Direct and Indirect Effects of Floral Defoliation on Photochemical and Non‐Photochemical Chlorophyll Fluorescence Dynamics of a Semiarid Bunchgrass
Erik P. Hamerlynck, Rory C. O'Connor

TL;DR
This study explores how clipping florets in crested wheatgrass affects photosynthesis and energy dissipation in remaining parts of the plant.
Contribution
The study reveals an integrated photosynthetic response in crested wheatgrass following floral defoliation.
Findings
Clipping basal florets increased regulated non-photochemical photoprotection without reducing PSII quantum yield.
Distal florets showed higher PSII quantum yield and reduced unregulated energy dissipation after clipping.
Crested wheatgrass compensates for tissue loss through an integrated reproductive photosynthetic system.
Abstract
Photosynthetic florets support reproductive development and energetic seed provisioning of semi‐arid bunchgrasses whose population dynamics rely mainly on sexually produced propagules. Photosynthetic gas exchange studies including crested wheatgrass ( Agropyron cristatum ) have found compensatory increases in seed‐head photosynthesis following floral defoliation are accompanied by reduced light‐adapted PSII quantum yield (ϕ PSII). We undertook a field experiment to ascertain if altered ϕ PSII and optimal PSII quantum yield (F v/F m) were concurrent with higher quantum yield of regulated (ϕ NPQ) or unregulated (ϕ NO) non‐photochemical PSII absorbed energy dissipation. We quantified these responses in directly affected basal florets, and in unclipped distal florets to establish indirect responses to tissue loss. Clipping basal florets reduced F v/F m and increased ϕ NPQ, indicating…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
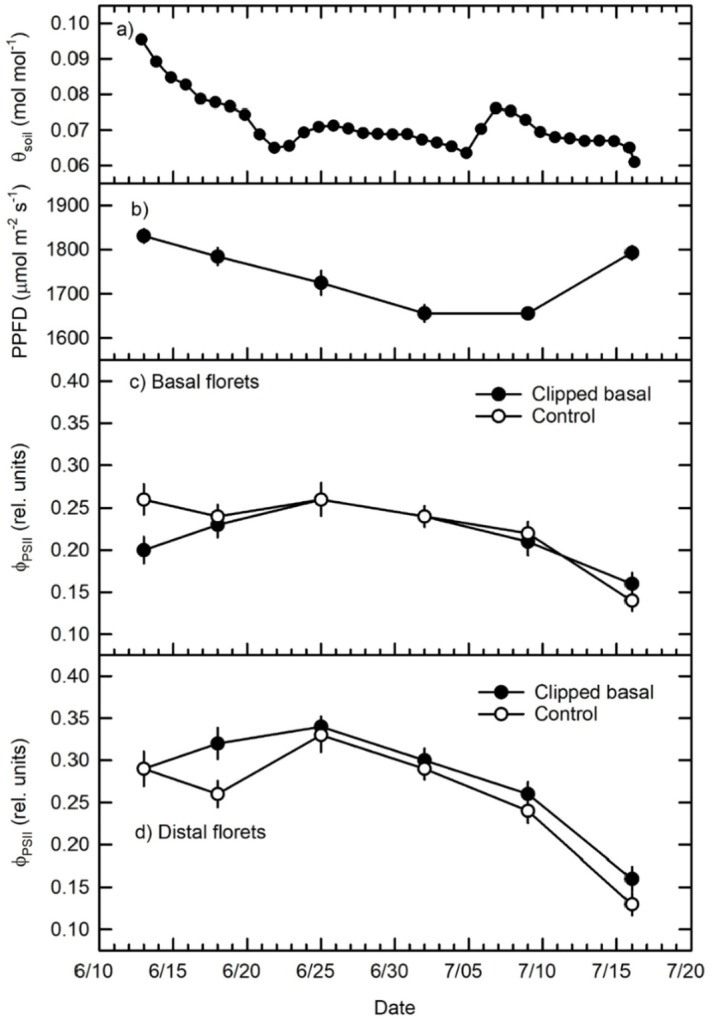 FIGURE 1
FIGURE 1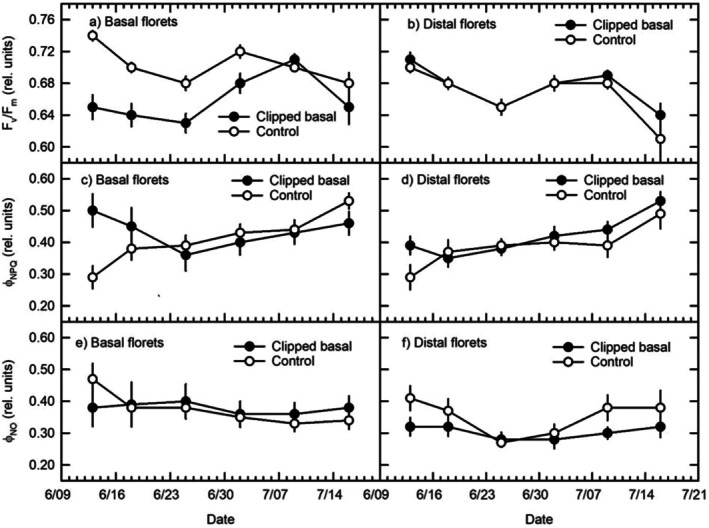 FIGURE 2
FIGURE 2| Structure‐parameter | Basal clip(1,30) | Phenological stage(2,60) | Clip × stage(2,60) |
|---|---|---|---|
| Basal floret— | 1.44 |
| 2.21 |
| Basal floret— |
| 0.62 |
|
| Basal floret— | 1.17 |
|
|
| Basal— | 0.14 | 1.49 | 1.15 |
| Distal floret— |
|
| 0.25 |
| Distal floret— | 1.98 |
| 1.53 |
| Distal floret— | 2.32 |
| 0.62 |
| Distal floret— |
|
| 1.00 |
| Position | Pre‐Anthesis | Anthesis | Post‐Anthesis | |||
|---|---|---|---|---|---|---|
| Base clip | Control | Base clip | Control | Base clip | Control | |
| Basal | ||||||
|
| 0.21 (0.011)b | 0.25 (0.011)a | 0.25 (0.011)a | 0.25 (0.001)a | 0.18 (0.011)c | 0.18 (0.011)c |
|
|
|
|
|
|
| 0.69 (0.007)b |
|
|
|
|
| 0.41 (0.021)ab |
| 0.48 (0.021)a |
