In vitro biofilm formation by a beneficial bacterium partially predicts in planta protection against rhizosphere pathogens
Yang Liu, Alexandra D Gates, Zhexian Liu, Quinn Duque, Sierra S Schmidt, Melissa Y Chen, Corri D Hamilton, George A O’Toole, Cara H Haney

TL;DR
This study explores how biofilm formation by a beneficial soil bacterium affects its ability to protect plants from pathogens.
Contribution
The study reveals that biofilm formation in vitro does not fully predict in planta protection against pathogens.
Findings
Only a subset of biofilm-related mutants of P. brassicacearum WCS365 lost protection against the pathogen N2C3.
Mutants defective in Lap biosynthesis, flagellar function, and O-antigen biosynthesis failed to protect plants.
Biofilm formation on abiotic surfaces does not fully correlate with pathogen suppression in planta.
Abstract
Plant roots form associations with beneficial and pathogenic soil microorganisms. Although members of the rhizosphere microbiome can protect against pathogens, the mechanisms are poorly understood. We hypothesized that the ability to form a biofilm on the root surface is necessary for the exclusion of pathogens; however, it is not known if the same biofilm formation components required in vitro are necessary in vivo. Pseudomonas brassicacearum WCS365 is a beneficial strain that is phylogenetically closely related to an opportunistic pathogen Pseudomonas sp. N2C3 and confers protection against N2C3 in the rhizosphere. We used this plant-mutualist-pathogen model to screen collections of P. brassicacearum WCS365 increased attachment mutants (iam) and surface attachment defective (sad) transposon insertion mutants that form increased or decreased biofilm on abiotic surfaces, respectively.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
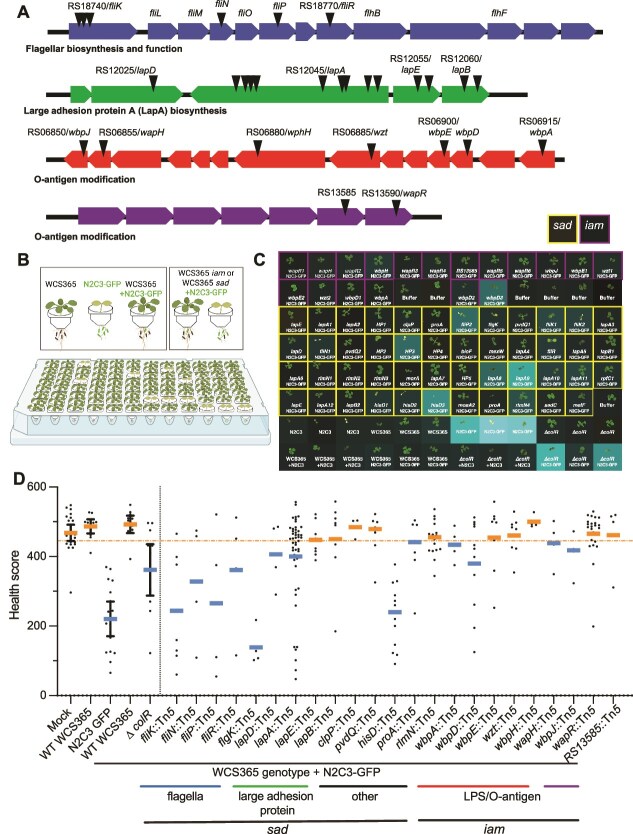 Figure 1
Figure 1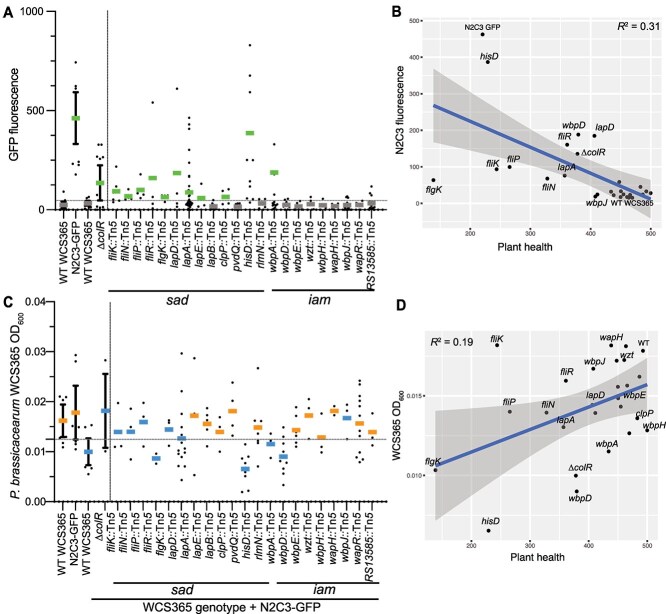 Figure 2
Figure 2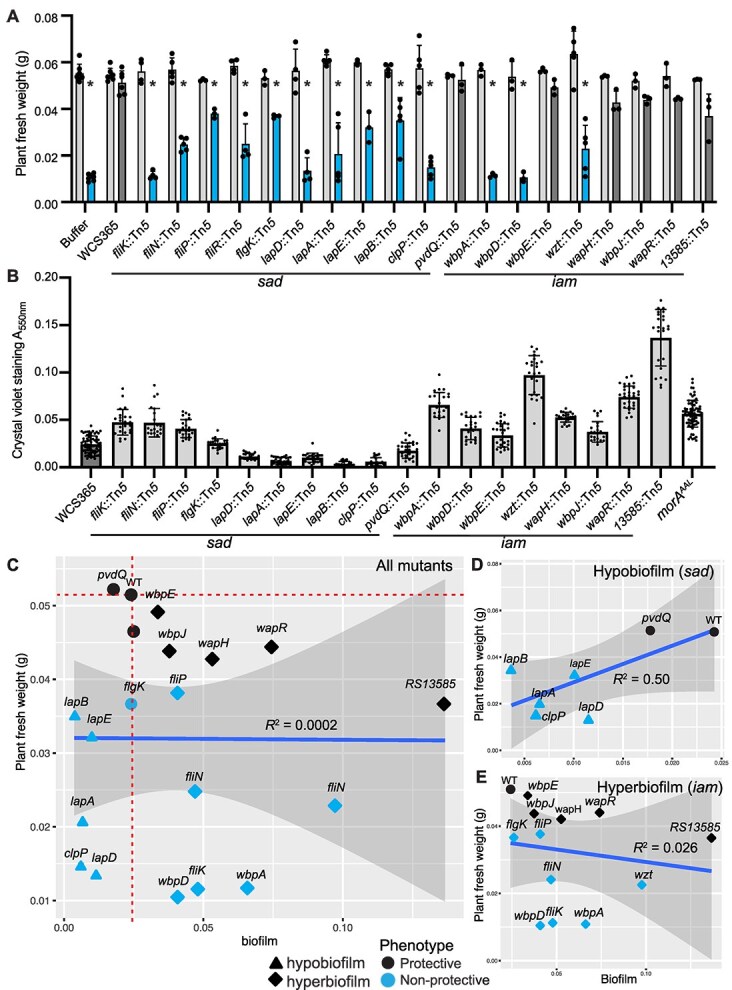 Figure 3
Figure 3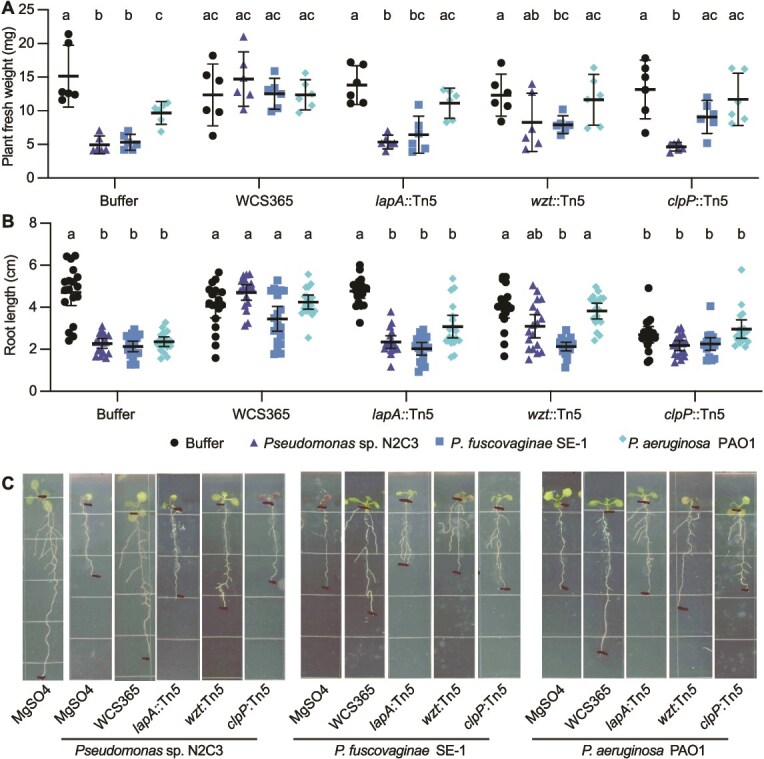 Figure 4
Figure 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant-Microbe Interactions and Immunity · Legume Nitrogen Fixing Symbiosis · Plant Pathogenic Bacteria Studies
Introduction
Microbiota play a key role in plant and animal defense against pathogens, both by modulating the immune system of their host and by excluding pathogens [1, 2]. Pathogen exclusion can occur through antagonism and the production of antimicrobials, or through niche competition and exclusion [3]. Niche exclusion can occur through direct physical competition, for instance by occupying space, or through more efficient use of nutrients. Although many antimicrobials made by microbiota that target pathogens have been identified, how microbiota exclude pathogens is poorly understood.
Biofilm formation has been implicated in both microbial virulence, as well as microbiota-mediated exclusion of pathogens [4]. Biofilms are comprised of mechanistically diverse extracellular matrices consisting of proteins and exopolysaccharides that are formed by microbes for biotic and abiotic surface attachment [5, 6]. For plant-associated microbiota, biofilm formation is required for rhizosphere colonization. For instance, a reverse genetics screen of Bacillus biofilm determinants identified that many in vitro biofilm components are also required to colonize plants [7]. Similarly, a forward genetic screen in a beneficial Pseudomonas strain found