Targeting surface cell antigen 2 increases sensitivity of Rickettsia typhi detection
Weerawat Phuklia, Kaisone Padith, Koukeo Phommasone, Vilada Chansamouth, Mayfong Mayxay, Susath Vongphachanh, Paul N. Newton, Matthew T. Robinson, Elizabeth A. Ashley

TL;DR
A new qPCR test targeting the sca2 gene improves detection of Rickettsia typhi, which causes murine typhus, especially in cases with low bacterial levels.
Contribution
A novel qPCR assay targeting the repetitive sca2 gene increases sensitivity for detecting Rickettsia typhi in patient blood.
Findings
The sca2 qPCR assay showed 59.09% sensitivity compared to 36.36% for the ompB assay.
DNA copy numbers detected by sca2 were 2.933 log units higher than those detected by ompB.
The sca2 assay is more effective for detecting low concentrations of R. typhi in patient samples.
Abstract
Murine typhus is a flea-borne disease caused by Rickettsia typhi that typically presents as an acute febrile illness. The diagnosis is often missed, leading to delays in appropriate treatment. A qPCR targeting a single gene (ompB) for R. typhi is widely used for diagnosis; however, it has low sensitivity for detecting bacterial DNA in patients’ blood. We aimed to increase sensitivity of detection of R. typhi using qPCR by targeting a gene containing repetitive sequences, sca2 (surface cell antigen 2). We compared diagnostic accuracy with the standard assay targeting single sequence ompB (outer membrane protein B). Specificity, sensitivity, and bacterial load measurement of both assays were compared using stored EDTA-anticoagulated buffy coat samples from 88 patients with febrile illness at Mahosot Hospital or provincial hospitals in Laos. Among these, 55 cases were confirmed as…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Fig 1
Fig 1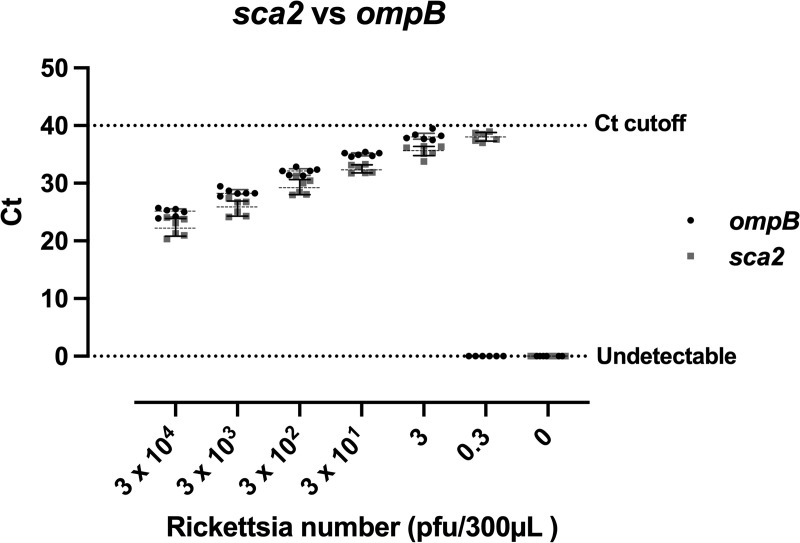 Fig 2
Fig 2 Fig 3
Fig 3- —http://dx.doi.org/10.13039/100010269Wellcome Trust
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVector-borne infectious diseases · Leptospirosis research and findings · Salmonella and Campylobacter epidemiology
Introduction
Rickettsia typhi is an obligate intracellular bacterium that causes the neglected disease murine typhus. Both the rat flea (Xenopsylla cheopis) and the cat flea (Ctenocephalides felis), play a role in transmitting the disease [1]. The disease has a worldwide distribution, including in the Lao PDR (Laos) [2]. Although the disease is frequently mild and responds to key antibiotics, the mortality rate in untreated patients is approximately 0.4% [3]. Misdiagnosis often leads to inappropriate treatment. In the absence of a single gold diagnostic standard, a combination of serology and PCR performed on acute samples is widely accepted for the diagnosis of murine typhus. However, some serological methods cannot provide results quickly enough for early clinical decision-making [4–6]. In contrast, quantitative PCR (qPCR) targeting R. typhi-specific genes is a valuable tool for early diagnosis, as it can detect bacterial DNA in acute patient samples [7]. The outer membrane protein B (ompB) gene is commonly used as a target for R. typhi detection [8]. However, it targets a single-copy region of the gene and its sensitivity may be limited, especially in patients with a low bacterial load [9]. In the case of Orientia tsutsugamushi, another obligate intracellular bacterium, targeting repetitive gene sequences has been shown to improve diagnostic sensitivity [10]. We therefore proposed applying a similar strategy for murine typhus.
