Morpho-phylogenetic evidence reveals five novel species of Pestalotiopsis (Sporocadaceae, Amphisphaeriales) from southern China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
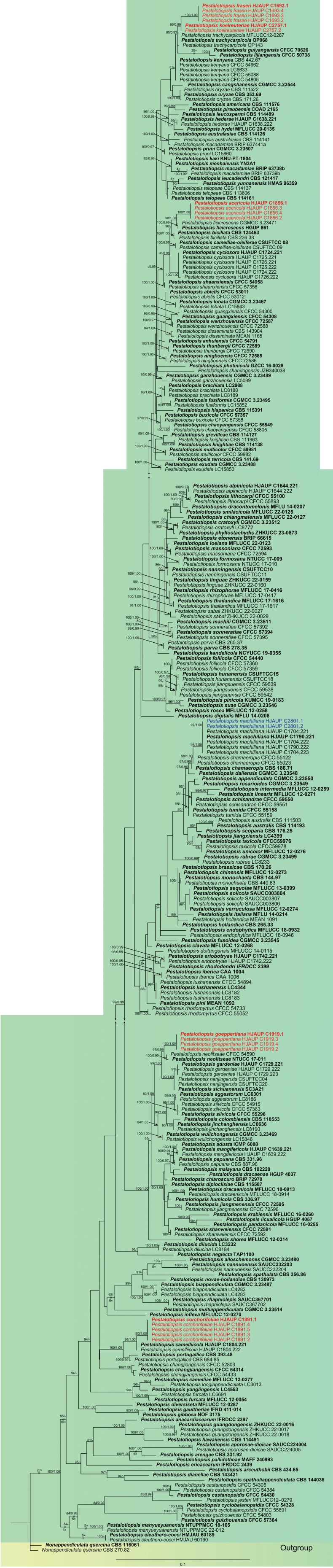 Figure 1
Figure 1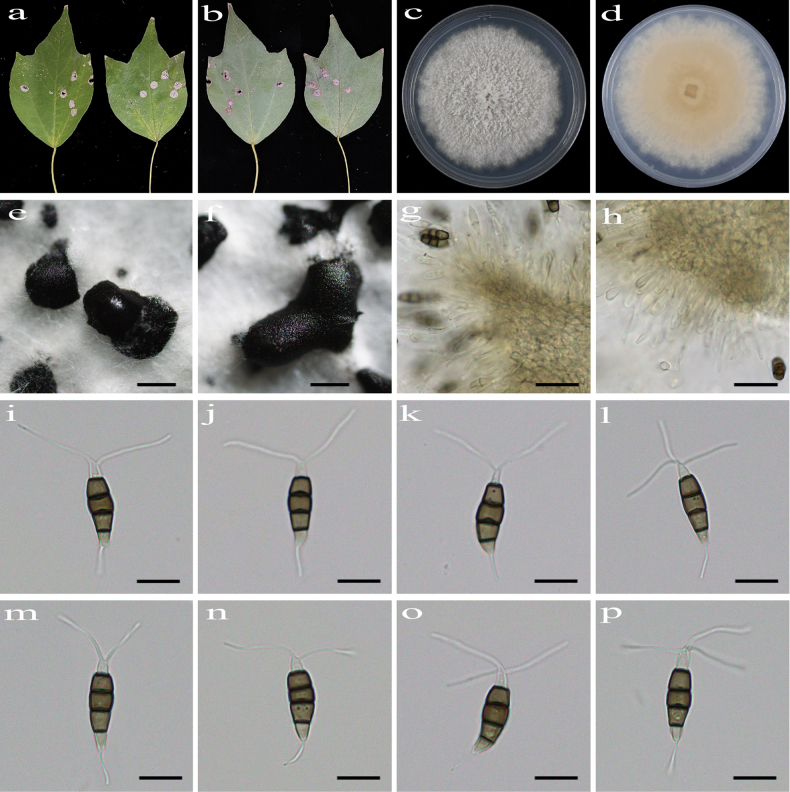 Figure 2
Figure 2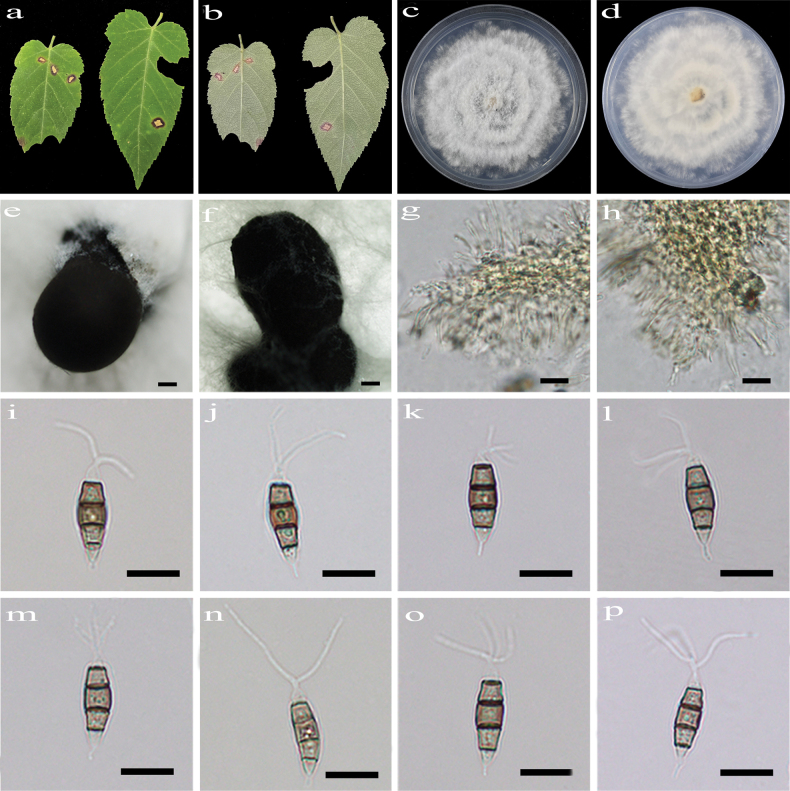 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5 Figure 6
Figure 6 Figure 7
Figure 7| Locus | Primers | PCR program | |
|---|---|---|---|
| Name | Sequence 5'–3' | ||
| ITS | ITS5 | GGAAGTAAAAGTCGTAACAAGG | 95 °C: 3 min, (95 °C: 15 s, 54 °C: 15 s, 72 °C: 30 s) × 35 cycles, 72 °C: 5 min |
| ITS4 | TCCTCCGCTTATTGATATGC | ||
| EF1-728F | CATCGAGAAGTTCGAGAAGG | 95 °C: 3 min, (95 °C: 15 s, 56 °C: 15 s, 72 °C: 30 s) × 35 cycles, 72 °C: 5 min | |
| EF1-986R | TACTTGAAGGAACCCTTACC | ||
| Bt2a | GGTAACCAAATCGGTGCTGCTTTC | 95 °C: 3 min, (95 °C: 15 s, 54 °C: 15 s, 72 °C: 30 s) × 35 cycles, 72 °C: 5 min | |
| Bt2b | ACCCTCAGTGTAGTGACCCTTGGC | ||
| Species | Strain number | Host/substrate | Locality | GenBank accession number | References | ||
|---|---|---|---|---|---|---|---|
| ITS | |||||||
|
| CFCC 53011T |
| China |
|
|
|
|
|
| CFCC 53012 |
| China |
|
|
| |
|
| ICMP 6088T | Refrigerator door | Fiji |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
|
|
|
| |
|
|
|
|
|
|
|
| |
|
| LC 6301T |
| China |
|
|
|
|
|
| LC 8186 |
| China |
|
|
| |
|
| CGMCC 3.23480T |
| China |
|
|
|
|
|
|
| China |
|
|
|
| |
|
|
| China |
|
|
| ||
|
| CBS 111576T | USA |
|
|
|
| |
|
| IFRDCC 2397T |
| China |
|
|
|
|
|
| CFCC 54791T |
| China |
|
|
|
|
|
| SAUCC224004T |
| China |
|
|
|
|
|
| SAUCC224005 |
| China |
|
|
| |
|
| CGMCC 3.23550T |
| China |
|
|
|
|
|
| CBS 434.65T |
| USA |
|
|
|
|
|
| CBS 331.92T |
| Singapore |
|
|
|
|
|
| CBS 114126T | New Zealand |
|
|
|
| |
|
| CBS 114141 | New South Wales |
|
|
| ||
|
| CBS 111503 | South Africa |
|
|
|
| |
|
| CBS 114193T | New South Wales |
|
|
| ||
|
| CGMCC 3.23487T | China |
|
|
|
| |
|
| LC4282 | China |
|
|
| ||
|
| LC4283 | China |
|
|
| ||
|
| CBS 124463T |
| Slovakia |
|
|
|
|
|
| CBS 236.38 | Italy |
|
|
| ||
|
| LC 2988T | China |
|
|
|
| |
|
| LC 8188 | China |
|
|
| ||
|
| LC 8189 | China |
|
|
| ||
|
| CBS 170.26T |
| New Zealand |
|
| – |
|
|
| CFCC 57357T |
| China |
|
|
|
|
|
| CFCC 57358 |
| China |
|
|
| |
|
|
| China |
|
|
|
| |
|
|
| China |
|
|
| ||
|
| MFLUCC 12–0277T |
| China |
|
|
|
|
|
| CSUFTCC 08T |
| China |
|
|
|
|
|
| CSUFTCC 09 |
| China |
|
|
| |
|
| CGMCC 3.23544T |
| China |
|
|
|
|
|
| CFCC 54430T |
| China |
|
|
|
|
|
| CFCC 54305 |
| China |
|
|
| |
|
| CFCC 54384 |
| China |
|
|
| |
|
| CBS 186.71T |
| Italy |
|
|
|
|
|
| CFCC 55122 |
| China |
|
|
|
|
|
| CFCC 55023 |
| China |
|
|
| |
|
| CFCC 54314T |
| China |
|
|
|
|
|
| CFCC 54433 |
| China |
|
|
| |
|
| CFCC 52803 |
| China |
|
|
| |
|
| CFCC 55549T |
| China |
|
|
|
|
|
| CFCC 58805 |
| China |
|
|
| |
|
| MFLUCC 22–0127T |
| Thailand |
|
|
|
|
|
