Social Processing in the Amygdala: Single-Nucleus RNA-Sequencing Reveals Distinct Neuronal Responses to Dominant and Subordinate Cues
Madeleine F. Dwortz, Hans A. Hofmann, James P. Curley

TL;DR
This study uses RNA sequencing to identify specific neurons in the mouse amygdala that respond differently to social dominance and subordinate cues.
Contribution
The study reveals distinct glutamatergic and GABAergic neuronal responses to social olfactory cues in the amygdala using snRNA-seq.
Findings
A glutamatergic cluster aligned with Slc17a7 (VGLUT1) medial amygdala neurons responds preferentially to dominant cues.
A larger glutamatergic Slc17a6 (VGLUT2) cluster responds more to subordinate cues, highlighting the medial amygdala's role in social olfactory processing.
A GABAergic cluster with elevated Drd2 expression responds to dominant cues, linking dopamine receptor 2 to threat responses.
Abstract
The amygdala serves as a critical neural hub for interpreting social cues, with its distinct subregions and diverse neuronal populations playing specialized roles in processing these signals. Here, we employ single-nucleus RNA sequencing (snRNA-seq) to characterize neuronal responses in the mouse amygdala to olfactory cues associated with social dominance, uncovering distinct activation patterns within glutamatergic and GABAergic populations. We find that a glutamatergic cluster closely aligned with Slc17a7 (VGLUT1) medial amygdala (MeA) neurons preferentially responds to dominant cues. In contrast, a larger glutamatergic Slc17a6 (VGLUT2) cluster associated with MeA, cortical, and basomedial amygdala neurons exhibits a heightened response to subordinate cues, underscoring the MeA’s role in processing social olfactory information. Additionally, a glutamatergic cluster resembling dorsal…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
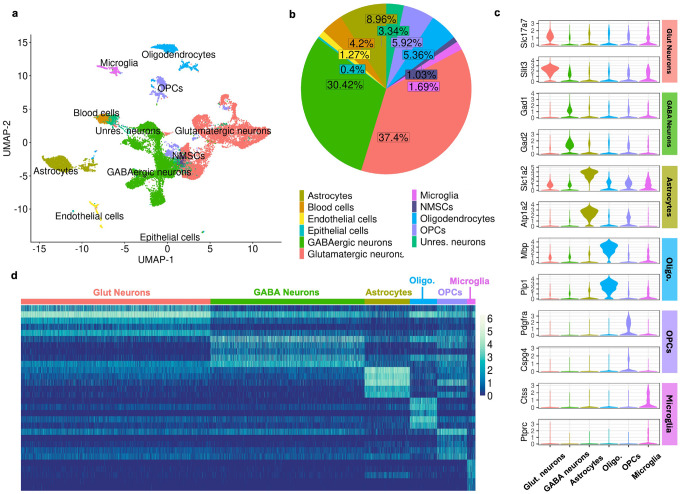 Figure 1
Figure 1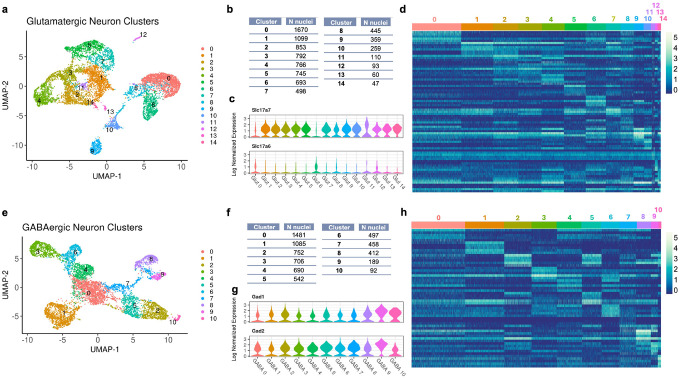 Figure 2
Figure 2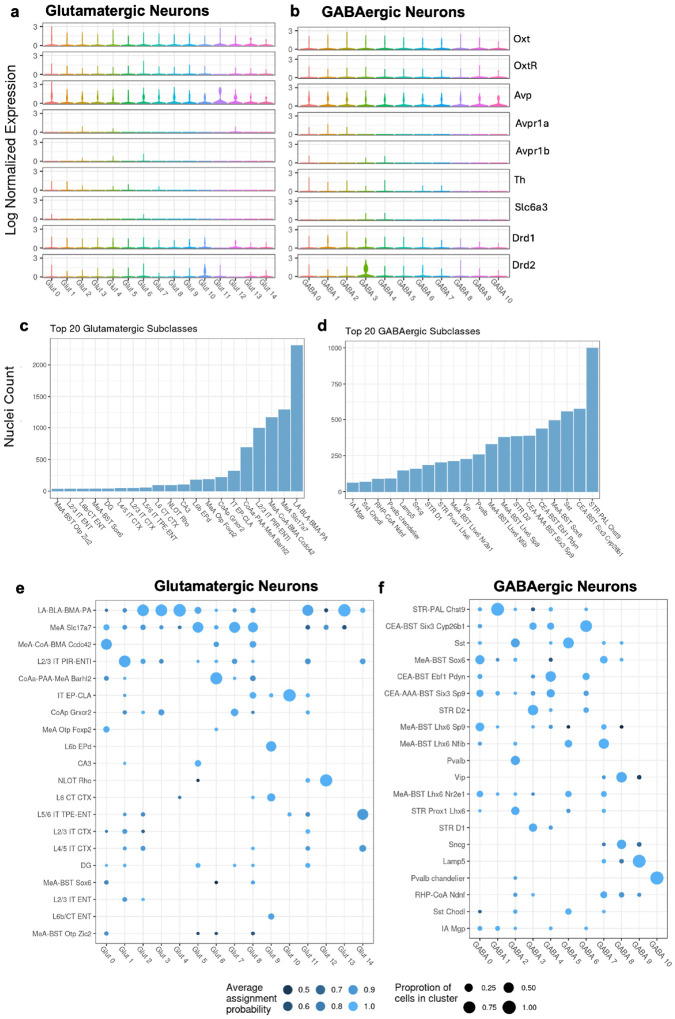 Figure 3
Figure 3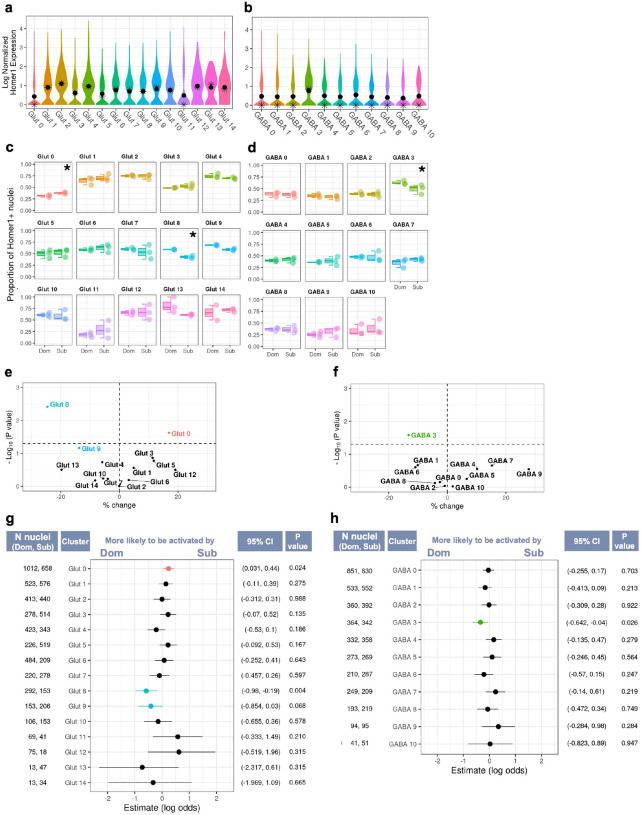 Figure 4
Figure 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsOlfactory and Sensory Function Studies · Neuroendocrine regulation and behavior · Memory and Neural Mechanisms
Introduction
Social perception is a critical neural function for animals living in social groups. The brain’s ability to perceive social context crucially guides appropriate social behaviour, enabling animals to form and maintain strong social relationships (Adolphs 2001, O’Connell and Hofmann 2011, Dwortz et al. 2022). This is especially important in a dominance hierarchy, a common form of social organization, where the contextually appropriate expression of dominant or subordinate behaviour determines an individual’s access to resources and chances of survival. Decades of research show that the amygdala is crucial for proper social functioning in dominance hierarchies and that it may be especially important for the perception of dominance cues (Rosvold, Mirsky, and Pribram 1954, Adolphs 2003). For instance, activity in the primate basolateral amygdala (BLA) is positively associated with the dominance status of perceived individuals’ faces (Zink et al. 2008; Munuera, Rigotti, and Salzman 2018). In mice, amygdala subregions innervated by olfactory neurons, such as the posteroventral medial amygdala (MeApv), have been shown to increase their activity in response to dominant olfactory cues (W. Lee et al. 2021; Veyrac et al. 2011). Our recent work in mice suggests that similar response patterns extend to dorsal and anteroventral MeA subregions (MeAd, MeAav), BLA, and dorsal endopiriform nucleus (EPd) (Dwortz and Curley 2025). While many specific subregions within the amygdala contribute to processing social cues, the extensive heterogeneity of this brain region suggests that interconnected nuclei and diverse neural processes underpin social perception.