the large adhesion protein A (LapA) is required to colonize corn roots [8]. LapA is also required for biofilm formation in vitro [9] indicating that there might be overlapping mechanisms between biofilm formation and host association. However, several Pseudomonas ogarae F113 mutants affecting cyclic di-GMP biosynthesis and in vitro biofilm formation resulted in similar or enhanced in planta fitness and pathogen protection [10, 11]. Whereas biofilms have been extensively studied in vitro, there are limited data testing the degree to which biofilm formation in vitro and plant colonization share mechanistic overlap.
Reductionist model plant-microbiota-pathogen systems have facilitated the identification of mechanisms by which microbiota can protect hosts from pathogens [12]. A previously described model system consisting of the beneficial Pseudomonas brassicacearum strain WCS365, a closely related pathogen Pseudomonas sp. N2C3 [13], and the model plant Arabidopsis thaliana (Arabidopsis) was used to identify mechanisms required by mutualists for protection against pathogens [14]. For example, plant colonization through a two-component system ColR/S and LPS core polysaccharide modification was shown to be required for WCS365-mediated protection against N2C3 [14]. Although P. brassicacearum WCS365 is an efficient plant colonizer, it forms discontinuous microcolonies on the root surface [15, 16], which are reminiscent of microcolonies formed during early biofilm formation [5]. We hypothesized that mechanisms that contribute to biofilm and microcolony formation by the beneficial strain P. brassicacearum WCS365 would be required for protection against pathogenic N2C3.
To identify bacterial biofilm components necessary for pathogen protection, we screened two previously described, but only partially characterized, collections of P. brassicacearum WCS365 biofilm transposon insertion mutants for pathogen protection [9, 17]. These include mutants with decreased (sad, surface attachment defective mutants) and increased (iam, increased attachment mutants) biofilm formation on abiotic (plastic and glass) surfaces [9]. From this screen, mutations in genes encoding the large adhesion protein A (LapA) system were described as promoting biofilm formation by P. brassicacearum [9]. Because LapA was also previously implicated in plant association [8], and because the majority of mutants in the sad and iam collections have had limited characterization, we hypothesized that these libraries are a source of novel rhizosphere colonization determinants required to exclude pathogens. Furthermore, the iam mutants provide the opportunity to determine if increased biofilm formation can enhance pathogen protection, or rather, will result in colonization defects due to mis-regulation of biofilm formation [18].
By mapping the genetic location of transposon insertions in the P. brassicacearum WCS365 iam and sad biofilm libraries, we identified mutations in both previously described and novel biofilm formation components*.* Whereas the iam and sad mutants had altered biofilm formation in vitro, we found only a subset of these mutants lost the ability to protect against pathogens in planta. These results suggest that only a subset of biofilm components required in vitro are required for plant-protective functions in planta, and that in vitro biofilm formation and in planta pathogen protection use only partially overlapping mechanisms.