|
| 0.38 (0.045)a | 0.43 (0.031)a | 0.38 (0.033)a | 0.34 (0.020)a | 0.37 (0.025)a | 0.34 (0.019)a |
| Distal | ||||||
|
| 0.30 (0.012)ab | 0.28 (0.013)b | 0.32 (0.010)a | 0.31 (0.012)a | 0.21 (0.014)c | 0.18 (0.013)c |
|
| 0.69 (0.006)a | 0.69 (0.005)a | 0.67 (0.006)b | 0.66 (0.007)bc | 0.67 (0.009)b | 0.64 (0.013)c |
|
| 0.37 (0.020)bc | 0.33 (0.028)c | 0.40 (0.018)bc | 0.40 (0.015)bc | 0.48 (0.020)a | 0.44 (0.031)ab |
|
|
|
| 0.28 (0.018)c |
| 0.31 (0.019)bc |
|
- —Agricultural Research Service10.13039/100007917
- —Oregon State Agricultural Experiment Station
- —Agricultural Research Center10.13039/501100024856
- —Oregon State University10.13039/100009612
- —Oregon State University10.13039/100009612
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBioenergy crop production and management · Turfgrass Adaptation and Management · Weed Control and Herbicide Applications
Introduction
1
In semiarid sagebrush steppe bunchgrasses, photosynthetic activity in the seed‐head is an important determinant of seed quality, as field experiments have shown seed‐head shading markedly reduces seed specific mass, an indicator of parental energetic provisioning (Hamerlynck and O'Connor 2021, Hamerlynck et al. 2024). This is important as these grasses' population dynamics rely almost exclusively on sexual reproduction (Liston et al. 2003; Hamerlynck and Davies 2019) and seed specific mass is associated with seedling traits that contribute to successful establishment (Quigley et al. 2023). The highly successful exotic Eurasian bunchgrass crested wheatgrass, Agropyron cristatum ([L.] Gaertn.), shows compensatory increases in photosynthesis in damaged florets that serve to enhance its reproductive effort in the remaining undamaged florets (Hamerlynck et al. 2023), a result not observed in native bunchgrasses (Quigley et al. 2024). In both these studies, floral defoliation decreased light‐adapted PSII yield (ϕ PSII; Genty et al. 1989), depending on reproductive phenological stage (Hamerlynck et al. 2023), or genotypic variation associated with selection for high reproductive output (Quigley et al. 2024). Herbivory‐induced alterations to ϕ PSII are well‐documented across a wide variety of plants and different modes of herbivory and are often associated with engagement of non‐photochemical photoprotective mechanisms (Nabity et al. 2012; Jänkäpää et al. 2013; Barton 2016; Henschel et al. 2024). While well‐studied in vegetative tissue, photoprotective activity in response to defoliation in graminoid reproductive structures has not been well‐documented.
