Discovery of a Human Metabolite That Mimics the Bacterial Quorum-Sensing Autoinducer AI‑2
Emilee E. Shine, Julie S. Valastyan, Vanessa Y. Ying, Jonathan Z. Huang, Mohammad R. Seyedsayamdost, Bonnie L. Bassler

TL;DR
Scientists discovered a human metabolite, l-xylosone, that mimics a bacterial signaling molecule called AI-2, which could help explain how human cells communicate with gut bacteria.
Contribution
The study identifies l-xylosone as a new human metabolite that mimics the bacterial autoinducer AI-2, expanding our understanding of host-microbe communication.
Findings
l-xylosone and l-xylulose were identified as AI-2 mimics using a reactivity-based metabolomics approach.
The l-configuration of xylosone is required for bacterial receptor recognition, similar to AI-2.
l-xylosone is a new addition to the human metabolome and suggests the existence of other reactive molecules in host-microbe interactions.
Abstract
Bacteria use small molecules to orchestrate collective behaviors in a process called quorum sensing (QS), which relies on the production, release, and group-wide detection of extracellular signal molecules referred to as autoinducers. One QS autoinducer, termed AI-2, is broadly used for interspecies bacterial communication, including in the mammalian gut. AI-2 consists of a family of interconverting compounds and adducts originating from 4,5-dihydroxy-2,3-pentanedione. This complex speciation, coupled with the inherent instability of AI-2 congeners, have complicated isolation efforts. It has been known that mammalian epithelial cells produce an AI-2 mimic to which bacteria respond. However, the identity of the AI-2 mimic has remained elusive, presumably due to its instability, similar to that of known AI-2 compounds. Here, we developed a reactivity-based metabolomics approach to capture…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 1
1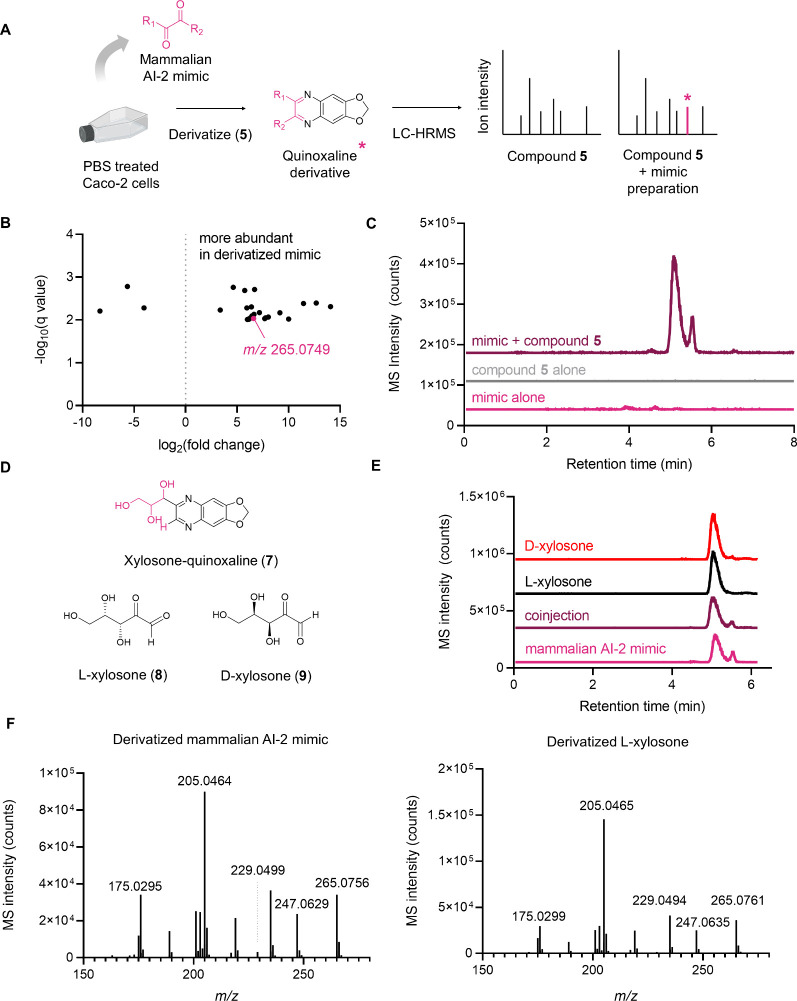 2
2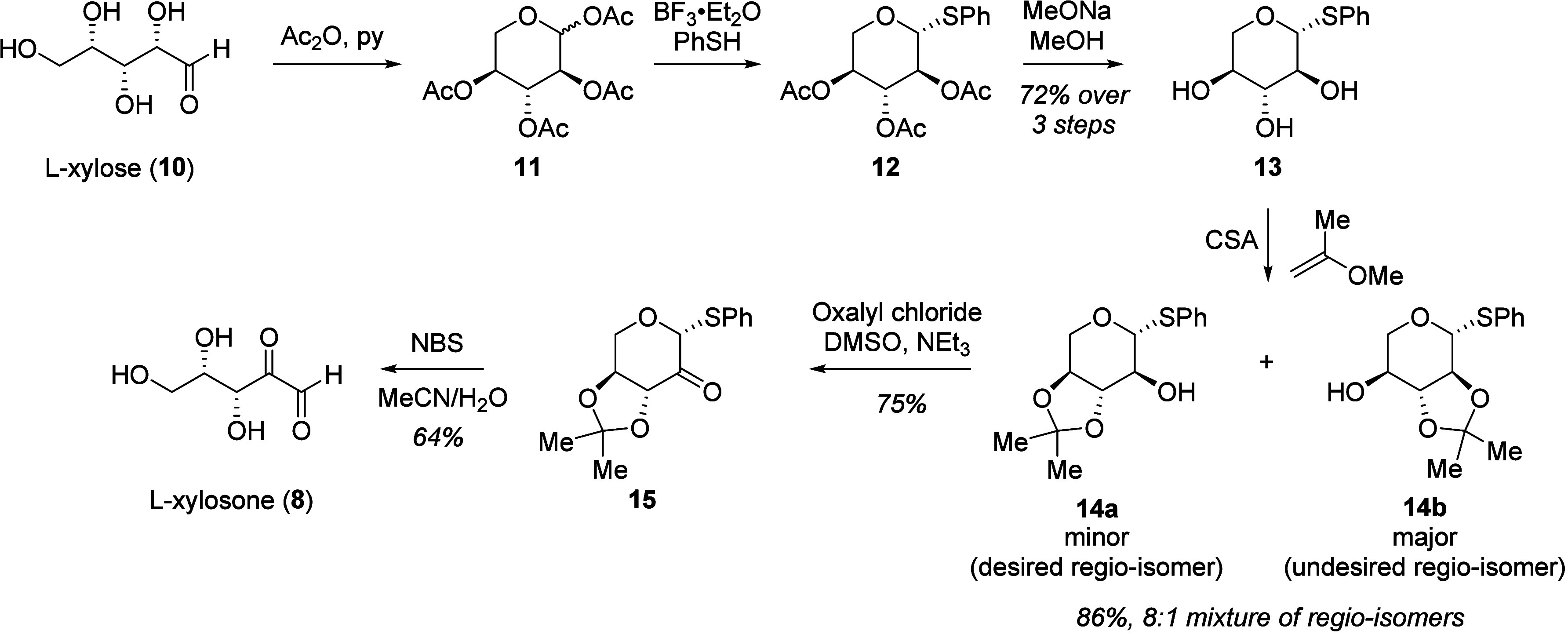 3
3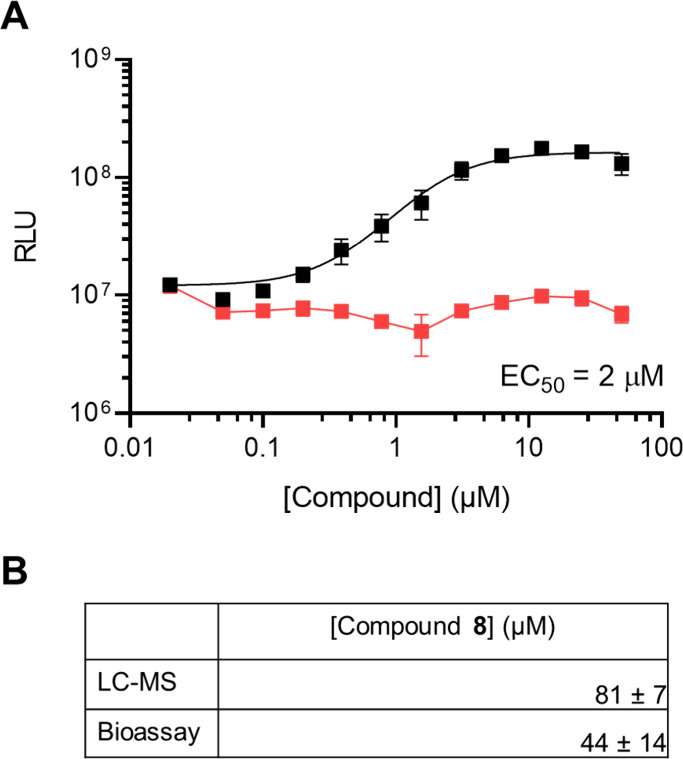 4
4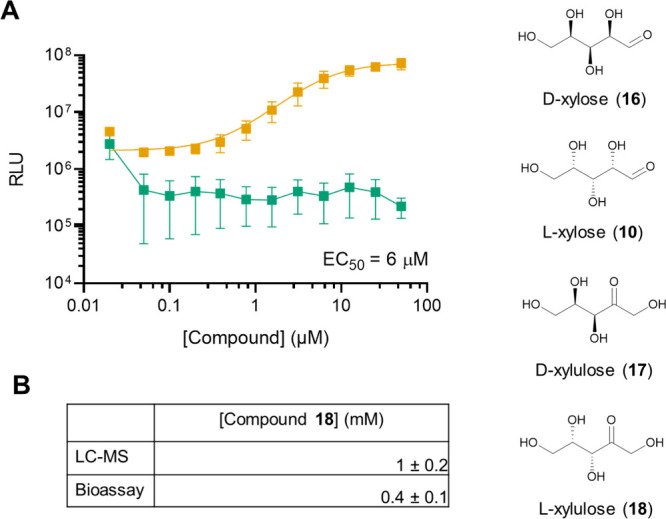 5
5- —Howard Hughes Medical Institute10.13039/100000011
- —Life Sciences Research Foundation10.13039/100009559
- —National Science Foundation (NSF)NA
- —National Institutes of Health (NIH)NA
- —National Institutes of Health (NIH)NA
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial biofilms and quorum sensing · Bacterial Genetics and Biotechnology · Cystic Fibrosis Research Advances
Introduction
The human gut microbiome consists of thousands of bacterial species that profoundly influence health and disease. ?−? ? Collectively, gut microbes possess far greater metabolic capacity than the host and synthesize a diverse array of metabolites, the identities of which continue to be uncovered. Some have been shown to drive microbe–microbe ?,? and host–microbe interactions, ?,? but the roles of many of these microbially derived metabolites remain to be defined. The molecular mechanisms underlying these chemically mediated interactions, which can be both beneficial or detrimental to human health, are another topic of intense interest. Microbiome metabolites are often challenging to identify in biological matrices due to low abundance ?,? or chemical instability. ?,?