achieved preferential replication within tumors, reducing tumor burden by 2–3-fold compared to controls and exhibiting minimal hepatotoxicity. These results highlight the potential of miRNA-regulated oncolytic viral therapy as a precision strategy for HCC [70].
Nevertheless, the approach presents some relevant translational constraints, including reduced viral replication in cirrhotic tissue, innate immune activation and anti-adenovirus immunity, which makes the approach as a proof-of-concept strategy requiring further optimization.
5. Conclusions
The TG221 transgenic mouse model has been proven essential for elucidating the oncogenic role of miR-221 in hepatocarcinogenesis. Its miRNA expression profile closely mirrors human HCC, including upregulation of oncomiRs (miR-221 and miR-21) and downregulation of tumor suppressors miR-122 and miR-199a-3p. Beyond mechanistic value, TG221 is highly versatile for translational research, enabling evaluation of miRNA inhibition, miRNA replacement, oncolytic strategies, and drug-combination regimens.
The incorporation of a cirrhotic background (CCl_4_/TG221) further enhances translational relevance, providing a unique platform for testing chemopreventive interventions in high-risk liver disease.
Compared with the preclinical HCC models summarized in a recent published review [71], the TG221 transgenic mouse offers a distinct and complementary way to study liver cancer. Table 2 highlights how TG221 differs from commonly used mouse models, and Table 3 illustrates the specific gaps this model helps to address.
First, unlike chemically or diet-induced models (such as DEN, CCl_4_, Western diet, or NASH-like regimens), which depend on random DNA damage and long-term inflammation to create genetically diverse tumors, TG221 provides a genetically defined system. It is driven by overexpression of miR-221, one of the most consistently upregulated oncomiRs in human HCC. This leads to tumors that are more controlled, reproducible, and easier to interpret mechanistically.
Second, while xenograft and patient-derived xenograft (PDX) models reproduce human tumor architecture, they require immunodeficient mice, making it impossible to study interactions between the tumor and the immune system. In contrast, TG221 mice develop tumors in a fully immunocompetent host, and the model can be paired with cirrhosis-inducing treatments to better mimic the inflammatory environment of advanced liver disease.
Third, compared with traditional genetically engineered mouse models based on activating single oncogenes or deleting tumor suppressors (such as Myc, β-catenin, or PTEN), TG221 uniquely combines metabolic injury with miRNA dysregulation. As a result, the mice develop steatohepatitic features and a miRNA expression profile that closely mirrors human HCC.
Finally, although hydrodynamic transposon models (SB/HTVI) can quickly generate tumors driven by defined sets of oncogenes, they generally lack the chronic liver damage seen in human disease. TG221, however, can model both spontaneous and injury-accelerated tumor development. Exposure to DEN or CCl_4_ produces strong synergy, leading to highly penetrant and faster-forming tumors.
Taken together, these characteristics position TG221 as a valuable middle ground between classical GEMs and highly variable chemical or dietary models, biologically relevant, miRNA-driven, immunocompetent, and metabolically altered system with strong translational potential.
This model also presents some important limitations [72,73]. In particular, TG221 tumorigenesis is driven by a single dominant oncomiR (miR-221), which may not fully capture the genetic complexity and multi-driver evolution that characterize human HCC. Moreover, although microRNA sequences are highly conserved across species, their basal expression levels, effective target repertoires and regulatory networks may differ between species (https://www.mirbase.org/) [74,75,76]. In addition, there are relevant species-specific differences in liver metabolism and immune responses that must be considered when translating findings from TG221 mice into clinical settings. Species-dependent variations in hepatic metabolism, drug clearance and immune regulation can significantly influence pharmacokinetics and the immune-mediated effects, in human hepatocarcinogenesis. For example, mice and humans express largely overlapping cytochrome P450 (CYP) enzyme families; however, the relative abundance of individual isoenzymes differs markedly between species. In addition, key enzymes such as CYP2E1 display distinct kinetic properties and regulatory mechanisms, while several isoforms (e.g., Cyp3a11, Cyp3a13) are mouse-specific. Consistent with these differences, mice exhibit faster drug metabolism and higher hepatic clearance, resulting in shorter drug half-lives. This explains why higher mg/kg doses are often required in murine models to achieve systemic exposures comparable to those observed in humans. Furthermore, the murine liver is more prone to rapid steatosis development, reflecting species-specific differences in lipid and glucose metabolism [77,78,79]. From an immunological perspective, mice display notable species-specific differences in hepatic immune responses, including higher basal activation of Kupffer cells and distinct expression patterns and sensitivities of Toll-like receptors (TLRs). These differences contribute to hepatic inflammation and tumor microenvironments that are not fully overlapping between mice and humans. Differences also extend to adaptive immunity, with substantial variation in T helper cell subsets (Th1/Th2/Th17) and in the expression and regulation of immune checkpoint molecules such as PD-1 and CTLA-4. Collectively, these factors can result in divergent responses to immuno-oncological therapies and underscore the limited predictive power of murine models for immunotherapy in HCC [80,81]. Recognizing these limitations may help to better contextualize the appropriate applications of the model while preserving its strengths.
