AKT1 but not AKT2 single nucleotide polymorphisms are associated with the risk of microscopic polyangiitis
Lizhen Li, Huifang Tan, You Peng, Liepeng Chu, Jing Yang, Wenlv Tang, Kui Tan, Shuangshuang Fu, Meili Huang, Meijun Xu, Jinlan Rao, Chao Xue, Yinyin Chen

TL;DR
This study finds that specific genetic variations in the AKT1 gene are linked to a lower risk of microscopic polyangiitis, a type of autoimmune vasculitis.
Contribution
The study identifies AKT1, but not AKT2, as a novel genetic contributor to microscopic polyangiitis risk through SNP and haplotype analyses.
Findings
AKT1 variants rs2498786 and rs1130233 are significantly associated with reduced MPA risk, especially in P-ANCA-positive patients.
AKT1 gene-level analysis shows a strong protective effect (OR = 0.884, P = 0.002), while AKT2 shows no association.
Protective AKT1 alleles correlate with increased expression in immune tissues, suggesting a regulatory mechanism.
Abstract
Microscopic polyangiitis (MPA), a severe antineutrophil cytoplasmic antibody associated vasculitis (ANCA-associated vasculitis, AAV), demonstrates strong clinical association with myeloperoxidase/perinuclear anti-neutrophilic cytoplasmic antibodies (MPO/P-ANCA). While genetic factors are known to contribute to MPA susceptibility, the potential roles of AKT signaling components remain incompletely characterized, with limited data available for AKT1 and even less for its homologous gene AKT2 in this specific disease context. This case-control analysis included 798 participants (202 MPA patients and 596 controls, the latter comprising 387 individuals from the 1,000 Genomes Project), with control groups pooled after confirmation of genetic homogeneity. Genotypes of seven single-nucleotidepolymorphisms (SNPs) (four in AKT1, three in AKT2) with divergent allele frequencies across populations…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
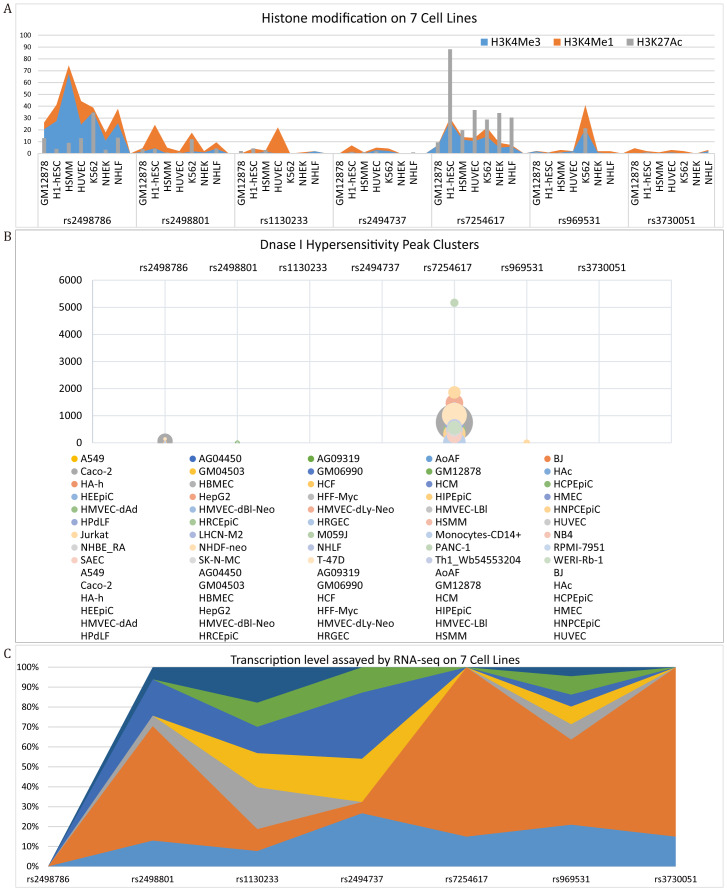 Figure 1
Figure 1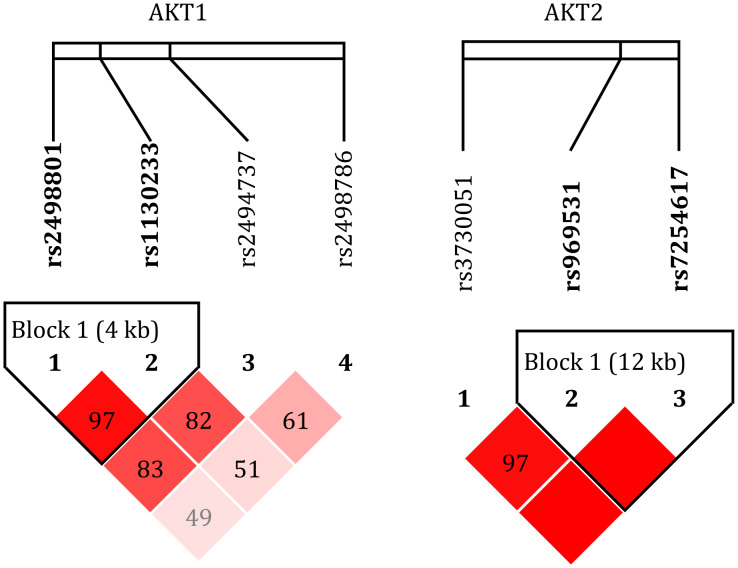 Figure 2
Figure 2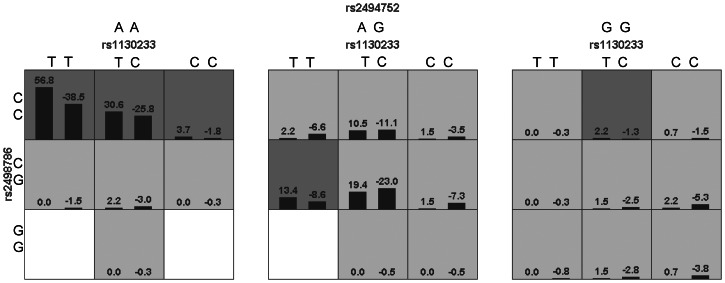 Figure 3
Figure 3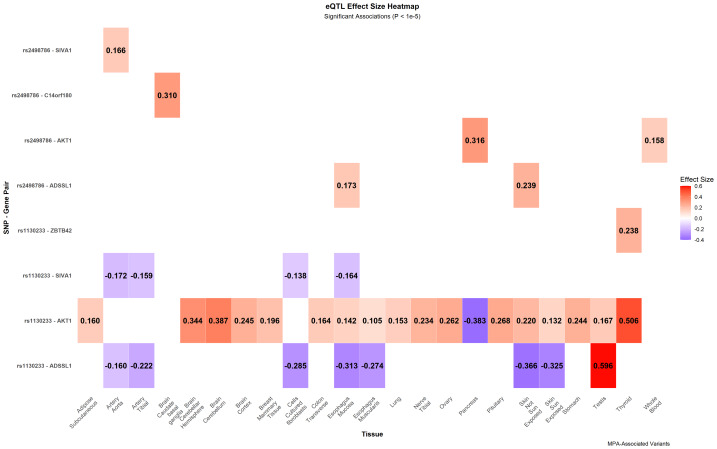 Figure 4
Figure 4- —Guangxi Natural Science Foundation
- —Hunan TCM Administration Research Project
- —Hunan Health Commission Research Project
- —Hunan Education Department Research Project
- —Natural Science Foundation of Hunan Province
- —Natural Science Foundation Youth Project of Hunan Provincial
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVasculitis and related conditions · Otitis Media and Relapsing Polychondritis · Immunodeficiency and Autoimmune Disorders
Introduction