| BRIP 72970T |
| Australia |
|
|
|
|
|
| MFLUCC 12–0273T | NA | China |
| – | – |
|
|
| MFLUCC 12–0268T | China |
|
|
|
| |
|
| CBS 118553T |
| Colombia |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
|
|
|
| |
|
|
|
|
|
|
|
| |
|
|
|
|
|
|
|
| |
|
| CGMCC 3.23512T |
| China |
|
|
|
|
|
| LC8772 |
| China |
|
|
| |
|
| CFCC 54328T |
| China |
|
|
|
|
|
| CFCC 55891 |
| China |
|
|
| |
|
|
| China |
|
|
|
| |
|
|
| China |
|
|
| ||
|
|
| China |
|
|
| ||
|
|
| China |
|
|
| ||
|
|
| China |
|
|
| ||
|
|
| China |
|
|
| ||
|
| CGMCC 3.23548T |
| China |
|
|
|
|
|
| CBS 143421T | Australia |
| – |
|
| |
|
| MFLU 14–0208T |
| New Zealand |
| – |
|
|
|
| LC3232T |
| China |
|
|
|
|
|
| LC8184 |
| China |
|
|
| |
|
| CBS 115587T |
| China |
|
|
|
|
|
| CBS 143904 |
| New Zealand |
|
|
|
|
|
| MEAN 1165 |
| Portugal |
|
|
|
|
|
| MFLUCC 12–0287T | China |
|
|
|
| |
|
| MFLUCC 14–0115 | Thailand |
|
|
|
| |
|
| HGUP 4037T |
| China | – |
|
|
|
|
| MFLUCC 18–0913T | Thailand |
|
|
|
| |
|
| MFLUCC 18–0914 | Thailand |
|
|
| ||
|
| MFLU 14–0207 |
| Thailand |
|
| – |
|
|
| HMJAU 60189T |
| China |
| – | – |
|
|
| HMJAU 60190 |
| China |
| – |
| |
|
| MFLUCC 18–0932T |
| Thailand |
|
| – |
|
|
| MFLUCC 18–0946 |
| Thailand |
|
| – | |
|
| IFRDCC 2439T |
| China |
|
|
|
|
|
|
| China |
|
|
|
| |
|
|
| China |
|
|
| ||
|
| BRIP 66615T |
| Australia |
|
|
|
|
|
| CGMCC 3.23488T |
| China |
|
|
|
|
|
|
|
| China |
|
|
| |
|
| HGUP 861T |
| China |
|
|
|
|
|
| CGMCC 3.23471 |
| China |
|
|
|
|
|
| CFCC 54440T |
| China |
|
|
|
|
|
| CFCC 57359 |
| China |
|
|
| |
|
| CFCC 57360 |
| China |
|
|
| |
|
| NTUCC 17–009T | China |
|
|
|
| |
|
| NTUCC 17–010 | China |
|
|
| ||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
|
|
|
| |
|
|
|
|
|
|
|
| |
|
| MFLUCC 12–0054T |
| Thailand |
|
|
|
|
|
| LC6691 |
| China |
|
|
|
|
|
| CGMCC 3.23495T | China |
|
|
|
| |
|
|
| China |
|
|
| ||
|
| CGMCC 3.23545T |
| China |
|
|
|
|
|
| CGMCC 3.23489T |
| China |
|
|
|
|
|
| LC5089 |
| China |
|
|
| |
|
|
| China |
|
|
|
| |
|
|
| China |
|
|
| ||
|
|
| China |
|
|
| ||
|
| IFRD 411–014T |
| China |
|
|
|
|
|
| NOF 3175T |
| Canada |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
|
|
|
| |
|
|
|
|
|
|
|
| |
|
| CBS 114127T | Australia |
|
|
|
| |
|
| ZHKUCC 22–0016T |
| China |
|
|
|
|
|
| ZHKUCC 22–0017 |
| China |
|
|
| |
|
| ZHKUCC 22–0018 |
| China |
|
|
| |
|
| CFCC 54308T |
| China |
|
|
|
|
|
| CFCC 54300 |
| China |
|
|
| |
|
| CFCC 70626T |
| China |
|
|
|
|
|
| CFCC 54803 |
| China |
|
|
|
|
|
| CFCC 57364T |
| China |
|
|
| |
|
| CBS 114491T | USA |
|
|
|
| |
|
|
| China |
|
|
|
| |
|
|
| China |
| – |
| ||
|
| CBS 115391T | Spain |
|
|
|
| |
|
| CBS 265.33T |
| Netherlands |
|
|
|
|
|
| MEAN 1091 |
| Portugal |
|
|
|
|
|
| CBS 336.97T | Soil | Papua New Guinea |
|
|
|
|
|
| CSUFTCC15T |
| China |
|
|
|
|
|
| CSUFTCC18 |
| China |
|
|
| |
|
| MFLUCC 20–0135T |
| Thailand |
|
|
|
|
|
| CAA 1004T |
| Spain |
|
|
|
