Hypermethylation-mediated silencing of RASD1 drives multiple myeloma pathogenesis
Chenfeng Yi, Nana Ren, Yuxi Cai, Jiajia Zhang, Yonghuai Feng

TL;DR
This study finds that RASD1 is silenced in multiple myeloma due to hypermethylation, linking this to worse outcomes and suggesting RASD1 acts as a tumor suppressor.
Contribution
The novel contribution is identifying RASD1 as a tumor suppressor silenced by hypermethylation in multiple myeloma.
Findings
RASD1 mRNA and protein are significantly downregulated in multiple myeloma patients.
RASD1 promoter hypermethylation is associated with adverse clinical features in MM.
Demethylating agents restore RASD1 expression and increase apoptosis in MM cells.
Abstract
The role of Ras-related dexamethasone-induced 1 (RASD1) in multiple myeloma (MM) pathogenesis remains unclear. This study investigated the expression profile, clinical significance, and epigenetic regulation of RASD1 in MM. Bone marrow samples were collected from 26 newly diagnosed patients with MM and 8 healthy controls. RASD1 messenger RNA (mRNA) and protein expression were analyzed using reverse transcription quantitative polymerase chain reaction (RT-qPCR) and immunohistochemistry, respectively. DNA methylation status was assessed via methylation-specific PCR (MSP). The U266 MM cell line was treated with the demethylating agent decitabine (DAC) to evaluate its effects on RASD1 expression and apoptosis. RASD1 mRNA and protein expression were significantly downregulated in patients with MM compared to healthy controls (P < 0.001). Low RASD1 mRNA levels correlated significantly with…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
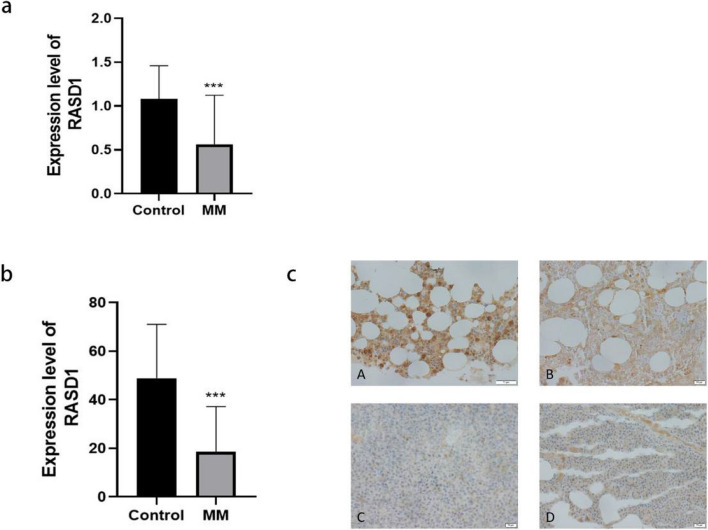 Figure 1
Figure 1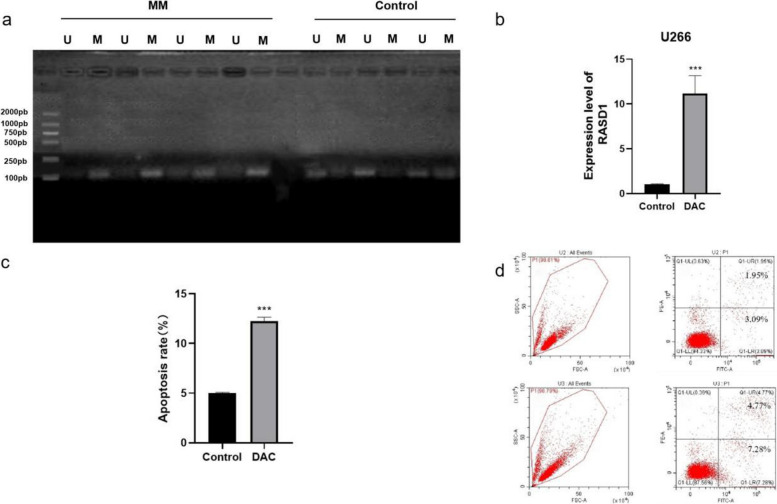 Figure 2
Figure 2- —Guizhou Science and Technology Department ZK [2021-406].