Antineutrophil cytoplasmic antibody (ANCA) are a group of autoantibodies targeting neutrophil cytoplasmic antigens, and are classified into Proteinase 3/Cytoplasmic ANCA (PR3/C-ANCA) and MPO/P-ANCA based on their target antigens and immunofluorescence staining patterns (Bossuyt et al., 2020). ANCA-associated vasculitis is a group of autoimmune disorders characterized by necrotizing inflammation of small-to-medium-sized vessels, with diagnosis contingent upon ANCA serological testing and specific histopathological alterations (Kronbichler et al., 2024). As a core subtype within this disease spectrum, microscopic polyangiitis (MPA) is distinguished by the absence of granuloma formation; its strong association with MPO/P-ANCA not only serves as a diagnostic hallmark (Kronbichler et al., 2024; Guchelaar et al., 2021) but also helps elucidate molecular pathways underlying the pathophysiology of vasculitis at the level of pathogenetic mechanisms (Kitching et al., 2020). In addition to ANCA, genetic factors such as SNPs are also believed to contribute to the pathogenesis of MPA (Trivioli et al., 2022). Numerous studies have demonstrated that polymorphisms in the human leukocyte antigen (HLA) gene were associated with susceptibility to MPA. For example, the HLA-DQB1 (rs1049072) allele was linked to MPA risk in the American population (Merkel et al., 2017), while HLA-DQ (rs5000634) was associated with MPA in the European population (Lyons et al., 2012). Additionally, the HLA-DRB109:01 and HLA-DQA103:02 alleles were identified as risk factors for MPA in the Japanese population (Kawasaki et al., 2023). Research has also established a connection between HLA gene polymorphisms and ANCA. Specifically, HLA-DQA2 (rs3998159, rs7454108) and HLA-DQB1 (rs1049072) were associated with MPO/P-ANCA in the American population (Merkel et al., 2017), whereas HLA-DQA103:02* and DQB103:03* were linked to MPO-AAV in the Chinese population (Wang et al., 2019). Beyond HLA genes, polymorphisms in other genes have also been implicated in MPA. For instance, PTPN22 (rs2476601) (Cao et al., 2015; Martorana et al., 2012; Merkel et al., 2017) and PTPN22 (rs6679677) (Merkel et al., 2017) were associated with MPA in the European population, and FCGR3B copy number variations (Fanciulli et al., 2007; Martorana et al., 2016) have been linked to MPA. Furthermore, the TLR9 (3-SNP haplotype) was associated with both MPA and MPO/P-ANCA (Husmann et al., 2014; Wang et al., 2020), while BACH2 (rs78275221) was found to correlate specifically with MPO/P-ANCA (Dahlqvist et al., 2022). Other genetic factors that may influence MPA risk include CTLA4 (rs3087243), KIR2DS3, and LILRA2 (rs2241524) (Trivioli et al., 2022).
