Comprehensive evaluation of EEG spatial sampling, head modelling and parcellation effects on network alterations in idiopathic generalized epilepsy
Christina Stier, Markus Loose, Carmen Loew, Marysol Segovia-Oropeza, Sangyeob Baek, Holger Lerche, Niels K N Focke

TL;DR
Low-density EEG can detect brain network changes in idiopathic generalized epilepsy, though high-density EEG is better for detailed spatial mapping.
Contribution
The study shows that low-density EEG arrays can reliably detect global alterations in idiopathic generalized epilepsy, particularly in signal power.
Findings
Lower-density EEG arrays reliably identified global group differences in power and connectivity.
Spatial similarity dropped for connectivity with fewer than 64 channels.
Coarser brain parcellations increased stability for low-density maps.
Abstract
Idiopathic generalized epilepsy is characterized by marked brain network alterations as assessed using electrophysiology. Logistical challenges and the need for a volumetric MRI often hinder the clinical application of high-density EEG or magnetoencephalography. This study investigates the influence of EEG channel density and the head model on brain metrics derived from 256-channel EEG and 19-channel routine EEG in two samples balanced for age and sex. First, we evaluated resting-state data from 35 individuals with idiopathic generalized epilepsy and 54 healthy controls collected using the 256-channel setup. Data were analysed at full density and then iteratively downsampled to lower densities. Source reconstruction was performed either using individual MRI data or a standard brain template and dynamic imaging of coherent sources. We assessed EEG power and connectivity (imaginary part…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
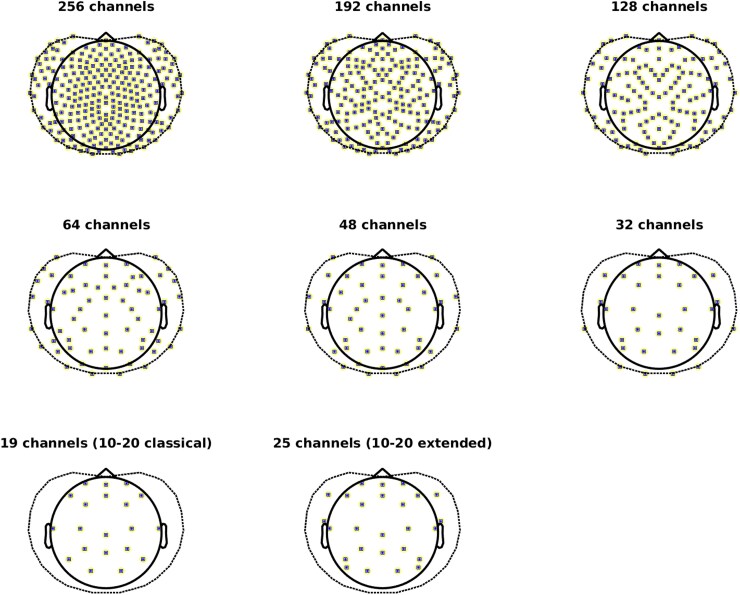 Figure 1
Figure 1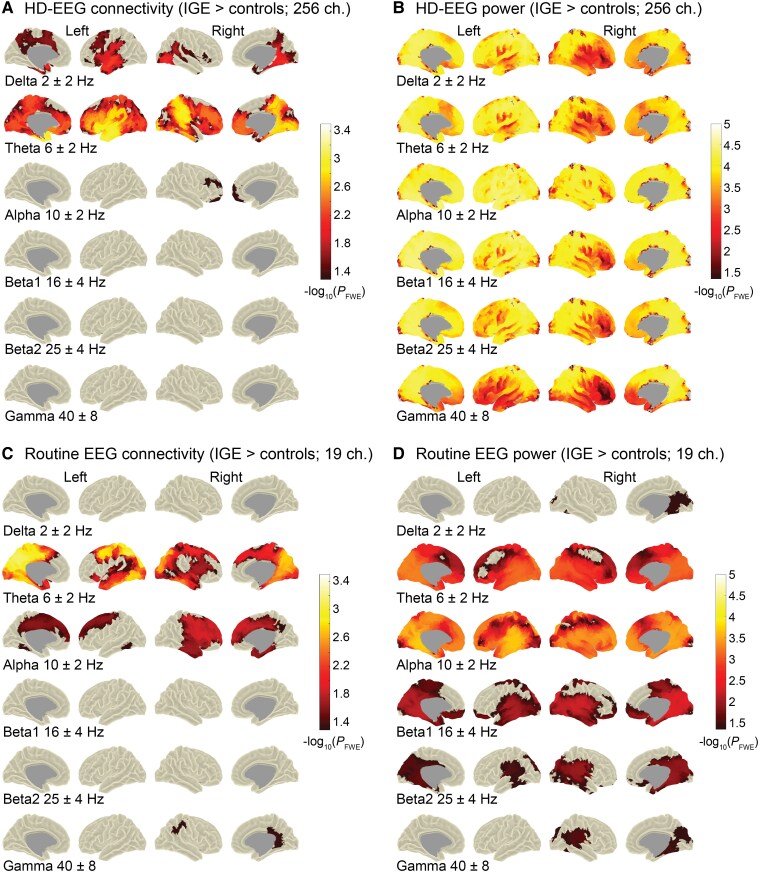 Figure 2
Figure 2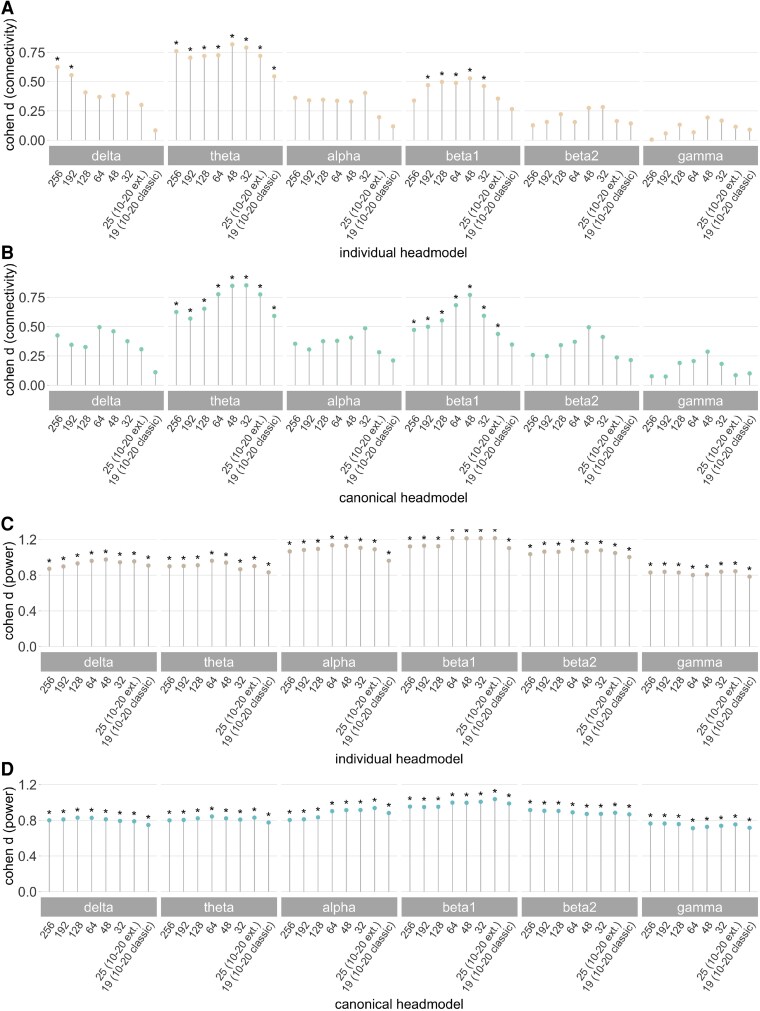 Figure 3
Figure 3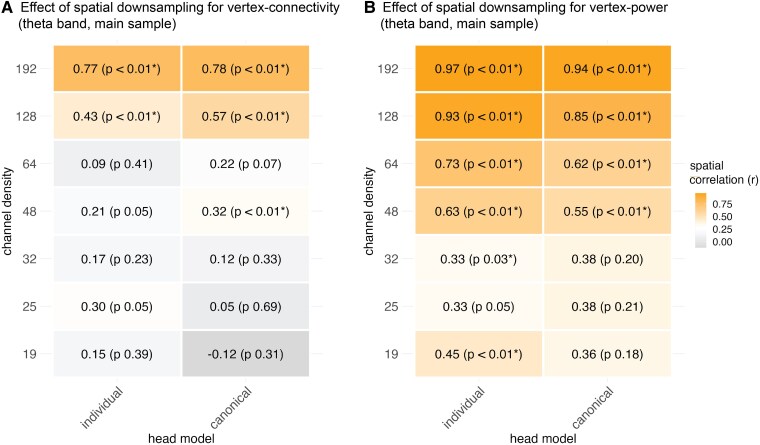 Figure 4
Figure 4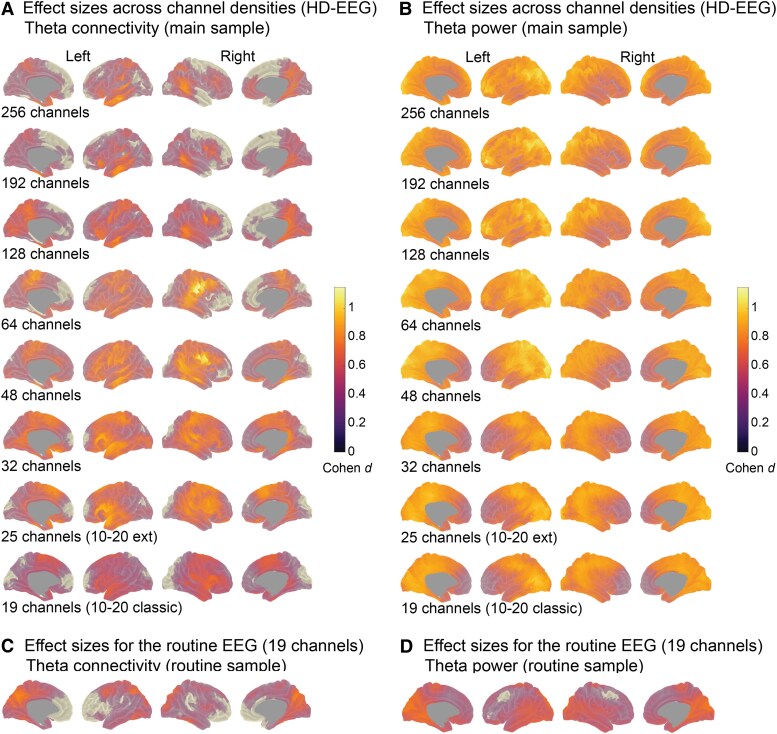 Figure 5
Figure 5| HD-EEG sample | (Clinical) routine sample | |||
|---|---|---|---|---|
| Patients | Controls | Patients | Controls | |
| Total ( | 35 | 54 | 71 | 43 |
| Site: total | 10/25 | 15/39 | 71/0 | 43/0 |
| Female, | 18 (72) | 27 (49) | 35 (49) | 26 (60) |
| Mean age (y), (SD) | 32 (11.6) | 31 (12.2) | 29 (10.2) | 28 (8.8) |
| GSWD during EEG recordings, | 13 (37) | 0 (0) | 19 (27) | 0 (0) |