Results
Rescreening a collection of P. brassicacearum biofilm mutants identified genes required for pathogen protection
We set out to determine if P. brassicacearum WCS365 mutants with increased or decreased biofilm formation [9] could still protect plants from the closely related Pseudomonas sp. N2C3 [14]. In vivo biofilm formation is challenging to quantify directly, and we reasoned that protection against pathogens would provide a readout for the symbiotic ability of the P. brassicacearum WCS365 biofilm mutants. As not all of the P. brassicacearum WCS365 iam and sad library transposon insertion sites were mapped previously, we first mapped the transposon insertions by arbitrary primed PCR [9]. The PCR products were sequenced and aligned to the P. brassicacearum WCS365 reference genome to determine the location of the transposon insertions [19]. Among a total of 62 mutants, we identified insertions in 35 unique genes, the majority of which were previously implicated in biofilm formation (Table S1, Fig. 1A). The majority of insertions were within four genetic loci including nine insertions in a flagellar synthesis and function operon (fliKLMNOPQR; [20, 21]), 15 insertions in large adhesion protein biosynthesis locus (lapDAEB; [19, 22]), seven insertions in an LPS biosynthesis operon (RS13585/warR; [23]), and nine insertions within a predicted O-antigen biosynthesis operon (wbpADE/wzt/wphH/wapH/wbpJ [24, 25]) (Fig. 1A). Additional genes with multiple insertions from the screen include pvdQ [26], hisD, proA, and rlmN (Table S1). There were seven insertions in genes and operons that only had a single insertion including in intergenic regions or hypothetical proteins (Table S1). As we had less confidence that these insertions were responsible for the observed phenotype, these singletons were not considered further except for clpP [9, 27], which has been previously characterized in WCS365 biofilm formation. The diversity of mutants and biofilm processes captured in the iam and sad libraries will allow us to test whether in vitro biofilm formation predicts plant colonization.
A high throughput screen identified P. brassicacearum WCS365 biofilm mutants that cannot protect plants from a pathogen. (A) Schematic of four major operons identified and characterized in the screen including flagellar biosynthesis and function, large adhesion protein, and two operons involved in O-antigen modification. Triangles indicate approximate locations of transposon insertions. Insertions that are only affected with a single gene within an operon including clpP, hisD, proA, pvdQ, and rlmN, are not illustrated. (B) Schematic of high-throughput pathogen protection screen where plants are grown in 96-well plates and treated with either a pathogen, a mutualist, or in combination. Increased attachment mutants (iam) and surface attachment defective (sad) mutants of WCS365 were screened using a 96-well plant health quantification assay. Plants were inoculated with a 1:1 ratio of WCS365 iam or sad libraries and the N2C3 pathogen expressing GFP from a plasmid. (C) Plant health and bacterial fluorescence was quantified. Plant health was quantified by scanning plants with a high-resolution scanner and quantifying plant size and color. Growth of the bacterial pathogen was quantified by reading GFP fluorescence. Level of GFP signal is depicted as a heatmap where brighter indicates level of fluorescence. (D) Plant health was quantified as a function of how large and how green leaves are at the end of the assay period. Using this metric, wildtype WCS365 results in largely healthy plants even with the N2C3 pathogen is present. A previously described WCS365 ∆colR mutant that cannot protect plants was used as a control; the dotted orange line indicates the threshold to detect ∆colR mutants with 95% confidence, which was used as a cutoff (dashed line) in this preliminary screen. Each dot represents a single plant from four independent replicates, and genes with multiple insertions are pooled. Bars indicate the mean, and then error bars represent the 95% confidence intervals of the control.
Correlation between in vitro biofilm formation and plant protection
We hypothesized that biofilm formation would be positively correlated with plant colonization and protection against pathogens. However, we also hypothesized that increased biofilm formation, could potentially be detrimental for plant colonization and pathogen protection. To screen the P. brassicacearum WCS365 iam and sad mutants for protection against the pathogenic Pseudomonas sp. N2C3, we made use of a high-throughput plant protection assay [28]. In this assay, seedlings are germinated in 3D printed MYCroplanters and then transferred to wells of 96-well plates containing bacteria (Fig. 1B). Under these conditions, WCS365 protects against N2C3 and results in healthier plants and low levels of N2C3 abundance; both bacterial abundance and plant health can be readily quantified (Fig. 1B). We inoculated plants with wildtype WCS365 or individual iam or sad mutants in combination with Pseudomonas sp. N2C3 containing a plasmid expressing GFP in four biological replicates (Fig. 1B). The previously described colonization-deficient WCS365 ∆colR mutant was used as a control, as it cannot protect against N2C3 [14, 29]. We then quantified plant health using a Python script to extract seedling size and color from scanned plate images, where larger, greener plants are scored as healthier [28]. We also quantified N2C3-GFP fluorescence to determine if the WCS365 mutants were no longer able to protect the plant and if N2C3 was able to grow in the rhizosphere (Fig. 1B and 1C).
We found that a subset of iam and sad mutants could no longer protect plants against disease caused by Pseudomonas sp. N2C3 as measured by a decreased plant health score (Fig. 1C, D). Interestingly, nonprotective iam and sad mutants included both those with increased and decreased biofilm formation. Nonprotective sad mutants included insertions in genes involved in large adhesion protein biosynthesis and localization (lapD and lapA), consistent with previous descriptions of a role for the Lap adhesion system in plant colonization [7] and flagellar synthesis and function (fliKNPR and flgK). Nonprotective iam mutants included those with insertions in lipopolysaccharide (LPS) modification (wbpAD). Conversely, a number of other mutants including those involved in LPS modification retained protective ability. Collectively, these findings indicate that some, but not all, genes required for in vitro biofilm formation are required for pathogen protection in planta.
Decreased in planta fitness partially explains loss of protection against pathogens
In our high throughput plant protection assay, each well contains plant root exudates that can support the growth of bacteria. Under these conditions, wildtype P. brassicacearum WCS365 outcompetes pathogenic N2C3 in the rhizosphere resulting in WCS365 dominating the final community in each well [14, 28]. Here, co-inoculation with WCS365 resulted in low growth of N2C3, and N2C3-GFP fluorescence was not detectable above the background detection limit (Fig. 2A). We hypothesized that nonprotective iam and sad mutants would be associated with increased growth of the N2C3 pathogen, but potentially not a change in growth of the WCS365 mutant.