The amygdala is a highly heterogeneous structure, and recent advancements in single-cell profiling technologies have greatly enhanced our understanding and appreciation of its cellular diversity (Swanson and Petrovich 1998; Yu et al. 2023; Hochgerner et al. 2023). The amygdala contains a diverse array of neuronal cell types, including excitatory principal neurons and inhibitory interneurons. These cell types are intricately modulated by a complex network of neuromodulators, including dopamine (DA) and neuropeptides such as oxytocin (Oxt) and vasopressin (Avp), which play crucial roles in regulating social behaviours and emotional responses (de la Mora et al. 2010; Yao et al. 2017; Arakawa, Arakawa, and Deak 2010; Fadok, Dickerson, and Palmiter 2009; Lee, Lee, and Kim 2017). The expression of Oxt and Avp receptors is particularly prominent in the medial and central nuclei of the amygdala, areas that are key to processing social stimuli (Dębiec 2005; Ferguson et al. 2001; Ferretti et al. 2019). Moreover, the amygdala hosts a variety of projection neurons that connect different nuclei within the amygdala to other brain regions (Janak and Tye 2015; McGarry and Carter 2017; Kita and Kitai 1990; Dong, Petrovich, and Swanson 2001). Accounting for this cellular diversity is critical for elucidating how the amygdala processes social information.
In the present study, we captured the amygdala’s response to social dominance cues using single-nucleus RNA-sequencing (snRNA-seq). This method allowed us to simultaneously analyze responses of multiple amygdala subregions and characterize activated neurons at a granular, cell-type-specific level. Specifically, we exposed mid-ranking mice living in dominance hierarchies to urinary cues from familiar dominant and subordinate individuals and then used single-nucleus RNA-sequencing (snRNA-seq) to measure transcriptional responses of multiple amygdala subregions. To characterize activated neurons at a granular, cell-type-specific level we examined the expression of Homer1, an immediate early gene (IEG). IEGs are a class of genes that are rapidly and transiently transcribed upon cell membrane depolarization and, therefore, facilitate the identification of activated neurons. We further analyzed neurotransmitter gene expression and mapped neurons to specific amygdala sub-regions with the Allen Brain Cell database to aid in interpreting the functional implications of our findings. We hypothesized that exposure to dominant and subordinate social cues would elicit transcriptionally distinct patterns of neuronal activation across amygdala subregions. Specifically, we expected that dominant social cues would preferentially recruit excitatory neurons within the medial amygdala, reflecting specialized encoding of socially salient olfactory information. Our results confirm the heterogeneity of the amygdala, revealing a significant presence of both glutamatergic and GABAergic neurons, each distinguished by distinct subtypes. Specifically, we observed that glutamatergic VGLUT1 neurons whose transcriptional signatures were most closely aligned with MeA reference profiles exhibited heightened responses to the dominant olfactory cue. We also found that a cluster of glutamatergic VGLUT2 neurons with profiles matching those in the medial-cortical and basomedial amygdala regions showed a stronger response to the subordinate olfactory cue. This cluster notably co-expressed both Oxt and Avp preprohormones at relatively higher levels compared to other clusters. Additionally, we found that Drd2-expressing striatal-like GABAergic neurons predominantly responded to dominant olfactory cues. These results demonstrate that multiple mechanisms within the amygdala work together to process social cues.