−? ? Advances in mass spectrometry have provided high-throughput routes to compound detection. ?−? ? ? Nonetheless, it remains challenging to assign structures to the majority of ions detected in metabolomics data sets. ?,? Connecting function and regulation to known metabolites provides a further hurdle in unraveling chemically mediated interactions in the microbiome.
Small molecules underpin the cell-to-cell communication process called quorum sensing (QS), which enables bacteria to orchestrate collective behaviors. QS involves production, release, accumulation, and group-wide detection of extracellular signal molecules called autoinducers. ?,? QS controls numerous collective behaviors in diverse bacteria, including bioluminescence, virulence factor production, antibiotic biosynthesis, and biofilm formation. There is increasing evidence that QS is fundamental to interactions occurring in the context of the human microbiome.?
QS autoinducers can be species-specific, meaning that a single bacterial species produces and detects a particular autoinducer. Other autoinducers are more “universal”; they are produced and detected by many bacterial species. Autoinducer-2 (AI-2) is among the latter class and is produced by the LuxS synthase, which is widely conserved in bacteria. ?−? ? AI-2 consists of a set of interconverting compounds derived from 4,5-dihydroxy-2,3-pentanedione (DPD, 1 in FigureA), an intermediate in the S-adenosylmethionine (SAM)-dependent methylation recycling pathway.? Following release of an activated methyl group from SAM to an acceptor substrate, the byproduct, S-adenosylhomocysteine (SAH), undergoes a two-step transformation to DPD: SAH is converted to S-ribosylhomocysteine (SRH) by the enzyme Pfs, followed by further processing by LuxS, yielding homocysteine and compound 1. In aqueous solution, compound 1 rapidly interconverts between linear, cyclic, and hydrated forms that coexist in equilibrium. ?−? ? ? The active AI-2 moiety that is recognized depends on the particular bacterial receptor, as well as the chemical environment. The boron-rich marine environment promotes formation of a boron adduct (2S,4S)-2-methyl-2,3,3,4-tetrahydroxytetrahydrofuran-borate (S-THMF-borate, 2), which is the active AI-2 recognized by the LuxP receptor in Vibrio spp.? In the absence of boron, compound 1 rearranges to (2R,4S)-2-methyl-2,3,3,4-tetrahydroxytetrahydrofuran (R-THMF, 3), which is the active AI-2 recognized by the LsrB receptor in enteric bacteria.? LsrB, first identified in Salmonella enterica ssp. typhimurium, is structurally similar to LuxP and occurs in Escherichia coli as well as some members of the Clostridiaceae and Bacillacaeae families. ?,? The ubiquitous pathogen, Pseudomonas aeruginosa, encodes a receptor with a dCache_1 domain that is reported to bind AI-2.? Over 1,500 transmembrane proteins harboring the dCache_1 domain are known among bacteria and archaea, providing opportunities for discovery of potentially new AI-2 structures and functions. ?,?