|
|
| CAA 1006 |
| Spain |
|
|
| |
|
| MFLUCC 12–0270T | Unidentified tree | China |
|
|
|
|
|
| MFLUCC 12–0259T | Unidentified tree | China |
|
|
|
|
|
| MFLU 14–0214T |
| Italy |
|
|
|
|
|
| MFLUCC12–0279 |
| China |
|
|
|
|
|
| CFCC 72595T |
| China |
|
|
|
|
|
| CFCC 72596 |
| China |
|
|
| |
|
| CFCC 59538 |
| China |
|
|
|
|
|
| CFCC 59539 |
| China |
|
|
| |
|
| CFCC 59542 |
| China |
|
|
| |
|
| LC4399T | China |
|
|
|
| |
|
| LC6636T |
| China |
|
|
|
|
|
| LC8190 |
| China |
|
|
| |
|
| KNU–PT–1804T |
| Korea |
|
|
|
|
|
| NCYUCC 19–0355T |
| China |
|
|
|
|
|
| CBS 442.67T | Kenya |
|
|
|
| |
|
| LC6633 |
| China |
|
|
| |
|
| CFCC 54962 |
| China |
|
|
|
|
|
| CFCC 54805 |
| China |
|
|
| |
|
| CFCC 55088 |
| China |
|
|
| |
|
| CBS 111963 | New Zealand |
|
|
|
| |
|
| CBS 114138T | New Zealand |
|
|
| ||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| |
|
| MFLUCC 16–0260T | Thailand |
|
|
|
| |
|
| CBS 121417T | South Africa |
|
|
|
| |
|
| CBS 114489T |
| USA |
|
|
|
|
|
| HGUP 4057T |
| China |
|
|
|
|
|
| CFCC 50738T | China |
|
|
|
| |
|
| MFLUCC 12–0271T | China |
|
|
|
| |
|
| ZHKUCC 22–0159T |
| China |
|
|
|
|
|
| ZHKUCC 22–0160 |
| China |
|
|
| |
|
| CFCC 55100T |
| China |
|
|
|
|
|
| CFCC 55893 |
| China |
|
|
| |
|
| CGMCC 3.23467T |
| China |
|
|
|
|
|
|
|
| China |
|
|
| |
|
| MFLUCC 22–0123T | Dead leaves | Thailand |
|
|
|
|
|
| LC3013 |
| China |
|
|
|
|
|
| LC4344T | China |
|
|
|
| |
|
| LC8182 | China |
|
|
| ||
|
| LC8183 | China |
|
|
| ||
|
| CFCC 54894 |
| China |
|
|
|
|
|
| BRIP 63738bT |
| Australia |
|
|
|
|
|
| BRIP 63739b |
| Australia |
|
|
| |
|
| BRIP 637441a |
| Australia |
|
|
| |
|
| CGMCC 3.23511T | China |
|
|
|
| |
|
|
| China |
|
|
|
| |
|
|
| China |
|
|
| ||
|
|
| China |
|
|
| ||
|
|
| China |
|
|
| ||
|
|
| China |
|
|
| ||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| |
|
| CBS 102220T |
| Malaysia |
|
|
|
|
|
|
| China |
|
|
|
| |
|
|
| China |
|
|
| ||
|
| NTUPPMCC 18-165T | China |
|
|
|
| |
|
| NTUPPMCC 22-012 | China |
|
|
| ||
|
| CFCC 72593T |
| China |
|
|
|
|
|
| CFCC 72594 |
| China |
|
|
| |
|
| YN3A1T |
| China |
|
|
|
|
|
| CBS 144.97T |
| Netherlands |
|
|
|
|
|
| CBS 440.83 |
| Netherlands |
|
|
| |
|
| CGMCC 3.23514T | NA | China |
|
|
|
|
|
| CFCC59981T |
| China |
|
|
|
|
|
| CFCC59982 |
| China |
|
|
| |
|
| CSUFTCC20 |
| China |
|
|
|
|
|
| CSUFTCC04 |
| China |
|
|
| |
|
| CSUFTCC10T |
| China |
|
|
|
|
|
| CSUFTCC11 |
| China |
|
|
| |
|
| SAUCC232203T | Unknown host | China |
|
|
|
|
|
| SAUCC232204 | Unknown host | China |
|
|
| |
|
| TAP1100T |
| Japan |
|
|
|
|
|
| NTUCC 17–011T |
| China |
|
|
|
|
|
| CFCC 54590 |
| China |
|
|
|
|
|
| CFCC 72585T |
| China |
|
|
|
|
|
| CFCC 72586 |
| China |
|
|
| |
|
| CBS 130973T |
| Australia |
|
|
|
|
|
| CBS 111522 | USA |
|
|
|
| |
|
| CBS 171.26 | NA | Italy |
|
|
| |
|
| CBS 353.69T |
| Denmark |
|
|
| |
|
| MAFF 240993T |
| Japan |
|
|
|
|
|
| MFLUCC 16–0255T | Thailand |