Decrease in mutualist fitness and increase in pathogen growth in planta contribute to loss of protection. (A) Quantification of N2C3-GFP signal in competition with P. brassicacearum. The horizontal dashed line indicates the background detection limit. (B) Increased growth of the N2C3-GFP strain partially explains a decrease in plant health (R2 = 0.31, P = 0.0035). (C) Growth of P. brassicacearum WCS365 iam and sad mutants in competition with N2C3. The dashed line shows the 95% confidence interval of wildtype WCS365 growth. Bars are colored by whether strains were protective (orange) or non-protective (blue) from Fig. 1. (D) Decreased growth in planta explains some but not all of the loss of protection (R2 = 0.19, P = 0.028).
We first measured pathogen growth by quantifying N2C3-GFP signal in each well. We found that several WCS365 iam or sad mutants that resulted in decreased health scores when competed against N2C3 also had increased N2C3-GFP fluorescence (Fig. 1C and 2A), suggesting that the ability to limit pathogen growth may play an important role in mutualist-mediated plant health. To test whether all WCS365 mutants that lost protection also had corresponding increases in N2C3 abundance, we performed a linear regression between N2C3-GFP fluorescence and plant health. We found a significant negative correlation between N2C3 abundance as measured by GFP fluorescence signal and plant health (R^2^ = 0.31, P = 0.0035; Fig. 2B). Interestingly, insertions in the hisD auxotrophic mutants resulted in a similar level of N2C3-GFP fluorescence as N2C3-GFP alone suggesting these mutants likely had low growth in the rhizosphere (Fig. 2A). In contrast, the majority of flagellar mutants failed to protect plants, but without a corresponding increase in GFP signal (Fig. 2B). This suggests that the flagellar mutants may still inhibit N2C3 growth, but that loss of flagellar motility may result in a loss of pathogen protection due to decreased plant colonization. Altogether, these results are consistent with loss of protection through diverse mechanisms.
Increase in N2C3 growth and decrease in plant health in the presence of iam and sad mutants could be a result of rhizosphere fitness defects or due to loss of niche occupation because of altered biofilm formation*. In planta,* we found that a subset of the nonprotective strains had reduced rhizosphere growth consistent with rhizosphere fitness defects (Fig. 2C). We found a positive correlation between plant health and growth of WCS365, although less robust than the correlation between N2C3 abundance and plant health (R^2^ = 0.19, P = 0.028; Fig. 2D). This finding suggests that some iam or sad mutants, such as the hisD mutants, were unable to grow in planta, which explains their inability to exclude the N2C3 pathogen. Others, such as a strain carrying a mutation in the fliK gene, still grew to fairly high levels but cannot protect plants against N2C3. Interestingly, similar to the previously described colonization deficient WCS365 ∆colR mutant, the wbpAD mutants, which are involved in O-antigen biosynthesis, grow to an intermediate level in the rhizosphere. ColR regulates genes involved in LPS modification, although not specifically wbpAD [29]. The co-clustering of these mutants for rhizosphere fitness and plant health suggests that wbpAD and colR genes may contribute to rhizosphere fitness through related processes.
To rule out the possibility that decreased plant health was caused by the biofilm formation mutants directly, we measured the plant health scores following incubation with individual WCS365 iam or sad mutants in the absence of pathogen. We also measured the growth of each individual strain by measuring the final OD600. None of the WCS365 biofilm formation mutants resulted in a significant decrease in plant health or had a significant difference in total growth relative to wildtype bacteria (Fig. S1). To determine if growth defects of iam or sad mutants were rhizosphere specific, or the mutants have generalized growth defects, we performed in vitro growth curves with these mutants in LB medium. We found that the majority of mutants grew to similar levels as wildtype bacteria (Fig. S2) and that those with significant, although modest changes in growth did not correlate with those that could no longer protect (R^2^ = 0.0024, P = 0.82*;* Fig. S2). Collectively, these findings indicate that both pathogen exclusion and competitive fitness in the rhizosphere contribute to WCS365-mediated protection.
Some biofilm mechanisms are conditionally required for protection against pathogens
We previously found that there is not a perfect correlation between genes required for pathogen protection in hydroponics compared to a solid surface assay [14, 29]. Although the hydroponic assay has the benefit of being high throughput, it may not recapitulate all aspects of growing in planta, including the necessity to move across liquid-air interface and solid surfaces. As a result, we repeated protection assays for 19 mutants representing the majority of genes and operons identified in the screen using a solid surface plant assay [14]. In these assays, plant fresh weight is a read-out for protection. Our results show N2C3 significantly stunts plant growth, whereas plants treated with WCS365 or a 1:1 ratio of WCS365 and N2C3, have similar fresh weights as buffer-treated plants (Fig. 3A). To test for pathogen protection, we co-inoculated P. brassicacearum WCS365 or WCS365 mutants with Pseudomonas sp. N2C3 in a ratio of 1:1 along the plant roots.
*The extent of in vitro biofilm formation does not predict protection against a pathogen in planta. (A) From the 65 P. brassicacearum WCS365 biofilm formation mutants in the iam and sad mutant collections (Table S1), 21 mutants were re-tested for their ability to protect the rhizosphere against disease caused by the Pseudomonas sp. N2C3 pathogen. In a competition assay using P. brassicacearum WCS365 mutants with Pseudomonas sp. N2C3 in a ratio of 1:1, 12 mutants lost protection in the rhizosphere. Each dot represents an average of three technical replicates as one independent biological replicate. The assay was repeated at least three times. Statistical significance was determined by two-way ANOVA comparing different strains with N2C3 treatment, P < 0.001 = *. Error bars show standard deviation. (B) in vitro biofilm was quantified using a crystal violet assay. The sad-51/lapA::Tn5 and morAAAL mutations are previously described hypo- and hyper- biofilm controls, respectively. Each dot represents a technical replicate from one independent biological replicate and each biological replicate includes eight technical replicates. The assay was repeated at least three times. Statistical significance was determined by one-way ANOVA comparing different strains with WCS365 treatment followed by a Tukey’s HSD test P < 0.05. All error bars show standard deviation. (C) Correlation between in vitro biofilm formation versus plant weight. The plant weight was used as a readout for protection and whether the tested mutant can outcompete Pseudomonas sp. N2C3 in the rhizosphere or not (R2 = 0.0002, P = 0.98). Each dot represents the average of at least three independent biological replicates quantifying biofilm formation and plant weight. (D-E) Independent linear regressions were performed for just the sad (D) or iam (E) mutants. Linear regression analysis was performed in R by ggplot2. A weak positive correlation between the in vitro hypobiofilm formation and protection (R2 = 0.50, P = 0.077), but not hyerbiofilm (R2 = 0.046, P = 0.46), was found.