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEpigenetics and DNA Methylation · Multiple Myeloma Research and Treatments · PI3K/AKT/mTOR signaling in cancer
Introduction
Multiple myeloma (MM) is a malignant neoplasm of clonal plasma cells and represents the second most common hematological malignancy [1]. Despite therapeutic advances, including the introduction of novel agents and stem cell transplantation, MM remains largely incurable due to the inevitable development of relapsed or refractory disease [2]. A deeper understanding of its molecular pathogenesis is crucial for identifying new therapeutic targets.
Ras-related dexamethasone-induced 1 (RASD1) is a member of the Ras superfamily of small G-proteins. Unlike its well-characterized oncogenic counterparts (e.g., K-Ras, H-Ras), RASD1 exhibits context-dependent roles in cancer and can function as a tumor suppressor, exerting growth-inhibitory effects in lung adenocarcinoma and breast cancer cells [3, 4]. Conversely, it may promote progression in malignancies such as osteosarcoma [5]. Evidence regarding the role of RASD1 in hematological cancers is limited and contradictory. Some studies suggest that RASD1 suppresses B-cell proliferation [6], whereas others indicate that it promotes growth in B-cell acute lymphoblastic leukemia [7].
Epigenetic silencing via promoter hypermethylation represents a key mechanism for inactivating tumor suppressor genes. In MM, Nojima et al. reported that RASD1 hypermethylation was associated with dexamethasone resistance [8]. However, the clinical relevance of RASD1 expression and methylation in a well-defined cohort of patients with MM remains underexplored.
This study aimed to comprehensively evaluate RASD1 in MM. The mRNA and protein expression were analyzed in patient samples and correlated with clinical parameters, while the role of DNA methylation in regulating RASD1 expression was investigated. Furthermore, the functional consequences of demethylation on RASD1 expression and cell survival were examined in vitro.
Materials and methods
Patient samples and cell culture
Bone marrow aspirates and biopsy tissues were obtained from 26 newly diagnosed patients with MM and 8 healthy controls at the Affiliated Hospital of Zunyi Medical University between June 2019 and October 2020, after obtaining written informed consent. The study was approved by the Ethics Committee of the Affiliated Hospital of Zunyi Medical University (KLLY-2019-209), and was conducted in accordance with the Declaration of Helsinki. All patients with MM were diagnosed according to the International Myeloma Working Group criteria [9]. Peripheral blood mononuclear cells (PBMCs) were isolated using Ficoll density gradient centrifugation, and CD138^+^ plasma cells were subsequently purified from patient PBMCs using magnetic-activated cell sorting. The human MM cell line U266 was obtained from the American Type Culture Collection, and was maintained in Dulbecco’s Modified Eagle Medium supplemented with 10% fetal bovine serum.
RNA extraction and quantitative real-time PCR (RT-qPCR)
Total RNA was extracted from PBMCs, CD138^+^ cells, and U266 cells using TRIzol reagent, following the manufacturer's protocol. Complementary DNA (cDNA) was synthesized using a reverse transcription kit. RT-qPCR was performed with SYBR Green I Master Mix on a Cobas z480 system. GAPDH served as the endogenous control. The primer sequences for RASD1 were: Forward, 5′-CCA CCG CAA GTT CTA CTC CAT-3′; Reverse, 5′-CCA GGA TGA AAA CGT CTC CTG T-3′. Relative gene expression was calculated using the 2^(-ΔΔCt) method [10].
DNA extraction and methylation-specific PCR (MSP)
Genomic DNA was extracted using a commercial kit, and bisulfite conversion was performed with the EZ DNA Methylation-Gold™ Kit. MSP was conducted using primers specific for methylated (M) and unmethylated (U) alleles of the RASD1 promoter, as previously described, with modifications [8].
- Methylated Forward: 5′-TTTCGTAGTAGCGTGGATC-3′
- Methylated Reverse: 5′-GTAATCCGAACTCGAACTTT-3′
- Unmethylated Forward: 5′-AGTTTTTGTAGTAGTGTGGATT-3′
- Unmethylated Reverse: 5′-CTCATAATCCAAATCAAACTTT-3′
Immunohistochemistry (IHC)
Formalin-fixed, paraffin-embedded bone marrow biopsy sections were stained with an anti-RASD1 antibody, following previously described protocols [11]. Staining was evaluated using an immunoreactivity score (IRS), calculated by multiplying the score for the percentage of positive cells (0 = 0%, 1 = 1–25%, 2 = 26–50%, 3 = 51–75%, 4 = 76–100%) by the staining intensity score (0 = negative, 1 = weak, 2 = moderate, 3 = strong). An IRS > 4 was defined as high expression.