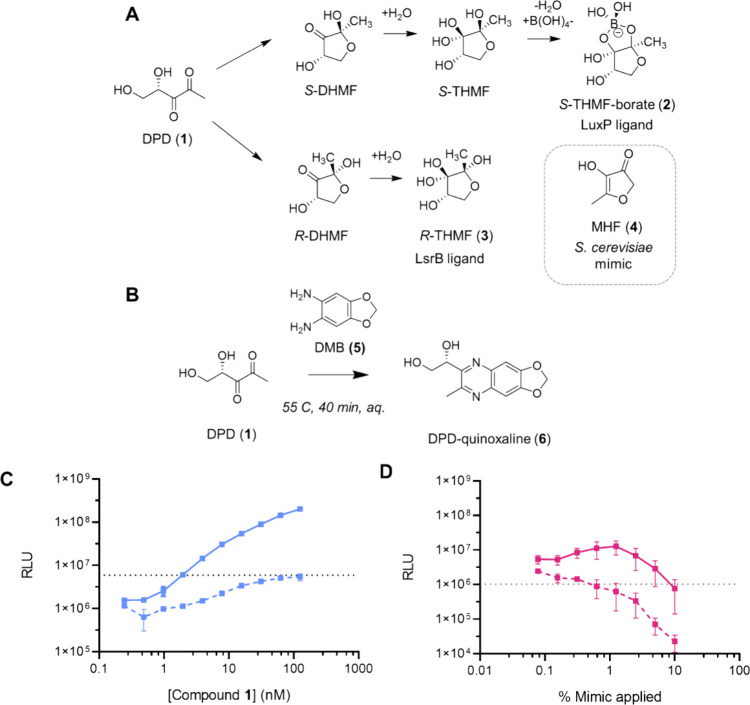
Derivatization of α-diketone compounds as a chemical screening strategy to identify the mammalian AI-2 mimic. A) Diagram showing the structure of the AI-2 precursor DPD (1), known interconverting AI-2 moieties (2) and (3), and the S. cerevisiae AI-2 mimic MHF (4). B) The o-diaminobenzene derivatization reagent (5) and the reaction scheme used for derivatizing compound 1 to form the functionalized quinoxaline product (6). C) Monitoring reaction completion by measuring the activity of compound 1 via light output from the V. harveyi TL-26 bioassay. Mixtures containing a 5 μM solution of compound 1 incubated with either control solution (solid line) or 3 mM compound 5 (dashed line) were applied in serial dilution to V. harveyi TL-26. The concentrations designated on the x-axis are relative to the initial concentration of compound 1 in the control reaction. D) Light output from V. harveyi TL-26 in response to 50 μL of culture fluids from Caco-2 cells treated 1:1 (v/v) with either control solution (solid line) or 3 mM compound 5 (dashed line). At high concentrations, both the control and reacted material cause declines in light output from the reporter, presumably due to the presence of inhibitors/toxic compounds. In C and D, RLU denotes relative light units, which are bioluminescence/OD600, the dotted lines show the baseline level bioluminescence in the assay, and error bars represent standard deviations of biological replicates, n = 3.
Many strains of gut-associated bacteria across the Bacteroidetes, Firmicutes, and Proteobacteria phyla possess the AI-2 biosynthetic enzyme LuxS. ?−? ? The yeast Saccharomyces cerevisiae produces an AI-2 mimic, 4-hydroxy-5-methylfuran-3(2H)-one (MHF, compound 4), that agonizes LuxP.? The yeast MHF-synthase Cff1p has homologues across archaeal, bacterial, and fungal genomes. The human host also participates in AI-2-driven communication. Specifically, in response to the secreted bacterial cytolytic toxin aerolysin, tight-junction disruption, or nutritional stress, mammalian intestinal epithelial cells produce an AI-2 mimic, i.e., a compound harboring AI-2 activity.? Prior to this report, the identity of the mammalian AI-2 mimic had not been reported, but the active compound appeared not to be one of the previously identified AI-2 structures for the following reasons: First, eukaryotic genomes do not possess luxS. Second, supplementation of SRH to mammalian cell culture did not promote increased AI-2 mimic production, indicating that epithelial cells cannot convert SRH to compound 1. Third, HPLC fractionation of epithelial cell culture fluids showed that the mammalian AI-2 mimic activity did not elute with compound 1, indicating that the compounds are not identical.
In this study, we developed a reactivity-based metabolomics workflow based on chemical derivatization that allowed us to capture a chemically unstable mammalian AI-2 mimic, thus enabling its identification and characterization. We identify the metabolite l-xylosone as an AI-2 mimic from human Caco-2 epithelial cells. We find that l-xylosone is recognized by the bacterial LuxP AI-2 receptor with micromolar affinity, whereas the d-enantiomer is not. Examination of compounds with structural features similar to l-xylosone revealed that l-xylulose, a primary metabolite in the glucuronate-xylulose pathway, also harbors AI-2 activity. Our identification of two mammalian-produced autoinducer mimics begins to define the chemical lexicon employed in cross-domain host-bacterial communication.
Results
Development of an α-Diketone-Based Derivatization Strategy
to Trap a Mammalian AI-2 Mimic
AI-2 has posed significant challenges for spectroscopic detection and characterization. As noted above, the AI-2 precursor (1) exists in an equilibrium mixture of linear and cyclic forms that undergo subsequent hydration events. ?−? ? ? There is no UV chromophore in the set of molecules comprising AI-2 and they ionize poorly using standard MS techniques. For these reasons, traditional activity-guided compound isolation did not reveal the structures of AI-2; rather, crystallization and structure elucidation of the LuxP-ligand and LsrB-ligand complexes provided the identities of the active AI-2s. ?,? Likewise, the mammalian AI-2 mimic did not yield to traditional purification and characterization methods, and its structure has remained unknown.
Previous analytical strategies to detect and quantify compound 1 in biological samples have hinged on derivatization, primarily relying on an *o-*diaminobenzene tag that reacts specifically with the α-diketone moiety of compound 1. ?−? ? ? The resulting quinoxaline product provides sensitive and reliable UV detection, as well as a specific m/z signature. We reasoned that the mammalian AI-2 mimic must be structurally similar to compound 1 and would also contain an α-diketone functional group. Leveraging this reactivity for tagging and detection could make the mammalian AI-2 mimic amenable to LC-MS-based metabolomics.