Non‐photochemical dissipation of excess light energy is a critical eco‐functional feature of all oxygenic‐photosynthetic organisms (Demmig‐Adams and Adams III 1992; Osmond 1994; Derks et al. 2015), and is a function of the quantum yields of physiologically regulated (ϕ NPQ) and unregulated constituent quenching processes (ϕ NO; Klughammer and Schreiber 2008), both of which vary in response to environmental conditions and between species with distinct ecological tolerances (Proctor and Smirnoff 2011; Samson et al. 2019). Photosynthetic activity within the seed‐head plays a central role in the energetic provisioning and attendant reproductive effort of aridland bunchgrasses (Hamerlynck and O'Connor 2021; Hamerlynck et al. 2024). Understanding how floret ϕ NPQ and ϕ NO respond to defoliation may be important, as these may modulate compensatory increases in photosynthesis in damaged florets associated with increased reproductive potential in the remaining unaffected florets (Hamerlynck et al. 2023).
Here, we present an experimental field study assessing the photochemical and non‐photochemical responses of crested wheatgrass to simulated floral herbivory. As stated earlier, crested wheatgrass has demonstrated capacity to increase affected floret photosynthesis in response to defoliation, and that this is accompanied by a marked decrease in ϕ PSII (Hamerlynck et al. 2023). In addition to ϕ PSII, we measured optimal PSII quantum yield (F v/F m; Genty et al. 1989) to establish the full photochemical response to defoliation. We expected clipping to induce a decrease in F v/F m and ϕ PSII and it would be concurrent with increased ϕ NPQ, indicating these declines are photoprotective and are under physiological regulation. Moreover, we also tracked F v/F m, ϕ PSII, ϕ NPQ and ϕ NO in florets distal to clipped and unclipped controls to establish any indirect effects of floral tissue removal on photochemical and non‐photochemical processes.
Materials and Methods
2
Field work was performed from June 10 to July 17, 2025, in a level area of intact Wyoming big sagebrush ( Artemisia tridentata [Nuttall] spp. wyomingensis Beetle & Young) in a stand of naturally established crested wheatgrass on the Northern Great Basin Experimental Range (NGBER; 119.691709 W, 43.472600 N, 1395.4 m ASL), located ca. 70 km from Burns, OR. Supporting environmental measurements of daily average rooting zone volumetric soil moisture (θ soil) at 10 cm depth were made at a soil moisture monitoring array established in 2018 (see Hamerlynck and Ziegenhagen 2020 for installation and data acquisition details). Precipitation data was obtained from a National Atmospheric Deposition Program/National Trends Network (NADP/NTN) site (Site OR07; https://nadp.slh.wisc.edu/precipitation/) located 206 m SE from the sampling location.