| Seizure free > 12 months, | 19 (54) | 55 (77) | ||
| Drug intake at measurement, | 2 (6) | 13 (10) | ||
| Epilepsy syndrome, | ||||
| CAE | 5 (14) | 3 (4) | ||
| JAE | 10 (29) | 7 (10) | ||
| JME | 7 (20) | 23 (32) | ||
| GTCS | 6 (17) | 19 (27) | ||
| IGE | 7 (20) | 19 (27) | ||
| Individual head model | Canonical head model | ||||||
|---|---|---|---|---|---|---|---|
| Channels |
|
|
|
|
|
| |
| Connectivity | 192 | 1.02 | 1.29 | 0.74 | 1.05 | 1.22 | 1.07 |
| (theta band) | 128 | 0.46 | 0.47 | 0.18 | 0.65 | 0.65 | 0.76 |
| 64 | 0.09 | 0.20 | 0.03 | 0.22 | 0.20 | 0.21 | |
| 48 | 0.21 | 0.19 | −0.12 | 0.33 | 0.31 | 0.01 | |
| 32 | 0.17 | 0.37 | 0.33 | 0.12 | 0.23 | 0.12 | |
| 25 | 0.31 | 0.22 | 0.20 | 0.05 | 0.17 | 0.04 | |
| 19 | 0.15 | 0.18 | −0.09 | −0.12 | 0.09 | −0.34 | |
| Power | 192 | 2.09 | 2.65 | 2.65 | 1.74 | 2.09 | 2.65 |
| (theta band) | 128 | 1.66 | 1.83 | 1.95 | 1.26 | 1.66 | 2.30 |
| 64 | 0.93 | 1.02 | 1.53 | 0.73 | 0.95 | 1.74 | |
| 48 | 0.74 | 0.78 | 1.07 | 0.62 | 0.73 | 1.33 | |
| 32 | 0.34 | 0.18 | 0.02 | 0.40 | 0.41 | 0.41 | |
| 25 | 0.34 | 0.22 | −0.01 | 0.40 | 0.40 | 0.37 | |
| 19 | 0.48 | 0.40 | 0.33 | 0.38 | 0.38 | 0.48 | |
| Channel sets (Individual ∼ canonical head model maps) |
|
|
|
|
| |
|---|---|---|---|---|---|---|
| Connectivity | 256 | 0.50 | <0.01 | 0.55 | 0.81 | 1.00 |
| (theta band) | 192 | 0.60 | <0.01 | 0.69 | 1.07 | 1.38 |
| 128 | 0.62 | <0.01 | 0.73 | 1.02 | 0.33 | |
| 64 | 0.57 | <0.01 | 0.65 | 1.00 | 0.62 | |
| 48 | 0.52 | <0.01 | 0.58 | 0.76 | 0.00 | |
| 32 | 0.60 | <0.01 | 0.69 | 0.93 | 0.81 | |
| 25 | 0.73 | <0.01 | 0.93 | 1.22 | 1.07 | |
| 19 | 0.65 | <0.01 | 0.78 | 1.00 | 1.02 | |
| Power | 256 | 0.63 | <0.01 | 0.75 | 1.21 | 1.62 |
| (theta band) | 192 | 0.66 | <0.01 | 0.79 | 1.22 | 1.42 |
| 128 | 0.65 | <0.01 | 0.77 | 1.11 | 1.49 | |
| 64 | 0.68 | <0.01 | 0.84 | 1.38 | 1.95 | |
| 48 | 0.77 | <0.01 | 1.02 | 1.80 | 2.16 | |
| 32 | 0.68 | <0.01 | 0.82 | 1.08 | 1.01 | |
| 25 | 0.70 | <0.01 | 0.87 | 1.33 | 1.26 | |
| 19 | 0.73 | <0.01 | 0.94 | 1.33 | 1.62 |
- —German Research Foundation10.13039/501100001659
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFunctional Brain Connectivity Studies · Epilepsy research and treatment · EEG and Brain-Computer Interfaces
Introduction