We evaluated multiple o-diaminobenzene containing derivatization agents and selected 1,2-diamino-4,5-methylenedioxybenzene (DMB, 5, FigureB) for our initial analyses as the derivatization reaction proceeds in complex aqueous biological matrices and because 5 is commercially available.? The reaction between compounds 1 and 5 was optimized, and quinoxaline product formation was monitored by LC-MS (compound 6, Figure S1). Reaction completion was assessed by applying the reaction mixture to a Vibrio harveyi strain that reports on AI-2 activity. This V. harveyi strain, called TL26, carries a luxS deletion (ΔluxS) and is therefore incapable of AI-2 production.? V. harveyi TL26 emits bioluminescence only when supplied with exogenous AI-2. Reacting compound 1 with excess compound 5 eliminated all AI-2 activity in the sample as judged by the V. harveyi TL26 bioassay (FigureC), suggesting that AI-2 had been completely derivatized under these conditions. The presence of compound 5 is not toxic, nor does it interfere with bioluminescence. Specifically, compared to 1 alone, addition of compounds 1 and 5 together at the start of the bioassay did not decrease V. harveyi TL26 light output (Figure S1).
Preparation of the mammalian AI-2 mimic for analyses followed a previously reported procedure. Briefly, medium from cultured Caco-2 cells was replaced with phosphate buffered saline (PBS), the cells were incubated for 48 h, and cell-free culture fluids harvested. Treatment of Caco-2 cells with PBS drives AI-2 mimic production while not affecting Caco-2 cell viability.? We reacted multiple such preparations of the mammalian AI-2 mimic with 5. When we applied the derivatization reaction mixtures to V. harveyi TL26, all bioassay activity was eliminated (FigureD). To confirm the selectivity of this reactivity-based chemical screen, we incubated compound 5 with MHF (compound 4), the yeast AI-2 mimic which, notably, lacks an α-diketo-moiety. No fluorescence emission indicative of a quinoxaline derivative was observed in reactions between compounds 4 and 5 (Figure S2).? When this reaction mixture was applied to V. harveyi TL26, light output was nearly identical to the untreated sample. Indeed, even a reaction with 100-fold molar excess of 5 did not eliminate activity of compound 4 in the bioassay (Figure S2). The reactivity profile of 5, coupled with its ability to inactivate mammalian AI-2 mimic activity, support our hypothesis that an α-diketone-containing compound primarily accounts for the mammalian AI-2 mimic activity.
Identification of Xylosone as a Mammalian AI-2 Mimic
We employed a reactivity-guided metabolomics screen to identify the mammalian AI-2 mimic (FigureA). Samples for our chemical screen were prepared as follows: Preparations of mammalian AI-2 mimic were incubated with compound 5 at 55 °C for 40 min and analyzed by LC-MS. As a control, compound 5 was likewise incubated in PBS followed by LC-MS analysis. The initial data set revealed 2,897 differentially abundant features (Figure S3). To identify biologically relevant signals, we applied stringent filters to limit our focus to metabolites with an LC-MS intensity threshold of 10,000, exhibiting at least a 10-fold increase compared to the control, and possessing q values ≤ 0.01 and m/z > 175, all features of compounds that could contain the quinoxaline core resulting from reaction with 5.
Comparative LC-MS analysis of mammalian AI-2 mimic samples derivatized with 5 identifies xylosone (8 and 9). A) Reactivity-based screening workflow. B) Volcano plot in which each data point represents a distinct molecular species detected by LC-MS. Features were filtered to include only those with m/z > 175, a minimum MS intensity threshold of 10,000, >10-fold changes compared to the control, and p values < 0.001. The observed ion for xylosone-quinoxaline (7, see panel D) is highlighted in pink (observed m/z 265.0749, calculated m/z 265.0819). C) Extracted ion chromatogram (EIC) spectra for m/z 265.0819 from mammalian AI-2 mimic samples treated with (maroon) or without (pink) 5, and of compound 5 alone (gray). D) Proposed chemical structure of the xylosone-quinoxaline product, compound 7 and l- and d-xylosone (compounds 8 and 9, respectively). E) EIC spectra (m/z 265.0819) of mammalian AI-2 mimic (pink), synthetic 8 (black), or synthetic 9 (red), each derivatized with 5. Also shown is a 1:1 v/v coinjection of 5-derivatized mammalian AI-2 mimic and compound 8 (maroon). F) MS/MS spectra (m/z 265.0819) of 5-derivatized AI-2 mimic and compound 8. Collision energy = 20 eV.
We mined the resulting 25 molecular features (Table S1) for predicted m/z corresponding to quinoxaline derivatives of previously reported α-diketone metabolites.? This strategy revealed a unique mass feature corresponding to m/z 265.0749 (FigureB), detected as two resolved peaks, one major (retention time = 5.1 min) and one minor (retention time = 5.5 min). This species was present in mammalian AI-2 mimic preparations derivatized by compound 5, but not in samples that had not been subjected to derivatization (FigureC). The molecular formula of the putative α-diketone-containing metabolite was determined to be C_5_H_8_O_5_, matching that of xylosone, a rare sugar that is new to human cells and can exist as l- or d-enantiomers (8 and 9, FigureD).
To verify xylosone as the active component and differentiate between the two enantiomers, we developed a synthetic route to obtain pure 8 and 9 (Figure). To obtain 8, we began with l-xylose (10), converted it to the acetyl-protected, cyclic form (11) and installed the (1R)-thioketal regio- and diastereoselectively by using boron trifluoride etherate and thiophenol (12).? The remaining acetate groups in 12 were then deprotected, followed by acetonide protection of vicinal diols in 13 that resulted in the formation of two regio-isomers 14b and 14a in an 8:1 ratio and an 86% combined yield.? The desired product 14a is the minor regio-isomer formed during the acetonide protection, which was separated from 14b and subjected to Swern oxidation using oxalyl chloride, DMSO, and triethylamine to furnish ketone 15 in 75% yield. Global deprotection of the thioketal and acetonide groups in 15 using N-bromosuccinimide afforded l-xylosone (8) in 64% yield. This route also allows for the preparation of d-xylosone (9) from d-xylose.