Four days prior to chlorophyll fluorescence measurements, eight reproductive culms with recently emerged seed‐heads were tagged at the base on eight randomly selected plants. Two days later, four of the culms on each plant had ca. 50% of their basal floret area ca. 3 cm from the distal tip of the inflorescence clipped using scissors as per Hamerlynck et al. (2023). Night‐time dark‐adapted (ca. 2:00–4:30 AM, PDT) and mid‐morning light‐adapted (ca. 8:30–10:00 AM PDT) chlorophyll fluorescence (F) measurements were made on basal and distal florets with a Li‐600‐N porometer/fluorometer (LiCOR Instruments, Lincoln, NE). On each of the six seasonal sampling dates, two clipped and two control culms were randomly selected for night‐time measurements from marked culms on each plant (n = 16 per treatment type, total n = 32 culms for each time point), with their identifying information recorded for subsequent daytime remeasurement. Tissue was enclosed across the full length of a 0.15 × 0.05 cm cuvette and exposed to a modulated F measuring beam provided by 2 LED light sources focused on the cuvette, filtered by a 750 ± 40 nm band‐pass filter, with fluorescence detected from 700 to 780 nm. Determination of baseline fluorescence (F_o_) in the dark and light‐adapted steady‐state fluorescence (F_s_) under incident ambient sunlight were established via a sub‐saturating modulated beam pulsed at 4–8 Hz over 2–5 s. This was followed by a saturating flash of 10,000 μmol m^−2^ s^−1^ intensity with a pulse width of 667 ns and 250–750 kHz frequency to determine dark‐adapted maximum fluorescence (F_m_) and light‐adapted maximum fluorescence yield (F_m_′). F_s_ and F_m_′ were determined using incident photosynthetic photon flux density (PPFD in μmol m^−2^ s^−1^) measured concurrently with a Si‐photodiode quantum sensor integrated into the Li‐600‐N, as the actinic light source, with all samples oriented perpendicular to the sun to receive similar levels of incident actinic PPFD. From these, maximum PSII quantum efficiency (F v/F m (F m – F o)/F m) and light‐adapted PSII quantum yield ((ϕ PSII; F m′ – F s)/F m′) were determined. F m and F m′ were used to estimate the quantum yield of non‐photochemical energy loss (NPQ) in PS II that is physiologically regulated (ϕ NPQ; ϕ NPQ = (F s/F m′) – (F s/F m)) and unregulated (ϕ NO; ϕ NO = F s/F m) (Klughammer and Schreiber 2008).
Split‐plot repeated measures analysis of variance (RM‐ANOVA) (Statisix v.8.0; Analytical Software, Tallahassee, FL) was used to test for differences in photochemical and non‐photochemical performance of clipped and unclipped basal florets, and intact florets distal to these. Individual culms were considered replicates for our statistical analyses, using data from basal and distal florets from 16 clipped and 16 unclipped culms on each sampling date (total n = 32 culms). We did so considering the independent and modular nature of plant functional structure and light interception at a canopy level in perennial bunchgrasses (Sprugel et al. 1991; Ryel et al. 1993). As we were not interested in comparing basal and distal floret performance but rather the effects of basal clipping within these, we ran separate RM‐ANOVA for each position. Whole‐plot between treatment effect was clipping treatment (clipped vs. unclipped control), with the clip‐by‐replicate culm interaction as the whole‐plot F‐test as error term. We grouped the six sampling dates into three phenological stages: pre‐anthesis (June 13 and 18), anthesis (June 25 and July 2), and post‐anthesis (July 9 and 16), and used these three phenological stages as the sub‐plot effect, along with the stage‐by‐clipping interaction, using the stage‐by‐clip‐by‐replicate culm interaction as F‐test denominator. F‐test results were considered significant at associated p < 0.05; with post hoc means comparisons made using α‐adjusted least significant difference (LSD) test.