Surface cell antigen (sca) proteins belong to the autotransporter protein family in Rickettsia. The R. typhi genome encodes five Sca proteins; sca1, sca2, sca3, sca4 and sca5. These proteins are essential for bacterial survival. Although R. typhi contains relatively few repetitive sequences in its genome [11], the sca2 gene has six tandem repeats within a 150-base-pair region. This protein may facilitate bacterial adhesion to host cells [12]. Given its essential role in R. typhi’s intracellular replication, the sca2 gene represents a promising new target for diagnostic assays. In this study we aimed to develop a qPCR assay for R. typhi detection by targeting a gene region containing repetitive sequences (sca2) and to compare its performance for detection of R. typhi with the existing qPCR assay that targets ompB.
Methods
Ethics statement
These samples were part of previous studies that were approved by the National University of Laos, National Ethics Committee for Health Research (NECHR) and National Institute of Public Health (NIOPH), Vientiane and Oxford Tropical Research Ethics Committee (OxTREC), Faculty of Medical Sciences. For all studies, written informed consent was obtained from all adult participants and from parents or legal guardians of child participants prior to sample collection.
Assay development
Searching for repetitive sequences.
The sca2 gene, encoding the 190-kDa surface cell antigen (sca2) of Rickettsia typhi strain Wilmington, was identified from the complete genome sequence (GenBank accession number AE017197.1) at genomic positions 68,152–72,603 and corresponds to locus tag RT0052. To investigate repetitive sequence within the R. typhi sca2 gene, the full length of nucleotide sequence (4,452 bp) was submitted to the Tandem Repeat Finder (https://tandem.bu.edu/trf/trf.basic.submit.html). The sequence was analyzed as described [13] and four regions containing repetitive sequences are illustrated in S1 Fig. The region with the highest number of repetitive sequences was selected as the target for PCR detection. Primers were designed to amplify this region using Primer 3 (https://bioinfo.ut.ee/primer3-0.4.0/), as shown in S1 Fig. PCR amplification using these primers generates products whose size may vary depending on the number of repeat copies in the target region. The primer sequences were sca2F 5’- TGGAATGGACAGTAAAACGACAG-3’ and sca2R 5’- CGTTCTGCTGCCTCTTCTGA-3’.
Determination of assay specificity.
To test the specificity of the selected primers, stored DNA from several strains of cultured R. typhi (Wilmington, AZ331 and GER), as well as DNA from other bacteria including Rickettsia prowazekii, Rickettsia felis, Rickettsia conorii, O. tsutsugamushi, Burkholderia thailandensis, Burkholderia cepacia, Anaplasma phagocytophilum, Leptospira spp., and Neorickettsia sennetsu were subjected to PCR and the bands were observed using 2% agarose gel electrophoresis.