Scoring was performed independently by two board-certified pathologists who were blinded to the sample groups (patients with MM vs. healthy controls). Discrepancies, observed in ≤ 10% of cases, were resolved by consensus.
Cell treatment and apoptosis assay
U266 cells were treated with 50 μM decitabine (DAC) for 48 h. Apoptosis was measured by flow cytometry using Annexin V-YF647A/PI staining, according to the manufacturer's instructions [12].
Statistical analysis
Data were analyzed using SPSS version 18.0. Group comparisons were performed using Student's t-test or the chi-square test. Receiver operating characteristic curve analysis was performed to assess the diagnostic value of RASD1 mRNA expression. A P value < 0.05 was considered statistically significant.
Results
RASD1 expression is downregulated in MM
RASD1 mRNA expression was significantly reduced in bone marrow samples from patients with MM (0.42 ± 0.31) compared to healthy controls (1.28 ± 0.67, P < 0.001, Fig. 1a). Immunohistochemistry was employed to assess RASD1 protein expression. The results revealed high RASD1 expression in 7 out of 8 (87.5%) healthy controls, with only one case showing low expression. In contrast, high expression was observed in only 6 out of 26 (23.1%) patients with MM, while the remaining 20 cases exhibited low expression. Statistical analysis demonstrated that RASD1 protein expression was significantly reduced in patients with MM compared to healthy controls (P < 0.001, Fig. 1b, c).Fig. 1a Relative RASD1 mRNA levels (mean ± SD) in patients with MM versus healthy controls (HC), normalized to GAPDH (***P < 0.001). b Quantification of RASD1 protein expression (IRS scores, mean ± SD) in patients with MM versus HC (***P < 0.001). c Representative IHC images (400× magnification); Panels A and B: high RASD1 expression in HC bone marrow biopsies; Panels C and D: low RASD1 expression in bone marrow biopsies from patients with MM
Clinical correlations of RASD1 expression
Reduced RASD1 mRNA expression was significantly associated with advanced DS stage (P = 0.01) [13], anemia (P = 0.04), hypercalcemia (P = 0.03), and elevated M-protein levels (P = 0.04, Table 1). Similarly, low RASD1 protein expression was correlated with hypercalcemia (P = 0.04) and hyperglobulinemia (P = 0.04). Table 1. Correlation between RASD1 mRNA Expression and Clinical Characteristics in Patients with MMClinical CharacteristicTotal (n)RASD1 Expressionχ ^2^PHighLowGender Male166100.0160.9 Female1046Age (years) < 60185132.8210.09 ≥ 60853DS Stage Stage I & II10375.9870.01 Stage III1679ISS Stage Stage I & II10282.341.13 Stage III1688Anemia Yes209113.8690.04 No615White Blood Cell Normal249150.1220.72 Abnormal211Platelet Normal15690.0350.85 Abnormal1147Serum Corrected Calcium (mmol/L) ≤ 2.75191454.3980.03 > 2.75725Serum Phosphorus Normal13670.650.42 Abnormal1349M-protein (g/L) < 309273.3130.04 ≥ 3017710
RASD1 is silenced by promoter hypermethylation
MSP analysis revealed a high frequency of RASD1 promoter methylation in patients with MM, whereas samples from healthy controls were predominantly unmethylated (Fig. 2a). To functionally validate this mechanism, U266 cells were treated with DAC. This treatment significantly restored RASD1 mRNA expression (11.18 ± 1.98 vs. 1.04 ± 0.06; P < 0.001; Fig. 2b) and markedly increased apoptosis (12.08% vs. 5.04%, P < 0.01; Fig. 2c, d) compared to controls.Fig. 2a MSP gel electrophoresis showing methylation (M) and unmethylation (U) status in representative samples. b RASD1 mRNA expression in U266 cells following DAC treatment. c Quantification of apoptosis rates (mean ± SD) in U266 cells (**P < 0.01). d Representative flow cytometry plots of apoptosis (Annexin V-YF647A/PI staining); Total apoptosis: 5.04% in the control group and 12.08% in the DAC-treated group
Discussion
Multiple myeloma (MM) remains an incurable plasma cell malignancy despite advances in novel therapeutic agents [1, 2]. The emergence of drug resistance and clonal evolution continues to pose major clinical challenges, highlighting the need for new molecular targets and biomarkers [13]. The present study provides compelling evidence that RASD1 acts as a tumor suppressor gene epigenetically silenced through promoter hypermethylation in MM.