Results
3
Volumetric soil moisture (θ soil) over the course of the study followed a typical seasonal drying trend, reducing from high levels attained over early to mid‐spring rainfall (Figure 1a). These θ soil are very similar to those experienced in previous studies in this location (Hamerlynck and O'Connor 2022; Hamerlynck et al. 2023; Hamerlynck et al. 2024). Incident photosynthetic photon flux densities (PPFD; Figure 1b) were consistently above 1500 μmol m^−2^ s^−1^, well above documented photosynthetic light saturation points for crested wheatgrass seedheads (Hamerlynck and Ziegenhagen 2020). Two days following basal floret defoliation, ϕ PSII in clipped florets declined below control floret levels, then recovered to control floret levels throughout the rest of the reproductive period (Figure 1c). Florets distal to clipped florets started at ϕ PSII similar to controls, then attained higher ϕ PSII even as levels declined through the rest of the study (Figure 1d). Despite the initial difference, basal floret ϕ PSII did not differ significantly between clipped and unclipped controls pooled across the study but rather changed significantly between phenological stages (Table 1), being significantly lower over the post‐anthesis compared to similar higher ϕ PSII over the pre‐anthesis and anthesis stages (Table 2). Basal clipping resulted in significant differences between distal floret ϕ PSII (Table 1). This in part was due to higher ϕ PSII in florets distal to clipped at the end of the pre‐anthesis, and ϕ PSII in these florets being consistently higher than those above intact basal florets throughout the reproductive period (Figure 1d; Table 2), resulting in significantly higher ϕ PSII in distal to clipped (0.28 ± 0.009 SE) compared to those distal to controls pooled across the study (0.26 ± 0.009 SE; LSD < 0.05). As in basal florets, distal floret ϕ PSII were high and similar over the pre‐anthesis and anthesis stages and these were significantly higher than ϕ PSII attained over the post‐anthesis (Figure 1d; Table 2).
Reproductive period (a) volumetric soil moisture (θ soil) (b) photosynthetic photon flux density (PPFD) and light adapted PSII quantum efficiency (ϕ PSII) of (c) clipped and unclipped basal and (d) florets distal to these; PPFD and ϕ PSII are means of 32 and 16 measurements, respectively; error bars indicate ± one SE of the mean. Statistical analyses for phenological stages are: Pre‐anthesis (measurements 1 and 2), anthesis (measurements 3 and 4), and post‐anthesis (measurements 5 and 6).
TABLE 1: Repeated‐measures analysis of variance F‐test results comparing basal (direct) and distal (indirect) photochemical (F v/F m and ϕ PSII) and non‐photochemical (ϕ NPQ and ϕ NO) responses to basal floret clipping treatment over pre‐anthesis, anthesis, and post‐anthesis reproductive phenological stages. Degrees of freedom for each F‐test in parentheses, bold results significantly differ at p < 0.05.
TABLE 2: Means and standard errors of basal floret (direct) and distal floret (indirect) photochemical (F v/F m and ϕ PSII) and non‐photochemical (ϕ NPQ and ϕ NO) responses to basal floret clipping over pre‐anthesis, anthesis, and post‐anthesis reproductive phenological stages. Superscripted letters differ at p < 0.05 (LSD), bold results indicate significant clipping treatment differences within a phenological stage, italic and underlined results indicate significant differences between phenological stages within a clipping treatment.
Optimal PSII quantum yield (F v/F m) in clipped basal florets was depressed below control floret levels over the first four samples, corresponding to pre‐anthesis and anthesis stages, followed by convergence to controls over the post‐anthesis (Figure 2a). Pre‐anthesis low F v/F m was concurrent with increased ϕ NPQ in clipped basal florets (Figure 2c), while ϕ NO was at similar levels, as it was throughout the full study period (Figure 2e). Pooled across the study, there was a significant difference between clipped and unclipped basal floret F v/F m, with a significant stage‐by‐clip interaction (Table 1). Overall, clipped basal floret F v/F m (0.66 ± 0.006 SE) was lower than in unclipped counterparts (0.70 ± 0.004 SE; LSD p < 0.05). The two‐way interaction was due to: (i) significantly lower clipped basal floret F v/F m over the pre‐anthesis and anthesis stages, with significant differences over the post‐anthesis (Table 2) and (ii) increasing F v/F m going from pre‐ to post‐anthesis in clipped florets while control floret F v/F m was less variable across the three phenological stages (Table 2; Figure 2a). Seasonally pooled basal ϕ NPQ significantly differed between treatments (0.43 ± 0.019 SE for clipped and 0.41 ± 0.014 SE for controls, respectively), also with a stage‐by‐clip interaction (Table 2). This was due to pre‐anthesis basal floret ϕ NPQ being significantly higher than in control florets over the anthesis and post‐anthesis stages (Table 2) despite ϕ NPQ levels being quite close between treatments by the end of the pre‐anthesis stage (Figure 2c). There were also within‐treatment stage differences, with clipped basal florets having high ϕ NPQ during pre‐anthesis, followed by low levels during anthesis, then increasing again post‐anthesis, while control basal florets had a trend of increasing ϕ NPQ going from pre‐ to post‐anthesis (Table 2; Figure 2c). There was no significant variation in basal floret ϕ NO between clipping treatments or phenological stages across the reproductive period (Table 2; Figure 2e).