Since the invention of EEG, the EEG systems have undergone a constant development and remained an essential tool in the diagnostics of epilepsy. Today, systems with a dense spatial coverage are available and generally expected to lead to a better spatial resolution than sparse arrays. The current guidelines by the International Federation of Clinical Neurophysiology (IFCN) in 2017 have been tailored for the clinical diagnostic use, which recommends the use of at least 25 electrodes in a standard array, and the 10-10 system or high-definition systems with 64 to 128 electrodes for source localization purposes.^1^ Several studies have demonstrated improvements in the source localization of interictal or ictal epileptic discharges using high-density recordings.^2-4^ In other (single) cases, the localization of seizure onset zones^5^ and the mapping of the language areas improved,^6^ and epileptic oscillations at rapid scales could be identified.^6^ Others argue that high-density EEG (HD-EEG) not necessarily improves diagnostic accuracy if the number of spikes is sufficiently high^7^ or data recordings long enough.^8^
In general, evaluations of high-density over low-density systems have been limited to diagnostics in the pre-surgical settings and patients with suspected focal epilepsies. However, in recent years, a network perspective of epilepsy and its mechanisms has emerged and prompted a series of studies investigating whole-brain networks in generalized and focal seizure types. In this context, the effects of spatial sampling on brain-wide mapping of source-reconstructed EEG signals have not been sufficiently examined. While there is accumulating evidence for altered interictal, spectral network alterations in patients with IGE,^9-12^ it is unclear to what extent these results can be expected for data derived from a clinical setting. The clinical standard in epilepsy monitoring units is often still limited to the conventional low-density recordings as it is less costly and time-intensive than a high-density montage. To leverage the wealth of clinical EEG databases and assess the potential of resting-state brain dynamics for diagnostic purposes, a side-by-side evaluation of data acquired in clinical and research settings using similar methods is essential. Advanced brain-wide modelling should also provide spatial insights into the underlying effects, enabling differentiation between epilepsy syndromes and other neurological disorders. Furthermore, in resource-restricted countries and individuals with IGE, volumetric MRI scans may not be available for precise source reconstruction.
