Plastic Germination, Temporal Niche Partitioning and Emergent Assortative Mating in Annual Plants
Max Schmid, Katja Tielbörger, Amaël Daval, Charles Mullon

TL;DR
The paper shows how plants can evolve to germinate at different times, which helps them coexist and diversify into new species.
Contribution
The study reveals that plastic germination can drive genetic diversification and ecological speciation through temporal niche partitioning and assortative mating.
Findings
Adaptive plasticity in germination evolves via genetic associations with fecundity traits.
Plastic germination enhances temporal niche partitioning and promotes divergence into specialized morphs.
Plasticity generates temporal assortative mating, maintaining trait associations without genetic linkage.
Abstract
Temporal fluctuations in the environment can promote coexistence via the storage effect, where competing variants are buffered during unfavourable years. In annual plants, this can arise from seed dormancy: seeds remain in the seed bank across years and germinate under suitable conditions. Here, we investigate how plasticity in germination timing (where seeds use environmental cues to adjust when they germinate) affects genetic diversification and ecological speciation. Using eco‐evolutionary models, we show that adaptive plasticity readily evolves via genetic associations between germination and fecundity traits, allowing seeds to germinate preferentially in years favourable for reproduction. This enhances temporal niche partitioning and promotes divergence into specialised morphs. Because these morphs germinate in different years, plasticity generates temporal assortative mating and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
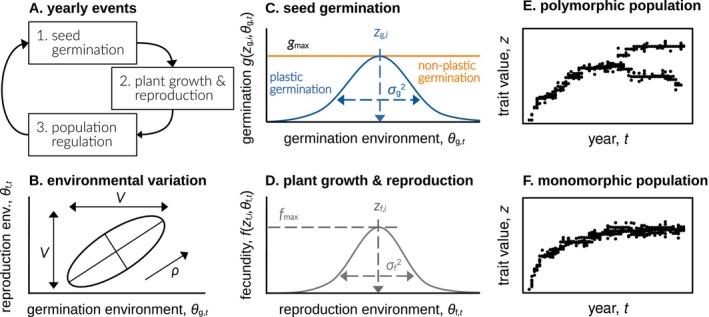 FIGURE 1
FIGURE 1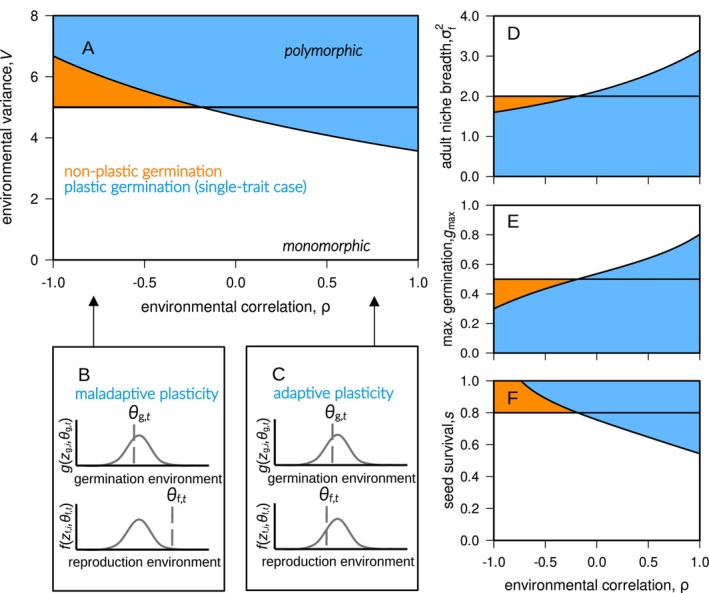 FIGURE 2
FIGURE 2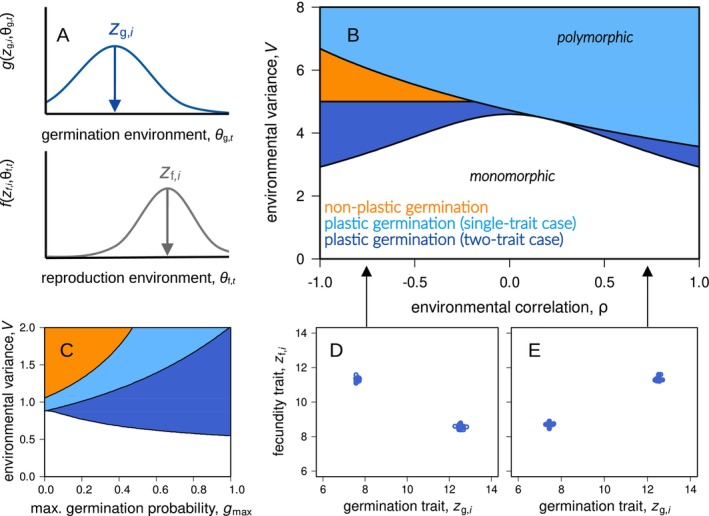 FIGURE 3
FIGURE 3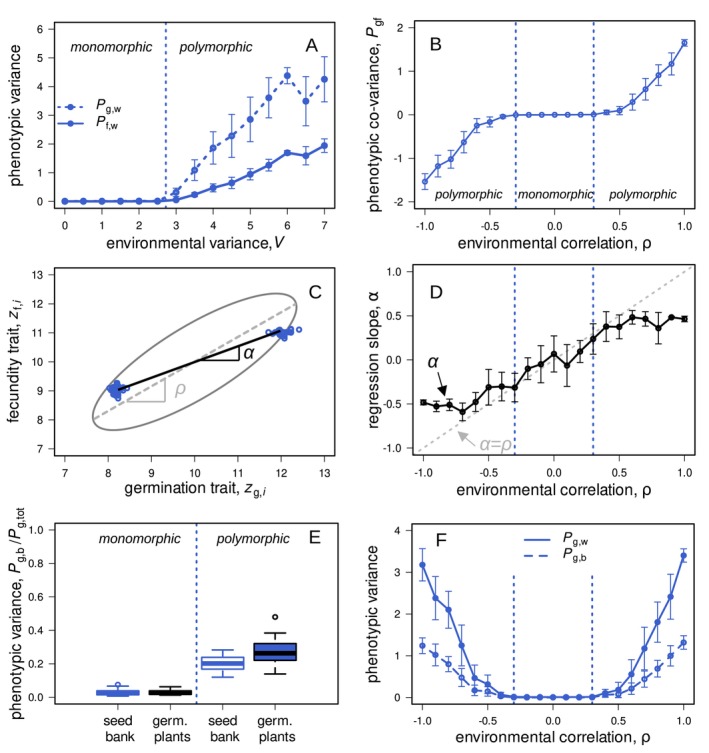 FIGURE 4
FIGURE 4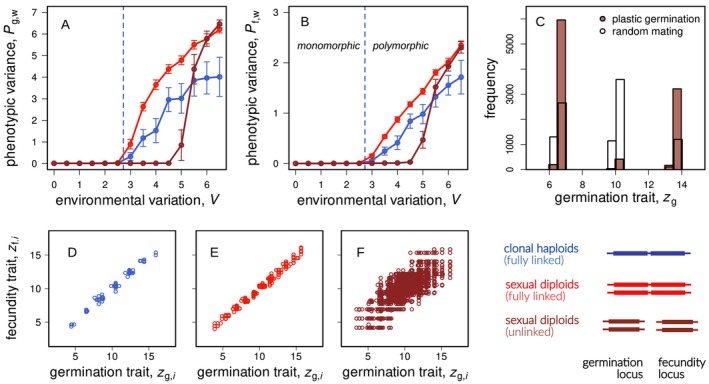 FIGURE 5
FIGURE 5- —Deutscher Akademischer Austauschdienst10.13039/501100001655
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant and animal studies · Evolution and Genetic Dynamics · Seed Germination and Physiology
Introduction
1
Most communities experience fluctuations in environmental conditions, with year‐to‐year variation in temperature, rainfall or disturbances such as fires or floods (Grant et al. 2017; Ummenhofer and Meehl 2017; He et al. 2019; Tielbörger et al. 2014; Pausas and Lamont 2022). Such temporal variability can promote biodiversity by maintaining genetic and/or phenotypic variation in two main ways. First, individuals can evolve to respond plastically to prevailing conditions, resulting in environmentally induced phenotypes (polyphenism; Woltereck 1909; Bradshaw 1965; Scheiner 1993; Schlichting 1986; Via et al. 1995). The evolution of adaptive plasticity requires reliable cues that predict the selective environment (Moran 1992; Tufto 2000; Bonamour et al. 2019), as well as