R. typhi quantification by quantitative real time PCR (qPCR).
DNA from R. typhi culture or patients’ buffy coat was extracted using GeneJet Genomic DNA purification kit (ThermoFisher Scientific, UK), following the manufacturer’s instructions. The DNA copy number of R. typhi was measured using qPCR targeting either sca2 or ompB. The primers for sca2 were described above, and the Taqman probe for the sca2 gene was sca2P: 5’-FAM- ACAGATAACATAGCAGCAGAATCT-BHQ1–3’. The primers and Taqman probe for ompB were Rt557F: 5’- TGGTATTACTGCTCAACAAGCT-3’, Rt678R: 5’-CAGTAAAGTCTATTGATCCTACACC-3’ and Rt640 BP: 5’-FAM- CGCGATCGTTAATAGCACCAGCATTATCGCG -BHQ1–3’. The qPCR mixture was composed of 1X qPCRBIO Probe Mix (qPCR Probe MIX LO-ROX, PCR Biosystems, UK), 0.4 μM forward and reverse primers, 0.2 μM probe, sterile distilled water and 1μL of extracted DNA. The sca2 qPCR assay was performed in the presence of 0.4 μg/mL bovine serum albumin (BSA) per reaction to improve amplification efficiency. BSA helps stabilize the DNA polymerase and mitigate the inhibitory effects of secondary structures often present in repetitive sequences, enhancing reproducibility and sensitivity of the assay [14,15]. Real-time PCR was performed on a CFX96 real-time PCR detection system (Bio-Rad Laboratories) using the following conditions: initial denaturation at 95°C for 2 min, followed by 45 cycles of denaturation at 95°C for 15 sec and combined annealing and extension at 60°C for 30 sec with the acquisition of fluorescence. Rickettsial DNA copies per ml of blood were calculated using 10-fold serial dilution of known concentrations of R. typhi sca2 and ompB fragments cloned into the pGEM-T Easy vector (ranging from 10^6^ to 10^0^ copies/μL).
Limit of detection (LOD) for qPCR assays using R. typhi spiked blood.
Limit of detection (LOD) is defined as the minimum number of DNA copies in a sample that can be reliably detected with the assay. We used the plaque assay, the gold standard for Rickettsia quantification [16], to determine rickettsial load in terms of plaque forming units (pfu). R. typhi strain Wilmington was used to spike whole blood collected from healthy donors (n = 2). Purified R. typhi was serially diluted ten-fold and spiked into blood samples. Specifically, 30μL of media containing R. typhi at concentrations ranging from 10^1^-10^6^ pfu/mL was added to 270 μL of blood. The resulting 300 μL mixture containing R. typhi ranging from 3 x 10^-1^ to 3 x 10^4^ pfu/total blood was incubated at room temperature for 30 minutes, after which DNA was extracted from 300 μL of blood using GeneJet Genomic DNA Purification Kit as described by manufacturer’s instruction (ThermoFisher Scientific, UK). R. typhi DNA in each dilution was quantified using both the ompB and sca2 qPCR assays.
Diagnostic accuracy testing
Clinical samples.
Three groups of stored buffy coat samples were used to validate and compare the specificity and sensitivity of the sca2 and ompB assays. Relevant studies included “A multicentre, open-label randomized trial comparing 3-day doxycycline, 7-day doxycycline, and 3-day azithromycin for scrub and murine typhus” (OxTREC no. 003–03 and 1413/FMS), “The pathophysiology of typhus in Laos” (OxTREC no. 015–04 and 010/NECHR) and “A cross-sectional prospective study to identify the pathogenicity of acute non-malaria febrile illness” (OxTREC no. 015–10 and 134/NECHR). Samples in this group were confirmed by indirect immunofluorescence assay (IFA) and ompB PCR from cell culture or by ompB PCR from buffy coat. The second group of samples came from patients confirmed to have murine typhus (n = 10) from a fever surveillance study in three provinces in Laos (OxTREC no. 027–14 and 026 NIOPH/NECHR). Diagnosis in this group was confirmed by IFA using paired sera (acute and convalescent). The third group consisted of buffy coat samples from patients (n = 17) who tested negative for murine typhus RDT (ImmunoDOT Rickettsia typhi test (GenBio)) and scrub typhus by RDTs (Scrub Typhus Detect IgM Rapid System (Dipstick) (InBios)) in a prospective study of the causes of fever amongst hospitalized patients in Lao PDR (Tables 1 and S1).