We therefore set out a study to compare resting-state brain activity and synchronization between patients with IGE and controls measured using a 256-channel EEG. We focused on two well-established metrics such as power^9,13-15^ reflecting the squared signal amplitude, and the imaginary part of coherency^16^ as our connectivity measure, since liability to pathological synchronization has been described even during the interictal state.^17-20^ We subsequently repeated the group-level analyses after virtually reducing the number of channels, applying conventional source reconstruction based either on individual MRI anatomy or on a brain template (hereinafter referred to as the ‘canonical’ head model). Although accurate tissue boundaries generally improve source reconstruction solutions^21,22^ and can enhance localization in focal epilepsy,^4^ the added value for whole-brain, group-level studies in generalized epilepsies remains unclear. To mirror previous work in IGE, we first tested group differences at brain surface vertices and then repeated the analyses after aggregating data into anatomical regions and functional networks, evaluating whether coarser parcellations yield more consistent results across channel densities. Finally, we validated the findings in an independent cohort recorded with a routine 19-channel EEG, thereby assessing the effects under typical clinical conditions. The overarching aim of this work is to provide an overview of spatial sampling effects on brain metrics commonly used to quantify neurophysiological alterations in epilepsy.
Materials and methods
Participants
We considered healthy individuals and individuals diagnosed with IGE according to the International League Against Epilepsy^23^ from two sites. Participants included in the HD-EEG sample were recruited and measured through the Department of Neurology, University Hospital of Tübingen, Germany, between 2013 and 2019, or through the Clinic of Clinical Neurophysiology, University Medical Center of Göttingen, Germany, between 2018 and 2020. The local ethics committee of the Medical Faculties in Tübingen (ethics number 646/2011BO1) and Göttingen (ethics number 16/10/17) approved the studies, compliant with the Declaration of Helsinki. In total, 35 patients and 54 controls were included in the HD-EEG sample, of which data from 22 patients and 34 controls were re-analysed from a previous study.^11^ The second sample included 71 patients and 43 controls undergoing a routine clinical procedure (see next section). These data were acquired between 2007 and 2021 in the Clinic of Clinical Neurophysiology, Göttingen. The need for informed consent was waived for this retrospective study by the local ethics committee (ethics number 2/5/21). In general, only participants with normal MRI scans or non-specific findings (e.g. cysts) were considered in the study. Clinical information can be found in Table 1. All controls were free of neurological or psychiatric conditions, and none of them were taking any medication at the time of the measurement. The two samples were comparable in age (FHD-EEG = 0.17, PHD-EEG = 0.68; Froutine = 0.92, Proutine = 0.34) and sex (χ^2^HD-EEG = 0.0, PHD-EEG = 1; χ^2^routine = 0.93, Proutine = 0.33).
EEG recordings
Individuals from the HD-EEG sample underwent 30 min of continuous resting-state recordings using a 256-channel system (GES400; Magstim EGI) with a sampling frequency of 1 kHz. All individuals were instructed to keep their eyes closed while relaxing, not to fall asleep, and not to think of anything in particular. For the routine sample, a 19-channel EEG with classical 10–20 montage^24,25^ was used with sampling frequency of 500 Hz and 20 min of recording time conducted according to clinical standard in the routine neurophysiological laboratory. As such, the patients of this sample underwent a routine clinical procedure with periods of hyperventilation, tests for the Berger effect, and in a few cases intermittent photic stimulation. Controls for the clinical sample were recorded in the same laboratory with the same equipment, and tests for Berger effect were equally applied, but no hyperventilation and photic stimulation.
Head models
In the HD-EEG study, each individual had a sagittal T1-weighted MRI acquisition (3D-MPRAGE, repetition time = 2.3 s, echo time = 3.03 ms, flip angle = 8° (Tübingen) or 9° (Göttingen), voxel size = 1 × 1 × 1 mm) subsequent to the EEG recordings either using a 3T Siemens Trio (12-channel headcoil; 12/54 controls; 4/35 patients) or Prisma/Prisma^fit^ scanner (64-channel headcoil; 42/54 controls; 31/35 patients). As anatomical information was not available for all individuals of the routine sample, we used an MRI template for all individuals based on 225 T1 and FLAIR images with a large field-of-view customized for EEG head model generation.^26^ To project sensor level data onto cortical surfaces, we applied FreeSurfer reconstruction to T1 images and used SUMA^27^ to resample the cortical surfaces (density factor = 10) based on a standard template (‘fsaverage’). This procedure allowed anatomical correspondence between individuals at 1002 vertices per hemisphere serving as our source points. Volume conduction models were built using the individual or template-derived cortical meshes, brain segmentations via SPM12 processing,^28^ and the EEG electrodes realigned to the anatomical landmarks (nasion and pre-auricular points) and projected onto the scalp mesh. We used a three-layer boundary element model (‘openmeeg’^29^) to compute the leadfields using Fieldtrip^30^ in MATLAB (R2018b).