sufficiently flexible developmental programs (DeWitt et al. 1998; Lande 2014; Dupont et al. 2024). Second, populations can evolve genetic polymorphisms, or species within communities can diverge via character displacement, leading to temporal niche partitioning. Here, different variants (alleles or species) specialize on contrasting yearly conditions (Chesson and Warner 1981; Ellner and Hairston 1994; Yamamichi et al. 2023). The evolution of temporal niche partitioning requires a mechanism buffering variants against selection in unfavorable years, known as the temporal storage effect (P. L. Chesson 1983; Svardal et al. 2015; Yamamichi et al. 2023). Determining how these processes shape genetic and phenotypic variation is essential for understanding biodiversity and its maintenance in temporally variable environments.
Adaptive plasticity and genetic polymorphism are typically viewed as alternatives but these processes may also interact (e.g., West‐Eberhard 1989; Crispo 2008; Conover et al. 2009; Ghalambor et al. 2015; Gulisija et al. 2016; Schmid and Guillaume 2017; Sommer 2020). One context where this interaction might be particularly relevant is temporal niche partitioning in annual plants. Under interannual fluctuations, niche partitioning readily evolves when seeds exhibit between‐year dormancy, whereby a fraction remains dormant for multiple years before germinating (P. L. Chesson 1994; Ellner and Hairston 1994; P. Chesson 2000; Barabás et al. 2018; P. Chesson 2020; Johnson and Hastings 2022), generating the necessary storage effect. Seed germination itself can show adaptive plasticity, with seeds germinating preferentially under conditions predicting favourable subsequent growth and reproduction (e.g., Angert et al. 2009; Donohue et al. 2010; Cohen 1967). Such predictive germination (Venable and Lawlor 1980; Pake and Venable 1996) can reinforce character displacement, causing different species to germinate in different years, thereby facilitating coexistence via the storage effect (Kortessis and Chesson 2021). However, existing theory has so far treated predictive germination mainly as a mechanism maintaining differences among already diverged species (Venable et al. 1993; Snyder and Adler 2011; Mathias and Chesson 2013; Kortessis and Chesson 2021). What remains unclear is whether the same process can also create diversity in the first place, by initiating genetic diversification and ecological speciation within initially homogeneous populations.
Here, we extend current theory to investigate whether the evolution of predictive germination facilitates the emergence and divergence of genetic morphs within annual plant species. We show that predictive germination enhances the conditions under which temporal niche partitioning evolves. We further explore evolutionary constraints imposed by sexual reproduction and recombination, revealing that adaptive divergence in germination timing generates temporal assortative mating, thus setting the stage for ecological speciation.