AKT, also known as protein kinase B (PKB), is a serine/threonine kinase and a central regulator of the PI3K/AKT/mTOR pathway, controlling cell survival, metabolism, proliferation, and autophagy (Manning & Toker, 2017). By inhibiting TSC2, a negative regulator of mTORC1, AKT sustains mTORC1 activation (Dibble & Cantley, 2015; Inoki et al., 2002). Consequently, mTORC1 phosphorylates ULK1 at Ser757, preventing its interaction with AMPK and suppressing autophagy initiation (Kim et al., 2011). Additionally, AKT directly phosphorylates ULK1 at Ser774, impairing its autophagy-inducing function, and phosphorylates Beclin-1 at Ser295, disrupting its association with VPS34, which is essential for autophagosome formation (Wang et al., 2012). Furthermore, AKT phosphorylates FoxO1 and FoxO3a, promoting their degradation. Since FoxO transcription factors regulate autophagy-related genes (e.g., LC3, Atg5, Atg7), their inhibition further suppresses autophagy (Schmitt-Ney, 2020). AKT-driven autophagy inhibition is implicated in autoimmune diseases such as rheumatoid arthritis (Keller, Adamopoulos & Lünemann, 2023), systemic lupus erythematosus (Keller, Adamopoulos & Lünemann, 2023; Qi, Zhou & Zhang, 2019), and inflammatory bowel disease (Foerster et al., 2022), exacerbating immune dysregulation and tissue damage. In addition, AKT plays a pivotal role in inflammation by activating the NF-κB pathway, which induces the production of pro-inflammatory cytokines such as TNF-α, IL-6, and IL-1β (Cadwell, 2016; Deretic & Levine, 2018; Matsuzawa-Ishimoto, Hwang & Cadwell, 2018). It influences macrophage polarization, promoting the inflammatory M1 phenotype while suppressing the anti-inflammatory M2 phenotype (Linton, Moslehi & Babaev, 2019; Matsuzawa-Ishimoto, Hwang & Cadwell, 2018). In adaptive immunity, AKT enhances the survival and activation of autoreactive T and B cells, contributing to autoimmune disorders (Cadwell, 2016; Deretic, 2021; Matsuzawa-Ishimoto, Hwang & Cadwell, 2018). Dysregulated AKT signaling disrupts immune balance, leading to persistent inflammation and exacerbating tissue damage in autoimmune diseases (Cadwell, 2016).
ANCA play a critical role in driving inflammation in AAV, a group of autoimmune disorders marked by excessive neutrophil activation and vascular injury (Walulik et al., 2023). Through induction of neutrophil degranulation and reactive oxygen species (ROS) production, ANCA amplify inflammatory cytokine release and endothelial damage (Nakazawa et al., 2019). Autophagy serves a dual function in AAV, balancing neutrophil survival and immune regulation. Chen et al. (2010) identified AKT1 and AKT2—two homologous but distinct genes—as the predominant AKT family members expressed in neutrophils. Given the geographic and gender variations in AAV clinical subtypes and ANCA specificity, we selected four SNPs in AKT1 and three in AKT2 from the 1000 Genomes Project based on their divergent allele frequencies across populations. This selection enabled investigation of the relationship between AKT gene variations and MPA risk, followed by subgroup analyses stratified by P-ANCA status and gender.
