Hidden reservoir of highly adaptable multi-host plasmids that propagate antibiotic genes in healthy human populations
Na Han, Xianhui Peng, Tingting Zhang, Yujun Qiang, Xiuwen Li, Wen Zhang

TL;DR
The study reveals that plasmids like pGut1, found in healthy human guts, can spread antibiotic resistance genes across diverse bacteria, posing a risk for multidrug-resistant pathogens.
Contribution
The discovery of a hidden reservoir of multi-host plasmids, including pGut1, that silently propagate antibiotic resistance genes in healthy human populations.
Findings
A global plasmid pGut1 was found in over 50% of individuals and across diverse bacterial species.
pGut1 contains a conserved backbone and a variable region with antibiotic resistance genes like cfr(C), erm(B), and aphA.
pGut1 was detected in 93 bacterial strains across 49 genera, suggesting cross-species transmission.
Abstract
Plasmids are key vectors for disseminating antibiotic resistance genes, yet their diversity and dynamics in the healthy human gut microbiome remain largely unexplored. Using fecal metagenomes from two cohorts (n = 498 samples), we constructed a comprehensive atlas of the healthy human gut plasmidome. We observed a polarization: while 97.4% of 19 151 plasmid clusters exhibited low prevalence (<5%), we identified 17 plasmid clusters that were detected in >30% of individuals. Among these, the plasmid pGut1 emerged as a paradigm of a stealth vector. Prevalent globally (>50% in independent cohorts), pGut1 possesses a minimal 4-kb conserved backbone ensuring stability and a hypervariable region acting as a “plug-and-play” module. We documented 40 distinct cargo inserts, including multiple antibiotic resistance genes such as cfr(C), erm(B), and aphA, across individuals, within individuals over…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
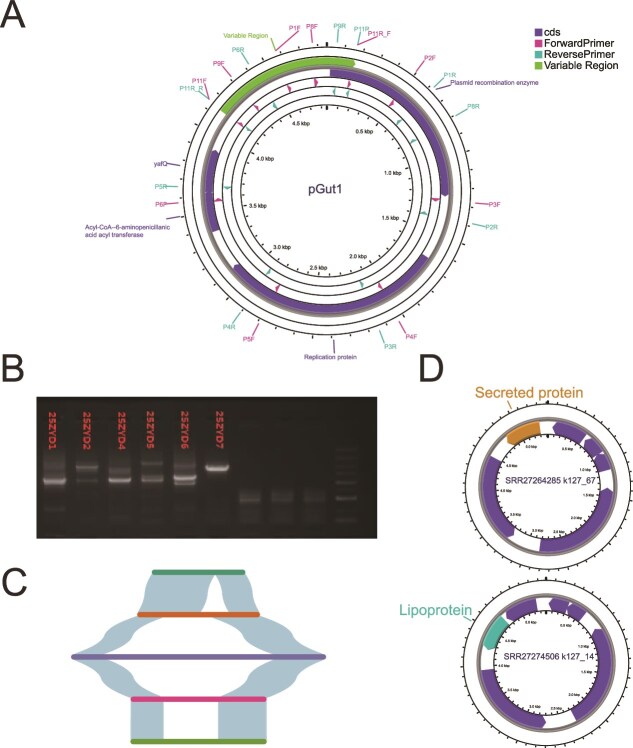 Figure 1
Figure 1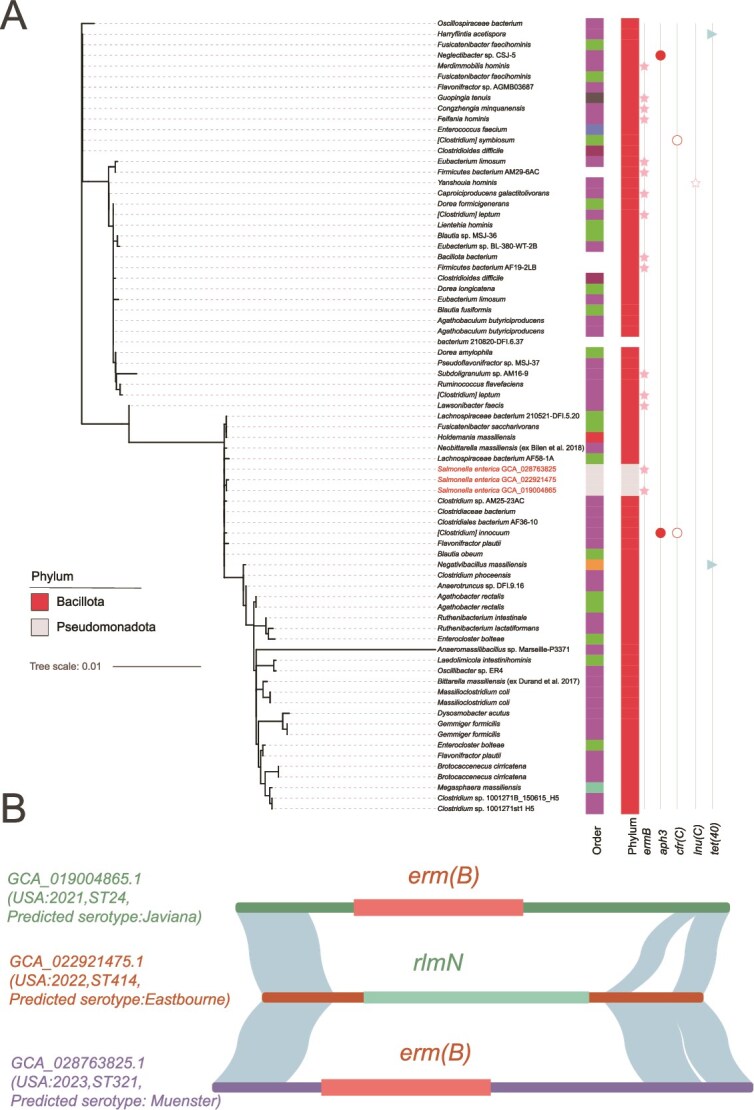 Figure 2
Figure 2- —The National Key Research and Development Program of China