Table 1: Repetitive sequence regions of sca2 gene investigation.
DNA extraction.
DNA was freshly extracted from the frozen buffy coat samples using the GeneJet Genomic DNA Purification Kit (ThermoFisher Scientific, UK), following the manufacturer’s instructions. Briefly, 200 μl of the buffy coat was lysed and precipitated with ethanol, and the mixture was passed through a spin column for DNA binding. The final elution volume was 100 μl per sample. DNA was stored at −80°C for long-term preservation and at 4°C between qPCR runs.
Data analysis
The sensitivity and specificity of the qPCR assay targeting ompB and sca2 were calculated. Statistical significance was tested by Fisher’s exact test. The 95% confidence interval of sensitivity and specificity were calculated using Wilson-Brown method. As the qPCR copy number data were not normally distributed and included outliers, median and interquartile range (IQR) of Rickettsial loads determined by both assays was compared using Wilcoxon test. Graphical representations were calculated and generated using GraphPad Prism (version 10).
Results
Repetitive sequences on R. typhi sca2 sequence
Repetitive sequences were identified at four distinct regions along the full-length R. typhi sca2 gene. These regions contain approximately 2.3, 1.8, 2.1 and 6 copies repetitive elements, respectively, with varying sequence lengths, as summarized in Table 1. Among these, the region located at position 2360–3262 had the highest number of repeats (6 copies) and the largest sequence size. Due to its high repeat number and sequence length this region was selected as the target for qPCR assay development.
Specificity testing
To evaluate the specificity of the designed primers for R. typhi, we first conducted in silico analysis comparing R. typhi strains to other Rickettsia species. The analysis demonstrated that the primers were specific to R. typhi genomes and did not amplify sequences from other species, including R. prowazekii.
We then validated these findings experimentally by performing conventional PCR using DNA extracted from R. typhi strains Wilmington, AZ331, and GER, as well as from other bacterial species. The result confirmed that sca2 assay specifically amplified DNA from R. typhi strains, with no amplification observed in non-R. typhi samples (S2 Fig).
Optimization of qPCR targeting sca2 assay and ompB
To determine the limit of detection (LOD) for both qPCR assays, serial 10-fold dilutions (10^6^ to 10^0^ copies/μL) of R. typhi sca2 and ompB fragments cloned into pGEM-T Easy vector were used. The result showed that the LOD for the ompB assay was 10 copies/μL, while the sca2 assay demonstrated higher sensitivity with a LOD of 1 copy/μL (Fig 1).
Standard curve for Rickettsia typhi qPCR assay.Assay used plasmid (pGEM-Teasy) carrying ompB or sca2 fragment of R. typhi as target sequences, with concentrations ranging from 1 to 106 copies/μL. The line with the blank triangle represents patient samples.
Limit of detection for qPCR assays targeting ompB or sca2 using R. typhi spiked blood
We also compared the limit of detection between ompB and sca2 assays by spiking whole blood with partially purified R. typhi. The result showed that qPCR using ompB was able to detect R. typhi 3 pfu/ 300 μL of whole blood whereas sca2 was able to detect R. typhi 0.3 pfu/mL as demonstrated in Fig 2 and raw data for Cycle threshold (Ct) value are provided in S1 Data.
Limit of detection of sca2 and ompB qPCR assay.R. typhi ranging from 0.3 to 3 x 104 PFU of 30 μL of culture media containing R. typhi was spiked into 300 μL of whole blood from two healthy donors. Whole blood without R. typhi was included as the 0-PFU control. R. typhi was detected using different two assays targeting the sca2 (grey) and ompB (black) genes. Each dot represents a Ct value obtained from qPCR for sca2 or ompB. Dashed lines indicate the median with interquartile range (IQR) as error bars. Experiments for each gene were performed in triplicate per donor. The horizontal dashed line at Ct = 40 indicates the positivity cut-off (Ct ≤ 40).