Methods
Participants
This study included 798 adults, comprising 202 MPA patients and 209 healthy controls recruited from the Second Affiliated Hospital of Guangxi Medical University (September 2009–October 2023), along with 387 Chinese controls from the 1000 Genomes Project (https://www.internationalgenome.org). To enhance statistical power, the two control groups were pooled after verification of genetic homogeneity via Chi-square tests in SPSS and logistic regression analysis in SNPStats, which confirmed no significant differences in genotype distributions after accounting for sex (Tables S1–S3). The inclusion criteria for MPA patients were: (a) Diagnosis of MPA based on the 2012 Chapel Hill Consensus Conference on Vasculitis criteria. (b) No history of malignancy, infection, or medication-induced vasculitis. Participant demographic features are summarized in Table 1. The MPA group comprised 76 males and 126 females (mean age: 55.19 ± 14.50 years), and 68.81% (139/202) were P-ANCA positive. The control group consisted of 266 males and 330 females.
Table 1: Demographic features of participants.
Ethical approval
The study protocol was approved by the Ethics Committee of the Second Affiliated Hospital of Guangxi Medical University (NO. 2018 KY-0100 and 2024 KY-0782) and conducted in strict accordance with the ethical principles outlined in the Declaration of Helsinki. Written informed consent was obtained from all participants prior to their inclusion in the study.
Methods of obtaining genotypes
Peripheral blood samples were collected from 411 hospital-recruited participants, and DNA was extracted using a Blood DNA Extraction Kit (Tiangen, Beijing, CA). DNA quality was assessed with a Nanodrop 2000 spectrophotometer (Thermo Fisher Scientific, USA), ensuring A260/280 ratios between 1.8–2.0. PCR primers for AKT1 and AKT2 were designed and synthesized (Sangon Biotech, Shanghai, CA). The PCR-amplified DNA was purified using AMPure XP beads, followed by quality assessment via agarose gel electrophoresis. Whole-genome sequencing was conducted using HiSeq XTen (Illumina, USA). The sequencing reads were processed using Cutadapt (v1.2.1) and PRINSEQ-lite (v0.20.3) to remove adapter sequences and filter low-quality reads. SNP genotyping was performed through BWA (v0.7.13-r1126) for alignment and Samtools (v0.1.18) for variant calling. Additionally, SNP genotype data from 387 individuals were obtained from the 1000 Genomes Project (https://www.internationalgenome.org), serving as a reference population.
Gene-level associations analysis
Gene-level associations were assessed through two complementary strategies. First, a likelihood-ratio test was implemented in R by comparing logistic regression models with and without the four AKT1 SNPs, while adjusting for AKT2. Second, a set-based association test was performed using PLINK (v1.9), in which the four AKT1 SNPs and three AKT2 SNPs were grouped into separate gene sets. We further conducted exploratory analyses—including linkage disequilibrium estimation, haplotype reconstruction, and SNP-SNP interaction testing—to examine underlying genetic relationships. Finally, tissue-specific cis-eQTL information for relevant SNPs was retrieved from the 3DSNP database (v2.0).
Statistical analysis
Data statistics were performed via SPSS (version 25.0) and SNPStats (https://www.snpstats.net/start.htm). OR value was calculated by logistic regression analysis. Linkage disequilibrium and Haplotype analysis were performed via SNPStats and Haploview (version 4.1). Interaction analysis was tested by Generalized multifactor dimensionality reduction (GMDR) (version 0.9). To control the false discovery rate (FDR) in multiple comparisons, we used the Benjamini–Hochberg procedure (BH). Statistical significance was determined by P-value <Q-value adjusted by FDR (BH).
Results
Information of SNPs
In this study, we analyzed seven SNPs from the 1000 Genomes Project, whose allele frequencies exhibited significant variations across different populations (Fig. S1). These included four SNPs in AKT1 (rs2498786, rs2498801, rs1130233, rs2494737) and three SNPs in AKT2 (rs7254617, rs969531, rs3730051). A comprehensive characterization of the SNPs was conducted, detailing their genomic locations, predicted functional consequences, and allele frequencies (Table 2, Table S4). This genetic profile was complemented by functional genomic analyses confirming the SNPs’ presence in active regulatory regions (Fig. 1). The rs2498786 and rs7254617 are promoter variants linked to histone modifications (H3K4me3, H3K4me1, H3K27Ac; A) and DNase I hypersensitivity peaks (B) (ENCODE). The rs2498801 and rs969531 function as enhancers. And the rs1130233 (exon) has high transcriptional activity (C). In contrast, the intronic SNPs rs2494737 and rs3730051 were characterized by low levels of associated histone modifications and chromatin accessibility in the ENCODE datasets. The genotypic data for all 798 participants and their association with MPA risk are summarized in Table 2. In this study, the minor allele frequency (MAF) exceeded 0.05 in both the MPA and control groups, thereby satisfying the MAF threshold (MAF > 0.05). Additionally, the Hardy-Weinberg equilibrium (HWE) was maintained (HWE p > 0.05) in the control group. Statistically significant associations with MPA risk were identified for specific alleles of AKT1, namely rs2498786 (P = 0.003) and rs1130233 (P = 0.003). Genotype-based analysis further revealed significant associations for rs2498786 (P = 0.015) and rs1130233 (P = 0.013). In contrast, neither the alleles nor the genotypes of AKT1 rs2494737 or any of the AKT2 SNPs (rs7254617, rs969531, and rs3730051) showed any association with MPA susceptibility (all p > 0.05). Importantly, after FDR-BH adjustment, the initial associations observed for rs2498801 were no longer statistically significant.