Model
2
Life Cycle and Traits
2.1
We model a large population of annual plants that shows between‐year seed dormancy (though this model can equally apply to other taxa, such as Daphnia with resting eggs or fungi with dormant spores). Seeds may remain dormant in the seed bank for multiple years, while plants that germinate are short‐lived and die after reproduction.
The population may experience environmental conditions during germination and reproduction that vary between years (e.g., fluctuations in humidity, temperature or herbivore density). We represent these fluctuations with two random variables per year t: θg,t, describing the environment during the germination period, and θf,t, describing the environment later in the year during reproduction. These conditions influence germination and reproduction through interactions with specific plant traits, as detailed below.
Seed Germination
2.1.1
At the beginning of each growing season, seeds in the seed bank either remain dormant or germinate. The probability that a seed indexed i, characterised by its germination trait zg,i, germinates in year t depends on the early‐year environment θg,t (e.g., temperature at the time of rainfall, see Baskin et al. 1993) as follows:
Equation 1 indicates that a seed germinates with a maximum probability, 0<gmax≤1, when its trait matches environmental conditions (i.e., when θg,t=zg,i; Figure 1C). In this view, trait zg,i could be seen as a physiological or morphological seed character that regulates the preferred environment for germination. Germination probability declines with increasing mismatch between a seed's trait and the environment, at a rate determined by σg2>0.
Graph A shows the yearly life‐cycle events, where a fraction of seeds germinates, reproduces, and the plant population then experiences density regulation. The ellipsoid in graph B illustrates the environmental distribution where environmental conditions during germination θg,t (x‐axis) and reproduction θf,t (y‐axis) are drawn from a bivariate normal distribution, each year anew. Both environmental conditions show between‐year variance V and within‐year environmental correlation ρ. Graph C contrasts plastic seed germination (blue line) with non‐plastic seed germination (orange line, where σg2→∞). Graph D shows how the individual fecundity changes with the match between θf,t and the individual fecundity trait zf,i. The evolutionary dynamics in zg and zf then could either lead to a polymorphic plant population under disruptive selection (E), or to a monomorphic population with stabilising selection (F).
Equation 1 sets a reaction norm describing how seed germination responds to the environment with the parameter σg2 tuning the degree of germination plasticity. A small σg2 corresponds to high plasticity, meaning germination probabilities change strongly in response to environmental variation (blue line in Figure 1C; e.g., see Simons 2014, for empirical data supporting bell‐shaped germination functions in response to temperature). In contrast, large σg2 corresponds to low plasticity, meaning germination probabilities remain relatively constant across a broad range of environmental conditions. In the limiting case (σg2→∞), germination becomes completely independent of the environment (non‐plastic), reducing the reaction norm to a horizontal line with intercept gmax (orange line in Figure 1C; e.g., see Ellner and Hairston 1994).
Plant Growth and Reproduction
2.1.2
After germination, plants grow and reproduce. We assume that the fecundity of plant i, characterised by its fecundity trait zf,i, in response to the reproduction environment θf,t (e.g., temperature during flowering), is given by.
where maximum fecundity fmax is reached when the fecundity trait matches environmental conditions during reproduction (i.e., when zf,i=θf,t; Figure 1D). Trait zf,i could be seen as a quantitative trait in germinated plants that governs the preferred environment for reproduction (like root morphology or stomata density). Fecundity declines with increasing mismatch between trait and environment at a rate determined by σf2>0. This parameter σf2 measures the breadth of the fecundity niche, with large σf2 indicating a broad niche (plants achieve high fecundity across a wide range of conditions) and small σf2 indicating a narrow niche (high fecundity is limited to specific environmental conditions). Equation 2 represents a classical assumption in many theoretical models of adaptation to environmental variables (e.g., Via and Lande 1985; Day 2000; Kisdi 2002; Chevin and Lande 2011; Svardal et al. 2015; Miller and Klausmeier 2017; Yamamichi et al. 2019; Ohtsuki et al. 2020; Orive et al. 2023; Saltini et al. 2023).
Regulation
2.1.3
After reproduction, all germinated plants die. Seeds that did not germinate remain dormant, surviving to the next year with probability s and dying with probability 1−s. Seed‐bank slots are freed when seeds germinate (and thus leave the seed bank) or when dormant seeds die. Newly produced seeds then compete for these vacant slots. We assume the seed‐bank size is constant, with N seeds present at the start of each year (with N large), and that recruitment into vacant slots is random with respect to trait values, following a lottery model (Chesson and Warner 1981).
The assumption of a constant seed‐bank size, which greatly facilitates our mathematical analysis, may seem contrived and biologically unrealistic. However, simulations where we relax this assumption and let the size of the seed bank fluctuate year‐to‐year (in a similar way to Kortessis and Chesson 2021) indicate that it has little effect on our model's results (see Appendix D.4).
Environmental Heterogeneity and Genetic Constraints
2.2
We assume environmental conditions during germination (θg,t) and reproduction (θf,t) fluctuate randomly among years around the same long‐term mean and with equal variance (Eθg,t=Eθf,t=θ¯ and Vθg,t=Vθf,t=V for all t; Supporting Information Appendix A). Conditions during germination and reproduction within the same year may be correlated (Corrθg,tθf,t = ρ for all t; Figure 1B; e.g., see Kortessis and Chesson 2021). A positive within‐year correlation (ρ>0) implies predictive germination cues (e.g., dry germination periods predict dry reproductive periods), whereas a negative correlation (ρ<0) indicates an inverse relationship (e.g., dry germination periods predict wet reproductive periods). Weak correlations (ρ≈0) imply limited predictability between conditions at germination and reproduction, likely impeding the evolution of predictive germination.