We found that 13 of the 19 P. brassicacearum WCS365 mutants lost the ability to protect against Pseudomonas sp. N2C3 as indicated by a significant decrease in fresh weight relative to the protective wildtype P. brassicacearum WCS365. These include mutants with insertions in genes coding for the Lap system (lapDAEB) [30], motility (fliK, fliN, fliP, fliR, flgK), lipopolysaccharide (LPS) modifications (wbpAD, wzt) [31, 32], and a protease encoded by the clpP gene [9] (Fig. 3A). The remaining P. brassicacearum WCS365 mutants maintained pathogen protection as indicated by no significant difference in plant weight when co-inoculated Pseudomonas sp. N2C3 (Fig. 3A). Whereas all iam and sad mutants defective in plant protection against N2C3 in hydroponics were also required in a solid surface plant assay, several additional genes, when mutated, were shown to be conditionally required on the solid surface plant assay including clpP, wzt, and lapEB. These results indicate a potential differential requirement for some components of biofilm formation involved in pathogen protection on plants growing on a solid surface. As plant growth on solid agar may more closely mimic soil, where microbes must navigate air-liquid interfaces and movement along soil particles, the solid surface plant assay may more closely mimic soil conditions and reveal the importance of additional biofilm genes in pathogen protection.
Because the hydroponic assay also quantified total bacteria in both the rhizosphere (in the well) and on the rhizoplane (on the root), we tested whether all non-protective mutants were completely outcompeted by N2C3 at the rhizoplane. We co-treated plants with a subset of WCS365 biofilm formation mutants and N2C3 expressing lacZ, and ground roots and plated bacteria to quantify the relative abundance of each strain [14]. We found that in competition with both wildtype WCS365 and the protective mutant *wapR::*Tn5, less than 1% of the final root-associated community was N2C3 (Fig. S3). We found that the nonprotective mutants *lapA::*Tn5 and *clpP::*Tn5 as well as the ∆colR colonization deficient control were completely outcompeted by N2C3. The remaining nonprotective mutants lapD::Tn5, wbpD::Tn5 and wzt::Tn5 were able to coexist with N2C3 (Fig. S3). Collectively, these results indicate that plant protection involves both the ability to compete in the rhizosphere and colonize, as well as additional factors like niche occupation that are not captured by bacterial abundance alone.
There is no correlation between the extent of biofilm formed in vitro and pathogen protection in planta
We found that a subset of P. brassicacearum WCS365 mutants from both the iam and sad libraries lost protection against Pseudomonas sp. N2C3, and therefore there did not appear to be a correlation between the extent of biofilm produced and protection. Using the same subset of 19 mutants retested on the solid plate assay, we robustly quantified in vitro biofilm formation and in planta competition.
Using a modified crystal violet assay [33] we validated the previously described increased or decreased biofilm phenotypes for the 19 P. brassicacearum WCS365 mutants in vitro. The assay conditions were nearly identical to the original assay including the same media composition, and the biofilm was allowed to proceed overnight for 18 h [9] to reflect the longer interaction time with a plant [9]. We found 13 mutants, including components of LPS modification and flagellar synthesis and function, formed significantly higher amounts of biofilm than wildtype P. brassicacearum WCS365 (Fig. 3B). We found six mutants, primarily in the Lap system, formed lower biofilm amounts (Fig. 3B). It is noteworthy that in the repeat of the biofilm assays the flagellar mutants formed similar or increased biofilm relative to wildtype WCS365, although they were originally described as reduced for biofilm formation [9]. Quantitative differences in biofilm formation have previously been described for Pseudomonas flagellar mutants as a function of time and media [34], which is consistent with the observation that flagella are required for early stages of attachment, but not mature biofilm formation [5]. Although the replicate assay was performed in the same medium as the initial assay but over 18 h instead of 10 h as in the initial screen, we conducted a 10-h in vitro biofilm assay using several representative mutants to test if the incubation time could explain the distinct results. We found that most of these mutants exhibited similar levels of biofilm formation at 10 h as they did at 18 h, with the exception of fliP, which showed reduced biofilm formation at the shorter time point as was in the initial screen (Fig. S4). These results are consistent with distinct roles of flagella at distinct times during biofilm formation. The remaining mutant phenotypes are consistent with descriptions of biofilm phenotypes from the iam and sad mutants and previous descriptions of biofilm components [9].
To determine if the extent of the in vitro biofilm formed could predict in planta pathogen protection, we performed a linear regression between the plant weight, as a readout for protection, and in vitro biofilm formation at 18 h. No significant correlation between the two phenotypes was found, suggesting that the extent of the biofilm formed by a strain in vitro does not directly predict its protective ability in planta (R^2^ = 0.0002, P = 0.98; Fig. 3C). As non-protective WCS365 mutants included both decreased (sad) and increased (iam) attachment mutants, we formed separate regression analyses to test if there was a correlation between mutants that formed decreased or increased biofilm. Interestingly, we found a trend between decreased biofilm formation and decreased protection (R^2^ = 0.50, P = 0.077; Fig. 3D) suggesting that loss of biofilm formation results in inability to protect plants from N2C3. In contrast, there was no correlation between increased WCS365 biofilm formation and protection against N2C3 (R^2^ = 0.046, P = 0.46; Fig. 3E). These results are consistent with diverse mechanisms of biofilm formation in vitro, that may not precisely correlate with functions in planta.