Synthetic scheme used to access enantiopure l- and d-xylosone. The scheme is shown for l-xylosone (8) starting with l-xylose (10), but the identical approach was used to obtain d-xylosone (9) starting from d-xylose. l-Xylose (10) is first converted into 11 using a combination of acetic anhydride (Ac2O) and pyridine (py). The thioketal in 12 was then installed regio- and diastereoselectively by activating the anomeric acetate group in 11 using boron trifluoride etherate (BF3·Et2O) and thiophenol (PhSH). Deprotection with sodium methoxide (MeONa) in methanol yielded 13. The vicinal diols in 13 were then acetonide-protected using camphor sulfonic acid (CSA), resulting in the formation of two regio-isomers 14b and 14a in 8:1 ratio and 86% combined yield. The desired product 14a is the minor regio-isomer formed during the acetonide protection, which was separated from 14b and subjected to Swern oxidation conditions using oxalyl chloride, DMSO and triethylamine (NEt3) to furnish ketone 15 in 75% yield. Global deprotection of the thioketal and acetonide groups in using N-bromosuccinimide (NBS) in acetonitrile/water afforded l-xylosone (8) in 64% yield.
With the two enantiomers in hand, we prepared the quinoxaline derivatives of synthetic l-xylosone and d-xylosone, which exhibited identical chromatographic properties and were not separable by HPLC with a retention time of 5.1 min. Importantly, this elution profile matched the retention time of the derivatized mammalian AI-2 mimic (FigureE). Coinjection of the derivatized synthetic material and the mammalian AI-2 mimic also showed identical retention times (FigureE). Finally, MS/MS analysis confirmed that the derivatized synthetic material was indistinguishable from mammalian-derived xylosone-quinoxaline using a range of collision energies (20 to 50 eV) (FigureF, Figure S4, Tables S2, S3). These data verify xylosone as an active AI-2 mimic.
Verification that l-Xylosone Possesses AI-2 Activity
We next assessed each xylosone enantiomer for AI-2 mimic activity using the V. harveyi TL26 bioassay (see Figures S5–S9 for quantitation methods).? l-xylosone (8) was active with a half-maximal effective concentration (EC_50_) of ∼2 μM, while d-xylosone (9) was inactive (FigureA, Figure S16). The stereochemical selectivity for the l-xylosone enantiomer aligns with prior findings from the crystal structure of the LuxP-AI-2 complex.? The hydroxyl group on the C-4 of 2 forms two hydrogen bonds with LuxP residues Trp82 and Gln77. Inversion from the S- to the *R-*configuration renders the hydroxyl group inaccessible for hydrogen bonding.? Compound 8 retains the C-4 configuration that is required for binding to the LuxP receptor. The human intestinal pathogen and QS bacterium Vibrio cholerae detects AI-2 to control traits including virulence and biofilm formation. ?−? ? ? A V. cholerae AI-2 reporter strain? responded to exogenous compound 8 with an EC_50_ of ∼8.3 μM, comparable to the measured EC_50_ of ∼1 μM for the native ligand, compound 1 (Figure S10).
l-Xylosone (8) has AI-2 activity, while d-xylosone (9) does not. A) Light output from the V. harveyi TL-26 bioassay in response to indicated amounts of 8 (black) and 9 (red). The EC50 for 8 is shown. Error bars represent standard deviations of technical replicates, n = 3. B) Quantitation of 8 from Caco-2 AI-2 mimic preparations from 2,000,000 cells grown for 2 days. Concentrations are calculated from LC-MS measurements and the V. harveyi TL-26 bioassay. Data are reported as averages ± standard deviations of biological replicates, n = 3.
Analogous to other AI-2s, compound 8 likely cyclizes to form a furanose that mimics compound 2 in the binding pocket of LuxP. Indeed, the ^13^C NMR spectrum of compound 8 displays 6 major signals suggestive of the formation of at least 3 acetal and/or hemiacetal moieties of the α-ketoaldehyde in xylosone (Figure S11). We conducted boron binding studies with 8 with the rationale that cyclization and subsequent hydration of the C-3 carbonyl of 2 could lead to formation of a borate diester that drives maximal AI-2 activity in the V. harveyi bioassay. ?−? ? Similarly, addition of boric acid to the medium is required to achieve high potency of compound 8 in the bioassay (Figure S12A, B), supporting the notion that l-xylosone complexes with borate across a cis-diol. Borate esters of 1,3-diols and 1,2-diols display characteristic chemical shifts, allowing distinction between these moieties.? Surprisingly, however, no ^11^B or ^13^C NMR signals indicative of borate-diester formation were detected when borate was incubated with compound 8 (Figures S13, S14). It is possible that formation of the borate diester complex of 8 only occurs in the LuxP binding pocket.
We used two methods to quantify the amount of xylosone present in mammalian culture fluids. First, using the V. harveyi TL26 bioassay, we estimated concentrations based on measurements of activity from known quantities of pure 8. Second, mammalian AI-2 mimic samples were derivatized with 5; then, using LC-MS we determined the concentration from a standard curve generated from known amounts of 8 derivatized with 5. The two methods yielded similar inferred concentrations: After 2 days of growth in PBS, Caco-2 cells produced 44 ± 14 μM and 81 ± 7 μM AI-2 mimic as determined by bioassay and LC-MS, respectively (Figure). For comparison, recent GC-MS based quantitation of compound 1 levels in the cecal contents of specific pathogen free mice ranged from 0.07–0.21 μM,? consistent with reported K_d_ values for LsrB receptors from various bacterial species.? Together, these data confirm l-xylosone as the active AI-2 mimic and provide production titers that appear physiologically relevant and significantly higher than the EC_50_ values determined in the V. harveyi TL26 and V. cholerae bioassays (Figure).
l-Xylulose, a Compound Structurally Related to l-Xylosone Possesses AI-2 Activity
We tested two commercially available compounds, xylose and xylulose, with structural features similar to compound 8 for AI-2 mimic activity. These compounds differ from 8 only in the oxidation state of a single carbon atom. Neither the naturally occurring d-xylose (16) nor l-xylose (10) possessed detectable AI-2 activity (Figure S15). Likewise, d-xylulose (17) is also inactive, consistent with prior reports (FigureA).? By contrast, l-xylulose (18) was active, with an EC_50_ ∼ 6.0 μM (FigureA, Figure S17). l- and d-xylulose are naturally occurring sugars generated as intermediates in the glucuronate-xylulose pathway leading to d-xylulose-5-phosphate. Analogous to other LuxP ligands, addition of boric acid to the medium is required for optimal activity (Figure S12C) and a small amount of complex could be detected by ^11^B NMR (Figure S16 ; chemical shift δ 10.6 ppm), consistent with previously reported values.? With respect to detection by the V. harveyi QS apparatus, compound 1 is most potent, followed by compounds 8 and then 18 (Figure S17).
Structurally related ketopentose sugar l-xylulose (18) possesses AI-2 activity. A) Light output from the V. harveyi TL-26 bioassay in response to indicated amounts of 17 (green) and 18 (orange). The EC50 for 18 is provided. Error bars represent standard deviations of technical replicates, n = 3. B) Quantitation of 18 levels in Caco-2 AI-2 mimic preparations from 2,000,000 cells grown for 2 days. Concentrations are calculated from LC-MS measurements and the V. harveyi TL-26 bioassay. Data are reported as averages ± standard deviations of biological replicates, n = 3.