We explore how genetic constraints interact with environmental correlations to influence adaptation and diversification. We consider the joint evolution of the germination (zg) and fecundity (zf) trait, which together determine environmental conditions at germination and subsequent reproduction. We compare two scenarios: (i) the single‐trait case, where germination and fecundity traits are genetically constrained to be identical (zg,i=zf,i; such that each individual maximises germination and subsequent fecundity under the same conditions), and (ii) the two‐trait case, where traits evolve independently (such that each individual may adapt to different conditions for germination and for reproduction). Comparing these scenarios allows examination of how genetic constraints influence the evolution of temporal niche partitioning under different degrees of environmental correlation.
Analyses
2.3
We aim to identify the ecological and genetic conditions under which germination and fecundity traits become polymorphic, thus leading to temporal niche partitioning. We use two complementary approaches.
First, we conduct evolutionary invasion analyses (Supporting Information B and C), assuming haploid, clonally reproducing plants. Each trait is encoded by a single locus evolving under the continuum‐of‐alleles model. Mutations are rare and have small phenotypic effects such that evolution occurs in two phases (Geritz et al. 1998). Initially, directional selection drives the population toward convergence‐stable trait values with the population remaining largely monomorphic. Subsequently, the population either remains monomorphic under stabilising selection (Figure 1F) or undergoes evolutionary branching, becoming genetically and phenotypically polymorphic under disruptive selection (Figure 1E). Our goal is to identify conditions favouring evolutionary branching.
Second, we use individual‐based simulations, relaxing key analytical assumptions by explicitly incorporating diploidy, sexual reproduction and varying genetic linkage between germination and fecundity loci. These simulations test the robustness of our analytical predictions and allow us to investigate how sexual reproduction and recombination influence the evolution of trait polymorphism and temporal niche partitioning (Supporting Information Appendix D). They also allow us to analyze the emergence of temporal assortative mating and its consequences for ecological speciation.
Results
3
Germination Plasticity Facilitates Polymorphism Under Positive Environmental Correlations and Genetic Constraints
3.1
We first investigate the single‐trait case where individuals achieve maximum germination and maximum reproduction at the same environmental condition (i.e., zg,i=zf,i due to genetic constraints). As a baseline, we compare our results to the classical scenario without plasticity (σg2→∞; Figure 1C), for which polymorphism evolves under the condition 1−gmaxsV>σf2 (orange areas in Figure; P. L. Chesson 1983; Ellner and Hairston 1994; Svardal et al. 2015). Polymorphism in this baseline scenario is thus favoured by high between‐year environmental variability (V), narrow fecundity niches (small σf2), and extensive seed dormancy (small gmax, large s).
When allowing plastic germination (σg2<∞), conditions for evolutionary branching are more involved mathematically (see equation B16 in Supporting Information Appendix B for an explicit analytical solution) but can be easily interpreted graphically (Figure 2). Plastic germination broadens conditions leading to polymorphism under positive within‐year environmental correlations (blue areas in Figure 2, ρ>0). Here, plasticity allows plants to germinate preferentially in years reliably favourable for subsequent reproduction; that is, plasticity in this case is adaptive (following Gotthard and Nylin 1995's conceptual distinction). For instance, plasticity evolves such that plants adapted to warm conditions germinate and reproduce predominantly in warm years, while those adapted to cooler conditions germinate predominantly in cooler years. By allowing plants specialised to distinct environmental conditions to reproduce mostly in separate years, plastic germination here reduces between‐morph competition and enhances temporal niche partitioning.
Graphs A, D–F present analytical results for the single‐trait case (zg,i = zf,i) where plants evolve genetic/phenotypic polymorphism in the coloured areas, but stay monomorphic in the white areas. The case of plastic seed germination is presented in blue (when σg2 = 20; according to equation B13 with components B16), the case of non‐plastic germination is shown in orange (when σg2→∞; following 1−gmaxsV>σf2). If not specified in graph B and D–F, the parameter values are: gmax = 0.5, s = 0.8, σf2 = 2, V = 5, and ρ = 1. Graphs B and C conceptualise how environmental correlation leads to adaptive and maladaptive germination plasticity. With ρ<0 and large differences between θg,t and θf,t within years, a specialised plant morph (zg≠θ¯) might experience suitable conditions for germination, but detrimental conditions during reproduction (graph B). With ρ>0, environmental conditions do not change much within years (θg,t≈θf,t) such that plants with zg,i=zf,i could combine high germination probabilities with high fecundities.
Conversely, plastic germination restricts polymorphism relative to the non‐plastic scenario under negative environmental correlations (blue areas in Figure 2, ρ<0). In this situation, the same environmental preference during germination and fecundity cannot simultaneously match contrasting environmental conditions within a year (i.e., θg,t≠θf,t when ρ→−1; see equation A11). This mismatch means seeds germinate into conditions detrimental for reproduction, so germination plasticity here is maladaptive (Figure 2B). For example, warm‐adapted plants germinate preferentially under warm conditions but then experience cold conditions during reproduction. Such mismatch in turn hinders temporal niche partitioning.
Overall, we see that adaptive germination plasticity promotes polymorphism while maladaptive plasticity opposes it. This parallels findings by Kortessis and Chesson (2021), who showed that predictive germination promotes divergence between differentiated species when reliable cues are present and when the germination schedule can be aligned to beneficial conditions for plant growth.