Methods
Subjects and housing
We used 18 male CD-1 mice (Mus musculus domesticus) aged 7 to 8 weeks from Charles River Laboratory (Houston, TX, USA). Upon arrival, mice were marked for identification using blue non-toxic markers (Stoelting Co.) and housed in groups of three in standard cages with pine shaving bedding. For the duration of the experiment (25–26 days), subjects were housed in standard cages with pine shaving bedding under a reverse dark–light cycle and provided standard chow and water ad libitum. All procedures were approved by the University of Texas at Austin Institutional Animal Care and Use Committee (IACUC; Protocol No. AUP-2019–00338), were performed in accordance with relevant institutional guidelines and regulations, and are reported in accordance with the ARRIVE guidelines.
Behavioural observations and dominance analysis
Groups were observed for one hour each day for the first five days of group housing and then no less than one hour every other day for the remainder of the study, between 11:00 and 17:00 during the dark phase of the reverse light cycle when mice were most active. During observation periods, observers used all-occurrence sampling to document fighting, chasing, mounting, submissive postures, and fleeing behaviours, noting winners and losers (see Table S1 for ethogram). Behavioural data analysis was then conducted in R Studio using the Compete package (Curley 2016). The cumulative wins and losses for each individual over the housing period were compiled into frequency win/loss sociomatrices for each cohort (Fig S1). From these sociomatrices, we calculated the Directional Consistency (DC) of dominance interactions, and David’s Scores (DS) (de Vries 1995). DC assesses the degree to which all agonistic interactions in a group occur in the direction of the more dominant individual to the more subordinate individual within each relationship. It is equal to (H- L)/(H + L) where H is the frequency of behaviours occurring in the most frequent direction and L is the frequency of behaviours occurring in the least frequent direction within each relationship. The significance of DC values was evaluated using a randomization test (Leiva, Solanas, and Salafranca 2008). DS provides an individual dominance rating and ranking for each individual in a group, determining the overall success of each individual at winning contests relative to the success of all other individuals. Briefly, it is derived from the proportion of wins and losses of each individual and is corrected for the frequency of agonistic interactions (de Vries 1995). DS above 0 typically reflects that an animal is socially dominant whereas a DS below 0 typically indicates that an animal is socially subordinate. The most dominant and subordinate individuals in each hierarchy had the highest and lowest DS, respectively.
Urine collection and odor exposures
Urine collection began one week prior to the odor presentations, starting on days 17 and 18 of group housing, and concluded one day before the odor presentations on days 24 and 25. Urine was collected from the most dominant and subordinate group members between 7 and 10 am, during the light cycle. Urine was collected by gently scruffing the mouse, applying a small amount of pressure on the bladder and collecting deposited urine directly into an Eppendorf tube. Samples were then immediately stored at −80° C.
On the morning of odor presentations, urine collected over the previous week was thawed on ice and then pooled into a single sample that remained at room temperature for approximately one hour. Stimuli were matched by delivery volumes and exposure durations, and donors were familiar home-cage members, reducing novelty-related confounds. We did not perform chemical normalization (e.g., creatinine or total protein) of pooled urine, thereby preserving compositional differences inherent to urine derived from dominant versus subordinate animals and more closely replicating stimuli as they occur naturally in a group-living environment. Odor presentations occurred between the hours of 11 am and 1 pm, during the dark phase of the light cycle. Mid-ranked subjects were first transferred to a separate behavioural testing room and placed in a testing chamber with a thin layer of fresh pine shaving bedding. They habituated to the chamber for 30 minutes and were then gently scruffed and the experimenter pipetted 15 μl of urine onto the nasal groove. Subjects were presented with urine samples from either the most dominant or subordinate member of their home-cage (n = 3). They were then placed back into the chamber, and the experimenter pipetted 1000 μl of urine onto the bedding. Subjects remained in the chamber for an additional 30 minutes and were sacrificed by rapid cervical dislocation followed by decapitation. Immediately upon sacrifice, brain tissue was flash-frozen in dry ice-cooled hexanes and stored at −80°C until further processing.
Amygdala microdissection
Brain tissue was transferred to a − 20° C cryostat chamber (Leica Microsystems), where it was embedded in OCT and incubated for approximately 30 minutes before amygdala dissection. We used the publicly available Allen Mouse Brain Atlas (Allen Reference Atlas – Mouse Brain, atlas.brain-map.org) to identify and section regions of interest. Four 300 μm slices were made onto glass slides, and the amygdala was dissected bilaterally using sterile surgical blades (approximately slides 65 to 77 of the Allen Mouse Brain Atlas). The amygdalae from each of the three mid-rank subjects per stimulus group (dominant and subordinate stimuli) were pooled into a single 0.2 ml RNAse-free tube and stored at −80°F until further processing.
Nuclei dissociation, library preparation and sequencing
Samples were removed from the − 80°C freezer and briefly brought to room temperature. We then isolated nuclei using a procedure adapted from Salem et al. 2024 (Salem et al. 2024). Samples were homogenized in 1.5 ml Nuclei EZ Lysis Buffer (Sigma # NUC101) with 0.2 U/ul RNAse inhibitor (NEB # ML314L) in pre-chilled 2mls KIMBLE Dounce tissue grinders (Sigma Aldrich D8938). Three up-and-down motions of the dounce sufficiently homogenized the samples. Lysate was then filtered through a 35-μm cell strainer and centrifuged at 900 rcf for 5 minutes at 4°C. The pellet was resuspended in 700 μl of a 25% iodixanol mixture (2% BSA in 1X PBS supplemented with 0.2 U/ul RNAse inhibitor mixed with 60% iodixanol Optiprep medium, Sigma-Aldrich # D1556). This suspension was then layered on top of 700 μl of a 29% iodixanol cushion in a new 1.5 ml tube and centrifuged at 8,000 rcf for 30 minutes at 4°C. Supernatant was discarded and the pellet was resuspended in 150 μl buffer (2% BSA in 1X PBS supplemented with 0.2 U/ul RNAse inhibitor). This suspension was again filtered through a 35-μm cell strainer twice. The final eluent with nuclei was stained with DAPI and the concentration of nuclei was approximated using a Countess Automated Cell Counter (Thermo Fisher Scientific Single nuclei libraries were prepared by the Genomic Sequencing and Analysis Facility at UT Austin using the Chromium Next GEM Single Cell 3’ Kit v3.1 (10× Genomics, PN-1000128) with a target cell count of 10,000 cells. Libraries then were sequenced on a NovaSeq 6000 using an S4 flow cell.