Beyond their mechanistic role, miR-221 upregulation and miR-199a-3p loss represent clinically relevant expression patterns that may be leveraged as biomarkers for HCC risk stratification and patient selection. Meta-analytic and cohort-level evidence supports an association between elevated miR-221 and adverse prognosis [64,82], while reduced miR-199a-3p is repeatedly observed in HCC and aligns with tumor-suppressive pathway disruption. In the liquid biopsy setting, circulating miRNA panels that include miR-221 show moderate diagnostic value [83,84]. Finally, because these miRNAs map onto actionable signaling programs, miR-221 and/or miR-199a-3p profiling could support biomarker-guided trial enrichment for miRNA-targeted or rational combinations therapeutics [60,85]. Insights from TG221 studies could guide the identification of patient subgroups most likely to benefit from miRNA-targeted preventive or therapeutic strategies. Future work should further focus on preventive strategies in cirrhotic TG221 models and on combinatorial regimens targeting resistance pathways. Advances in delivery systems, including AAV vectors, innovative nanotechnologies approaches and polymeric carriers, will be critical for effective, liver-targeted modulation of pathogenic miRNA circuits.
Overall, TG221 represents a significant model for the preclinical study of miRNA-driven hepatocarcinogenesis and provides a robust bridge between molecular discovery and clinical translation.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bray F. Laversanne M. Sung H. Ferlay J. Siegel R.L. Soerjomataram I. Jemal A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries CA Cancer J. Clin.20247422926310.3322/caac.2183438572751 · doi ↗ · pubmed ↗
- 2Llovet J.M. Kelley R.K. Villanueva A. Singal A.G. Pikarsky E. Roayaie S. Lencioni R. Koike K. Zucman-Rossi J. Finn R.S. Hepatocellular carcinoma Nat. Rev. Dis. Primers 20217610.1038/s 41572-020-00240-333479224 · doi ↗ · pubmed ↗
- 3Cheng A.L. Qin S. Ikeda M. Galle P.R. Ducreux M. Kim T.Y. Lim H.Y. Kudo M. Breder V. Merle P. Updated efficacy and safety data from I Mbrave 150: Atezolizumab plus bevacizumab vs. sorafenib for unresectable hepatocellular carcinoma J. Hepatol.20227686287310.1016/j.jhep.2021.11.03034902530 · doi ↗ · pubmed ↗
- 4Llovet J.M. Ricci S. Mazzaferro V. Hilgard P. Gane E. Blanc J.F. de Oliveira A.C. Santoro A. Raoul J.L. Forner A. Sorafenib in advanced hepatocellular carcinoma N. Engl. J. Med.200835937839010.1056/NEJ Moa 070885718650514 · doi ↗ · pubmed ↗
- 5Kudo M. Finn R.S. Qin S. Han K.H. Ikeda K. Piscaglia F. Baron A. Park J.W. Han G. Jassem J. Lenvatinib versus sorafenib in first-line treatment of patients with unresectable hepatocellular carcinoma: A randomised phase 3 non-inferiority trial Lancet 20183911163117310.1016/S 0140-6736(18)30207-129433850 · doi ↗ · pubmed ↗
- 6Brown Z.J. Heinrich B. Greten T.F. Mouse models of hepatocellular carcinoma: An overview and highlights for immunotherapy research Nat. Rev. Gastroenterol. Hepatol.20181553655410.1038/s 41575-018-0033-629904153 · doi ↗ · pubmed ↗
- 7Li Y. Tang Z.Y. Hou J.X. Hepatocellular carcinoma: Insight from animal models Nat. Rev. Gastroenterol. Hepatol.20129324310.1038/nrgastro.2011.19622025031 · doi ↗ · pubmed ↗
- 8Heindryckx F. Colle I. Van Vlierberghe H. Experimental mouse models for hepatocellular carcinoma research Int. J. Exp. Pathol.20099036738610.1111/j.1365-2613.2009.00656.x 19659896 PMC 2741148 · doi ↗ · pubmed ↗