Table 2: Information of SNPs in AKT1 and AKT2 genes and MPA risk (MPA, n = 202; Control, n = 596).
Annotation of SNPs from UCSC (ENCODE).The annotation information for the SNPs includes histone modifications (H3K4me3, H3K4me1, H3K27Ac) (A), DNase I hypersensitivity peak clusters across different cell lines (B) and transcription levels in seven cell lines (C).
MPA risk analysis
Association analysis of AKT1 SNPs with MPA risk was performed under four genetic models (Codominant, Dominant, Recessive and Overdominant) using SNPStats (Table 3). The rs2498786 G and rs1130233 C alleles exhibited a negative association with MPA risk in the Codominant, Dominant, and Recessive models. However, after FDR-BH correction, the association of rs2498801 under the Recessive model was no longer statistically significant. By contrast, neither AKT1 rs2494737 nor any of the AKT2 SNPs were associated with MPA risk in any model (Table S5).
Table 3: Association between the SNPs in AKT1 and MPA risk (n = 798, adjusted by sex).
Subgroup analysis of MPA patients with serum P-ANCA positivity is summarized in Table 4. The rs2498786 polymorphism showed consistent associations with P-ANCA-positive status under Codominant, Dominant, and Recessive models (P = 0.024, P = 0.024, and P = 0.032, respectively), aligning with its association with overall MPA risk. The rs1130233 C allele was negatively associated with P-ANCA positivity in Codominant and Recessive models (P = 0.042 and P = 0.032). No significant associations were observed for the remaining SNPs in any genetic model (Table S6), consistent with their lack of association in the primary analysis.
Table 4: SNPs in AKT1 and MPA patients with blood P-ANCA (+) susceptibility analysis (n = 737, adjusted by sex).
A significant interaction with gender was observed for the rs2498786 genotype (P-interaction = 0.007; Table 5), whereas no such interaction was identified for any other tested SNPs. This analysis was conducted via logistic regression on the SNPStats platform, with adjustments for the relevant SNP and gender.
Table 5: Interaction analysis between SNPs in AKT1 (rs2498786) and gender (n = 796).
Gene-level evidence from multivariable logistic regression and set-based analysis
A gene-level analysis was conducted to assess the combined contribution of AKT1 polymorphisms. Using a likelihood-ratio test that compared a full model incorporating all AKT1 and AKT2 SNPs against a reduced model containing only AKT2 variants, we found the AKT1 gene block to be independently associated with MPA susceptibility (χ^2^ = 24.885, df = 4, P = 5.31 × 10^−5^), supporting its role as a risk locus (Table S7). Then, gene-set analyses were performed using PLINK to examine the aggregated effects of AKT1 and AKT2 variants. The analysis identified a significant association between the AKT1 gene set (4 SNPs) and decreased MPA risk (OR = 0.884, 95% CI [0.820–0.953]; P = 0.002), which persisted after FDR adjustment (P-adjusted = 0.004). Conversely, the AKT2 gene set (3 SNPs) demonstrated no significant relationship with disease susceptibility (OR = 1.028, 95% CI [0.902–1.171]; P = 0.686; P-adjusted = 0.686) (Table S8). These findings collectively indicate a protective function specifically associated with AKT1, with no parallel effect observed for AKT2 in MPA development.
Exploratory analysis of SNP-SNP interactions
To evaluate potential genetic relationships, we performed a triad of genetic analyses: linkage disequilibrium assessment, haplotype construction, and SNP-SNP interaction testing.
Linkage disequilibrium analysis
Analysis revealed no significant linkage disequilibrium between SNPs located in the AKT1 and AKT2 gene. However, strong linkage was observed within each gene: the four AKT1 SNPs (rs2498786, rs2498801, rs1130233, and rs2494737) and the three AKT2 SNPs (rs7254617, rs969531, and rs3730051) constituted independent linkage blocks. Specific D’ measurement values are annotated in the corresponding boxes in Fig. 2.