Biofilm components are required for protection against diverse rhizosphere pathogens
Pseudomonas sp. N2C3 is an opportunistic pathogen of plants and produces a lipopeptide toxin [13]. Previously described P. brassicacearum WCS365 genes required for protection against N2C3 are also required for protection against a virulent rice pathogen Pseudomonas fuscovaginae SE-1 [14], which uses a similar toxin-based virulence mechanism to cause disease. Pseudomonas aeruginosa is an opportunistic pathogen of both plants and animals but uses distinct virulence mechanisms from N2C3 and SE-1 [35]. We tested whether the same genes required to protect against N2C3 were also required to protect against P. fuscovaginae SE-1 and P. aeruginosa PAO1 by choosing three mutants affecting diverse processes, including *lapA::*Tn5 (large adhesion protein, hypobiofilm), wzt::Tn5 (LPS modification, hyperbiofilm), and clpP::Tn5 (protein turnover, hypobiofilm). We found that wildtype WCS365 robustly protects from both plant biomass decreases (Fig. 4A) and root stunting (Fig. 4B-C) by all three pathogens. We found consistent loss of protection by the *lapA::*Tn5, *wzt::*Tn5 and *clpP::*Tn5 mutants against both N2C3 and SE-1. Interestingly, none of the genes were required for protection against PAO1 as measured by no significant reduction in plant biomass relative to the mutant alone. The only exception was that the lapA mutant, which did not fully protect against PAO1-mediated root stunting. Interestingly, we found that clpP mutant itself resulted in plant root stunting, however the fresh weight of plants inoculated with the clpP mutant did not significantly differ from buffer treated plants (Fig. 4). Collectively these findings indicate that precise regulation of biofilm formation is required for mutualists to colonize plant roots and protect plants against various pathogens of agronomic importance.
Biofilm mutants have protection defects against diverse pathogens. To determine if mechanisms of P. brassicacearum WCS365 excluding pathogens are also important for Pseudomonas pathogens with distinct virulence mechanisms, we tested mutants in three distinct processes for their ability to protect against Pseudomonas fuscovaginae SE-1 and P. aeruginosa PAO1. Plant biomass (A) and root length (B) were quantified with representative images shown in panel C. Letters denote differences by ANOVA and Tukey’s HSD tests. Mean and 95% confidence intervals are shown.
Discussion
Bacteria form biofilms to attach to abiotic surfaces and colonize plants. However, whether bacteria employ the same mechanisms for biofilm formation in vitro and in planta remains largely unknown. To gain a broader understanding of mechanisms that are shared or distinct between biofilm formation in vitro and in host colonization, we screened a collection of P. brassicacearum WCS365 transposon insertion mutants with altered biofilm formation on abiotic surfaces using a previously described plant-mutualist-pathogen model [13, 14]. This collection of mutants spans a broad range of gene functions that impact biofilm formation on abiotic surfaces, including known genes involved in large adhesion proteins and flagellar biosynthesis and function, as well as hyperbiofilm formers involved in LPS biosynthesis that have not previously been implicated in biofilm formation or host colonization. By employing a high-throughput assay that measures fluorescence and absorbance, we were able to monitor the plant health as well as the individual growth of both a pathogen and a mutualist coexisting in the rhizosphere [24]. We found that not all the mutants in the iam and sad libraries lost protection of plants against a pathogen, suggesting that not all biofilm formation mechanisms in vitro are required for protection against pathogens in planta. This finding also indicates that bacteria may employ partially distinct mechanisms for biofilm formation in vitro versus in planta. We found that the plant health is more strongly correlated with the abundance of the pathogen than the mutualist as some biofilm mutants were able to maintain their growth but failed to provide protection. Our results suggest that protection requires close association of the protective microbe with the plant and not just growth in the rhizosphere.
We tested protection against the N2C3 pathogen in two assays, one in hydroponics and one on the solid agar plates. Previous work identified some discrepancy between bacterial colonization of plants using these two assays, specifically that genes required for LPS modification were more important for colonization on solid surfaces [14, 29]. Similarly, in this study, we found that the solid rhizosphere condition altered the protection phenotype of certain biofilm mutants including those in the wzt and clpP genes. This finding might suggest that the function of these genes is more important for survival or competition with other microbes on solid surfaces or in an air-liquid interface assay.
We found that the mutants that failed to protect plants from the Pseudomonas sp. N2C3 pathogen make similar, higher, or lower amounts of biofilm than wildtype P. brassicacearum WCS365 on abiotic surfaces, indicating that the extent of in vitro biofilm formation does not itself correlate with protection in planta. However, we observed a positive correlation between hypobiofilm formation in vitro and reduced plant weight, indicating that decreased biofilm production indeed diminishes the protective effect. Thus, the inability to properly form a biofilm in vitro impacts rhizosphere fitness of these mutants, which likely influences their capability to confer plant protection.
We found that loss of the large adhesion protein A of P. brassicacearum WCS365 (encoded by lapA gene) resulted in strains unable to protect plants against Pseudomonas sp. N2C3 and also formed significantly lower amounts of biofilm compared wildtype WCS365 in vitro (Fig. 1D and Figure 3AB). The Lap system was previously shown to be required for biofilm formation in P. brassicacearum WCS365 [17, 22]. The lapA gene encodes a large adhesion protein which is required for attachment to plant roots and abiotic surfaces [8, 9]. LapEB are components of the type I secretion system which is required for LapA secretion [19, 30]. The lapD gene, which encodes an inner membrane receptor of the intracellular signaling molecule c-di-GMP, determines the bacterial lifestyle transition between biofilm formation and dispersion by controlling the retention or the release of the LapA protein [19, 30]. Loss of protection by the P. brassicacearum WCS365 lapDAEB mutants suggests that the entire Lap system and the resulting secretion and cell-surface localization of LapA are likely necessary for protection against the Pseudomonas sp. N2C3 pathogen, which may be due to the reduced rhizosphere colonization (Fig. 2C).
Both biofilm formation and motility are important for successful rhizosphere colonization; however, they are often inversely regulated where downregulation of motility coincides with decreased initial attachment, but increase in biofilm-forming ability [36–38]. The fliK (encodes a polar flagellar hook-length control protein) [21, 39], the fliN (encodes a flagellar motor switch protein) [8, 40], and the fliP (flagellar axial protein export apparatus) mutants [41] showed significantly increased biofilm formation at 18 h (Fig. 3B). Both fliK and fliP (flagellar axial protein export apparatus) mutants showed a loss of protection in the rhizosphere, but not rhizosphere colonization, suggesting that P. brassicacearum WCS365 requires functional motility for competitive rhizosphere colonization [14]. These findings suggest that motility is necessary for competitive colonization, likely due to a defect in early attachment.