Initial snRNA-seq data processing
We used the Cell Ranger pipeline (10x Genomics) to align raw sequencing data to the mouse genome (GRCm39, Ensembl release 2024-A), perform cell calling, and generate gene-by-cell UMI count matrices. These matrices were then imported into R (RStudio, v4.3.1) and processed using Seurat (v4.3.0.1) (Hao et al. 2024; 2021; Stuart et al. 2019; Butler et al. 2018). We used the Souporcell pipeline (Heaton et al. 2020) to demultiplex each sample pool and identify three putative individuals (Fig S2a,b). We then filtered each dataset to ensure comparable cell count and quality between samples. Souporcell also aided in the detection and removal of doublets. (Fig S2c). Poor-quality nuclei were identified based on mitochondrial DNA content and MALAT1 expression. Using MALAT1, a nuclear-retained non-coding RNA universally expressed across cell types, as a quality control metric is a recommended approach in processing single-cell and single-nucleus data, as low MALAT1 expression is indicative of droplets lacking nuclei (Clarke and Bader 2024). We retained cells with 500–7,000 expressed genes, less than 1% mitochondrial DNA, and log normalized MALAT1 expression greater than 5 (Fig S3). Gene counts were normalized with Seurat’s NormalizeData function, which divides by total expression, scales to a common factor (10,000), and applies log transformation (Stuart et al. 2019).
Data integration, clustering and annotation
We integrated the datasets derived from dominant- and subordinate-urine exposed subjects to identify common cell-type clusters using Seurat (Fig S5). This integration reduced batch effects and enabled direct comparison of shared cell-type clusters across conditions, facilitating the identification of cellular heterogeneity and accurate cell-type annotation. To ensure that clustering was based on cell-type similarity rather than activation states, we first removed IEGs. Based on existing literature, the following IEGs were excluded: Btg2, Jun, Egr4, Fosb, Junb, Gadd45g, Fos, Arc, Nr4a1, Npas4, Coq10b, Tns1, Per2, Ptgs2, Rnd3, Tnfaip6, Srxn1, Tiparp, Ccnl1, Mcl1, Dnajb5, Nr4a3, Fosl2, Nptx2, Rasl11a, Mest, Sertad1, Egr2, Midn, Gadd45b, Dusp6, Irs2, Plat, Ier2, Rrad, Tpbg, Csrnp1, Peli1, Per1, Kdm6b, Inhba, Plk2, Ifrd1, Baz1a, Trib1, Pim3, Lrrk2, Dusp1, Cdkn1a, Pim1, Sik1, Frat2, Dusp5, Egr1, Homer1, Bdnf, Nr4a2 (Hochgerner et al. 2023). Next, a set of 2,000 highly variable features was selected to identify consistent patterns of variation across datasets using Seurat’s FindVariableFeatures function. We then computed PCA projections, determining the number of principal components based on the point where the explained variance reached an optimal balance. Seurat’s FindNeighbors function was then used to calculate the nearest neighbors for each cell, followed by the FindClusters function to apply the Louvain algorithm and identify clusters of transcriptionally similar cells (Fig. 1a).
We assigned cell types to the identified clusters using multiple annotation approaches. First, Seurat’s FindAllMarkers function was used to identify cluster-specific marker genes. This function uses a Wilcoxon Rank Sum test to compare gene expression levels between each cluster and all other cells and provides Bonferroni-corrected p-values. In particular, this analysis revealed a distinct cluster of blood cells characterized by high expression of Ttr, Hbb-bs, and Hba-a1 (Hochgerner et al. 2023). To perform reference-based classification, we applied SingleR with reference datasets from Celldex (Aran et al. 2019) to assign broad cell classes including neurons, astrocytes, oligodendrocytes, endothelial cells, and epithelial cells. We next used ScType (Ianevski et al. 2022) to refine annotations of specific subtypes, such as oligodendrocyte precursor cells (OPCs), based on curated marker databases. Finally, we employed the Allen Brain Cell “Map My Cells” (MMC) tool to achieve finer anatomical resolution and facilitate comparability with future datasets. MMC uses a hierarchical reference-mapping algorithm based on the Allen Brain Atlas to assign nuclei to specific cell types and returns a bootstrapped probability score reflecting assignment confidence (Z. Yao et al. 2023).
We examined the results of these various annotation methods in the 10x Loupe Browser (10x Genomics) and assigned cells based on agreement across methods. Cells with conflicting annotations were re-clustered using the full integration and clustering pipeline, enabling the selection of more biologically relevant genes for feature selection. We next analyzed glutamatergic and GABAergic neurons separately, re-running the clustering workflow within each group to resolve finer neuronal subtypes. To identify overexpressed genes associated with each cluster, we again employed Seurat’s FindAllMarkers function.
Identifying differentially activated neuronal populations
We aimed to identify differences in neuronal activity between dominant and subordinate stimuli by comparing the proportions of *Homer1-*expressing (Homer1^+^) neurons within each glutamatergic and GABAergic cluster for each individual mouse genotype from the sample pools. Previous work from our laboratory using in situ IEG labeling (catFISH) demonstrated that activity-induced expression of the IEG isoform Homer1a is selectively associated with social stimulus exposure occurring approximately 30 minutes prior to brain extraction and fixation (Dwortz & Curley, 2025). Given 10× 3’ snRNA-seq quantifies gene-level UMIs, we cannot resolve Homer1a versus constitutive Homer1 isoforms; therefore, we interpret gene-level Homer1 as a proxy for recent activation. Within our dataset we also found that Homer1 was the most highly expressed IEG in neurons and showed positive correlations with other IEGs (Fig S6c,d). Furthermore, Homer1^+^ nuclei were enriched for cyclic-nucleotide-mediated signaling and postsynaptic processes (Fig S7; Table S10), consistent with an activated state.
We applied binomial generalized linear mixed-effect models (GLMMs) to evaluate the association between proportions of Homer1^+^ neurons and social stimuli, incorporating the total number of cells per cluster per individual as weights to account for varying cell counts across clusters and individual subjects. To explore the functional characteristics of differentially ‘activated’ cell populations, we analyzed the MMC subclass annotations assigned to each neuronal cluster. We also used a candidate gene approach and examined neurotransmitter gene expression across subclusters. Additionally, we performed differential gene expression analysis between Homer1^+^ and Homer1^−^ neurons using Seurat’s FindAllMarkers function and Gene Ontology (GO) Enrichment analysis to confirm that processes related to cell activation were activated in these nuclei.