EEG processing and source analysis
Using Fieldtrip,^30^ we filtered the raw data (first-order Butterworth filter at 1 and 70 Hz), applied a discrete Fourier-transform filter to remove line-noise at 50 Hz and its harmonics (100 and 150 Hz), downsampled to 150 Hz, demeaned and segmented the data into trials of 10 s length. All EEGs were re-referenced to a common average reference online. Trials were visually inspected and those containing artefacts (movements, muscle artefacts and sensor jumps) rejected. In case of spike-wave discharges occurring during the EEG recordings, the respective, the preceding and following trial were removed. In the routine EEG sample, only trials with eyes-closed and trials without external stimulation parameters plus 90 s recovery were considered for further analysis. We applied independent component analyses to detect and (manually) reject components with cardiac and eye movements. All remaining trials were reviewed a second time and vigilance of the individuals was rated according to the criteria of the American Academy of Sleep Medicine (https://aasm.org/). Thirty clean data trials rated as awake (>50% alpha activity within a trial) were then randomly drawn for each individual as we have shown that EEG power and connectivity metrics are reliable for 5 min of data.^31^ To project the data from the electrode-level to the surface points, we applied fast Fourier spectral analyses using multi-tapers (DPSS) for six frequency bands (centre frequency ± smoothing: delta: 2 ± 2 Hz, theta: 6 ± 2 Hz, alpha 10 ± 2 Hz, beta1 16 ± 4 Hz, beta2 25 ± 4 Hz and low gamma 40 ± 8 Hz). Based on this, cross-spectral densities and EEG power were estimated and frequency-dependent source projection performed using Dynamic Imaging of Coherent Sources^32^ (regularization: 5%). At each frequency band, coherency between all pairs of vertices (n = 2004) and power was computed for each vertex. The absolute imaginary part of the coherency coefficient was used as connectivity measure in this study to account for potential fieldspread.^16^ All connections between vertices were then averaged to yield an estimate of synchronization of a vertex, and, for individual global estimates, also across all vertices.
Channel reduction and statistical procedure
To test the impact of spatial sampling on statistical comparisons between the patients and controls, electrode layouts were created with 192-, 128-, 64-, 32-channel maps, and maps with 19 and 25 channels based on the original 10–20^24,25^ and extended 10–20 montage,^1^ respectively, following the guidelines of the IFCN. The full 256-channel layout was used as original template. For the 192 to 48 channel-maps, channels from the original 256-layout were removed iteratively based on the distance to neighbouring channels. Corresponding channels positioned on the other hemisphere were equally removed. Channels assigned to the extended 10–20 positions were kept in the reduction process (see Fig. 1). For each channel set, the cleaned electrode-level data of each individual in the HD-EEG study sample was selected, leadfields re-computed and source projection repeated.
Channel compositions at different densities. Shown are the channel layouts used to test the impact of spatial sampling on group comparisons in the HD-EEG study sample. All compositions rely on the original EGI Magstim 256-layout as a template. The number of channels was reduced in an iterative process, yielding compositions of different densities. Channels assigned to the positions of the classical 10–20 montage were kept throughout (see Methods in the main manuscript for further details). The outline of the layout was plotted using the Fieldtrip toolbox30/GPL license.
Group-level statistics
To assess whether patients differed from controls in power and connectivity, we tested one-sided contrasts using permutation analyses of linear models using PALM^33^ at each of the surface vertices and once for the global metrics. Age, sex and scanner site were included as regressors of no interest. P-values were computed from the resulting empirical distribution and threshold-free cluster enhancement (TFCE),^34^ and familywise error (FWE) corrected at the cluster level. We used tail approximation with 5000 permutations for accelerated inference.^35^ The significance level was set to −log_10_(P) = 1.3 (equivalent to P < 0.05). This statistical procedure was carried out for each channel set in the HD-EEG sample as well as for the routine sample measured using low-density EEG. To determine standardized group mean differences, we used Cohen d computed on the t-values of the group factor of the linear models adjusted for the effects of the covariables. A d ≥ 0.8 indicates a large, d = 0.5 a medium, and d = 0.2 a small effect.
Spatial correlations of effect size maps
To assess the similarity of the group-level results among the different channel sets, we correlated the original effect size map (256 channels) with the maps for the reduced channel sets (192 to 19 channels). We focused on the EEG differences in the theta band, as this has yielded the strongest and most consistent results in the present study and others.^36^ We applied the spin-test^37^ to assess statistical significance of the Spearman rank coefficients based on 1000 random rotations of the spherical cortical surface of each hemisphere separately as it takes the spatial embeddedness of the correlated cortical maps into account.
Channel densities across parcellation schemes
To evaluate group differences (IGE versus controls) at different cortical resolutions, we remapped individual power and connectivity by averaging across vertices within each of 68 brain regions defined by Desikan et al.^38^ and 14 functional resting-state networks defined by Yeo et al.^39^ We then ran separate analyses for each parcellation scheme using PALM (see section ‘Group-level statistics’) and, as described above, computed spatial correlations between the results for the 256-channel maps and the maps for the virtually reduced channel sets (192 to 19 channels). To ensure comparability of the results among the different resolutions, we standardized the correlation coefficients using Fisher’s z transformation^40^ and estimated the probability of the z-differences between the vertex-space and the Desikan regions and Yeo networks, respectively, using R.^41^ We controlled the false discovery rate (FDR) for seven comparisons (original versus lower-density maps) at each type of head model and EEG metric.
Impact of the head model type
We further tested whether the group-level results for the HD-EEG sample were different depending on whether individual or canonical head models were used. For this purpose, the respective Cohen d maps for the results in the theta band were correlated within each channel set using the spin-test as described in 2.7. (1000 permutations). Again, Fisher’s z transformation of the coefficients and FDR correction of the P-values were applied at the level of channel sets (n = 8) to compare the effects across parcellation schemes (vertices, regions and networks).
Results
Group contrast at vertex-based resolution (individual head models)
Group contrasting at the vertex-resolution using a 256-channel EEG and individual head models yielded significantly higher connectivity and power in patients with IGE than in controls. For connectivity, this was mainly the case in the theta frequency band in frontal, centrotemporal and posterior brain regions (Fig. 2A) and also globally (dtheta = 0.76, Ptheta < 0.001), indicating overall increased levels in the patients. Weaker patterns were also observed in the delta and alpha frequency bands (Fig. 2A), which reached significance at the global level only in delta (ddelta = 0.62, Pdelta < 0.01). There were no vertex-wise or global significant differences in the beta and gamma bands (P > 0.05). Increased power in the patients was significant across the brain and the frequency spectrum (delta to gamma, global d: 0.83–1.12, P < 0.001), with emphasis on posterior brain regions (Fig. 2B).