In the absence of environmental correlation within years (ρ=0), germination plasticity can either facilitate or oppose polymorphism relative to the non‐plastic scenario, depending on parameters (Figure 2, horizontal line at ρ=0; equation B19). Two opposing effects explain this result. On the one hand, increased plasticity (smaller σg2) reduces the long‐term average germination probability, increasing seed dormancy and thus strengthening the storage effect, thereby favouring polymorphism. On the other hand, increased plasticity induces stabilising selection on the trait during germination when environmental variability (V) or seed survival (s) is limited (in fact, selection is stabilising if sV<σg2 when fecundity selection is weak, σf2→∞). The interplay of these two effects determines whether germination plasticity promotes or inhibits polymorphism when ρ=0.
Temporal Niche Partitioning Under Positive and Negative Environmental Correlation When Germination and Fecundity Traits Evolve Independently
3.2
We next allow germination and fecundity traits to evolve independently (two‐trait case). Allowing independent trait evolution broadens conditions leading to polymorphism, now occurring under both strongly positive and strongly negative environmental correlations within years (high absolute values of ρ, dark blue areas in Figure 3B). Under negative correlation (ρ<0), polymorphism emerges because germination and fecundity traits can evolve in opposite directions, each trait matching distinct environmental conditions within the same year. For example, some plants evolve to germinate preferentially under cool conditions but reproduce best under warm conditions, while others evolve the opposite pattern. This generates negative covariance between germination and fecundity traits (zg,i and zf,i; Figure 3D, see also equation C19 in Supporting Information Appendix C for the explicit analytical expression). Conversely, under positive environmental correlation (ρ>0), covariance between germination and fecundity traits becomes positive, aligning germination timing with conditions beneficial for reproduction (Figure 3E). Germination plasticity is thus adaptive at both positive and negative correlation, enhancing temporal niche partitioning whenever environmental correlations within years are sufficiently strong.
Graph A illustrates the two‐trait case where the germination trait and the fecundity trait could evolve independently and differ from each other within individuals (such that zg,i≠zf,i). The analytical results for the two‐trait case in graphs B and C present conditions leading to trait polymorphism and niche partitioning (dark blue area), and conditions leading to monomorphic plant populations (white area; from the Hessian matrix C6 with components C17‐C19). Note that the orange and light blue colours in graph B and C mask the dark blue areas underneath. Graphs D, E illustrate how plants could diversify with negative (positive) environmental correlation by the evolution of negative (positive) covariance between individual zg,i and zf,i values. Parameter values for graphs (B, D, E) are: gmax = 0.5, s = 0.8, σf2 = 2, σg2 = 20, V = 5, with ρ=1 (graph E) and ρ = −1 (graph D). The parameters for (graph C) are: s = 0.95, σf2 = 1, σg2 = 5, V = 5 and ρ = 1.
In the two‐trait scenario, high maximum germination probabilities (large gmax) can facilitate polymorphism, particularly under strong environmental correlations (large ∣ρ∣) and high seed survival (s; dark blue area in Figure 3C). This contrasts with our single‐trait scenario and with earlier models (e.g., Wisnoski and Shoemaker 2022), where increased dormancy (lower gmax) promotes polymorphism by enhancing the storage effect. This difference arises because independent evolution of the two traits enables the evolution of highly adaptive plasticity when ∣ρ∣ is large: specialised genotypes evolve to germinate almost exclusively in conditions that reliably predict high fecundity. As a consequence, increasing the maximum germination probability (gmax) allows these specialised genotypes to express their traits more frequently when conditions are favourable. Seed survival during dormancy (s) modulates this effect by enabling seeds to persist through unfavourable periods, thus maintaining the specialised genotypes across years and sustaining the storage effect.
Plastic Seed Germination Drives Phenotypic Variation Within and Among Years
3.3
We next examined the phenotypic distributions at mutation‐selection‐drift equilibrium that arose in the individual‐based simulations. We measured phenotypic variance in the germination and fecundity traits (Pg,w and Pf,w; see equation D5) in germinated plants. Conditions favouring polymorphism also led to greater phenotypic variance. Specifically, high environmental variability (V) and strong environmental correlation (∣ρ∣) resulted in increased phenotypic variance in both germination and fecundity traits (Figure 4A,F). This occurred because these conditions generated strong selection for coordinated trait combinations that were specialised to distinct yearly environmental conditions. When selection was sufficiently strong, more than two plant morphs could evolve and coexist (Figure 5D).
These graphs show simulation results for the two‐trait case with clonal reproduction in haploid plants. Graph A depicts within‐year phenotypic variance in the germination trait (dashed lines) and the fecundity trait (solid lines) as a function of interannual environmental variation V. The vertical dotted lines (—) present the threshold between stabilising and disruptive selection as predicted by the mathematical model (following the Hessian matrix C6 with components C17‐C19). Graph B illustrates the extent of trait covariance Pgf (see equation D7) as a function of environmental correlation, where polymorphism could evolve with positive and negative ρ. Graph C conceptualises the linear regression slope for the phenotypic distribution (α) and for the environmental distribution (ρ), where plants not always evolve α=ρ (graph D). Graph E shows simulation results of the between‐year phenotypic variance (Pg,b/Pg,tot) for seeds in the seed bank and for germinated plants only. Graph F presents the between‐year phenotypic variance as a function of environmental correlation. The parameter values for graphs A‐F are: V = 4, gmax = 0.5, s = 0.8, σf2 = 2, σg2 = 10 (with ρ = 0.8 for graph A and C; and ρ = (0,1) for graph E). The whiskers in graphs A, B, D, F illustrate variation among replicates (covering one standard deviation).