Weighted Gene Co-expression Network Analysis
We also conducted a weighted gene co-expression network analysis for high dimensional data (hdWGCNA) using the hdWGCNA R package to identify modules of co-expressed genes across the integrated datasets for glutamatergic and GABAergic nuclei (Morabito et al. 2023). We selected soft power thresholds of 12 and 10 for glutamatergic and GABAergic nuclei, respectively, and only included genes expressed in at least 5% of nuclei in each cell type. We computed module eigengenes (MEs) using hdWGCNA’s ModuleEigengenes function. Within this function, we adjusted for potential sample quality and technical biases by regressing out the influence of total counts per cell, the percentage of mitochondrial genes expressed, and Malat1 expression. GO enrichment analysis was then conducted using the Enrichr package to determine active biological pathways associated with each module. For each module, the corresponding gene list was queried against the GO:Biological Process, GO:Cellular Component and GO:Molecular Function databases. (Chen et al. 2013). We examined the expression of MEs and pathway activity across neuronal clusters and analyzed the overlap between cluster-specific gene markers and module-associated genes.
Results
Dominance
All six social groups of male mice established dominance hierarchies with a mean Directional Consistency (DC) of 0.752 ± 0.198 (Fig S1; Table S2). The stability and strength of each dominance relationship determined how subjects were assigned a dominant or subordinate stimulus. For instance, in cohort C, the mid-ranking mouse defeated the most dominant individual in nine interactions, resulting in a low DC score of 0.443. However, this mouse consistently defeated the most subordinate individual, indicating a stable dominance relationship with that opponent. Consequently, the mid-ranking mouse in cohort C received urine from the subordinate animal (see Fig S1 for stimulus assignments and Table S2 for full dominance results).
Neuronal diversity within the amygdala
We identified 22,695 nuclei that passed the quality control filters (11,597 cells in the dominant-stimulus sample and 11,098 in the subordinate-stimulus sample) containing 125,267,275 total reads and 33,696 unique genes. These nuclei included neurons, astrocytes, oligodendrocyte precursor cells (OPCs), oligodendrocytes, blood cells, endothelial cells, non-myelinating Schwann cells (NMSCs), and epithelial cells (Fig. 1; see Table S3 for list of cell type gene markers). Neurons clustered robustly into glutamatergic (8,489 cells) and GABAergic (6,904 cells) clusters (Fig. 1a). Ultimately, only a small fraction of unresolved neurons could not confidently be assigned to glutamatergic or GABAergic classes (Fig. 1b). Glutamatergic and GABAergic populations were further clustered, and we identified 16 glutamatergic cell clusters (Fig. 2a–d) and 11 GABAergic cell clusters (Fig. 2e–h; see Tables S4 and Table S5 for lists of neuronal cluster gene markers). Across neuronal clusters, there was some variability in the relative contributions of each genotype by random chance (Fig S5c,f).
We observed cluster-specific expression of Slc17a7 (VGLUT1) and Slc17a6 (VGLUT2) subtypes among glutamatergic clusters (Fig. 2c). Specifically, VGLUT2 neurons were more prominent than VGLUT1 in glutamatergic clusters 0 and 6. In contrast, expression of GABAergic marker genes Gad1 and Gad2 were uniformly expressed across GABAergic clusters (GABAergic marker gene Slc32a1 was very lowly expressed in nuclei) (Fig. 2g).
We also examined the expression of neurotransmitter and receptor genes implicated in social processing across glutamatergic and GABAergic clusters (Fig. 3a,b; Table S6). Oxt and Avp preprohormones were expressed across various subclusters (average log normalized expression in glutamatergic nuclei: Oxt: 0.0740 ± 0.281, Avp:0.218 ± 0.467; GABAergic nuclei: Oxt: 0.0798 ± 0.305, Avp: 0.223 ± 0.484). In contrast, the expression of neuropeptide receptor genes was generally low across all nuclei, which was not unexpected, given that many neuropeptide GPCR mRNAs are localized to distal processes rather than the nucleus or soma, making them less readily detected in snRNA-seq (Cajigas et al. 2012) (average log normalized expression in glutamatergic nuclei: Oxtr: 0.0181 ± 0.130, Avpr1a: 0.000 ± 0.015, Avpr1b: 0.000415 ± 0.0181; GABAergic nuclei: Oxtr: 0.0205 ± 0.148, Avpr1a: 0.001 ± 0.035, Avpr1b: 0.000435 ± 0.0211). However, dopamine receptor genes were detectable (average log normalized expression in glutamatergic nuclei: Drd1: 0.040 ± 0.19, Drd2: 0.0248 ± 0.166; GABAergic nuclei: Drd1: 0.055 ± 0.25, Drd2:0.0837 ± 0.329), GABAergic cluster 3 exhibited especially prominent Drd2 expression compared to other clusters (average log normalized expression: 0.523 ± 0.720) (Fig. 3b, Table S5).
To assess the relative prominence of each subtype within clusters we further analyzed the distribution of neurons across distinct neuroanatomical subtypes using the MMC tool (Fig. 3c–f). Clusters showed enrichment for specific MMC-derived assignments, which were annotated according to their reference brain region and characteristic gene markers. These assignments suggest that the transcriptional profiles of nuclei in our dataset closely resemble those cataloged in the Allen Brain Cell database. While definitive classification of these nuclei cannot be established, their expression patterns indicate strong similarity to known cell types. Most glutamatergic nuclei resembled cells originating from the centralized collection of amygdala nuclei consisting of the basolateral (BLA), basomedial (BMA), lateral (LA) and piriform (PA) amygdala: LA-BLA-BMA-PA. The next most prevalent sources of glutamatergic neurons were the medial (MeA), anterior cortical (CoAa), piriform (PAA) amygdala area, and piriform-entorhinal (PIR-ENT) neurons: MeA Slc17a7, MeA-CoA-BMA Ccdc42, L2/2 PIR-ENT, CoAa-PAA-MeA Barhl2 (Fig. 3c).
GABAergic neurons appeared to be comprised of two primary types: striatal or striatal-like neurons and local inhibitory neurons. The largest proportion was identified as striatal pallidal neurons (STR-PAL Chst9). Prominent sources of GABAergic nuclei also included the central (CEA), medial and anterior (AAA) amygdala and bed nuclei of the striatal terminalis (BST): (CEA-BST Six3 Cyp26b1, CEA-BST Ebf1 Pdyn, CEA-AAA-BST Six3 Sp9). Somatostatin-positive neurons (SST) were also prevalent (Fig. 3d).