Vertex-based group comparisons using high-density (256 channels) and low-density EEG (19 channels). Highlighted are vertices on the cortical surface for which the patients with IGE from the HD-EEG sample (n = 35) and the routine sample (n = 71), respectively, had significantly higher connectivity (A, C) and power (B, D) than the controls (nHD-EEG = 54, nroutine = 43). Age, sex and measurement site were included as covariates into the permutation-based and frequency-specific analysis of linear models. The significance level was set at −log10(P) = 1.3 (equivalent to P = 0.05). Shown are the results after family wise error correction (FWE) and threshold-free cluster enhancement (TFCE). For the HD-EEG sample (A, B), the analyses were performed using individual head models and 256 EEG channels. The routine sample (C, D) was measured using 19 channels with a classical 10–20 montage and analyses were performed using a template head model as described in the Methods. Similar to the results for the HD-EEG sample, the strongest effects were observable for theta connectivity. In the same vein, increased power in the patients was distributed across the frequency spectrum with a posterior focus, but with generally weaker effects than in the HD-EEG study sample. Ch. = channels; centre frequency ± smoothing using multi-tapers.
Differences in global levels among channel sets
To provide a global estimate and comparison of the contrast IGE > controls for different channel sets (Fig. 1), we report standardized group mean differences in Fig. 3 (Cohen d). For global connectivity, the significance threshold (PFDR < 0.05) was exceeded for all channel densities only in the theta band (d > 0.5), irrespective of the head model type used. Increased connectivity in the delta frequency band was observed for 256- and 192-channels using an individual head model (d > 0.5, PFDR < 0.05) and also in the beta1 band for both head models (individual: d > 0.4 for 192-48 channels; canonical: d > 0.4 for 256-25 channels; PFDR < 0.05). No significant differences in any of the channel sets and head model types were observed in the remaining frequency bands, i.e. alpha, beta2 and gamma (d < 0.37, PFDR > 0.05). Increased global power in the patients was significant for all channel sets and head models (individual: d > 0.8; canonical: d > 0.7).
*Group differences (IGE > controls) in global connectivity and power at different channel densities. Each coloured dot represents standardized effect sizes (Cohen d) for increased connectivity (A, B) and power (C, D) in the patients with IGE (n = 35) compared with the controls (n = 54) of the HD-EEG sample based on permutation analysis of linear models. Separate analyses were run for each channel density and head model type (beige: individual, turquoise: canonical). All d-values were computed based on the respective t-value of the group factor and corrected for the influence of age, sex and measurement site. A d ≥ 0.8 indicates a large, d = 0.5 a medium, and d = 0.2 a small effect and allows direct comparison of the results for EEG metrics and head model types. Statistically significant group comparisons after correction for multiple comparisons at the level of channel sets within each frequency band are indicated with an asterisk (PFDR < 0.05). Electrode layouts were created with 192-, 128-, 64-, 32-channel maps, and maps with 19 and 25 channels based on the original 10–20 (10–20 classic) and extended 10–20 montage (10–20 ext.).
Effect of spatial sampling for vertex-connectivity (theta band, individual head models)
To quantify to what extent the spatial EEG patterns vary depending on the channel density, we focused on the group contrasts in the theta frequency band (IGE > controls) as this turned out to be the strongest global effect across channel sets and head model types. The virtual channel reduction generally influenced the group contrast in source space stronger for connectivity than for power (Fig. 4). Correlations between connectivity maps (Cohen d) for 256 channels and those from 192- and 128- channels were high (r_s_ = 0.77-0.43, Pspin_fdr < 0.05), but dropped to r_s_ = 0.21-0.09 (Pspin_fdr > 0.05) with 64 channels and fewer (Fig. 4). To visualize spatial differences, we subtracted the effect size maps for each channel density from that of 256 channels (Supplementary Fig. 1). The high-density sampling revealed stronger connectivity differences in parietal and frontal brain areas compared to lower channel densities. Conversely, the effect sizes for frontocentral and temporal regions were larger at low channel densities than at high densities. In general, the sampling strategies differed at a maximum of d ± 0.6 from the full set.
Spatial correlation analyses of group contrasts in the theta band across channel densities. We used Spearman’s rank correlation (r) to quantify the spatial similarity between the effect size maps (Cohen d) for EEG levels (IGE > controls) based on the 256-channel density and each lower-density channel set (192-19 channels). The correlation analyses were performed for the group contrasts at 2004 cortical vertices for A) connectivity and B) power. * Indicates significance of the correlations for the vertex-space using the spin-test and after controlling the FDR at the level of channel set comparisons (n = 7). Pspin-values are indicated in brackets.
Effect of spatial sampling for vertex-power (theta band, individual head models)
For comparability with the connectivity results, we focus on the theta frequency band for power as well. The correlations between the effect size maps at 256 channels for theta power and the maps at the other channel sets were significant for all densities except for 25 channels (r_s_ = ∼0.97-0.45, Pspin_fdr < 0.05; r_s__25-channels = 0.33, n.s., Fig. 4). In terms of spatial variability, full sampling (256 channels) yielded stronger power differences in the prefrontal cortex than for the lower densities, particularly for 48 channels and below, but smaller differences in the middle temporal areas (Supplementary Fig. 2).
Sampling effects across parcellation schemes
We further tested whether remapping of EEG metrics from the vertices to anatomical regions or functional networks were more robust to the virtual reduction of channels. Thus, we statistically compared the transformed correlation coefficients derived from the comparisons between the d-maps at 256 channels and the other channel sets (Table 2). The correlations for connectivity did not differ between the vertex-resolution and the Desikan-Killiany parcellation and the Yeo network-resolution, respectively (Pfdr > 0.05, individual and canonical head models). The regional resolution yielded significantly higher coefficients than at the vertices for power at the 192- (individual and canonical head model) and 128-channel densities (canonical head model). Similarly, higher coefficients were observed for the network- than for the vertex-resolution for the 192- to 64-channel sets (canonical head models), suggesting that the group results for power at the high-density sets were more stable using anatomically defined parcellations.