Graphs A‐F show simulation results for the two‐trait case with clonally reproducing haploid plants (in blue) and for outcrossing diploid plants with and without genetic linkage (light and dark red curves, respectively). The vertical dashed lines in graphs A and B present the analytical threshold between conditions leading to monomorphic and polymorphic plant populations (from the Hessian matrix C6 with components C17‐C19). The whiskers represent variation among replicates (spanning one standard deviation) and the parameter values for these simulations are: gmax = 0.5, s = 0.8, σf2 = 2, σg2 = 10, ρ = 1. The histograms in graph C present the phenotypic distribution at the end of a single simulation run with temporal assortative mating due to germination plasticity (dark red bars) and with random mating among all plant seeds (transparent bars). Temporal assortative mating leads to a reduced hybridization between the outer morphs relative to the case of random mating. Simulation parameters are the same as in graphs A‐B with V = 6. Graphs D–F show the phenotypic distribution of single simulation runs for varying genetic architectures with parameter values: gmax = 0.2, s = 0.98, σf2 = 1, σg2 = 1, V = 10, ρ = 0.9.
We further investigated how closely the phenotypic distributions of germinated plants matched the underlying environmental distributions (Figure 1B). First, we measured the phenotypic covariance (Pgf,w; see equation D7) between individual germination and fecundity traits (zg,i and zf,i). The sign of this covariance consistently matched the sign of the environmental correlation ρ (Figure 4B), confirming our analytical predictions (equation C19 in Supporting Information Appendix C). Next, we compared the slope of the linear regression of germination on fecundity traits (α=Pgf,w/Pg,w; Figure 4C) to the environmental correlation ρ. Though α and ρ were closely aligned, they did not match perfectly (i.e., α≠ρ; Figure 4D). Such deviations occurred because trait evolution depended not only on adaptation to environmental fluctuations, but also on reducing competition between specialised morphs. As a consequence, plants evolved phenotypic trait combinations that slightly deviated from a perfect match to environmental conditions. Similar results were found by Kortessis and Chesson (2021), who observed reduced predictive germination as a consequence of competition among plants.
We also measured between‐year phenotypic variance (Pg,b and Pf,b; see equation D6). Polymorphic populations consistently exhibited higher between‐year variance (Figure 4E,F). Moreover, this variance was greater among germinated plants than among seeds in the seed bank (Figure 4E). This was because polymorphic populations could respond more strongly to changing selective pressures across years, causing rapid changes in morph frequencies that track environmental changes. Under plastic germination, only phenotypes well‐adapted to current environmental conditions germinated each year, even when the composition of the seed bank remained relatively stable. Thus, the phenotypic composition of germinated plants became uncoupled from that of the seed bank, leading to substantial year‐to‐year shifts. Between‐year variance among germinated plants accounted for up to approximately 30% of the total phenotypic variance (Figure 4E), indicating that a single‐year measurement captured only about 70% of the total long‐term phenotypic variance.
Outcrossing and the Rise of Temporal Assortative Mating
3.4
As a final extension, we examined the evolution of polymorphism in diploid plants with sexual reproduction. Here, germination and fecundity traits were controlled by separate genetic loci without physical linkage or pleiotropy, and mating was random. Under these conditions, polymorphism evolved under narrower environmental conditions compared to clonal reproduction (compare blue and dark red lines, Figure 5A,B). This was because recombination breaks down genetic associations between germination and fecundity traits, which were necessary for adaptation to correlated environments (ρ≠0). Consistent with this interpretation, when loci controlling germination and fecundity were completely linked, polymorphism evolved under conditions similar to those with clonal reproduction (light red lines, Figure 5A,B).
However, when environmental fluctuations were sufficiently strong, polymorphism emerged even without physical linkage between loci. This occurred through the evolution of temporal assortative mating driven by divergence in germination timing (zg, dark red curves in Figure 5A,B). Plants with differentiated germination traits germinated preferentially in different years, thus reducing gene flow between differentiated morphs. This temporal segregation enabled genetic associations (linkage disequilibrium) between germination and fecundity traits to establish despite free recombination (Figure 5F). Temporal assortative mating was also reflected clearly in the phenotypic distribution of the population (Figure 5C). Under sexual reproduction, two highly differentiated morphs coexisted, along with an intermediate morph that resulted from mating between them. However, this intermediate morph occurred less frequently than would be expected if matings were entirely random (Figure 5C, red vs. white histograms).
Phenotypic variance was typically larger in polymorphic populations under sexual reproduction compared to clonal reproduction, in particular in the germination trait (Figure 5A, dark red curves exceed blue curves for large V). This was because divergence in the germination trait (zg) was under additional selection in sexual populations as differentiated germination schedules led to temporal assortative mating, reducing the formation of maladapted hybrids (see Aubier et al. 2023, for a similar result). Additionally, sexual reproduction further led to a so‐called segregation variance (Slatkin and Lande 1994) which also contributed to greater Pg,w and Pf,w in our simulations with sexual reproduction (compare blue to red curves in Figure 5A,B).
Discussion
4
Our analyses indicate that adaptive plasticity in seed germination readily evolves and facilitates the emergence and maintenance of genetic diversity in annual plants under temporally fluctuating environments. Such predictive germination boosts the storage effect by allowing seeds to germinate preferentially in years favourable for growth and reproduction. This in turn promotes divergence among genetic morphs that can specialise to different yearly conditions and leads to temporal niche partitioning. In contrast, when genetic constraints lead to maladaptive plasticity, seeds often germinate into unfavourable conditions, opposing diversification. Our results thus extend existing theory on the storage effect (Venable et al. 1993; Snyder and Adler 2011; Kortessis and Chesson 2021), demonstrating that adaptive plasticity and genetic polymorphism can interact to reinforce biodiversity.