Differential activation across neuronal clusters
Consistent with the stimulus timeline, in which stimulus onset occurred approximately 30 min prior to fixation, Homer1 was the most highly expressed IEG in glutamatergic and GABAergic neurons (Fig S6a,b) and showed strong correlations with longer-timescale IEGs such as Bdnf (Tao et al., 1998) (Fig S6c,d). Homer1 expression was generally higher among glutamatergic neurons, with the highest expression in glutamatergic cluster 2 (Glut 2) (mean = 1.11 ± 0.84) (Fig. 4a). Among GABAergic neurons, cluster 3 (GABA 3) exhibited the highest expression (mean = 0.778 ± 0.747) (Fig. 4b).
To assess whether Homer1 expression is associated with neuronal activation and to identify additional genes associated with this active state, we compared transcriptional profiles of Homer1^+^ and Homer1^−^ nuclei (see Table S9 for full DEG results). We found overexpression of phosphodiesterase (PDE) genes in glutamatergic and GABAergic Homer1^+^ cells (Fig S7a-d). PDE’s control intracellular signaling by catalyzing second messenger cyclic nucleotides, which in turn control receptor-effector coupling, protein-kinase cascades, and transmembrane signal transduction. GO enrichment analysis of these overexpressed genes revealed terms related to “cyclic-nucleotide-mediated signaling” (see Table S10 for GO analysis results). We specifically observed overexpression of PDE10a in both glutamatergic and GABAergic Homer1^+^ neurons, along with Pde4b and Pde7b in glutamatergic and GABAergic Homer1^+^ neurons, respectively. Additional GO enrichment terms associated with Homer1^+^ neurons also underscored the processes related to post-synaptic activation, such as “regulation of postsynaptic membrane neurotransmitter receptor levels” and “protein localization to postsynaptic membrane”. This is consistent with the established role of Homer1^+^ in interacting with excitatory metabotropic glutamate receptors at the postsynaptic density (Xiao et al. 1998; Vazdarjanova et al. 2002). In contrast, genes overexpressed in Homer1^−^ nuclei were associated with GO terms related to cell homeostasis, such as “maintenance of location in cell” and “regulation of cellular component size”.
We applied general linear mixed-effect models (GLMMs) for binomially distributed data to proportions of Homer1^+^ nuclei within each individual mouse genotype and neuronal cluster to determine if there was an association with the social status of the olfactory stimulus. We observed significant differences in the proportion of Homer1^+^ nuclei in several glutamatergic and GABAergic clusters, indicating that specific populations respond more to urine derived from either dominant or subordinate donors (Fig. 4c–h; see Table S8 for full GLMM results).
Glutamatergic cluster 8 (Glut 8) exhibited significantly greater proportions of Homer1^+^ nuclei in response to dominant stimuli compared subordinate stimuli (Sub – Dom: β = −0.583 ± 0.202, P = 0.004, 95% CI: −0.980, −0.190). Notably, this cluster was enriched for excitatory MeA neurons. Within this cluster, 54.8% of nuclei were confidently identified as MeA Slc17a7 neurons with a 99.0% average probability of assignment accuracy (avg. probability), while MeA-CA-BMA Ccdc42 neurons constituted 14.3% (avg. probability = 95.6%) (Fig. 3e; see Table S7 for full MMC assignments). Additionally, proportions of Homer1^+^ nuclei in glutamatergic cluster 9 (Glut 9) trended higher in response to dominant compared to subordinate stimuli (Sub – Dom: β =−0.410 ± 0.225, P = 0.068, 95% CI: −0.854, 0.028). Glut 9 was the only cluster enriched for dorsal endopiriform neurons (EPd) at 58.6% (avg. probability = 98.2%) (Fig. 3e).
In contrast, glutamatergic cluster 0 (Glut 0) exhibited significantly greater proportions of Homer1^+^ nuclei in response to subordinate stimuli compared to dominant stimuli (Sub – Dom: β = 0.238 ± 0.105, P = 0.024, 95% CI:0.031,0.444). Cluster Glut 0 was predominantly composed of excitatory MeA-CA-BMA Ccdc42 neurons, accounting for 70.5% (avg. probability = 97.8%). (Fig. 3e, Table S7). Notably, this cluster exhibited elevated expression of Oxt and Avp preprohormone transcripts compared to other neuronal clusters. Specifically, it showed the highest Oxt preprohormone expression and the second highest Avp preprohormone expression (average log normalized expression of Oxt: 0.110 ± 0.363; Avp: 0.316 ± 0.602; Fig. 3a, Table S6).
We also observed stimulus-associated Homer1^+^ expression among GABAergic neurons. GABA 3 exhibited significantly greater proportions of Homer1^+^ nuclei in response to dominant compared to subordinate stimuli (Sub – Dom: β = −0.341 ± 0.153, P = 0.026, 95% CI: −0.642, −0.041). This cluster also exhibited robustly higher Drd2 expression compared to all other neuronal clusters (Fig. 3b). In line with this observation, this cluster was enriched for striatal DRD2-expressing neurons (STR D2) based on the MMC analysis (Fig. 3f).
To better understand and characterize the amygdala’s complex neuronal populations, we used hdWGCNA to identify and analyze modules of co-expressed genes distributed across neuronal clusters. Additionally, we performed Gene Ontology (GO) enrichment analysis on these modules, uncovering active biological processes in these clusters. We discovered five modules within glutamatergic neurons and seven within GABAergic neurons (Fig S8, Fig S9; see Table S11 and Table S12 for module gene assignments). We focused specifically on clusters exhibiting stimulus-associated Homer1^+^ expression. Glut 0, which showed a higher proportion of Homer1^+^ nuclei in response to subordinate stimuli, exhibited elevated expression of the brown module eigengene (ME) (Fig S9a,b). This was also supported by a significant overlap between its marker genes and those of the brown module (Fig S9g). GO terms associated with this module included terms included “chloride transport” and “regulation of postsynaptic membrane potential” (Fig S9e; see Table S13 for GO analysis results). In response to dominant stimuli, Glut 9 exhibited a higher number of Homer1^+^ nuclei and showed elevated expression of the green module, the smallest module composed of only 28 genes (Fig S9b, Fig S9g). GO terms associated with the green module were related to “cell-cell adhesion” and “cell junction assembly” (Fig S9e). Glut 8 did not exhibit elevated expression of any particular module, although it did share some marker genes with the brown and green modules (Fig S9g). GABA 3, which showed a higher proportion of Homer1^+^ nuclei in response to dominant stimuli, exhibited a high degree of overlapping marker genes with the blue module and elevated expression of the blue ME (Fig S9c,d,g). The top GO terms for the blue module included terms related to cell activation, including “regulation of cation channel activity”, “chemical synaptic transmission” and “protein phosphorylation” (Fig S9f).