- —National Science and Technology Major Project10.13039/501100018537
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacteriophages and microbial interactions · Clostridium difficile and Clostridium perfringens research · Antibiotic Use and Resistance
The healthy human gut microbiome is a vast reservoir of antibiotic resistance genes (ARGs), with plasmids serving as primary vectors for their horizontal transfer [1–4]. However, the comprehensive landscape of the gut plasmidome remains obscured by technical challenges in metagenomic assembly and database biases toward cultured clinical isolates [5–7]. To address this gap, we analyzed fecal metagenomes from two cohorts—a cross-sectional cohort (CMP_region, n = 258) and a longitudinal cohort (CMP_multitime, n = 240) (Fig. S1A)—to characterize the plasmid diversity in healthy individuals.
We identified 23,360 plasmid clusters (PLCs) from metagenome-assembled genomes, with only 6.4% showing high similarity to known plasmids in the plasmid database (PLSDB, https://ccb-microbe.cs.uni-saarland.de/plsdb2025/) [8] (Fig. S1B), highlighting a vast unexplored plasmid diversity (Fig. S1C and D). Analysis revealed extreme polarization in plasmid prevalence: 97.4% of PLCs were rare (<5% prevalence), while a small subset of 17 PLCs were detected in >30% of individuals (Fig. S1F). We term these high-prevalence, previously undetected elements “stealth plasmids”, as they have remained undetected due to technical limitations in conventional metagenomic analyses (Fig. S1F). These high-prevalence PLCs demonstrated significantly longer persistence in the longitudinal cohort compared to low-prevalence PLCs (mean 45.13 vs. 22.50 months; Wilcoxon-test, P < 0.05) (Fig. S1E), suggesting enhanced colonization fitness. Among ARG-carrying plasmids (0.8% of all PLCs), we observed plasticity in gene carriage. Individual PLCs exhibited context-dependent cargo flexibility, carrying different ARGs across hosts or ecological contexts (Fig. S2).