Linkage disequilibrium plot.The linkage disequilibrium plot of the SNPs (rs2498786, rs2498801, rs1130233, rs2494737, rs7254617, rs969531, and rs3730051) was generated using Haploview (version 4.1). The intensity of the color corresponds to the degree of linkage disequilibrium, with darker colors indicating values closer to 100, representing a higher likelihood of linkage disequilibrium. The AKT1 and AKT2 loci exhibited linkage imbalance independently.
Haplotype analysis
Haplotype analysis using the SNPStats platform identified two AKT1 haplotypes significantly associated with MPA risk. Haplotypes G-T-C-T (P-adjusted = 0.003) and C-C-C-A (P-adjusted = 0.025) both demonstrated clear protective effects (Table 6). This indicates that individuals carrying these specific haplotype combinations have a significantly reduced risk of developing MPA.
Table 6: Haplotypes analysis of AKT1 (rs2498786/rs2498801/rs1130233/rs2494737) and MPA risk (adjusted by sex).
Interaction analysis
Using the Generalized Multifactor Dimensionality Reduction (GMDR) method, we analyzed interactions among SNPs screened in this study and previous research (Li et al., 2023). Among various models, the combination of rs2498786, rs2494752, and rs1130233 performed most prominently, achieving a perfect cross-validation consistency score (10/10) and a testing balanced accuracy of 0.5805. The risk analysis schematic (Fig. 3) visually demonstrates that the genotype combination rs2498786 CC, rs2494752 AA, and rs1130233 TT elevates the risk of MPA to the highest level. Complete analysis data are provided in Table S9.
Interactions among SNPs related to MPA risk in the best model.The interactions among SNPs in the model (rs2498786/rs2494752/rs1130233) were visualized using generalized multifactor dimensionality reduction (GMDR, version 4.1). The left bars in each cell represent case positive scores, while the right bars indicate control negative scores. Higher positive scores correspond to an increased risk of MPA. The genotype combination rs2498786 CC, rs2494752 AA, and rs1130233 TT was associated with the highest risk of MPA (P = 0.001).
eQTL analysis of MPA-associated AKT1 variants
Expression quantitative trait locus (eQTL) analysis revealed that the MPA-protective alleles of AKT1 SNPs function as significant cis-eQTLs (Fig. 4). The rs2498786-G allele was associated with enhanced AKT1 expression in whole blood (P = 2.99 × 10^−10^) and pancreas (P = 8.96 × 10^−8^). Similarly, the rs1130233-C allele significantly upregulated AKT1 expression across multiple tissues, most notably in thyroid (P = 1.13 × 10^−40^) and whole blood. Both variants additionally influenced the expression of nearby genes, including SIVA1, in vascular and immune-relevant tissues. These findings indicate that the protective association of these SNPs likely operates through the tissue-specific transcriptional regulation of AKT1 and its regulatory network.
Tissue-specific eQTL landscape of AKT1 variants associated with MPA.Effect sizes of significant cis-eQTL associations for MPA-linked AKT1 SNPs (rs2498786 and rs1130233) across human tissues (3DSNP v2.0), with color intensity representing the direction of allele effect on expression (red: positive; blue: negative). eQTL, expression quantitative trait locus; MPA, microscopic polyangiitis.
Discussion
This study presents accumulating evidence suggesting that the AKT1 gene, in contrast to its homologous counterpart AKT2, may contribute to MPA susceptibility. Our multi-level analyses are consistent with a model in which specific AKT1 variants could confer protection through mechanisms relevant to AAV pathology, whereas AKT2 appears to lack a comparable association.
The functional characteristics of the investigated SNPs provide insights into their potential regulatory roles. The promoter locus rs2498786 reside within genomic regions marked by characteristic histone modifications and DNase I hypersensitivity, indicating their potential as regulatory elements (Lawrence, Daujat & Schneider, 2016; Field & Adelman, 2020). Notably, rs2498786 has been previously associated with altered AKT1 protein levels (Liu et al., 2015). The exonic variant rs1130233, characterized by high transcriptional activity, may influence mRNA stability or protein function (He et al., 2023), a notion further supported by observations of genotype-dependent protein levels (Zubair, Khan & Imran, 2022).
The protective associations of promoter variant rs2498786 and exonic variant rs1130233 with MPA risk, particularly their correlation with P-ANCA positivity—a key diagnostic marker observed in 68.81% of our cohort and 55–70% of historical MPA cases (Hunter et al., 2020; Walulik et al., 2023)—lend support to AKT1’s potential role in AAV-related immune dysregulation. The observed association between protective alleles of these SNPs and enhanced AKT1 expression in whole blood suggests a plausible biological mechanism. Within the context of AAV pathogenesis, where ANCA-mediated Fc-Fcγ receptor engagement promotes neutrophil activation and NETosis via NADPH oxidase-derived ROS (Lu et al., 2021; Shiratori-Aso & Nakazawa, 2023), AKT1’s established function in neutrophil regulatory processes (Chen et al., 2010) raises the possibility that these protective variants may moderate neutrophil hyperactivation and NET formation. This could potentially disrupt the self-perpetuating “ANCA-NETs-ANCA” cycle characteristic of severe vasculitis (Nakazawa et al., 2019), suggesting a potential mechanism linking genetic variation to disease pathology.