We found that transposon insertions that disrupt a subset of WCS365 LPS modification genes resulted in a loss of ability to protect against N2C3. As the major components of the Gram-negative bacterial outer membrane, LPSs are more than just a physical permeability barrier for toxic compounds. LPS have been shown to be involved in versatile biological processes, such as host immunity recognition and evasion [42, 43], colonization [44, 45], and establishing symbiosis [46, 47]. LPS consists of three regions: the membrane anchoring region lipid A, the middle region core oligosaccharide, and the outside region referred to O-polysaccharide or O-antigen [48]. Genes involved in O-antigen transportation (wzt) [31] and biosynthesis (wbpADE, wbpJ) [49, 50] were identified in our mutant library. Wzt is part of the Wzm/Wzt ATP-binding cassette (ABC) transporter that specifically transports O serotypes O8 and O9a in Escherichia coli [31] and A-band O-antigen LPS in P. aeruginosa PAO1 [51]. The Wbp pathway including the wbpADE and wbpJ genes are in the B-band LPS serotype-specific O-antigen biosynthesis cluster in P. aeruginosa PAO1 [49–51]. Whereas some components from the core oligosaccharide modification (wapHR), O-antigen transportation (wzt) [31] and biosynthesis (wbpADE, wbpJ) behaved differently under liquid or solid rhizosphere conditions, we found that the O-antigen of LPS plays a more important role in competition against the pathogen in planta.
Collectively, our work sheds light on the role of bacterial biofilm formation in colonization and plant pathogen protection in the rhizosphere. As in vivo biofilms are difficult to quantify, in vitro biofilm formation is often used as a proxy for in vivo biofilm production. Importantly, our screen of the iam and sad mutant libraries demonstrated that only a subset of genes involved in biofilm formation lost protection in planta indicating that in vitro biofilm formation does not always predict pathogen protection.
By testing a collection of biofilm mutants for their ability to colonize plants, we revealed common mechanisms required for biofilm formation in vitro and protection against pathogens, as well as biofilm components that are dispensable in planta. Future characterization of genes that lost protection and have not previously been implicated in host colonization would also provide new insights into mechanisms of mutualist-mediated protection of plants against pathogens. In addition, the characterization of the nonprotective mutants identified in this study will enhance our understanding of this process and provide better guidance to microbiome engineering.
Materials and methods
Plant materials and growth conditions
A. thaliana Col-0 seeds were surface sterilized by a mixture of 70% bleach followed by 10% ethanol, or 70% EtOH and 1.5% H_2_O_2_ solution on filter paper and allowed to dry for 30 min in a laminar flow hood. Sterilized seeds were then transferred into a centrifuge tube with 0.1% agar and stored in 4°C in the dark for 48 h before sowing. Seeds were germinated on square plates with ½ X Murashige and Skoog (MS) medium containing 1 g/L 2-(N-morpholino) ethanesulfonic acid (MES) buffer, 2% sucrose, and 1% agar for 5 days. The pH of the MS medium was adjusted to 5.7 with 1 M KOH. On Day 6, plants of similar size were transferred to plates with ½ X MS medium containing 1 g/L MES buffer and 1% agar without sucrose. On Day 7, plants were inoculated with 5 μl of bacterial culture with OD_600_ of 0.001. All the plant materials were grown at 22°C at 100 μM m^−2^ s^−1^ light under a 12-h light/dark cycle in a temperature-controlled growth room unless otherwise indicated.
Bacterial strains and growth conditions
The library of P. brassicacearum WCS365 increased attachment mutants (iam) and surface attachment defective (sad) transposon insertion mutants were previously described [9]. The library contains 65 mutants in total (Table S1). Overnight cultures were made in LB broth supplied with either 10 μg/ml gentamycin or 40 μg/ml tetracycline and grown at 28°C with shaking at 180 rpm. The bacterial overnight cultures were diluted and resuspended in 10 mM MgSO_4_ to the indicated OD_600_ prior to plant inoculation. P. fuscovaginae SE-1 [52] was grown in LB at 28°C and P. aeruginosa PAO1 [53] was grown in LB at 37°C.
In vitro bacterial growth
The wildtype P. brassicacearum WCS365 and the iam and sad libraries were grown overnight in LB broth in a 96-well plate. Optical density at 600 nm (OD_600_) was measured in a Biotek Epoch 2 plate reader. Cultures were diluted to an OD_600_ of 0.02 in fresh LB broth in 200 μl total in a 96 well plate. The plate was incubated and read in a Biotek Epoch 2 plate reader set to 28°C with continuous orbital shake for 24 h. OD_600_ was read every 15 mins.
Transposon insertion mapping by arbitrary PCR
The DNA sequences flanking the insertions in the transposon insertion mutants were determined by arbitrary PCR and Sanger sequencing as previously described [9, 54]. The DNA flanking regions were amplified by two rounds of PCR by using two sets of primers. In the first round of PCR, a primer Tn5Ext, which is unique to the transposon but more distal from the transposon end and an arbitrary primer (ARB1), which can hybridize the chromosomal sequences flanking the transposon were used to enrich the genomic DNA near the transposon. The second round of PCR used the PCR products from the first round of PCR as a template, a primer Tn5Int, which is also unique to the transposon but more proximal to the transposon end (around 60 bp from the transposon to the chromosome junction) and an ARB2 primer, which has identical 5’end as ARB1. The PCR products were purified either by PCR clean-up kit (QIAquick PCR purification kit) or by gel extract purification kit (QIAquick gel extract kit) and were then sent for Sanger sequencing. The insertion location was identified by BLAST using the P. brassicacearum WCS365 reference genome (NCBI Accession CP089973.1).
MYCroplanters
GFP-expressing Pseudomonas sp. N2C3 was generated using pSMC21 [55] via electropoartion. Briefly, an overnight culture of N2C3 in LB broth was pelleted and washed twice in 300 mM sucrose to generate electrocompetent cells. Transformed cells were plated on LB supplemented with 25 μg/ml kanamycin for selection.
Arabidopsis seeds were sterilized and germinated in the MYCroplanter system [28]. Five days after gemination, wildtype P. brassicacearum WCS365 and the iam and sad libraries was grown overnight in LB broth in a 96-well plate at 30°C with shaking at 200 rpm. Pseudomonas sp. N2C3-GFP was grown overnight in LB broth supplemented with 25 μg/ml kanamycin. On Day 6, the WCS365 strains were spun down, spent LB broth was removed, and cells were resuspended in the same volume of ½ MS supplemented with MES and no sucrose. The N2C3-GFP culture was spun down, washed once to remove kanamycin, and resuspended in ½ MS supplemented with MES and no sucrose. For all strains, OD_600_ was measured using Biotek Epoch 2 plate reader. For transferring seedlings, a fresh 96-well plate was filled with 275 μl at a 1:1 ratio of wildtype WCS365 or iam or sad mutants and N2C3-GFP in 50 000 cells total using the cell estimate that 1 ml of Pseudomonas culture at OD_600_ of 1 is 5 × 10^8^ cells. Experiments in mono-association with the WCS365 iam or sad libraries were performed as above, but with no N2C3 added. A single MYCroplanter was transferred to each well of the 96-well plate, the lid brace was added, and the 96-well plate was sealed with 3 M Micropore tape. The MYCroplanter system was incubated in a reach in growth chamber for 7 days.