Comparison of head model types
We assessed the influence of the head model (individual versus canonical) on the group-level results in the theta band (Table 3). In the vertex-space, the effect size maps based on individual head models significantly correlated with those based on the canonical head model for each channel set. This was the case for EEG connectivity (r_s_ = 0.50–0.73, Pspin_fdr < 0.05) and power (r_s_ = 0.63–0.77, Pspin_fdr < 0.05). The regional resolution yielded higher concordance between the types of head model than the vertex-resolution for connectivity (192-64 and 25 channels) and power (all channel sets). The network-based (Yeo) analysis also provided higher correlation coefficients than in the vertex-space, but only for power in a few channel sets (256-48 and 19 channels).
Validation in a clinical context
To validate the results derived from the virtual channel reduction approach, we conducted the same processing and analysis principles on data acquired in a clinical setting using a classical routine 10–20 EEG system and a canonical head model. As with the HD-EEG sample and the virtual 19-channel set, increased connectivity was observed in patients compared with controls with the most pronounced effects in the theta band and effects in alpha (Figs 2 and 5). In a few clusters, there was significantly increased connectivity in the gamma band, which was not the case in the HD-EEG analysis nor the virtual 19-channel set. Conversely, no effects were found for delta connectivity, as was the case for the HD-EEG analysis (compare Fig. 2A). The power analysis of the routine EEG data yielded similar results to those obtained using the 256-channel and 19-channel sets (compare Fig. 2B), except for the delta band, for which increased power patterns did not survive corrections for multiple comparisons.
Cohen d maps of vertex-based group comparisons (IGE > controls) for the HD-EEG study sample and the routine sample (canonical head models). Shown are standardized effect sizes for increased EEG levels in the patients with IGE compared with the controls. Note, only results for the theta frequency bands are presented and based on source-projected signals using canonical head models in the HD-EEG study sample (A and B) and the routine sample (C and D). All d-values were computed based on the respective t-values of the group factor of the linear models and corrected for the influence of age, sex and measurement site. d ≥ 0.8 indicates a large, d = 0.5 a medium and d = 0.2 a small effect.42 Sample sizes HD-EEG: nIGE = 35; ncontrols = 54; Routine EEG nIGE = 71; ncontrols = 43.
Discussion
This study provides a comprehensive overview of global and spatial differences in brain activity of individuals with IGE compared with controls depending on the number of EEG channels, type of head model and parcellation scheme. We systematically evaluated group-level outcomes and found that channel density had a stronger effect on EEG connectivity than on power, largely independent of the parcellation. The results for the head model type (individual versus canonical) were similar when conducted at brain vertices, but better concordance was achieved with a regional or network parcellation. In essence, the main electrophysiological signature of increased theta connectivity and power was similarly observed in the high-density and the clinical low-density setting, as validated in two different study samples.
Not many studies have evaluated the impact of spatial sampling on brain-wide approaches in generalized epilepsies before. Silva Alves and colleagues^43^ have found local topological network reorganizations using 256-channel HD-EEG that were still observable when using only a 25-electrode montage. This suggests that the most prominent effects in EEG signals of patients with IGE can be preserved with a few channels—in that study for ∼20 individuals per group and for effects in the delta, theta and beta bands in large cortical areas. Similarly, our analyses show that global differences between patients and controls, hence, the direction of the effect, can be detected with sparse arrays. Global differences were also the strongest in the delta, theta and beta bands, with Cohen’s d between ∼0.8 and 0.4 for connectivity across channel sets and for power across the spectrum with effect sizes between ∼1.2 and 0.75. These findings are largely in agreement with previous studies demonstrating markedly increased power in IGE^36,44^ and increased connectivity,^9-11,43^ with strong effects for the theta band. Moreover, we replicated these findings to some extent in a second sample of about 71 patients and 43 controls recorded in a clinical setting using the same methods and a routine, low-density EEG.
However, when it comes to the spatial topography of the found effects, a clear impact of the channel densities was observed. This was mainly the case for the connectivity estimates based on fewer than ∼64 channels. For signal power, reduced spatial coverage was less problematic, still yielding correlation coefficients of ∼0.3 for low-density maps (32-19 channels) with the original HD-EEG set. Crucially, while EEG power is less sensitive to noise and is usually estimated for a single location in the brain, the connectivity metric applied here reflects the overall synchronization between pair-wise signals across the brain. Hence, estimating complex markers such as connectivity is presumably more prone to spatial imprecisions. This aligns with earlier recommendations to use at least 64 channels when graph theoretical metrics are computed,^45^ which usually involves describing more long-distance network connections. Also, in the context of source localization in focal epilepsies, earlier studies suggested that at least 64 electrodes should be used to minimize the localization error.^3^ Here, spatial sensitivity is crucial, whereas, in the search for biomarkers using electrophysiology at the group level, large-scale network alterations are reported with high individual variability and spatial variations across IGE subtypes.^10,11^ Indeed, such brain-wide alterations were detected for most of the virtually reduced channel sets here, particularly in temporal and frontocentral areas, but with various effect sizes and more blurred in low-density maps.
Regarding our routine sample, the spatial topography for both the virtual and real 19-channel settings varied to some degree. Also, no group differences were observed for the delta frequency band. Several factors might have caused these differences: First, proportionally more patients with absence seizures (CAE, JAE) were included in the HD-EEG sample than in the clinical sample, and second, more GSWD occurred. Data trials containing GSWD (±10 s of data) were excluded for each individual, but it is conceivable that brain activity beyond this period is altered if GSWD are present.^17,19^ Further investigations with larger sample sizes are needed to disentangle network changes across IGE subtypes and other epilepsy syndromes and should address how group-level estimates can inform patient-tailored diagnostics in regular medical care. Third, the routine sample underwent a slightly different recording procedure than the HD-EEG sample (see Methods). Berger maneuvers and hyperventilation might have induced variation in the resting-state despite removing the respective data segments for analysis. Yet, our results suggest that data acquired in clinical settings with low-density and often less costly EEG systems can be informative and should be leveraged for a better understanding of network changes among epilepsies, but spatial precision should be regarded with caution.