|
|
|
| |
|
| CBS 331.96T | Coastal soil | Papua New Guinea |
|
|
|
|
|
| CBS 887.96 |
| Papua New Guinea |
|
|
| |
|
| CBS 265.37 |
| NA |
|
|
|
|
|
| CBS 278.35T |
| NA |
|
|
| |
|
| GZCC 16–0028T |
| China |
|
|
|
|
|
| ZHKUCC 23–0873T | NA | China |
|
|
|
|
|
| MEAN 1092T |
| Portugal |
|
|
|
|
|
| KUMCC 19–0183T |
| China |
|
|
|
|
|
| COAD 2165T |
| Brazil |
|
|
|
|
|
| CBS 393.48T | NA | Portugal |
|
|
|
|
|
| CBS 684.85 |
| New Zealand |
|
|
|
|
|
| CGMCC 3.23507T |
| China |
|
|
|
|
|
|
|
| China |
|
|
| |
|
| SAUCC367701T |
| China |
|
|
|
|
|
| SAUCC367702 |
| China |
|
|
| |
|
| MFLUCC 17–0416T |
| Thailand |
|
|
|
|
|
| MFLUCC 17–0417 |
| Thailand |
|
|
| |
|
| IFRDCC 2399T |
| China |
|
|
|
|
|
| CFCC 54733 |
| China |
|
|
|
|
|
| CFCC 55052 |
| China |
|
|
| |
|
| CGMCC 3.23549T |
| China |
|
|
|
|
|
| MFLUCC 12–0258T | China |
|
|
|
| |
|
| CGMCC 3.23499T |
| China |
|
|
|
|
|
| LC8233 |
| China |
|
|
| |
|
| ZHKUCC 22–0027 |
| China |
|
|
|
|
|
| ZHKUCC 22–0029 |
| China |
|
|
| |
|
| CFCC 59550T |
| China |
|
|
|
|
|
| CFCC 59551 |
| China |
|
|
| |
|
| CBS 176.25T | China |
|
|
|
| |
|
| MFLUCC 13–0399T |
| Italy |
| – | – |
|
|
| CFCC 54958T |
| China |
|
|
|
|
|
| CFCC 57356 |
| China |
|
|
| |
|
| CFCC 72591T |
| China |
|
|
|
|
|
| CFCC 72592 |
| China |
|
|
| |
| MFLUCC 12–0314T |
| Thailand |
|
|
|
| |
|
| SC3A21T |
| China |
|
|
|
|
|
| CFCC 55296T |
| China |
|
|
|
|
|
| CFCC 54915 |
| China |
|
|
| |
|
| CFCC 57363 |
| China |
|
|
| |
|
| MFLUCC 22–0125T | Thailand |
|
|
|
| |
|
| SAUCC003804T | Soil | China |
|
|
|
|
|
| SAUCC003806 | Soil | China |
|
|
| |
|
| SAUCC003807 | Soil | China |
|
|
| |
|
| CFCC 57392 |
| China |
|
|
|
|
|
| CFCC 57394T |
| China |
|
|
| |
|
| CFCC 57395 |
| China |
|
|
| |
|
| CBS 356.86T |
| Chile |
|
|
|
|
|
| CBS 144035T |
| Australia |
|
|
|
|
|
| CGMCC 3.23546T |
| China |
|
|
|
|
|
| CFCC59976T |
| China |
|
|
|
|
|
| CFCC59978 |
| China |
|
|
| |
|
| CBS 114137 | Australia |
|
|
|
| |
|
| CBS 114161T | Australia |
|
|
| ||
|
| CBS 113606 | Australia |
|
|
| ||
|
| CBS 141.69T | Soil | Pacific Islands |
|
|
|
|
|
| MFLUCC 17–1616T |
| Thailand |
|
|
|
|
|
| MFLUCC 17–1617 |
| Thailand |
|
|
| |
|
| CFCC 72589T |
| China |
|
|
|
|
|
| CFCC 72590 |
| China |
|
|
| |
|
| OP068T |
| China |
|
|
|
|
|
| OP143 |
| China |
|
|
| |
|
| MFLUCC 12-0267 | Unidentified tree | China |
|
|
|
|
|
| CFCC 55158T |
| China |
|
|
|
|
|
| CFCC 55159 |
| China |
|
|
| |
|
| MFLUCC 12–0276T | China |
| – |
|
| |
|
| MFLUCC 12–0274T | China |
|
| – | Maharachchikumbura et al. (2012) | |
|
| CFCC 72587T |
| China |
|
|
|
|
|
| CFCC 72588 |
| China |
|
|
| |
|
| CGMCC 3.23469T |
| China |
|
|
|
|
|
|
|
| China |
|
|
| |
|
| LC 4553T |
| China |
|
|
|
|
|
| HMAS 96359T |
| China |
| – | – |
|
|
| CBS 116061T |
| Italy |
|
|
|
|
|
| CBS 270.82 |
| Italy |
|
|
| |