Sensitivity and specificity testing of sca2 and ompB
Clinical samples were subjected to both qPCR assays to compare their sensitivity and specificity. Raw qPCR Ct values and IFA results are provided in S1 Data. The results showed that the sensitivity of sca2 assay was 59.09% (95% CI, 38.73-76.74%) and specificity was 100% (95% CI, 93.98-100) whereas the sensitivity of the ompB assay was 36.36% (19.73-57.05%) and specificity was 100% (95% CI, 93.98-100%) (Table 2)
Table 2: Sensitivity and specificity of sca2 and ompB qPCR assays testing.
Rickettsial target gene copy number determination from positive R. typhi buffy coat by ompB PCR
In order to compare rickettsial target gene copy number in buffy coat detected by sca2 and ompB PCR, we calculated the copy number for both targets using R. typhi sca2 or ompB fragments cloned into the pGEM-T Easy vector at concentrations ranging from 10^0^ to 10^6^ copies/μL. The median rickettsia target gene copy number detected using sca2 gene was 16,500 copies/μL (IQR, 13,045–40,000), whereas for the ompB assay and it was 19.25 copies/μL (IQR, 11.11-56.62) (Fig 3). Raw data of rickettsia target gene copy number are provided in S1 Data.
Comparison of Rickettsia target gene copy number using qPCR targeting ompB and sca2.Bacterial DNA from buffy coat samples (n = 13) were determined using ompB assay (grey) and sca2 assay (black). Grey and black bar represent the median Rickettsia target gene copy numbers with interquartile range (IQR). Difference between assays were compared using Wilcoxon signed-rank test. A p-value <0.05 was considered statistically significant.
Discussion
This study developed a qPCR assay targeting the sca2 gene, which contains six repetitive sequences, for the detection of R. typhi. Unlike the single-copy ompB gene, this assay leverages multiple template copies per genome, enhancing analytical sensitivity for detecting low-level bacteremia commonly observed in murine typhus patients (approximately 200 copies/μL) [9]. A multiple-copy gene approach has been used for detection of various pathogens, including Plasmodium falciparum and Plasmodium vivax in asymptomatic infections [17]. Chao et al also developed a qPCR assay targeting multiple genes to detect fifteen different strains of O. tsutsugamushi isolates and in patient samples [10]. Although multiple gene copies are not found in the R. typhi genome, genes containing repetitive sequences have been identified [18]. In this study, we applied a multiple-target approach by designing primers and a probe to bind all six repetitive sequences within the sca2 gene. This enabled the primers and probe to bind six times, compared to targeting a single-copy gene. Currently, qPCR assays for R. typhi detection in clinical samples typically target single sequence such as ompB. In our in vitro experiments using plasmid containing fragments of sca2 and ompB, the sca2-based assay could detect DNA at 10-fold lower concentrations than the ompB-based assay (1copy/μL versus 10 copies/μL). In spiked samples, serial dilutions of R. typhi showed that the sca2 assay had lower limit of detection (0.3 PFU) compared to ompB (3 PFU), indicating higher sensitivity. These findings were supported by accuracy testing using buffy coat samples with known R. typhi infection status. Another study used the parvulin-type PPIase (prsA) genes as the qPCR target and reported 10-fold greater sensitivity than ompB [7]. However, prsA testing was not evaluated in this study. Other Rickettsia genes, such as citrate synthase (gltA) and 17 kDa lipoprotein outer membrane antigens (17-kDa), were not evaluated in this study, as both assays were commonly used for genus-level screening; ompB and sca2 were selected for species-specific detection [19]. Quantification of rickettsial target gene copy number using sca2 and ompB assays revealed significantly higher bacterial copy numbers when using the sca2-based assay. This suggests that sca2 is a valuable target for detecting R. typhi in patients with low bacterial loads and for monitoring bacteria clearance in antibiotic treatment studies. Interestingly, the rickettsia target gene copy number detected by sca2 was approximately 800-fold higher than that determined by ompB, despite theoretical expectation of only a six-fold increase based on repeat copy number. Several factors may explain this discrepancy. First, the TaqMan probe can cleave a new probe during each PCR cycle, generating fluorescent signal more rapidly [20]. Second, secondary structures within the repeat sequences may make the amplicon more accessible, enhancing primer and probe binding [21]. Third, repeat sequences may amplify more efficiently than single-copy targets due to thermodynamic advantages [22]. Finally, in genes with tandem repeats, primers and probes can potentially rebind multiple times during extension, further increasing fluorescence [23].