Beyond innate immunity, the observed gender interaction with rs2498786, combined with AKT1’s fundamental roles in T and B cell survival and activation (Di Lorenzo et al., 2009; Nagai, Kurebayashi & Koyasu, 2013), extends its potential influence to adaptive immune dysregulation in AAV. The allele-dependent eQTL signals in vascular tissues (aorta, tibial artery) indicate an additional endothelial protective mechanism under ANCA-mediated stress, which aligns with the recognized importance of endothelial integrity in AAV pathogenesis (Dömer et al., 2021; Massicotte-Azarniouch et al., 2022). The coordinated regulation of SIVA1, a gene involved in apoptosis and immune signaling (Shamshul & Yoshio, 2020; Coccia, Solé & Comella, 2021), further supports AKT1’s position within a broader regulatory network maintaining vascular integrity during inflammatory challenge (Lucotte et al., 2023).
Our gene-level analyses, though based on a set of seven SNPs, provide preliminary yet important evidence for the specific involvement of AKT1. The significant association of the AKT1 gene set with reduced MPA risk, contrasted with the null finding for the AKT2 set, underscores the biological specificity of this relationship. This divergence aligns with the well-documented functional differences between these homologous genes: AKT1 drives fundamental immune processes and is highly expressed in immune and vascular tissues (Manning & Toker, 2017), whereas AKT2 exhibits tissue-specific expression with greater emphasis on metabolic regulation and primarily participates in cellular metabolism rather than immune regulation within immune cells (Cho et al., 2001; Kohn et al., 1996). This fundamental distinction, evident in their roles in macrophage polarization (Arranz et al., 2012; Vergadi et al., 2017) and neutrophil regulation (Chen et al., 2010), may explain why AKT1 variants influence MPA susceptibility while AKT2 polymorphisms do not, suggesting our findings reflect specific biological mechanisms rather than general immune pathway disturbances (Chou et al., 2022; Hers, Vincent & Tavaré, 2011).
The haplotype and interaction analyses further substantiate AKT1’s central role in MPA susceptibility. The identification of protective haplotypes (G-T-C-T, C-C-C-A) and the high-risk genotype combination (rs2498786 CC, rs2494752 AA, rs1130233 TT) demonstrates that MPA risk is shaped by complex genetic interactions beyond individual variant effects. These epistatic relationships reveal the intricate genetic architecture underlying this autoimmune vasculitis and highlight the potential utility of multi-locus genetic profiles in risk prediction.
In conclusion, our integrated genetic analysis positions AKT1 as a significant genetic determinant of MPA susceptibility through mechanisms specifically relevant to AAV pathology. The protective effects appear to operate through modulation of neutrophil activation, regulation of adaptive immunity, and protection of vascular integrity—all central pathways in AAV pathogenesis. The consistent absence of association with AKT2 variants across all analytical approaches reinforces the specificity of this relationship. These findings not only advance our understanding of MPA genetics but also suggest that targeted modulation of AKT1 signaling may represent a promising therapeutic strategy for this autoimmune vasculitis.
Conclusions
In summary, this work contributes to growing evidence that AKT1, unlike AKT2, may play a role in MPA susceptibility. The protective associations observed may be mediated through pathways central to ANCA-associated vasculitis, such as those governing neutrophil behavior, immune regulation, and vascular function. These genetic observations help refine the MPA susceptibility landscape and could inform future exploration of AKT1-targeted therapeutic avenues. Further investigation in independent populations and functional characterization of these variants would strengthen these preliminary findings.
Supplemental Information
10.7717/peerj.20791/supp-1Supplemental Information 1Association analysis of AKT1 and AKT2 SNPs between the two control groups under different genetic models (N = 597, adjusted by sex)The two control groups were pooled after verification of genetic homogeneity via logistic regression analysis in SNPStats.
10.7717/peerj.20791/supp-2Supplemental Information 2Chi-square test analysis of genotype distributions between control groupsThe two control groups were pooled after verification of genetic homogeneity via Chi-square tests in SPSS.
10.7717/peerj.20791/supp-3Supplemental Information 3Chi-square test analysis of genotype distributions between control groups by genderThe two control groups were pooled after verification of genetic homogeneity via Chi-square tests in SPSS.