To determine the production titers of 18 by Caco-2 cells under our experimental conditions, we derivatized the cell-free fluids with N,N-diphenylhydrazine and used LC-MS to quantify the hydrazone derivative (FigureB).? We also estimated the concentration of compound 18 present in mammalian mimic preparations by comparison to the V. harveyi TL26 bioassay activity generated from known amounts of compound 18. These assays yielded a concentration of 0.4 ± 0.1 mM (bioassay) and 1 ± 0.2 mM (LC-MS). We suspect that these values are overestimates because the bioassay output will be the sum of activity from 8 and 18 and our LC-MS method does not distinguish between the hydrazone derivative of xylulose and the isobaric hydrazone derivative of xylose.
Discussion
The microbiome-mucosal interface is a crucial site for regulation of intestinal function. On the bacterial side, microbial products such as short-chain fatty acids,? secondary bile acids, ?,? and tryptophan metabolites ?,? function to maintain gut epithelial integrity. On the host side, intestinal epithelial cells detect pathogen-associated molecular patterns, including flagellin and lipopolysaccharide, on the outer surfaces of bacterial cells and they relay the garnered information to intestinal immune cells to launch defense mechanisms and drive immune tolerance.? Mammalian metabolites, including antimicrobial peptides, cytokines, and secreted IgA antibodies shape microbiome composition.? Adding to these bidirectional chemical interactions, we previously reported a mammalian AI-2 mimic of unknown structure that is detected by bacterial cells.? Here, our studies reveal that the new human metabolite l-xylosone functions as a QS autoinducer to which bacteria respond. Prior to this report, the only known role of xylosone was its involvement in nonenzymatic modifications of proteins to form advanced glycation end products (AGEs). ?−? ? Our work demonstrates that l-xylosone functions as a QS signal that bacteria detect; we do not yet know whether l-xylosone has additional metabolic and/or signaling roles affecting the mammalian host. Given that activation of QS in V. cholerae results in dispersal, production of xylosone by mammalian cells could clear the pathogen. The concentration of xylosone that we measure in cell culture and its EC_50_ suggest that this function can be physiologically relevant.
The most highly studied in vitro route to xylosone formation is from the spontaneous, oxidative degradation of ascorbic acid to dehydroascorbic acid. ?,? Under neutral or alkaline conditions, dehydroascorbic acid undergoes hydrolytic cleavage of the lactone ring to yield 2,3 diketo-l-gluconic acid, which subsequently decarboxylates to l-xylosone, which is highly reactive. Humans are incapable of synthesizing ascorbic acid and therefore obtain it from diet. Although often a common component of mammalian cell culture medium, we note that ascorbic acid is not present in media used here. Indeed, we were unable to detect ascorbic acid in our media nor in mammalian AI-2 mimic preparations using LC-MS. Thus, l-xylosone must originate from a nonascorbic acid route. Some eukaryotes, such as wood-degrading fungi, harbor pyranose 2-oxidase (P2Ox) which can produce xylosone.? P2Ox enzymes are flavoenzymes that, in the presence of oxygen, catalyze the regioselective C-2 oxidation of d-glucose, d-xylose, and various other mono- and disaccharides to yield the corresponding dicarbonyl sugars and hydrogen peroxide. ?−? ? P2Ox enzymes belong to the glucose-methanol-choline oxidoreductase superfamily.? A search of the InterPro database reveals two mammalian genes encoding proteins belonging to this class of enzymes: CHDH, encoding a choline dehydrogenase, and B4DMQ4, encoding a gene of unknown function similar to CHDH. Conceivably, one or both of these enzymes could catalyze C-1 oxidation of l-xylulose to produce l-xylosone. Lastly, it is also possible that l-xylosone production occurs spontaneously as is the case for other dicarbonyl compounds that are formed through dehydration and enolization of monosaccharides.
We also demonstrated that l-xylulose has AI-2 mimic activity. More is known about l-xylulose than l-xylosone in human metabolism. l-xylulose is a minor sugar produced in the glucuronate degradation pathway, an alternate pathway for glucose-6-phosphate oxidation.? Specifically, l-xylulose is produced by decarboxylation of β-keto-l-gulonate by the C11orf54 protein.? Next, l-xylulose is reduced to xylitol in a reversable reaction catalyzed by the dicarbonyl/l-xylulose reductase DCXR. Mutations in DCXR result in excretion of l-xylulose, the hallmark of the metabolic disorder called pentosuria.? DCXR also functions as a diacetyl reductase that detoxifies highly reactive α-dicarbonyl compounds. ?,? Indeed, DCXR deficient mice are more vulnerable to protein damage following diacetyl-induced cytotoxicity than are mice that are wildtype for DCXR.? One possibility is that alterations in DCXR activity, triggered by starvation or tight junction disruption, decrease DCXR diacetyl reductase activity and drive the accumulation and secretion of l-xylulose and l-xylosone.
We cannot exclude the possibility of additional mammalian AI-2 mimics, as our study specifically targeted α-diketone containing compounds. Indeed, l-xylulose is not derivatized in our chemical trapping strategy so it was not identified in our data set, yet we show it has AI-2 mimic activity. To our knowledge, there is no Caco-2 cell line incapable of producing l-xylulose and l-xylosone, which limits our ability to probe whether multiple AI-2 mimics exist. Other AI-2 mimics could also be produced when human cells are cultured under conditions that differ from those employed here. In this work, we induced epithelial cells to produce l-xylosone using nutrient deprivation stress. Earlier, we showed that AI-2 mimic production occurs following epithelial cell exposure to toxins or tight junction disruption. In those cases, we have not confirmed that l-xylosone or l-xylulose is the compound produced that harbors the AI-2 mimic activity.
Our findings highlight the utility of reactivity-guided metabolomics approaches to capture chemically unstable metabolites of interest. Reactivity-based screening approaches have been previously used to target labile functional groups such as α,β-unsaturated carbonyls, ?,? isonitriles,? epoxides,? and diazo? groups. Using our approach to discover α-diketone containing compounds, we detected the rare metabolite l-xylosone, which was not previously identified in the human metabolome database. Mammalian α-diketone containing compounds of intense research focus have historically centered on methylglyoxal, glyoxal, and 3-deoxyglucosone for their roles in the formation of AGEs in aging tissues. ?,?−? ? We speculate that there other highly reactive α-diketone containing compounds exist that are likewise absent from existing metabolomic data sets that possess novel and potentially fascinating biological roles.