Although sca2 is more sensitive than ompB and can detect lower copy numbers in clinical samples with a low bacterial burden, the cost is not expected to be higher, as the assay differs only in primer sequences and uses the same probe labeling chemistry (5′ FAM and 3′ BHQ1). Cost would increase only if longer oligonucleotides are required.
The sca2 qPCR assay is feasible for use in the tested population and can be applied alongside the ompB assay. Since murine typhus patients typically have low bacterial loads, ompB-based qPCR may fail to detect low levels of DNA. In such cases, sca2 qPCR serves as a complementary target, improving diagnostic confidence.
Murine typhus patients are usually treated with doxycycline after rapid antibody-based testing, before qPCR confirmation. This does not differentially affect qPCR reaction [24], as antibiotic treatment reduces bacterial load and impacts detection of all targets similarly. Therefore, improved detection with sca2 reflects enhanced analytical sensitivity at low bacterial loads rather than a gene-specific effect. In practice, qPCR positivity depends on the timing of sample collection relative to antibiotic initiation and the initial bacterial burden [25–27]. In this study, samples were collected at presentation or shortly after antibiotic therapy, reflecting routine diagnostic conditions.
This study had several limitations. Specificity testing was limited, as DNA from all Rickettsia spp. was not available. The number of clinical samples used for assay validation was small. The developed assay was optimized for detecting R. typhi in buffy coat samples, but not for other sample types or sources, including vectors and reservoirs. Additionally, only two assays were evaluated, and other genes with repeat sequences-such as those encoding ankyrin repeat-containing proteins, patatin-like phospoholipases were not included for comparison with sca2. Finally, rickettsial target gene copy number determined by sca2 and ompB were calculated only from qPCR-positive samples, and thus do not represent all patients who tested positive by the gold standard method (IFA).
Conclusion
We optimized a qPCR targeting sca2 gene, which contains repetitive sequences for diagnosing R. typhi infection in humans. The assay is specific to R. typhi and does not cross-react with other Rickettsia species or a panel of other bacteria. In vitro studies using plasmid standards and bacteria spiked into human blood showed that the sca2 assay had a limit of detection approximately 10-fold lower than that of ompB, the current commonly used qPCR target for R. typhi detection. This finding was supported by the sensitivity and specificity testing using patients’ stored buffy coat samples. Moreover, bacteria quantification using sca2 yielded approximately 2.9 log higher values compared to ompB assay. Overall, the qPCR assay targeting sca2 is a promising tool for diagnosis R. typhi in patients, who typically have low bacterial loads.
Future work will focus on optimizing the sca2 assay for different sample types, including vectors and reservoirs, to support surveillance studies, as well as validating the assay with a larger set of clinical samples. We will also investigate other target genes containing repetitive sequences and compare their performance with sca2 and ompB. Finally, we aim to use the selected target to develop a CRISPR-Cas–based lateral flow diagnostic assay and validate it with prospective patient samples.