Reproductive period responses to basal floret clipping of basal florets (direct effect) and distal florets (indirect effect) optimal PSII quantum yield (F v/F m), light‐adapted PSII quantum yield (ϕ PSII), and quantum yield of regulated (ϕ NPQ) and unregulated (ϕ NO) non‐photochemical quenching. Symbols are the average 16 measurements; error bars indicate ± one SE of the mean. Statistical analyses for phenological stages are: Pre‐anthesis (measurements 1 and 2), anthesis (measurements 3 and 4), and post‐anthesis (measurements 5 and 6).
Unlike distal floret ϕ PSII, F v/F m did not significantly differ between clipping treatments but only varied significantly between phenological stages (Table 1). Distal floret F v/F m were high and similar over pre‐anthesis and anthesis stages, and these were significantly greater than F v/F m attained post‐anthesis (Figure 2b, Table 2). ϕ NPQ in distal florets were similar between basal clipping treatments and showed significant increases progressing from pre‐anthesis to post‐anthesis stages (Figure 2d; Table 2). In contrast to basal florets, ϕ NO in distal florets differed significantly between basal floret clipping treatments (Table 1). Florets distal to clipped basal florets had significantly lower ϕ NO (0.30 ± 0.011 SE) compared to those distal to controls (0.35 + 0.016 SE) pooled across the study. The higher ϕ NO in distal to control florets apparent pre‐ and post‐anthesis (Figure 2f; Table 2) likely drove the significant differences in ϕ NO between the phenological stages (Table 1).
Discussion
4
In both basal and distal florets, F v/F m was below 0.75, a level indicative of the presence of priming and engagement of xanthophyll‐cycle mediated photoprotective thermal dissipation of excess absorbed PSII energy (Demmig‐Adams and Adams III 1992; Osmond 1994). Higher ϕ NPQ and lower F v/F m in clipped basal florets compared to those in unclipped controls are highly suggestive of high engagement of regulated photoprotective NPQ. These findings partially support our hypothesis that clipping would result in higher NPQ and down‐regulation of PSII quantum efficiency and suggest Hamerlynck et al. (2023) conjecture was correct, that floral defoliation induced down‐regulation of PSII photochemical function was mediated by non‐photochemical photoprotective mechanisms. However, contrary to our expectations and in contrast to findings by Hamerlynck et al. (2023) and Quigley et al. (2024), there was no evident down‐regulation of light‐adapted ϕ PSII in clipped basal florets. This may be due to differences in actinic light sources; our current study assessed ϕ PSII under full spectrum sunlight, while our previous results were obtained with an instrument using an artificial red/blue actinic light source for concurrent gas exchange and chlorophyll fluorescence measurements. Quantum yield can vary in response to both light quality and quantity (Hogewoning et al. 2012; Jokic et al. 2025), and the spectral differences in actinic light sources between our studies may have affected the attained ϕ PSII.