Experimental Methods
Mammalian AI-2 Mimic Production and Preparation for Analysis
Caco-2 cells were grown at 37 °C and 5% CO_2_. For maintenance, Caco-2 cells were grown in 1X DMEM (Gibco), 20% FBS (Corning), 1X Penstrep (Sigma), and 1X Plasmocin prophylactic (Invivogen). For mammalian AI-2 mimic production, Caco-2 cells were grown to confluence, washed twice with 1X Dulbecco’s PBS (Gibco), and detached from tissue culture plates using trypsin-EDTA (Corning). These cells were collected, washed once with maintenance medium to inactivate the trypsin, washed twice with 1X PBS, and subsequently placed in 5 mL of 1X PBS at a cell density of 2,000,000 cells/mL for 48 h. Cell-free culture fluids were harvested after centrifugation at 1,500 rpm for 5 min followed by filtration through a 0.22 μm filter. We call these samples mammalian AI-2 mimic preparations throughout this work. These preparations were stored at 4 °C prior to use.
AI-2 Bioassay
V. harveyi TL-26 was grown overnight in LM medium at 30 °C and diluted 1:1000 into AB medium supplemented with 0.1 mM boric acid. V. cholerae AB_Vc_542 (ΔcqsS ΔluxQ ΔvpsS ΔcqsR Δvc1807::PluxC-luxCDABE::Spec^R^) was grown overnight in LB medium at 30 °C and diluted 1:5000 into LB medium supplemented with 0.1 mM boric acid. The diluted cultures were aliquoted into wells of black-sided, clear-bottom 96-well plates (Corning). All compounds, derivatization reaction mixtures, and mammalian AI-2 mimic preparations were added at the indicated amounts. Plates were incubated at 30 °C with shaking for 6 h, followed by bioluminescence measurements using an Envision plate reader (PerkinElmer).
General LC-MS Methods
Low resolution data for optimization of compound 5 derivatizations were acquired using an Agilent MSD iQ System coupled to an Agilent 1290 Infinity II HPLC. Samples were separated on an Agilent Eclipse C18 1.8 μm (2.1 mm × 50 mm) column with an injection size of 2 μL. The mobile phase was a water–acetonitrile (MeCN) gradient containing 0.1% formic acid. Chromatography was performed as follows: 0–2 min 5% MeCN, 2–10 min 5–95% MeCN, 10–12 min 95% MeCN. The LC-MS iQ was carried out in positive mode scanning between m/z 100–1000. Source parameters for LC-MS acquisition were as follows: gas temperature 325 °C, gas flow 11 (L/min), capillary voltage 3500 V. Data were processed using Agilent OpenLAB CDS. Peaks were extracted by m/z, quantified by area under the curve. High resolution LC-MS data for the metabolomics workflow and quantitation of compounds 8 and 18 in mammalian AI-2 mimic samples were acquired on an Agilent 6546 LC-QTOF 1290 LC system. Samples were separated on a Phenomenex Polar RP 2.5 μm (100 mm × 3 mm) column with an injection size of 5 μL. The mobile phase was a water-MeCN gradient containing 0.1% formic acid. Chromatography was performed as follows: 0–2 min 5% MeCN, 2–10 min 5–95% MeCN, 10–11 min 95% MeCN, equilibrate to 5% MeCN. MS1 acquisition was carried out in positive mode scanning from 100 to 500 m/z. MS2 acquisition was carried out in positive mode scanning from 100 to 600 m/z with fixed collision energies at 20, 25, 55 eV. The MS1 scan rate was 3 spectra/sec and the MS/MS scan rate was 1 spectra/sec. Source parameters for both MS1 and MS2 acquisition were as follows: gas temperature 275 °C, gas flow 12 (L/min), capillary voltage 3500 V. Data were processed using Agilent MassHunter Qualitative Analysis 10.0. Peaks were extracted by m/z within a 20 ppm error window. Metabolites were quantified by peak integration and area under the curve.
Derivatization Reactions with Compound 5
Derivatization reagent solutions were prepared immediately prior to each experiment by dissolving 4 μmol of 4,5-methylenedioxy-1,2-phenylenediamine dihydrochloride (Sigma-Aldrich) and 56 μmol of sodium dithionite (Fisher Chemical) into 4 mL of deionized water to yield a final stock solution of 1 mM compound 5. This derivatization reagent solution was diluted to desired concentrations in control solution (14 mM sodium dithionite in water). To perform derivatization reactions, the following mixtures were prepared in screw capped glass vials: 1 mL of derivatization reagent solution and 1 mL of compound 1 (Jubilant Pharma) or 1 mL of derivatization reagent solution and 1 mL of compound 4 (AK Scientific), each diluted to the concentrations designated in figure legends. For mammalian AI-2 mimic derivatization reactions, 50 μL of samples prepared as above were combined with 50 μL of a 7 mM solution of compound 5. In all cases, mixtures were incubated at 55 °C for 40 min. Control reactions were prepared identically except that the derivatization reagent solution was replaced by 1 mL of deionized water. Reactions were terminated by cooling on ice for 5 min. To measure the quinoxaline products, aliquots of reactions were placed into wells of black-sided, clear-bottom 96-well plates (Corning) and fluorescence intensity was measured in a Synergy plate-reader (BioTek) (excitation/emission, 355 nm/393 nm).
Reactivity Guided Metabolomics
Derivatization reagent solutions were prepared immediately prior to each experiment by dissolving 12.6 μmol of 4,5-methylenedioxy-1,2-phenylenediamine dihydrochloride (Sigma-Aldrich) and 25.2 μmol of sodium dithionite (Fisher Chemical) into 1.8 mL of deionized water to yield a final stock solution of 7 mM compound 5. To perform derivatization reactions, screw capped glass vials containing 100 μL of compound 5 solution and 100 μL of mammalian AI-2 mimic preparations or 100 μL of 1X PBS were incubated at 55 °C for 40 min, cooled in ice for 5 min, and subjected to centrifugation at 13,000 rpm for 5 min to remove insoluble material. High resolution LC-MS analyses were performed using the parameters described above. Following LC-MS acquisition, Agilent.d files were converted into.mzXML files using MSCovertGUI (ProteoWizard). Data in the converted files were analyzed using? XCMS Online (Scripps Research Institute) in pairwise comparisons using the UPLC/UHD Q-TOF parameters. The peak picking algorithm was centWave, with significant features identified using unpaired parametric t-test (Welch t test) and a p-value threshold of 0.05. To identify highly significant molecular features, we sorted and filtered the data using the calculated q value, which provides the false discovery rate in multiple hypothesis testing.
NMR Methods
Nuclear magnetic resonance (NMR) spectra were acquired at the Princeton University Department of Chemistry Facilities. ^1^H, ^13^C and ^11^B NMR spectra were collected in the triple resonance cryoprobe of a Bruker Avance III 500 MHz NMR spectrometer, and were calibrated using residual undeuterated solvent as an internal reference (H_2_O: ^1^H NMR = 4.79; CDCl_3_: ^1^H NMR = 7.26, ^13^C NMR = 77.16; acetone-D_6_: ^1^H NMR = 2.05, ^13^C NMR = 29.84; CD_3_OD: ^1^H NMR = 3.31, ^13^C NMR = 49.00). ^1^H NMR spectra were tabulated as follows: chemical shift, multiplicity (s = singlet, d = doublet, t = triplet, q = quartet, p = pentet, dd = doublet of doublets, dt = doublet of triplets, m = multiplet, br = broad), coupling constant (Hz), and number of protons. ^13^C NMR spectra were tabulated by observed peak, and no special nomenclature is used for equivalent carbons. All NMR data were analyzed with MestReNova software.