At harvest, MYCroplanters were transferred to the scanning tray and scanned on an Epson Perfection V850 Pro scanner as stated in [28]. GFP fluorescence and OD_600_ of the 96-well plate without MYCroplanters were read in a BioTek Synergy H1 plate reader. Health score calculations were performed as described [28]. Briefly, each MYCroplanter containing a bacterial-treated plant was transferred to a scanning tray and scanned with a flatbed color scanner. Plant health was scored using a continuous metric that combines plant color and size derived from the scanned plant images. By generating a principal components plot of plant pixel size and color, we found that the data clustered plants into three groups that included green large plants that are “healthy”, red plants that are “stressed/sick”, and small yellow plants that are “dead”. We transformed pixel information (hue, saturation, value) into a single continuous variable where “Health Score” = (Huesaturationlog10(Plant pixels +1)*100. This health score was previously validated by human-derived assessments of plant images.
N2C3-GFP quantification was taken directly from the GFP fluorescence measurements. To calculate the fraction of WCS365 in the well, N2C3-GFP signal was converted to OD_600_ using a standard curve. For each well, the N2C3-GFP approximated OD_600_ was subtracted from the total OD_600_ reading to estimate the amount of WCS365 in the well.
Pathogen protection assay on solid agar
The bacterial overnight cultures were diluted and resuspended in 10 mM MgSO_4_ to a final OD_600_ of 0.001 inoculum for single-strain inoculations. For bacterial competitions, bacterial mixtures were prepared in a ratio of 1:1 with final OD_600_ of 0.002 (0.001 for each strain). Each 7-day old plant was inoculated with 5 μl of bacterial culture along the root. Plates were dried in a biosafety cabinet before sealing and moved to the growth chamber as described above. Seven days later, all the plates were imaged using an Epson V850 flatbed scanner and weighed by pooling three plants per treatment. To quantify WCS365 transposon mutant and N2C3 bacterial abundance in the root, pathogen protection assays were performed using N2C3-lacZ for blue-white screening. Plants were inoculated with WCS365 transposon mutans, N2C3-lacZ, or a mixture of both as described above and grown for 7 days. At harvest, roots were aseptically separated from leaf tissue, weighed by pooling three root systems, and washed once with 1 ml of 10 mM MgSO4. After washing, roots were homogenized in 1 ml of 10 mM MgSO4 with 4–5 sterile glass 2 mm beads. Root homogenate was serially diluted and plated on solid LB supplemented with 0.2 mg/ml 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside (X-gal) and 1 mM isopropyl-β-D-1-thiogalactopyranoside (IPTG) and resulting CFUs were counted and normalized to root weight.
Crystal violet assay
Bacterial biofilm formation on a 96-well U-shape plastic surface was measured by a crystal violet assay [33]. Bacterial overnight cultures (as described above) were spun down at 10 000 × g for 3 min and the pellet was washed and resuspended in 1 × M63 medium. 100 μl of bacterial culture with OD_600_ of 0.1 was added in a 96-well plate with eight technical replicates per treatment. Plates were subsequently incubated at 28°C for 18 h or 10 h, as indicated. After incubation, the bacterial culture was removed by inverting. The plate was then washed twice by gently submerging in a small tub of water and then inverting the plate to remove the water. 125 μl of a 0.1% crystal violet solution was added to the wells (including eight wells that were used as background controls) and left for 10 min at room temperature. The plates were rinsed 3 times by submerging in water as described above. All residual water was removed by firmly shaking the plate and allowing it to dry at room temperature. For biofilm quantification, 125 μl of 30% acetic acid was added to each well including the blank wells to solubilize the crystal violet. After a 10 min incubation, 100 μl of the solubilized crystal violet or the 30% acetic acid was transferred to a new dish with flat bottom and the absorbance was read at 550 nm.
Supplementary Material
Supplemental_Materials_wraf114
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Teixeira PJP, Colaianni NR, Fitzpatrick CR. et al. Beyond pathogens: microbiota interactions with the plant immune system. Curr Opin Microbiol 2019;49:7–17. 10.1016/j.mib.2019.08.00331563068 · doi ↗ · pubmed ↗
- 2Zheng D, Liwinski T, Elinav E. Interaction between microbiota and immunity in health and disease. Cell Res 2020;30:492–506. 10.1038/s 41422-020-0332-732433595 PMC 7264227 · doi ↗ · pubmed ↗
- 3Morales Moreira ZP, Chen MY, Yanez Ortuno DL. et al. Engineering plant microbiomes by integrating eco-evolutionary principles into current strategies. Curr Opin Plant Biol 2023;71:102316. 10.1016/J.PBI.2022.10231636442442 · doi ↗ · pubmed ↗
- 4Ramey BE, Koutsoudis M, Bodman. et al. Biofilm formation in plant–microbe associations. Curr Opin Microbiol 2004;7:602–9. 10.1016/J.MIB.2004.10.01415556032 · doi ↗ · pubmed ↗
- 5Sauer K, Stoodley P, Goeres DM. et al. The biofilm life cycle: expanding the conceptual model of biofilm formation. Nat Rev Microbiol 2022;20:608–20. 10.1038/s 41579-022-00767-035922483 PMC 9841534 · doi ↗ · pubmed ↗
- 6Stoodley P, Sauer K, Davies DG. et al. Biofilms as complex differentiated communities. Ann Rev Microbiol 2002;56:187–209. 10.1146/annurev.micro.56.012302.16070512142477 · doi ↗ · pubmed ↗
- 7Beauregard PB, Chai Y, Vlamakis H. et al. Bacillus subtilis biofilm induction by plant polysaccharides. Proc Natl Acad Sci USA 2013;110:E 1621–30. 10.1073/pnas.121898411023569226 PMC 3637697 · doi ↗ · pubmed ↗
- 8Yousef-Coronado F, Travieso ML, Espinosa-Urgel M. Different, overlapping mechanisms for colonization of abiotic and plant surfaces by pseudomonas putida. FEMS Microbiol Lett 2008;288:118–24. 10.1111/j.1574-6968.2008.01339.x 18783437 · doi ↗ · pubmed ↗