- —National Natural Science Foundation of China 501100001809 https://ror.org/01h0zpd94 http://doi.org/10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Yeasts and Rust Fungi Studies · Plant Pathogens and Resistance
Introduction
Fungi represent one of the most diverse groups of organisms globally, playing critical roles in areas such as culinary value, ecological regulation, nutrient cycling, and innovative applications in biotechnology, medicine, and ecological conservation (Niego et al. 2023). Projections derived from host-association models estimate global fungal diversity at 2.2 to 3.8 million species, whereas high-throughput sequencing approaches yield substantially higher estimates in the range of 11.7 to 13.2 million species (Hawksworth and Lücking 2017; Hyde 2022). However, only around 200,000 fungal species have been discovered and formally documented in Fungal Names (FN, https://nmdc.cn/fungalnames/).
The genus Pestalotiopsis Steyaert was segregated from Pestalotia De Not. by Steyaert (1949) to accommodate those taxa with 5-celled conidia based on conidial forms, and Pestalotiopsis maculans (Corda) Nag Raj (≡ Sporocadus maculans Corda) was designated as the type species (Nag Raj 1985; Maharachchikumbura et al. 2014; Wijayawardene et al. 2017). Pestalotiopsis species are easily characterized by fusiform or subcylindrical, 5-celled conidia with three pigmented median cells and the presence of apical and basal appendages (Ariyawansa and Hyde 2018; Yin et al. 2024). Maharachchikumbura et al. (2014) further proposed two segregated anamorphic genera from Pestalotiopsis, namely Neopestalotiopsis Maharachch., K.D. Hyde & Crous and Pseudopestalotiopsis Maharachch., K.D. Hyde & Crous, based on combined datasets (ITS, tef1-α, and tub2) and the conidial pigmentation of the three median cells. As a result, Neopestalotiopsis differs from Pestalotiopsis and Pseudopestalotiopsis by its conidia with versicolorous median cells, and Pseudopestalotiopsis can be distinguished from Pestalotiopsis by sequence data and its conidia with dark-colored concolorous median cells. Pestalotiopsis is a species-rich group with varied habitats distributed in tropical and temperate regions (Bate-Smith and Metcalfe 1957; Luo et al. 2024). To date, about 457 epithets of Pestalotiopsis are recorded in Index Fungorum (Index Fungorum 2025). Members of the genus are common phytopathogens causing a variety of plant diseases, also occurring as endophytes and saprophytes isolated from leaf tissues, soil substrates, and rotted leaves, with some species further implicated in human and animal infections (Zhang et al. 2012a; Monden et al. 2013; Li et al. 2024; Luo et al. 2024; Yin et al. 2024). Previous research on Pestalotiopsis across a spectrum of environments has revealed a range of ecological functions. For example, functioning as saprophytes, some species accelerate material decomposition, whereas as endophytes, some species serve as sources of novel compounds with medicinal, agricultural, and industrial relevance (Xu et al. 2010, 2014).