10.7717/peerj.20791/supp-4Supplemental Information 4Information of SNPs in AKT1 and AKT2A comprehensive characterization of the SNPs include their genomic locations, predicted functional consequences
10.7717/peerj.20791/supp-5Supplemental Information 5Association between the AKT2 genotypes and MPA risk (n = 798, adjusted by sex)Association between the AKT2 genotypes and MPA risk
10.7717/peerj.20791/supp-6Supplemental Information 6SNPs ( rs2498801, rs2494737, rs7254617, rs969531 and rs3730051) and MPA patients with blood P_ANCA (+) susceptibility analysis (n = 737, adjusted by sex)No significant associations were observed for the remaining SNPs ( rs2498801, rs2494737, rs7254617, rs969531 and rs3730051) in any genetic model in blood P_ANCA (+) susceptibility analysis
10.7717/peerj.20791/supp-7Supplemental Information 7Assessing the Collective Association of the AKT1 Gene Block with MPA Risk adjusted by sexthe AKT1 gene block to be independently associated with MPA susceptibility
10.7717/peerj.20791/supp-8Supplemental Information 8Association of AKT1 and AKT2 Gene Sets with MPA Risk adjusted by sexA significant association between the AKT1 gene set (4 SNPs) and decreased MPA risk.
10.7717/peerj.20791/supp-9Supplemental Information 9SNP-SNP interaction analysis performed by GMDR
10.7717/peerj.20791/supp-10Supplemental Information 10Annotion of SNPsData of Figs. 1 and 4
10.7717/peerj.20791/supp-11Supplemental Information 11Distribution of alleles among populationsThe allelic frequency distribution across major populations (East Asian, African, Amerindian, and European) in both the 1000 Genomes Project and this study cohort.Data for the 1000 Genomes populations were sourced from publicly available repositories hosted by the National Center for Biotechnology Information (NCBI; https://www.ncbi.nlm.nih.gov/) and the International Genome Sample Resource (IGSR; https://www.internationalgenome.org/).
10.7717/peerj.20791/supp-12Supplemental Information 12Details of VCF file conversions using the IGSR-supported Ensembl tool
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Arranz A Doxaki C Vergadi E Martinez dela Torre Y Vaporidi K Lagoudaki ED Ieronymaki E Androulidaki A Venihaki M Margioris AN Stathopoulos EN Tsichlis PN Tsatsanis C 2012 Akt 1 and Akt 2 protein kinases differentially contribute to macrophage polarization Proceedings of the National Academy of Sciences of the United States of America 1099517952210.1073/pnas.111903810922647600 PMC 3386059 · doi ↗ · pubmed ↗
- 2Bossuyt X Dillaerts D Mahler M Roggenbuch D Leinfelder U Hammar F Schlumberger W Olschowka N Damoiseaux J 2020 Standardisation of PR 3-ANCA and MPO-ANCA: evaluation of certified reference materials Annals of the Rheumatic Diseases 791520152210.1136/annrheumdis-2020-21741632546601 · doi ↗ · pubmed ↗
- 3Cadwell K 2016 Crosstalk between autophagy and inflammatory signalling pathways: balancing defence and homeostasis Nature Reviews Immunology 1666167510.1038/nri.2016.100PMC 534328927694913 · doi ↗ · pubmed ↗
- 4Cao Y Liu K Tian Z Hogan SL Yang J Poulton CJ Falk RJ Li W 2015 PTPN 22 R 620W polymorphism and ANCA disease risk in white populations: a metaanalysis The Journal of Rheumatology 4229229910.3899/jrheum.13143025448792 PMC 4314360 · doi ↗ · pubmed ↗
- 5Chen J Tang H Hay N Xu J Ye RD 2010 Akt isoforms differentially regulate neutrophil functions Blood 1154237424610.1182/blood-2009-11-25532320332370 PMC 2879106 · doi ↗ · pubmed ↗
- 6Cho H Mu J Kim JK Thorvaldsen JL Chu Q Crenshaw 3rd EB Kaestner KH Bartolomei MS Shulman GI Birnbaum MJ 2001 Insulin resistance and a diabetes mellitus-like syndrome in mice lacking the protein kinase Akt 2 (PKB beta)Science 2921728173110.1126/science.292.5522.172811387480 · doi ↗ · pubmed ↗
- 7Chou WC Rampanelli E Li X Ting JP 2022 Impact of intracellular innate immune receptors on immunometabolism Cellular & Molecular Immunology 1933735110.1038/s 41423-021-00780-y 34697412 PMC 8891342 · doi ↗ · pubmed ↗
- 8Coccia E SoléM Comella JX 2021 FAIM-L - SIVA-1: two modulators of XIAP in non-apoptotic caspase function Frontiers in Cell and Developmental Biology 982603710.3389/fcell.2021.82603735083225 PMC 8784879 · doi ↗ · pubmed ↗