Predictive germination facilitates diversity by generating positive correlations between germination timing and favourable conditions for reproduction. When seeds reliably detect cues predicting good years, they can align their germination with environments best suited for growth and fecundity. Such alignment reduces competition among emerging genetic morphs, as each specializes on a distinct subset of years (Chesson and Warner 1981; Ellner and Hairston 1994). This interplay between germination and fecundity traits produces strong phenotypic covariance, further promoting niche partitioning and enabling stable coexistence (Kortessis and Chesson 2021). Adaptive germination plasticity thus enhances temporal niche differentiation beyond what is possible with divergence in a single trait.
Our results also suggest that predictive germination can promote reproductive isolation, thus setting the stage for ecological speciation. Divergence in germination timing generates temporal assortative mating, reducing gene flow among plant morphs specialised on different annual conditions. In that sense, the germination trait in our model can be seen as a ‘magic trait’, simultaneously controlling adaptation to the environment and facilitating reproductive isolation (Servedio et al. 2011). Such traits are particularly effective drivers of ecological speciation because they prevent genetic recombination and sexual reproduction from breaking down adaptive trait combinations (Dieckmann and Doebeli 1999; Gavrilets and Vose 2007). Even when loci controlling germination and fecundity traits are unlinked, temporal assortative mating allows trait covariance to emerge and persist, circumventing recombination's homogenising effects (Dieckmann and Doebeli 1999; Aubier et al. 2023). When temporal assortative mating via plastic germination is insufficient to override recombination, selection may favour tighter genetic linkage or pleiotropy between germination and post‐germination traits, potentially promoting ‘supergenes’ that further stabilise adaptive trait combinations (Sinervo and Svensson 2002). Alternatively, other mechanisms strengthening temporal assortative mating, such as genetic incompatibility systems, could enhance reproductive isolation, avoid the production of unfit hybrids and thus facilitate diversification.
Our findings contribute to a broader discussion on how phenotypic plasticity affects genetic diversification. Adaptive plasticity is often viewed as an alternative adaptation to genetic diversification, potentially limiting genetic divergence by enabling a single genotype to exploit multiple environments (Ghalambor et al. 2007; Pfennig et al. 2010; Turcotte and Levine 2016; Schmid and Guillaume 2017). Predictive germination provides an interesting exception, showing instead how plasticity can facilitate genetic diversification. This illustrates a more general mechanism whereby the existence of traits or traits' associations that allow genotypes to be preferentially expressed in contexts where they perform best favours diversification (Avila and Mullon 2023; Lehmann and Mullon 2025). Our arguments therefore also apply more broadly to life‐history plasticity, where adaptive transitions between life stages may allow populations to exploit distinct temporal niches more effectively than divergence in a single trait alone. Rather than being mutually exclusive, adaptive plasticity and genetic diversification can thus act synergistically to enhance biodiversity (see also West‐Eberhard 1989 for an alternative mechanism; Hendry 2016 and Sommer 2020 for a review).
Plastic seed germination evolves in our model, but it rarely yields perfect tracking of favourable years. Such limitations to plasticity are well known (DeWitt et al. 1998), and two arise directly under our assumptions. First, genetic architecture can prevent germination from aligning with favourable growth conditions: in the single‐trait case with negative within‐year correlation (ρ<0), the germination trait cannot diverge sufficiently from the fecundity trait, while in the two‐trait sexual case with weak or no linkage, recombination erodes the build‐up of adaptive covariance between germination and fecundity. Second, plastic responses are limited by imperfect environmental information: when germination cues are only weakly predictive of the subsequent reproductive environment, as is commonly assumed for natural systems (Hendry 2016), seeds inevitably germinate in some years that later prove unfavourable. In addition to these constraints, we did not model explicit costs of plasticity. Such costs have been documented (van Buskirk and Steiner 2009) and may arise from the machinery required for cue detection and processing (Scheiner et al. 2020). If present, these costs would reduce survival or fecundity of plastic genotypes and would therefore be expected to further limit the evolution of strong germination plasticity and contract the parameter range supporting temporal niche partitioning. Importantly, costs would change the quantitative scope of plasticity evolution, but not the qualitative mechanism by which cue‐based germination creates temporal niche differences and thereby promotes the storage effect.
Empirical studies provide support for our findings on predictive germination. For example, some long‐term studies of desert annual plants indicate associations between germination timing and conditions favourable for reproduction, such as specific rainfall or temperature cues (Venable et al. 1993; Pake and Venable 1996; Gremer et al. 2016). Also, germination fractions within a year seem to correlate with the expected competitive environment (Tielbörger and Valleriani 2005; Tielbörger and Prasse 2009). Other potentially relevant examples come from fire‐prone ecosystems, where germination cues such as smoke or heat may reliably predict beneficial growth conditions, including enhanced nutrient availability and reduced competition (Keeley and Pausas 2022). In these systems, germination traits correlate with adaptive post‐germination traits like high‐light tolerance or rapid post‐fire growth (Pausas and Lamont 2022). More broadly, a diverse set of environmental cues has been linked to predictive germination (including salinity and light cues, see table 2 in Donohue et al. 2010), and environmentally induced seed germination is associated with high diversification rates in several plant lineages (Willis et al. 2014).