Our work further contributes with an evaluation of the head model type and anatomical resolution. A remapping of EEG metrics according to regions or functional networks did not substantially yield more stable results for channel reductions. The only exception was the results for HD-EEG power in combination with a canonical head model. This can partially be explained given that a canonical head model is built on a template brain and not on individually defined tissue boundaries, leading to less precise source estimation.^21,22^ Averaging vertex-level data smooths local inaccuracies and leverages group effects. In a similar vein, statistical outcomes tended to be more similar between the head model types with coarser anatomical resolutions. Thus, for group-level studies focusing on large-scale network alterations, canonical head models combined with regional or network-level parcellations provide a reasonable and cost-effective alternative to individual MRI-based models. For vertex-level analyses or studies requiring high spatial precision (e.g. pre-surgical planning, localization of epileptogenic foci), individual head models remain essential as discussed above. Of note, we used boundary element methods in combination with a simplified three-layer head model. It is conceivable that other methods considering more complex brain geometries^46^ would generate larger differences between individual and canonical head models.
Finally, it needs to be said that we defined the original 256-channel montage as the gold standard for our analyses. There was a slight tendency of intermediate arrays (∼64-48 channels) resulting in stronger group differences, particularly for connectivity. It is possible that intermediate channel densities represent a better trade-off between spatial resolution and signal quality. If we had chosen a lower-density array (e.g. 128-64 channels) as the primary reference, this would likely have increased spatial correspondence among lower-density configurations. Theoretically, more channels should improve the analysis, but only if the signal-to-noise ratio is high.^47^ In other words, using high-density arrays in presence of a high noise level may blur the underlying true signal. At the same time, more channels yield a higher spatial overlap of leadfields for the source projection, which can lead to ill-defined covariance matrices and needs to be regulated. We kept a regularization of 5% for all channel densities, which should minimize this problem. Critically, the ground truth or the true network alterations in IGE are not known and only indirectly accessible, such that the specificity and validity of interictal resting-state markers require further investigations. This clearly differs from work on spatial sampling in the pre-surgical setting, for which the success of the diagnostic accuracy and intervention usually is followed up or can be modelled. Nevertheless, as noted above, this and previous studies consistently point to wide-spread alterations in the theta frequency range. Intriguingly, Stevelink et al. found genetic correlations for EEG levels in theta and genetic risk for IGE.^48^ Others suggest that 5–9 Hz oscillations represent periods of increased seizure susceptibility based on genetic rat models,^49^ further supporting a mechanistic role of theta-band synchrony in the maintenance or propagation of pathological network states in humans. Ji et al.^50^ demonstrated remarkable seizure frequency reduction in patients with IGE upon deep brain stimulation of the centromedian nucleus of the thalamus, which has been identified as a key node in an IGE network.
Conclusion
To conclude, main group-level effects in individuals with IGE can be reliably estimated using low-density EEG arrays and amplitude-based measures such as power. However, caution is needed when probing the spatial profile of connectivity estimates, as low-density EEG (<64 channels) captures global alterations well but lacks the spatial resolution required for detailed local or network-level analyses. While incorporating individual anatomical data is ideal, canonical head models can serve as a reasonable alternative, albeit with reduced spatial precision.
Supplementary Material
fcag022_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Seeck M, Koessler L, Bast T, et al The standardized EEG electrode array of the IFCN. Clinical Neurophysiology. 2017;128(10):2070–2077.28778476 10.1016/j.clinph.2017.06.254 · doi ↗ · pubmed ↗
- 2Lantz G, De Peralta RG, Spinelli L, Seeck M, Michel C. Epileptic source localization with high density EEG: How many electrodes are needed? Clinical Neurophysiology. 2003;114(1):63–69.12495765 10.1016/s 1388-2457(02)00337-1 · doi ↗ · pubmed ↗
- 3Song J, Davey C, Poulsen C, et al EEG source localization: Sensor density and head surface coverage. J Neurosci Methods. 2015;256:9–21.26300183 10.1016/j.jneumeth.2015.08.015 · doi ↗ · pubmed ↗
- 4Brodbeck V, Spinelli L, Lascano AM, et al Electroencephalographic source imaging: A prospective study of 152 operated epileptic patients. Brain. 2011;134(10):2887–2897.21975586 10.1093/brain/awr 243PMC 3187544 · doi ↗ · pubmed ↗
- 5Staljanssens W, Strobbe G, Holen RV, et al Seizure onset zone localization from ictal high-density EEG in refractory focal epilepsy. Brain Topogr. 2017;30:257–271.27853892 10.1007/s 10548-016-0537-8 · doi ↗ · pubmed ↗
- 6Stoyell SM, Wilmskoetter J, Dobrota M, et al High density EEG in current clinical practice and opportunities for the future. J Clin Neurophysiol. 2021;38(2):112–123.33661787 10.1097/WNP.0000000000000807 PMC 8083969 · doi ↗ · pubmed ↗
- 7Vorderwülbecke BJ, Baroumand AG, Spinelli L, Seeck M, van Mierlo P, Vulliémoz S. Automated interictal source localisation based on high-density EEG. Seizure. 2021;92:244–251.34626920 10.1016/j.seizure.2021.09.020 · doi ↗ · pubmed ↗
- 8Justesen AB, Foged MT, Fabricius M, et al Diagnostic yield of high-density versus low-density EEG: The effect of spatial sampling, timing and duration of recording. Clinical Neurophysiology. 2019;130(11):2060–2064.31541983 10.1016/j.clinph.2019.08.007 · doi ↗ · pubmed ↗