Maharachchikumbura et al. (2014) revised the genus Pestalotiopsis based on combined ITS, tef1-α, and tub2 sequence data and morphological characteristics. These loci demonstrated strong phylogenetic resolution at the genus level and have been widely adopted in subsequent phylogenetic studies (Lin et al. 2023; Sun et al. 2023; Wang et al. 2025). In this study, samples of diseased leaves were collected from southern China. Accordingly, based on morphological observations and phylogenetic analyses (ITS, tef1-α, and tub2), 21 strains were assigned to Pestalotiopsis, including five novel species, namely P. acericola, P. corchorifolii, P. fraseri, P. goeppertiae, and P. koelreuteriae, and one known species, P. machiliana X.X. Luo & Jian Ma. Detailed illustrations and morphological descriptions of these taxa are provided below.
Materials and methods
Sample collection, fungal isolation, and morphological observation
Samples of diseased plant leaves were collected from botanical gardens and forest parks in southern China. The samples were placed in sealed bags with filter paper and transported to the laboratory. Fungi were isolated according to the tissue isolation method described by Gao et al. (2014). Leaf tissues were transferred onto potato dextrose agar (PDA; 200 g potato, 20 g glucose, 20 g agar, and 1000 mL water) plates and cultured at 25 °C under alternating light and dark conditions (8 h/16 h) for seven days, after which cultural characteristics, including color, shape, and size, were observed and recorded. The microscopic structures of isolated fungi were examined using an Olympus BX53 light microscope equipped with an Olympus DP27 digital camera (Olympus Optical Co., Ltd., Tokyo, Japan) to describe their morphological characteristics. All fungal strains were stored in 10% sterilized glycerin at 4 °C for further studies. The studied specimens and cultures were deposited in the Herbarium of Jiangxi Agricultural University, Plant Pathology, Nanchang, China (HJAUP).
DNA extraction, PCR amplification, and sequencing
After fungal cultures were grown at 25 °C under a light–dark cycle (8 h/16 h) for 14 days, fresh mycelia were scraped with a sterile needle and transferred into 2 mL centrifuge tubes for grinding with liquid nitrogen. Genomic DNA was extracted using the Solarbio Fungal Genomic DNA Extraction Kit (Beijing Solarbio Science & Technology Co., Ltd., Beijing, China) according to the manufacturer’s protocol. DNA was amplified in a 25 μL PCR system consisting of 12.5 µL of 2 × Power Taq PCR MasterMix, 9.5 µL of double-distilled water (ddH_2_O), 1 µL of DNA template, and 1 µL each of forward and reverse primers, targeting three loci: ITS, tef1-α, and tub2. The primer pairs and PCR programs are listed in Table 1. PCR amplification products were stained with ethidium bromide and examined by electrophoresis in 1% agarose gels. Purification and sequencing of the PCR products were performed by Hunan Youkanglai Biotechnology Co., Ltd. The obtained sequences were submitted to the GenBank database (www.ncbi.nlm.nih.gov, accessed on 1 December 2025; Table 2).
Phylogenetic analyses
According to recent publications (Hsu et al. 2024; Li et al. 2024; Luo et al. 2024; Zhao et al. 2024; Cao et al. 2025; Jiang et al. 2025; Wang et al. 2025), sequences of Pestalotiopsis species obtained from GenBank were selected for phylogenetic analyses together with the 21 strains in this study (Table 2). Nonappendiculata quercina (CBS 116061 and CBS 270.82) was used as the outgroup. All sequences were aligned using MAFFT v.7.526 (Katoh et al. 2019, https://mafft.cbrc.jp/alignment/server/). A combined multilocus sequence dataset comprising ITS, tef1-α, and tub2 was assembled and manually optimized to improve alignment and sequence accuracy using PhyloSuite v.1.2.2 (Zhang et al. 2020). Phylogenetic trees were constructed based on the concatenated dataset using maximum likelihood (ML) and Bayesian inference (BI) in PhyloSuite v.1.2.2 (Zhang et al. 2020). The optimal partition model (edge-linked) was selected using the ModelFinder function (Kalyaanamoorthy et al. 2017), with the BIC criterion applied for IQ-TREE construction and the AICc criterion implemented for MrBayes analysis. Maximum likelihood phylogenies were constructed using IQ-TREE (Nguyen et al. 2015) under an edge-linked partition model, with nodal support assessed through 10,000 ultrafast bootstrap replicates (Minh et al. 2013). The final phylogenetic tree was selected from suboptimal trees of each run through likelihood score comparison, with the following models applied to respective gene partitions: HKY+F+R5 for ITS, TIM2e+R4 for tef1-α, and TPM2u+F+I+G4 for tub2. Bayesian inference phylogenies were constructed using MrBayes v.3.2.6 (Ronquist et al. 2012) under a partition model configuration, with two parallel runs and 2,000,000 generations, while discarding the initial 25% of sampled data as burn-in. The best-fit models were HKY+F+I+G4 for ITS, GTR+F+G4 for tef1-α, and GTR+F+I+G4 for tub2. Phylogenetic trees were visualized using FigTree v.1.4.4 (http://tree.bio.ed.ac.uk/software/figtree), with subsequent editing and layout finalized in Adobe Illustrator CS v.5.