Among 17 “stealth plasmids” identified in this study, pGut1 (Accession ID: NMDCN000A675) was conspicuous for its dominance, being detected in 96.5% of samples. Its prevalence significantly exceeded that of the previously characterized pBI143 plasmid [5] (ranked 11th in our study), establishing pGut1 as a dominant yet previously overlooked member of the “stealth plasmid” group. To comprehensively evaluate the geographical and population distribution of pGut1*,* we analyzed 148 healthy individuals from the US Human Microbiome Project (HMP) [9], detecting pGut1 in 79 samples (53.4%). This finding confirms its widespread occurrence across geographically distinct populations. The difference in detection rates between our Chinese cohort (96.5%) and the HMP cohort (53.4%) may likely reflect geographical variations, dietary influences, or methodological differences in sample processing and sequencing protocols.
Through a combination of PCR amplification and Sanger sequencing, we resolved the complete pGut1 sequence. pGut1 possesses a compact, conserved backbone (~4006 bp) encoding essential genetic elements (Fig. 1A): a replication protein, a recombinase, and a YafQ-RelB toxin-antitoxin (TA) stabilization system. The backbone sequences displayed highly conservation across positive samples (average nucleotide p-distance = 0.00069). In contrast, a large hypervariable region exhibited extensive genetic diversity, harboring 40 distinct cargo insertions identified across individuals. These insertions included multiple ARGs (e.g., cfr(C), erm(B), aphA, linA) and various other functional genes.
The structural plasticity of pGut1 manifested across multiple biological scales. Longitudinal analysis revealed dynamic insertional polymorphism within individuals. For instance, in participant P4, pGut1 was detected in both August and September 2017 samples, with the September variant uniquely acquiring an additional erm gene within its variable region. Similarly, in participant P2, pGut1 maintained persistence across multiple timepoints but exclusively carried the pnuC gene in the January 2022 sample.
This plasmid heterogeneity extended to the single-sample level, where multiple pGut1 variants carrying divergent inserts coexisting within individual fecal specimens. We validated this intra-sample diversity through three orthogonal approaches: First, PCR amplification targeting the variable region yielded multiple distinct amplicon sizes in single samples (Fig. 1B), suggesting structure variation, though potential non-specific amplification required further verification. Second, Nanopore long-read sequencing of pGut1-positive sample 20ZYI1 generated 14 complete pGut1 sequence from 4 785 475 long reads (Avg length 1004 bp), which resolved into five structurally distinct variants carrying unique insertion profiles (Fig. 1C). Third, to confirm variant coexistence at cellular resolution, we reanalyzed single-cell sequencing data (PRJNA803937) from a healthy stool sample. Among 1995 assembled bacterial genomes in a sample, two discrete genomes (SRR27264285 and SRR27274506) each harbored a pGut1 plasmid with functionally divergent variable regions—one encoding a secreted protein and the other a lipoprotein in their variable regions, respectively (Fig. 1D). This profound intra-sample heterogeneity presented significant technical assembly challenges: the coexistence of multiple variable sequences generated complex repeat structures that prevented complete plasmid circularization of pGut1 in short-read metagenomic assemblies. This technical limitation likely contributed to pGut1’s historical obscurity despite its high prevalence. Our multi-platform analytical approach—combining PCR fragment analysis, long-read sequencing, and single-cell genomics—successfully overcame these obstacles, enabling the first comprehensive characterization of these elusive plasmids, and revealing unprecedented plasmid diversity within individual microbiomes.
To investigate the phylogenetic distribution and host range of pGut1, we conducted a comprehensive screen of 2,299,771 publicly available bacterial genomes retrieved from the NCBI genome database (accessed January 2025). pGut1 was identified in 217 genomes. Following filtering to exclude potentially chimeric metagenome-assembled genomes (MAGs), we confirmed its presence in 93 high-quality genomes spanning 53 species, 49 genera, and 2 phyla (Fig. 2A). This broad host range may contribute to the high positive rate of pGut1 observed in the human gut. Phylogenetic analysis based on conserved backbone revealed clustering of closely related pGut1 variants across diverse hosts (Fig. 2A), providing clues for candidate recent cross-species horizontal gene transfer (HGT) events. For instance, two distinct strains of Enterocloster bolteae were found to harbor pGut1 variants, each exhibiting the highest sequence similarity to pGut1 plasmids from different species-Ruthenibacterium lactatiformans and Flavonifractor plautii, respectively, suggesting potential independent inter-species plasmid transfer events.