Discussion
The amygdala serves as a central hub for processing social information and encompasses diverse subregions and neuronal types (Janak and Tye 2015, Li et al. 2017, Adolphs 2003, Hochgerner et al. 2023,). In the present study, we employed snRNA-seq to identify specific neuronal populations within the amygdala activated by social urinary olfactory cues. Our analysis revealed distinct clusters of glutamatergic and GABAergic nuclei, each characterized by neurotransmitter systems and enrichment for specific neuroanatomical cell types. The heterogeneity found in our dataset largely corroborates existing amygdala taxonomy revealed by single-cell RNA-seq studies (Hochgerner et al. 2023). This neuronal diversity highlights the need for cell-type-specific analyses of social processing in the brain.
We identified several neuronal clusters that were differentially activated, as indicated by Homer1 expression, in response to dominant and subordinate urinary olfactory cues under matched volume and exposure conditions. We intentionally did not chemically normalize urine across stimulus donors, as status-related differences in urinary composition constitute the natural sensory signals through which social status is conveyed. Consistent with this view, prior work, including studies from our laboratory (Lee et al. 2017), demonstrates that dominant males produce higher concentrations of specific urinary pheromones such as darcin, SBTs, and farnesenes, which function as status-linked chemosignals (Novotny et al. 1990, Roberts et al. 2010). Thus, neural activation patterns observed here are best interpreted as responses to biologically relevant cues encoding social status. While chemical profiling of urine is beyond the scope of the present study, the identification of status-responsive neuronal populations provides a foundation for future studies linking specific pheromonal components to circuit-level responses.
We found that a glutamatergic cluster, Glut 8, enriched for MeA Slc17a7 (VGLUT1) neurons, responded more to dominant stimuli. This finding aligns with our previous work showing that dominant urinary cues elevate c-fos immunoreactivity in the MeApv, a region with a high concentration of glutamatergic neurons (Keshavarzi et al. 2014; W. Lee et al. 2021). The MeA plays a critical role in processing social olfactory cues, acting as a pivotal relay station within the accessory or vomeronasal olfactory pathway, which primarily processes pheromonal cues (Raam and Hong 2021; Kevetter and Winans 1981; Dulac and Wagner 2006). Numerous studies have shown that activity within the MeA responds to socially-relevant olfactory cues (Li et al. 2017; Bergan, Ben-Shaul, and Dulac 2014; Carvalho et al. 2015; Choi et al. 2005). Our previous work supported the theory that the MeApv specifically plays a crucial role in discerning status-related cues early in the olfactory information processing stream, as it responded robustly to dominant cues regardless of familiarity with the individual (W. Lee et al. 2021). This context strengthens our current finding that Glut 8, as one of the few neuronal clusters in the amygdala that shows differential responses, aligns with the MeApv’s specific function.
Interestingly, we observed an opposite response pattern in Glut 0, which was enriched for MeA-CoA-BMA Ccdc42 neurons. Glut 0 exhibited the highest and second-highest levels of Oxt and Avp preprohormone expression among all neuronal clusters, respectively. Numerous studies have confirmed the critical role of the oxytocin receptor (OxtR) in the medial amygdala (MeA) for processing social odors and recognizing socially relevant cues (Li et al. 2017; S. Yao et al. 2017). Oxytocin (Oxt) release in the MeA is also implicated in social memory formation (Ferguson et al. 2001). In contrast, the function of vasopressin (Avp) release in the MeA is not as well understood. Similar to OxtR, Avp receptors (Avpr1a and Avpr1b) are essential for social recognition (Albers 2012; Wang et al. 2013) and facilitate social approach behaviours in the MeA (Arakawa, Arakawa, and Deak 2010). Moreover, Avp signaling in the amygdala is linked to aggression promotion (Bosch and Neumann 2010) and exhibits sexually dimorphic expression influenced by androgens, which may underpin its role in aggression (Tong, Abdulai-Saiku, and Vyas 2021). It is important to note that neuropeptide receptor transcripts appeared at low levels in our dataset, which is expected because many of these mRNAs are enriched in dendrites rather than in the nucleus (Cajigas et al., 2012). Since snRNA-seq isolates nuclear RNA only, it tends to underdetect such cytoplasm-localized transcripts (Bakken et al., 2018). Consistent with this, whole-cell scRNA-seq studies of the amygdala report higher receptor detection (O’Leary et al., 2020; Hochgerner et al., 2023).
Glut 0 was also one of two glutamatergic clusters that exhibited low VGLUT1 expression and more prominent VGLUT2 expression compared to other clusters. Indeed, VGLUT2 neurons are the primary glutamatergic subtype intermingled with GABAergic neurons in the MeA and biomedical amygdala (BMA), further validating this cluster’s MMC annotation (Hochgerner et al. 2023). Along with the observed increase in Glut 8 activity in response to dominant cues, these results imply that VGLUT1 and VGLUT2 expressing neurons in the amygdala may have distinct roles in processing social olfactory signals. One of the key distinctions between VGLUT1 and VGLUT2 transporters lies in their association with different glutamate release patterns. VGLUT1 is associated with a low probability of glutamate release, which is linked to a susceptibility to long-term potentiation (LTP), a process that strengthens synaptic connections and supporting learning and memory; in contrast, VGLUT2 is associated with to a high probability of glutamate release and is receptive to long-term depression (LTD), a mechanism that weakens synaptic connections and is also important for the refinement of learning and memory circuits. (Fremeau et al. 2004; Park et al. 2019). How these opposing yet complementary mechanisms underlie differential responses to social stimuli remains unclear. One possible explanation is that activation of VGLUT1-positive neurons in response to dominant cues may enhance the memory of these cues, reinforcing the importance of remembering dominant individuals. In contrast, VGLUT2-positive neuron activation in response to subordinate cues may engage LTD-related mechanisms associated with adaptive forgetting, in which weakening synaptic connections serves to selectively reduce the salience of less behaviorally relevant social information and maintain flexibility in ongoing social interactions (Ryan and Frankland 2022). Further research is required to clarify the unique contributions and potential interactions of these spatially proximal but functionally distinct neuronal populations in processing social cues.