Our findings also provide a potential explanation for the pronounced between‐year variation observed in some annual plant communities. In our model, predictive germination creates strong year‐to‐year changes in the phenotypic composition of germinated plants, despite a relatively stable composition of the underlying seed bank. Such between‐year fluctuations in community composition have indeed been reported from long‐term field surveys, with some species appearing abundantly in certain years and remaining virtually undetected in others (Pake and Venable 1996; Facelli et al. 2005; Levine et al. 2008; Tielbörger et al. 2014; Carrasco‐Puga et al. 2021). These dynamics could complicate biodiversity assessments because short‐term surveys may underestimate true diversity hidden in dormant seed banks (Carrasco‐Puga et al. 2021). Accurate evaluation of biodiversity under temporal fluctuations therefore calls either for long‐term monitoring data, comprehensive germination trials or genetic techniques such as environmental DNA analyses to fully capture community diversity.
Climate change could alter the ecological and evolutionary implications of predictive germination by modifying patterns of environmental predictability. As environmental cues may become less reliable with climate change (Kingsolver and Buckley 2018; Visser and Gienapp 2019; Edwards and Yang 2021), the capacity of seeds to match germination timing to optimal growth conditions may decline, reducing the effectiveness of temporal niche partitioning and potentially eroding biodiversity. In particular, increasingly erratic rainfall patterns, temperature shifts and altered disturbance regimes could disrupt previously stable associations between germination cues and favourable reproductive conditions. Such disruptions could disproportionately affect plant communities adapted to strongly correlated environmental fluctuations, possibly leading to declines in population sizes and shifts in species composition. Understanding how predictive germination strategies might respond to ongoing environmental change therefore emerges as a relevant question for future biodiversity dynamics.
Our model relies on several simplifying assumptions, and some modifications may be required to match particular systems. For instance, seed‐bank densities can vary substantially among years whereas we assume a constant size N. We relaxed this assumption in Supporting Information Appendix D.4 and still found that predictive germination strengthens the storage effect, consistent with Kortessis and Chesson (2021). In addition, while germination fractions in some systems, such as desert annuals, may increase monotonically with precipitation (e.g., see Lampei et al. 2017), we assume a bell‐shaped reaction norm; this affects quantitative details but not the core process by which predictive germination generates temporal niche differences. The model also omits parental effects (Tielbörger and Petrů 2010), which can contribute to temporal niche partitioning (Yamamichi and Hoso 2017); if adaptive, they would be expected to amplify the effects of predictive germination. Finally, germination and fecundity are often polygenic traits, whereas we modelled a one‐locus‐one‐trait architecture. Polygenic architectures may initially impede the build‐up of adaptive covariances between traits (e.g., Chebib and Guillaume 2022). However, if genetic modifiers are allowed to evolve, selection should favour a concentration of genetic effects that reduces recombination load, thereby facilitating the association between germination and fecundity traits.
To conclude, our study reveals that adaptive plasticity in seed germination promotes genetic diversification by facilitating temporal niche partitioning and reproductive isolation in temporally fluctuating environments. These results highlight that adaptive plasticity and genetic diversification are not mutually exclusive but instead can act together to enhance biodiversity. Understanding such interactions will help us better understand how natural populations might respond to future environmental change.
Author Contributions
Max Schmid, Amaël Daval, Charles Mullon and Katja Tielbörger conceived the project. Max Schmid performed the mathematical analyses with input from Charles Mullon, and Max Schmid ran the simulations with contributions from Amaël Daval. Max Schmid wrote the first draft of the manuscript, Charles Mullon revised the draft. All authors contributed to the final version.
Funding
This work was supported by SAGE, a Global Climate Centre, funded by the German Academic Exchange Service (DAAD).
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Data S1: ele70346‐sup‐0001‐supinfo.pdf.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Angert, A. L. , T. E. Huxman , P. Chesson , and D. L. Venable . 2009. “Functional Tradeoffs Determine Species Coexistence via the Storage Effect.” Proceedings of the National Academy of Sciences 106, no. 28: 11641–11645.10.1073/pnas.0904512106 PMC 271062219571002 · doi ↗ · pubmed ↗
- 2Aubier, T. G. , R. Bürger , and M. R. Servedio . 2023. “The Effectiveness of Pseudomagic Traits in Promoting Premating Isolation.” Proceedings of the Royal Society B: Biological Sciences 290: 20222108.10.1098/rspb.2022.2108 PMC 999305836883275 · doi ↗ · pubmed ↗
- 3Avila, P. , and C. Mullon . 2023. “Evolutionary Game Theory and the Adaptive Dynamics Approach: Adaptation Where Individuals Interact.” Philosophical Transactions of the Royal Society B 378: 20210502.10.1098/rstb.2021.0502 PMC 1002499236934752 · doi ↗ · pubmed ↗
- 4Barabás, G. , R. D'Andrea , and S. M. Stump . 2018. “Chesson's Coexistence Theory.” Ecological Monographs 88, no. 3: 277–303.
- 5Baskin, C. C. , P. L. Chesson , and J. M. Baskin . 1993. “Annual Seed Dormancy Cycles in Two Desert Winter Annuals.” Journal of Ecology 81, no. 3: 551–556.
- 6Bonamour, S. , L.‐M. Chevin , A. Charmantier , and C. Teplitsky . 2019. “Phenotypic Plasticity in Response to Climate Change: The Importance of Cue Variation.” Philosophical Transactions of the Royal Society, B: Biological Sciences 374, no. 1768: 20180178.10.1098/rstb.2018.0178 PMC 636587130966957 · doi ↗ · pubmed ↗
- 7Bradshaw, A. D. 1965. “Evolutionary Significance of Phenotypic Plasticity in Plants.” Advances in Genetics 13: 115–155.
- 8Carrasco‐Puga, G. , F. P. Díaz , D. C. Soto , et al. 2021. “Revealing Hidden Plant Diversity in Arid Environments.” Ecography 44: 98–111.