Lower F v/F m and higher ϕ NPQ (Figure 2a,c) in clipped basal florets were not always concurrent with decreased ϕ PSII; indeed, ϕ PSII was generally similar between clipped and unclipped controls (Figure 1c; Table 2). ϕ NPQ does not reflect the magnitude of NPQ engagement, but rather the proportion of absorbed light energy diverted safely away from PSII reaction centers, and ϕ PSII is considered a more sensitive indicator of physiological stress than F v/F m (Klughammer and Schreiber 2008). Lower F v/F m in clipped florets suggests stronger engagement and slower reversing of regulated NPQ, possibly due to greater regulatory involvement by the thylakoid‐bound PsbS protein (Li et al. 2002). Regardless, the degree of NPQ engagement was probably not sustained enough to reduce subsequent light‐adapted ϕ PSII. In a compensatory context, maintaining ϕ PSII makes sense, as electron transport rates would not be under additional photoprotective restraint, and up‐regulation of photosynthetic activity could be more readily attained. In addition, it may be that reproductive post‐defoliation physiological processes may not follow an obligate post‐defoliation dynamic and differ between species and environmental conditions. Hamerlynck et al. (2023) made measurements after an exceptionally large rain during the pre‐anthesis period, while Quigley et al. (2024) examined reproductive compensatory photosynthetic responses of bluebunch wheatgrass, which has distinctly different reproductive photosynthetic characteristics to crested wheatgrass (Hamerlynck et al. 2019; Hamerlynck and Ziegenhagen 2020). Therefore, interannual variation in reproductive period environmental conditions may affect the timing and magnitude of floral defoliation photosynthetic responses.
There were distinct indirect responses to clipping of basal florets apparent in the photochemical and non‐photochemical processes in florets distal to them. While F v/F m and ϕ NPQ did not differ in distal florets between clipping treatments, ϕ PSII and ϕ NO did, with higher overall ϕ PSII and lower ϕ NO in florets above clipped florets compared to those distal to controls (Figure 2 and Table 2). ϕ NO reflects constitutive PSII non‐photochemical processes (Klughammer and Schreiber 2008). High light exposure increases the rate of photochemical reactions, increasing the reduction state of the photosynthetic electron transport chain, principally at the primary electron acceptor, Q_a_, though other electron transport molecules can be affected as well, such that decreases in ϕ PSII are compensated for by an increase in ϕ NO (Samson et al. 2019). We found clipping of basal florets induced relaxation of PSII redox conditions in their distal florets such that ϕ PSII was maintained at modest but consistently higher levels than those distal to unclipped ssflorets (Figure 1d). Lower ϕ NO and higher ϕ PSII suggest smoother electron transfer through the entire photosynthetic electron transport system, but our results do not provide definitive insight as to the mechanism(s) by which this is attained. Photorespiration is an alternative electron transport system intimately linked to enzymatically mediated nitrogen dynamics and can affect gas exchange dynamics and NPQ, including ϕ NO (Heber et al. 1996; El‐Khatib et al. 2004; Proctor and Smirnoff 2011; Zhong et al. 2022). Higher ϕ NO expression over the pre‐ and post‐anthesis stages in florets where intra‐seed‐head competition between basal and distal florets is intact (Warringa et al. 1998) might reflect competition for assimilated nitrogen affecting photorespiratory processes. This is an attractive idea in that photorerespiratory metabolites associated with high NPQ capacity such as H_2_O_2_ can affect subsequent herbivore behavior (Jänkäpää et al. 2013). However, the accumulation of photorespiratory and other photoprotective metabolites are usually associated with increased ϕ NO (Proctor and Smirnoff 2011; Zhong et al. 2022), not its relaxation, as expressed in florets distal to clipped basal florets.
Another possibility is that the relaxation of ϕ NO is related to carbon assimilation. Hamerlynck et al. (2023) suggested carbon assimilates from damaged basal florets supplemented those assimilated by unaffected distal florets, thereby enhancing their reproductive effort. Over the pre‐anthesis stage this supplementation could help support a larger number of viable ovules, much as the flag leaf does in crested wheatgrass (Hamerlynck et al. 2024). This exogenous supply could reduce the endogenous demand of these, thereby relaxing redox pressures to electron transport. The ϕ NO relaxation with higher ϕ PSII over the post‐anthesis stage could also support higher net photosynthetic rates (A_net_) during energetic provisioning. Hamerlynck et al. (2023) observed slightly elevated post‐anthesis distal A_net_ but conjectured most of the carbon that increased distal seedhead specific mass came from damaged basal florets. While our results don't negate this supposition, they do suggest that photosynthetic activity by the distal florets themselves could be enhanced and contribute more to seed provisioning than previously thought.