Supporting information
S1 TableList of stored buffy coat samples for sensitivity and specificity testing.Buffy coat samples with confirmed rickettsial infection by at least one method from culture or indirect immunofluorescence assay (IFA) or Rapid Diagnostic Test (RDT) or qPCR.(DOCX)
S1 FigSchematic of the sca2 gene structure containing four regions containing repetitive sequences (green, yellow and blue) and the binding site of specific primer.(DOCX)
S2 FigGel electrophoresis analysis of DNA from sixteen laboratory strains of R. typhi and other bacteria using sca2.(DOCX)
S1 DataDataset.Raw data supporting the findings of this study. This dataset contains raw qPCR Ct values for sca2 and ompB, IFA titres (IgM and IgG) for patients confirmed by IFA, Ct values from spiked blood experiments, and bacterial loads measured by sca2 and ompB. Each type of data is provided in separate sheet within Excel file.(XLSX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Civen R, Ngo V. Murine typhus: an unrecognized suburban vectorborne disease. Clin Infect Dis. 2008;46(6):913–8. doi: 10.1086/527443 18260783 · doi ↗ · pubmed ↗
- 2Mayxay M, Castonguay-Vanier J, Chansamouth V, Dubot-Pérès A, Paris DH, Phetsouvanh R, et al. Causes of non-malarial fever in Laos: a prospective study. Lancet Glob Health. 2013;1(1):e 46-54. doi: 10.1016/S 2214-109X(13)70008-1 24748368 PMC 3986032 · doi ↗ · pubmed ↗
- 3Doppler JF, Newton PN. A systematic review of the untreated mortality of murine typhus. P Lo S Negl Trop Dis. 2020;14(9):e 0008641. doi: 10.1371/journal.pntd.0008641 32925913 PMC 7515178 · doi ↗ · pubmed ↗
- 4Lokida D, Sudarmono P, Kosasih H, Butar-Butar DP, Salim G, Antonjaya U, et al. Comparison of commercial enzyme-linked immunosorbent assay and immunofluorescence assay for diagnosis of acute Rickettsia typhi infections. Vector Borne Zoonotic Dis. 2020;20(2):93–9. doi: 10.1089/vbz.2019.2451 31448989 PMC 7041318 · doi ↗ · pubmed ↗
- 5Dhawan S, Robinson MT, Stenos J, Graves SR, Wangrangsimakul T, Newton PN, et al. Selection of Diagnostic Cutoffs for Murine Typhus Ig M and Ig G Immunofluorescence Assay: A Systematic Review. Am J Trop Med Hyg. 2020;103(1):55–63. doi: 10.4269/ajtmh.19-0818 32274984 PMC 7356422 · doi ↗ · pubmed ↗
- 6Phetsouvanh R, Thojaikong T, Phoumin P, Sibounheuang B, Phommasone K, Chansamouth V, et al. Inter- and intra-operator variability in the reading of indirect immunofluorescence assays for the serological diagnosis of scrub typhus and murine typhus. Am J Trop Med Hyg. 2013;88(5):932–6. doi: 10.4269/ajtmh.12-0325 23478577 PMC 3752761 · doi ↗ · pubmed ↗
- 7Papp S, Rauch J, Kuehl S, Richardt U, Keller C, Osterloh A. Comparative evaluation of two Rickettsia typhi-specific quantitative real-time PC Rs for research and diagnostic purposes. Med Microbiol Immunol. 2017;206(1):41–51. doi: 10.1007/s 00430-016-0480-z 27696011 · doi ↗ · pubmed ↗
- 8Henry KM, Jiang J, Rozmajzl PJ, Azad AF, Macaluso KR, Richards AL. Development of quantitative real-time PCR assays to detect Rickettsia typhi and Rickettsia felis, the causative agents of murine typhus and flea-borne spotted fever. Mol Cell Probes. 2007;21(1):17–23. doi: 10.1016/j.mcp.2006.06.002 16893625 · doi ↗ · pubmed ↗