LC-MS Quantitation of Compounds 8 and 18 in Mammalian AI-2 Mimic Samples
Mammalian AI-2 mimic samples were prepared by splitting cells into seeding densities of 2,000,000 cells/mL and incubating them in 5 mL of 1X PBS for 48 h followed by harvest. For LC-MS quantitation of compound 8, derivatization reactions were prepared using 100 μL of AI-2 mammalian mimic preparations mixed with 100 μL of a 7 mM solution of compound 5. The reactions were incubated at 55 °C for 40 min, cooled on ice for 5 min, and stored at −20 °C until LC-MS analysis. A standard curve was prepared by mixing 100 μL solutions (100 nM-1 mM) of compound 8 with a 7 mM solution of compound 5. For quantitation of compound 18, 400 μL of AI-2 mammalian mimic preparations were mixed with a 200 μL solution of 0.1 M N,N-diphenylhydrazine and 400 μL of MeOH. The N,N-diphenylhydrazine solution was prepared immediately prior to use by dissolving 1 mmol of diphenylhydrazine hydrochloride and 3 mmol of triethylamine in 10 mL water/MeCN (1:1, v/v). N,N-diphenylhydrazine derivatization reactions were allowed to stand for 24 h at room temperature prior to LC-MS analysis. Subsequently, samples were dried in vacuo (Genevac HT6 S3i Evaporator) followed by resuspension in 100 μL of MeOH for LC-MS analysis. N,N-Diphenylhydrazine was chosen as the derivatizing agent to transform the analyte into a xylulose-hydrazone for optimized LC-MS detection in positive mode. Concentrations of compound 18 were estimated from a standard curve generated from derivatizations of 400 μL compound 18 solutions (50 μM-10 mM) with a 0.1 M N,N-diphenylhydrazine solution.
Supplementary Material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Fan Y.Pedersen O.Gut Microbiota in Human Metabolic Health and Disease Nat. Rev. Microbiol 202119557110.1038/s 41579-020-0433-932887946 · doi ↗ · pubmed ↗
- 2Wirbel J.Pyl P. T.Kartal E.Zych K.Kashani A.Milanese A.Fleck J. S.Voigt A. Y.Palleja A.Ponnudurai R.Sunagawa S.Coelho L. P.Schrotz-King P.Vogtmann E.Habermann N.Niméus E.Thomas A. M.Manghi P.Gandini S.Serrano D.Mizutani S.Shiroma H.Shiba S.Shibata T.Yachida S.Yamada T.Waldron L.Naccarati A.Segata N.Sinha R.Ulrich C. M.Brenner H.Arumugam M.Bork P.Zeller G.Meta-Analysis of Fecal Metagenomes Reveals Global Microbial Signatures That Are Specific for Colorectal Cancer Nat. Med.20192567968910.1038/s 41591-019-0406-630936547 PMC 7984229 · doi ↗ · pubmed ↗
- 3Lloyd-Price J.Arze C.Ananthakrishnan A. N.Schirmer M.Avila-Pacheco J.Poon T. W.Andrews E.Ajami N. J.Bonham K. S.Brislawn C. J.Casero D.Courtney H.Gonzalez A.Graeber T. G.Hall A. B.Lake K.Landers C. J.Mallick H.Plichta D. R.Prasad M.Rahnavard G.Sauk J.Shungin D.Vázquez-Baeza Y.White R. A.Bishai J.Bullock K.Deik A.Dennis C.Kaplan J. L.Khalili H.Mc Iver L. J.Moran C. J.Nguyen L.Pierce K. A.Schwager R.Sirota-Madi A.Stevens B. W.Tan W.ten Hoeve J. J.Weingart G.Wilson R. G.Yajnik V.Braun J.Denson L. A.Jansson J. K.Knight R.Kugathasan S.Mc Govern D. P. · doi ↗ · pubmed ↗
- 4Zipperer A.Konnerth M. C.Laux C.Berscheid A.Janek D.Weidenmaier C.Burian M.Schilling N. A.Slavetinsky C.Marschal M.Willmann M.Kalbacher H.Schittek B.Brötz-Oesterhelt H.Grond S.Peschel A.Krismer B.Human Commensals Producing a Novel Antibiotic Impair Pathogen Colonization Nature 2016535511610.1038/nature 1863427466123 · doi ↗ · pubmed ↗
- 5Torres Salazar B. O.Dema T.Schilling N. A.Janek D.Bornikoel J.Berscheid A.Elsherbini A. M. A.Krauss S.Jaag S. J.Lämmerhofer M.Li M.Alqahtani N.Horsburgh M. J.Weber T.Beltrán-Beleña J. M.Brötz-Oesterhelt H.Grond S.Krismer B.Peschel A.Commensal Production of a Broad-Spectrum and Short-Lived Antimicrobial Peptide Polyene Eliminates Nasal Staphylococcus aureus Nat. Microbiol.2024920021310.1038/s 41564-023-01544-238110697 PMC 11310079 · doi ↗ · pubmed ↗
- 6Cohen L. J.Esterhazy D.Kim S. H.Lemetre C.Aguilar R. R.Gordon E. A.Pickard A. J.Cross J. R.Emiliano A. B.Han S. M.Chu J.Vila-Farres X.Kaplitt J.Rogoz A.Calle P. Y.Hunter C.Bitok J. K.Brady S. F.Commensal Bacteria Make GPCR Ligands That Mimic Human Signalling Molecules Nature 2017549485310.1038/nature 2387428854168 PMC 5777231 · doi ↗ · pubmed ↗
- 7Cao Y.Oh J.Xue M.Huh W. J.Wang J.Gonzalez-Hernandez J. A.Rice T. A.Martin A. L.Song D.Crawford J. M.Herzon S. B.Palm N. W.Commensal Microbiota From Patients with Inflammatory Bowel Disease Produce Genotoxic Metabolites Science 2022378 eabm 323310.1126/science.abm 323336302024 PMC 9993714 · doi ↗ · pubmed ↗
- 8Lee R.Ptolemy A. S.Niewczas L.Britz-Mc Kibbin P.Integrative Metabolomics for Characterizing Unknown Low-Abundance Metabolites by Capillary Electrophoresis-Mass Spectrometry with Computer Simulations Anal. Chem.20077940341510.1021/ac 061780 i 17222002 · doi ↗ · pubmed ↗