Results
Molecular phylogeny
Based on the concatenated sequences of the three barcode genes (ITS, tef1-α, and tub2), the phylogenetic tree (Fig. 1) was constructed using maximum likelihood (ML) and Bayesian inference (BI) to analyze the phylogenetic relationships of the 21 strains within the genus Pestalotiopsis. The phylogenetic analyses included 301 strains (Table 1), comprising 159 accepted Pestalotiopsis species and 21 strains from this study, with Nonappendiculata quercina (CBS 116061 and CBS 270.82) serving as the outgroup. The combined sequence consisted of 1472 nucleotide positions (ITS: 1–556, tef1-α: 557–1013, tub2: 1014–1472), comprising 901 distinct patterns, 662 parsimony-informative sites, 163 singleton sites, and 647 constant sites. Phylogenetic trees were constructed based on the concatenated sequences using ML and BI methods with essentially identical topological architectures. The optimal ML phylogenetic tree (lnL = −19450.797) is shown in Fig. 1, with node support values annotated adjacent to each branch. At each node, the first and second numerical descriptors correspond to ultrafast bootstrap support values from ML and posterior probabilities obtained from BI, respectively.
A maximum-likelihood phylogenetic tree of Pestalotiopsis constructed based on concatenated sequence data of ITS, tef1-α, and tub2. Bootstrap support values for ML greater than 80% and Bayesian posterior probabilities greater than 0.95 are shown near the nodes. The tree is rooted with Nonappendiculata quercina (CBS 116061 and CBS 270.82). New strains identified in this study are shown in blue; new species are shown in red. Some branches were shortened according to the indicated multipliers, and these are indicated by the symbol (//).
Taxonomy
Pestalotiopsis
acericola
Taxon classificationFungiAmphisphaerialesPestalotiopsidaceae
M.G. Liao & Jian Ma sp. nov.
7D2EAFC3-3BC4-575C-A1F2-7CE493E1F903
Index Fungorum: IF904707
Type.
China • Zhejiang Province, Hangzhou City, West Lake Scenic Area, on diseased leaves of Acer buergerianum, 22 October 2023, X.X. Luo (HJAUP M1856, holotype), ex-type living culture, HJAUP C1856.1 = HJAUP C1856.2 = HJAUP C1856.3 = HJAUP C1856.4.
Etymology.
Named after the genus, Acer, from which the fungus was isolated.
Description.
Regular leaf spots, grey white in center with blurred margin. Asexual morph on PDA: Conidiomata acervular, globose to clavate, 350–660 μm diam., superficial, solitary or aggregated in clusters, black. Conidiophores indistinct and reduced to conidiogenous cells. Conidiogenous cells hyaline, smooth, cylindrical to ampulliform. Conidia fusiform, straight or slightly curved, 16.1–24.5 × 5.2–6.9 μm (x̄ = 20.9 × 5.9 μm, n = 50), 4-septate, slightly constricted at the septa; basal cell conical, 3.1–5.3 μm (x̄ = 4.1 μm), hyaline or sometimes pale brown, smooth, thin-walled, with a single filiform appendage, unbranched, 4.6–10.7 μm (x̄ = 7.0 μm) long; three median cells doliiform to cylindrical, smooth, thick-walled, 10.4–14.6 μm (x̄ = 12.8 μm), concolorous, pale brown to brown, somewhat constricted at the septa, second cell from the base 3.6–5.0 µm (x̄ = 4.4 μm) long, third cell 3.0–5.0 µm (x̄ = 4.1 μm) long, fourth cell 3.6–5.8 µm (x̄ = 4.6 μm) long); apical cell conical to acute, hyaline, smooth, thin-walled, 2.6–4.6 µm (x̄ = 3.9 μm) long, with 2–3 filiform appendages, arising from the apex of the apical cell each at a different point, unbranched, 12.3–27.5 µm (x̄ = 17.7 μm) long. Sexual morph: not observed.
Pestalotiopsis acericola (HJAUP C1856.1, ex-type). a, b. Leaf of host plant (front and reverse); c, d. Culture on PDA (front and reverse); e, f. Conidiomata; g, h. Conidiogenous cells and conidia; i–p. Conidia. Scale bars: 400 µm (e, f); 20 µm (g, h); 10 µm (i–p).
Culture characteristics.
Colonies on PDA grow fast, filamentous, reaching 76–82 mm diam. after 5–7 days at 25 °C in light and dark conditions (8 h/16 h), white, with flocculent mycelium and entire edge, forming black, brown conidiomata, and reverse pale orange in center.
Note.