We also observed an increase in response to dominant stimuli in the Glut 9 cluster, which exhibited an expression profile similar to EPd neurons. This observation is consistent with recent studies showing glutamatergic EPd neurons exhibit heightened immediate early gene (IEG) expression following exposure to socially dominant individuals, compared to subordinate ones (Dwortz and Curley 2025). The EPd appears to be multifunctional, interfacing with areas involved in olfaction (e.g., olfactory bulb, medial amygdala, perirhinal cortex), emotional responses (BLA), memory (e.g., entorhinal cortex), and sensorimotor functions (Neafsey, Hurley-Gius, and Arvanitis 1986; Behan and Haberly 1999; Meis et al. 2008; Watson, Smith, and Alloway 2017). Emerging research also suggests that the EPd regulates interoceptive states, given its consistently high baseline activity during exploratory behaviour in mice and during slow-wave sleep in humans (Manjila et al. 2024; Ponomarenko, Korotkova, and Haas 2003). Additionally, some have proposed that EPd activation may contribute to conscious odor perception, as evidenced by situations in which human subjects not only detect an odor but also consciously acknowledge its presence (Traub and Whittington 2022). These findings collectively indicate that the EPd may be involved in integrating internal and external states. Thus, differential EPd activation in response to olfactory cues may indicate the recognition of relative dominance between the subject and the cue.
We also observed differential activity in specific GABAergic neurons. We identified a GABAergic cluster characterized by elevated expression of Drd2, which closely resembled striatal DRD2-expressing neurons. This raises the possibility that our dataset includes dorsal striatal neurons in proximity to the CEA. Alternatively, this cluster could consist of striatal-like DRD2-expressing GABAergic neurons identified within the CEA itself (McCullough et al. 2018; McDonald 1982). The significance of DRD2 signaling within the amygdala includes its role in modulating sociability (Ike et al. 2024), risk avoidance and impulsive behaviours (Kim et al. 2018; Casey et al. 2022). Thus, DRD2 activation in response to dominant odors may reflect social vigilance and avoidance of potentially agonistic interactions. Furthermore, polymorphisms in the Drd2 gene are linked to variations in amygdala activity and emotional responses to visual social cues in humans (Blasi et al. 2009). Given these insights, our findings suggest that activation of amygdala Drd2 neurons is involved in evaluating socially relevant cues across various sensory modalities.
We also performed hdWGCNA analysis on glutamatergic and GABAergic nuclei to investigate the organization of active biological processes in our dataset. This analysis identified modules of co-expressed genes whose expression patterns correlated with specific neuronal clusters, supporting the idea that these clusters represent biologically meaningful groups with specialized roles. Gene Ontology (GO) analysis further illuminated what these active processes might be. For instance, in Glut 9, corresponding to glutamatergic EPd neurons, we observed elevated expression of the green module, which is enriched for genes associated with cell-cell adhesion and junction assembly, suggesting that these processes may be particularly relevant for this population.
Interestingly, not all clusters exhibited overexpressed modules, implying that some clusters might contain neurons serving more generalized functions. For example, Glut 8, associated with glutamatergic MeA neurons, lacked distinctive module overexpression, indicating a potentially broader functional role. Collectively, these findings underscore the cellular diversity in the amygdala, comprising both highly specialized neuronal populations with distinct roles and more generalized populations that may support a range of processes and adapt flexibly to various contexts.
Conclusion
In conclusion, this study illustrates the utility of single-nucleus transcriptomic approaches for studying neural responses to socially relevant sensory cues. We observe stimulus-associated activation patterns in specific neuronal populations. Specifically, two groups of VGLUT1 neurons, with transcriptional profiles aligning with the MeA and EPd, respectively, respond more to dominant cues. Conversely, VGLUT2 neurons corresponding to the MeA-Coa-BMA region exhibited elevated responses to subordinate cues. The distinct responses of VGLUT1 and VGLUT2 neurons within the MeA region suggest a MeA-centered circuit may play a key role in mediating responses to social cues. Additionally, the increased expression of both Avp and Oxt preprohormone mRNAs in the MeA-Coa-BMA population underscores the importance of neuropeptide signaling in regulating social perception. Our results also implicate dopamine DRD2 signaling in inhibitory neuron populations in processing social cues. Future studies should expand the number of sequencing samples to replicate our findings. Ongoing advances in single-cell and single-nucleus RNA-sequencing technologies have improved scalability by reducing costs and increasing throughput, thereby facilitating larger and more comprehensive studies (Qu et al. 2023). Further research should not only confirm these transcriptional responses but also elucidate the causal relationships between olfactory cues and amygdala neuronal activity and circuit function.
Supplementary Files
This is a list of supplementary files associated with this preprint. Click to download.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Adolphs R. The Neurobiology of Social Cognition. Curr. Opin. Neurobiol. 11 (2), 231–239. 10.1016/S 0959-4388(00)00202-6 (2001).11301245 · doi ↗ · pubmed ↗
- 2Adolphs R. Is the Human Amygdala Specialized for Processing Social Information? Ann. N. Y. Acad. Sci. 985 (1), 326–340. 10.1111/j.1749-6632.2003.tb 07091.x (2003).12724168 · doi ↗ · pubmed ↗
- 3Albers H. E. The Regulation of Social Recognition, Social Communication and Aggression: Vasopressin in the Social Behavior Neural Network. Hormones and Behavior, Oxytocin. Vasopressin Social Behav. 61 (3), 283–292. 10.1016/j.yhbeh.2011.10.007 (2012). · doi ↗
- 4Allen Institute for Brain Science. Allen Reference Atlas – Mouse Brain [brain atlas] (Available from atlas.brain-map.org, 2011).
- 5Arakawa H., Arakawa K. & Deak T. Oxytocin and Vasopressin in the Medial Amygdala Differentially Modulate Approach and Avoidance Behavior toward Illness-Related Social Odor. Neuroscience 171 (4), 1141–1151. 10.1016/j.neuroscience.2010.10.013 (2010).20933576 · doi ↗ · pubmed ↗
- 6Aran Dvir, A. P. Reference-Based Analysis of Lung Single-Cell Sequencing Reveals a Transitional Profibrotic Macrophage. Nat. Immunol. 20 (2), 163–172. 10.1038/s 41590-018-0276-y (2019).30643263 PMC 6340744 · doi ↗ · pubmed ↗
- 7Behan M. & Haberly L. B. Intrinsic and Efferent Connections of the Endopiriform Nucleus in Rat. J. Comp. Neurol. 408 (4), 532–548. 10.1002/(SICI)1096-9861(19990614)408:4<532::AID-CNE 7>3.0.CO;2-S (1999).10340503 · doi ↗ · pubmed ↗
- 8Bergan J. F., Ben-Shaul Y. & Dulac C. Sex-Specific Processing of Social Cues in the Medial Amygdala. e Life 3 (June), e 02743. 10.7554/e Life.02743 (2014).24894465 PMC 4038839 · doi ↗ · pubmed ↗