In conclusion, this study highlights the importance of establishing reproductive ecophysiological processes, as these often serve as the origin point of population and community dynamics (Bazzaz 1979; Bazzaz et al. 1979). Moreover, these results are consistent with others showing drought and herbivory pressures produce co‐adaptive ecological strategies, especially in aridland bunchgrasses (Adler et al. 2004; Quiroga et al. 2010). This study and others we have made establish crested wheatgrass as possessing a remarkably integrated reproductive photosynthetic apparatus that facilitates its ability to produce seeds more capable of overcoming environmentally imposed demographic bottlenecks than native bunchgrasses (James et al. 2011; Hamerlynck and Davies 2019; Hamerlynck et al. 2019; Hamerlynck and O'Connor 2021; Hamerlynck et al. 2023; Quigley et al. 2023; Hamerlynck et al. 2024). Additionally, recent work has sought to formulate and implement seed‐based restoration strategies based on Grubb's regeneration niche concept (Grubb 1977; Larson et al. 2023, 2025). Our study suggests such approaches could be refined and improved with information regarding the environmental and biological constraints to parental‐plant reproductive ecophysiological performance.
Funding
This work was supported by the Agricultural Research Service and Oregon State Agricultural Experiment Station.
Conflicts of Interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Adler, P. B. , D. G. Milchunas , W. K. Lauenroth , O. E. Sala , and E. C. Burke . 2004. “Functional Traits of Graminiods in Semi‐Arid Steppes: A Test of Grazing Histories.” Journal of Applied Ecology 41: 653–663. 10.1111/j.0021-8901.2004.00934.x. · doi ↗
- 2Barton, K. E. 2016. “Low Tolerance to Simulated Herbivory in Hawaiian Seedlings Despite Induced Changes in Photosynthesis and Biomass Allocation.” Annals of Botany 117: 1053–1062. 10.1093/aob/mcw 021.27056973 PMC 4866310 · doi ↗ · pubmed ↗
- 3Bazzaz, F. A. 1979. “The Physiological Ecology of Plant Succession.” Annual Review of Ecology and Systematics 10: 351–371. 10.1146/annurev.es.10.110179.002031. · doi ↗
- 4Bazzaz, F. A. , R. W. Carlson , and J. L. Harper . 1979. “Contribution to Reproductive Effort by Photosynthesis of Flowers and Fruits.” Nature 279: 554–555. 10.1038/279554 a 0. · doi ↗
- 5Demmig‐Adams, B. , and W. W. Adams III . 1992. “Photoprotection and Other Responses of Plants to High‐Light Stress.” Annual Review of Plant Physiology and Plant Molecular Biology 43: 599–626. 10.1146/annurev.pp.43.060192.003123. · doi ↗
- 6Derks, A. , K. Schaven , and D. Bruce . 2015. “Diverse Mechanisms for Photoprotection in Photosynthesis. Dynamic Regulation of PSII Excitation in Response to Rapid Environmental Change.” Biochimica et Biophysica Acta 1847: 468–485. 10.1016/j.bbabio.2015.02.008.25687894 · doi ↗ · pubmed ↗
- 7El‐Khatib, R. T. , E. P. Hamerlynck , F. Gallardo , and E. G. Kirby . 2004. “Transgenic Poplar Characterized by Ectopic Expression of a Pine Cytosolic Glutamine Synthetase Gene Exhibit Enhanced Tolerance to Water Stress.” Tree Physiology 24: 729–736. 10.1093/treephys/24.7.729.15123444 · doi ↗ · pubmed ↗
- 8Genty, B. , J. M. Brantais , and N. R. Baker . 1989. “The Relationship Between the Quantum Yield of Photosynthetic Electron Transport and Quenching of Chlorophyll Fluorescence.” Biochemica et Biophysica Acta 990: 87–92. 10.1016/S 0304-4165(89)80016-9. · doi ↗