Epigenetic alterations, particularly DNA methylation, have emerged as critical drivers of MM pathogenesis and treatment resistance [14]. Recent studies have demonstrated that aberrant DNA methylation patterns not only silence tumor suppressor genes but also reshape the bone marrow microenvironment [15]. In this context, investigation of RASD1 is particularly relevant because it is located on chromosome 17p11.2, a region frequently altered in hematological malignancies [16].
Significant downregulation of RASD1 expression was observed at both transcriptional and translational levels in patients with MM compared to healthy controls. This finding aligns with recent reports identifying RASD1 as a potential tumor suppressor in other malignancies. For instance, a 2024 study by Ren M. et al. demonstrated that RASD1 restoration inhibits tumor growth in solid tumors through modulation of the MAPK pathway [17].
Mechanistically, DNA hypermethylation was identified as a direct regulator of RASD1 silencing in MM. Treatment with DAC reversed RASD1 expression and induced apoptosis, while MSP analysis confirmed promoter hypermethylation in primary patient samples. These findings are consistent with the current understanding of epigenetic regulation in MM, wherein DNA methyltransferase inhibitors have shown promise in reversing aberrant methylation patterns [18]. Notably, MSP is a qualitative assay; therefore, future studies using quantitative methylation techniques would further delineate the methylation burden of the RASD1 promoter in MM.
Clinically, low RASD1 expression was significantly correlated with established poor prognostic indicators, including advanced DS stage, anemia, and elevated M-protein levels [19, 20]. This clinical relevance is particularly important in the context of recent efforts to develop molecular biomarkers for risk stratification in MM [21]. The association with aggressive disease features suggests that RASD1 methylation status may serve as a valuable biomarker for identifying high-risk patients.
This study has several limitations. First, the sample size was small (26 patients with MM and 8 healthy controls), which limits the generalizability of the conclusions. Second, functional experiments relied on a single MM cell line (U266), and the DAC-induced increase in apoptosis was modest. Third, MSP is a qualitative assay that does not quantify methylation burden. Finally, external validation using publicly available datasets was not conducted. These limitations highlight the need for future studies incorporating larger sample sizes, additional cell lines, quantitative methylation assays, and independent validation to confirm the present findings.
Despite these limitations, the findings provide a foundation for future research. Recent advances in single-cell epigenomics and multiomics approaches present important opportunities to further elucidate the role of RASD1 in MM pathogenesis [22]. Additionally, combining epigenetic therapies with existing MM treatments represents a promising therapeutic avenue that merits further investigation [23].
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Agirre X, Castellano G, Pascual M, et al. Whole-epigenome analysis in multiple myeloma reveals DNA hypermethylation of B cell-specific enhancers. Genome Res. 2015;25(4):478-87.10.1101/gr.180240.114PMC 438152025644835 · doi ↗ · pubmed ↗
- 2Garcia-Gomez A, Li T, de la Calle-Fabregat C, et al. Targeting aberrant DNA methylation in mesenchymal stromal cells as a treatment for myeloma bone disease. Nat Commun. 2021;12(1):421.10.1038/s 41467-020-20715-x PMC 781386533462210 · doi ↗ · pubmed ↗
- 3Nojima M, Maruyama R, Yasui H, et al. Genomic screening for genes silenced by DNA methylation revealed an association between RASD 1 inactivation and dexamethasone resistance in multiple myeloma. Clin Cancer Res. 2009;15(13):4356-64.10.1158/1078-0432.CCR-08-333619549772 · doi ↗ · pubmed ↗
- 4Ren M, Pan H, Zhou X, et al. KIAA 1429 promotes gastric cancer progression by destabilizing RASD 1 m RNA in an m 6A-YTHDF 2-dependent manner. J Transl Med. 2024;22(1):584.10.1186/s 12967-024-05375-5PMC 1119126338902717 · doi ↗ · pubmed ↗
- 5Tirier SM, Mallm JP, Steiger S, et al. Subclone-specific microenvironmental impact and drug response in refractory multiple myeloma revealed by single-cell transcriptomics. Nat Commun. 2021;12(1):6960.10.1038/s 41467-021-26951-z PMC 863010834845188 · doi ↗ · pubmed ↗
- 6Kumar S, Kats LM, Gruber E. Epigenetic reprogramming in multiple myeloma-Challenges and opportunities. Int J Cancer. 2026;158(2):423-432.10.1002/ijc.35426 PMC 1262803640171810 · doi ↗ · pubmed ↗