Host analysis revealed 95.7% of pGut1-positive isolates belonged to the Bacillota phylum, predominantly comprising gut commensals, and anaerobes. However, we identified pGut1 in three strains of the pathogenic Gram-negative bacterium S. enterica (submitted by the US CDC in 2021–2023) (Fig. 2B). These strains possessed different Multi-Locus Sequence Types (MLST) and serotypes, indicating multiple independent acquisition events. Their pGut1 variants carried different inserts: two harbored the erm(B) gene, whereas the third carried rlmN (Fig. 2B). This supports repeated cross-species transfer (even potentially cross-phylum transfer, from Bacillota to Pseudomonadota) of pGut1 from commensal reservoirs into a clinical pathogen, with potential implications for antimicrobial resistance dissemination.
Our study establishes pGut1 as a paradigm for stealth plasmid-mediated resistance transmission. Its combination of high prevalence, genetic promiscuity, and multi-host colonization capacity creates a potentially dangerous vector for incubating pan-resistant infections. Collectively, these findings reveal that the healthy gut plasmidome contains highly adaptable, pre-adapted vectors that silently propagate ARGs, and are poised for cross-boundary dissemination into pathogens. Integrating plasmid-centric metagenomic surveillance into existing AMR monitoring programs is therefore crucial to intercept such emerging threats before they contribute to outbreaks.
Supplementary Material
SFig1_wrag004
SFig2_wrag004
Supplemental_Method_wrag004
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Castañeda-Barba S, Top EM, Stalder T. Plasmids, a molecular cornerstone of antimicrobial resistance in the one health era. Nat Rev Microbiol 2024;22:18–32. 10.1038/s 41579-023-00926-x 37430173 PMC 12440250 · doi ↗ · pubmed ↗
- 2Dabuo B, Abubakari A, Sankah FE. et al. Antibiotics and antimicrobial resistance genes in a gut microbiota as a reservoir—a review. Adv Microbiome Res 2025;2025:6574751.
- 3Debroas D . Global analysis of the metaplasmidome: ecological drivers and spread of antibiotic resistance genes across ecosystems. Microbiome 2025;13:77.40108678 10.1186/s 40168-025-02062-5PMC 11921664 · doi ↗ · pubmed ↗
- 4Smillie C, Garcillán-Barcia MP, Francia MV. et al. Mobility of plasmids. Microbiol Mol Biol Rev 2010;74:434–52. 10.1128/MMBR.00020-1020805406 PMC 2937521 · doi ↗ · pubmed ↗
- 5Fogarty EC, Schechter MS, Lolans K. et al. A cryptic plasmid is among the most numerous genetic elements in the human gut. Cell 2024;187:1206–1222.e 1216.38428395 10.1016/j.cell.2024.01.039PMC 10973873 · doi ↗ · pubmed ↗
- 6Maguire F, Jia B, Gray KL. et al. Metagenome-assembled genome binning methods with short reads disproportionately fail for plasmids and genomic islands. Microbial Genomics 2020;6. 10.1101/2020.03.31.997171 · doi ↗
- 7Watterson WJ, Tanyeri M, Watson AR. et al. Droplet-based high-throughput cultivation for accurate screening of antibiotic resistant gut microbes. Elife 2020;9:e 56998. 10.7554/elife.56998.sa 132553109 PMC 7351490 · doi ↗ · pubmed ↗
- 8Molano LG, Hirsch P, Hannig M. et al. The PLSDB 2025 update: enhanced annotations and improved functionality for comprehensive plasmid research. Nucleic Acids Res 2025;53:D 189–d 196. 10.1093/nar/gkae 109539565221 PMC 11701622 · doi ↗ · pubmed ↗