Phylogenetic analyses revealed that the four strains (HJAUP C1856.1^T^, HJAUP C1856.2, HJAUP C1856.3, and HJAUP C1856.4) of P. acericola form a distinct clade sister to P. ficicrescens (HGUP 861^T^ and CGMCC 3.23471) with 98% ML/1.00 BI bootstrap support (Fig. 1). Based on a BLASTn search of P. acericola (HJAUP C1856.1^T^) and P. ficicrescens (HGUP 861^T^) in GenBank, comparisons of nucleotides showed 2 bp (537/539, including two gaps), 1 bp (291/292, no gaps), and 4 bp (451/455, including one gap) nucleotide differences in the ITS, tef1-α, and tub2 regions, respectively. Moreover, P. acericola differs from P. ficicrescens Qi Yang & Yong Wang bis (Hyde et al. 2023) in its smaller conidiomata (350–660 μm diam. vs. 400–1200 μm diam.) and wider conidia (5.2–6.9 μm vs. 3–5.5 μm) with a longer fourth cell (3.6–5.8 µm vs. 3–5 µm) from the base, a longer basal appendage (4.6–10.7 μm vs. 3.5–7 µm), and longer apical appendages (12.3–27.5 μm vs. 10.5–18 µm). Furthermore, P. acericola can be distinguished from P. ficicrescens by its different host (Acer buergerianum vs. Ficus tikoua).
Pestalotiopsis
corchorifolii
Taxon classificationFungiAmphisphaerialesPestalotiopsidaceae
M.G. Liao & Jian Ma sp. nov.
AA172086-4AAB-5E13-A437-236403FC8465
861554
Type.
China • Zhejiang Province, Wenzhou City, Wenzhou Botanical Garden, on diseased leaves of Rubus corchorifolius, 24 October 2023, X.X. Luo (HJAUP M1891, holotype), ex-type living culture HJAUP C1891.1 = HJAUP C1891.2 = HJAUP C1891.3 = HJAUP C1891.4 = HJAUP M1891.5.
Etymology.
Named after the host species, Rubus corchorifolius, from which the fungus was isolated.
Description.
Regular leaf spots, grey white in center with brown at margin. Asexual morph on PDA: Conidiomata acervular, globose to clavate, 849–1375 μm diam., solitary or aggregated in clusters, black. Conidiophores indistinct and reduced to conidiogenous cells. Conidiogenous cells hyaline, smooth, cylindrical to ampulliform. Conidia fusiform, straight or slightly curved, 14.6–21.6 × 4.4–5.9 μm (x̄ = 17.4 × 5.3 μm, n = 50), 4-septate, slightly constricted at the septa; basal cell conical, 2.2–3.9 μm (x̄ = 2.6 μm), hyaline, smooth, thin-walled, with a single filiform appendage, unbranched, 0.9–3.1 μm (x̄ = 2.1 μm) long; three median cells doliiform to cylindrical, smooth, thick-walled, 10.5–14.0 μm (x̄ = 12.1 μm), concolorous, brown, somewhat constricted at the septa, second cell from the base 3.4–5.6 µm (x̄ = 4.1 μm) long, third cell 3.3–4.6 µm (x̄ = 4.1 μm) long, fourth cell 3.4–4.7 µm (x̄ = 4.0 μm) long); apical cell conical to acute, hyaline, smooth, thin-walled, 1.9–3.7 µm (x̄ = 2.7 μm) long, with 2–3 filiform appendages, arising from the apex of the apical cell each at a different point, unbranched, 6.7–17.4 µm (x̄ = 11.5 μm) long. Sexual morph: not observed.
Pestalotiopsis corchorifolii (HJAUP C1891.1, ex-type). a, b. Leaf of host plant (front and reverse); c, d. Culture on PDA (front and reverse); e, f. Conidiomata; g, h. Conidiogenous cells and conidia; i–p. Conidia. Scale bars: 200 µm (e, f); 10 µm (g–p).
Culture characteristics.
Colonies on PDA grow fast, filamentous, reaching 85–90 mm diam. after 5–7 days at 25 °C in light and dark conditions (8 h/16 h), white, with flocculent mycelium and entire edge, forming black conidiomata, and reverse white.
Note.
Phylogenetic analyses revealed that the five strains (HJAUP C1891.1^T^, HJAUP C1891.2, HJAUP C1891.3, HJAUP C1891.4, and HJAUP M1891.5) of P. corchorifolii form a distinct clade sister to P. camelliicola (HJAUP C1804.221^T^ and HJAUP C1804.222) with 98% ML/1.00 BI bootstrap support (Fig. 1). Based on a BLASTn search of P. corchorifolii (HJAUP C1891.1^T^) and P. camelliicola (HJAUP C1804.221^T^) in GenBank, comparisons of nucleotides showed 1 bp (630/631, including one gap), 5 bp (296/301, including three gaps), and 5 bp (476/481, including one gap) nucleotide differences in the ITS, tef1-α, and tub2 regions, respectively. Moreover, P. corchorifolii can be distinguished from P. camelliicola X.X. Luo & Jian Ma (Luo et al. 2024) by its larger conidiomata (849–1375 μm diam. vs. 470–1320 μm diam.) and smooth, narrower conidia (4.4–5.9 μm vs. 5.7–7.6 μm), and further differs in that the conidia of P. camelliicola have 2–4 tubular apical appendages arising from an apical crest or irregularly branched along their length, resulting in 2–3 branches. Furthermore, P. corchorifolii can be distinguished from P. camelliicola by its different host (Rubus corchorifolius vs. Camellia japonica).
Pestalotiopsis
fraseri
Taxon classificationFungiAmphisphaerialesPestalotiopsidaceae
M.G. Liao & Jian Ma sp. nov.
A3BC8BEE-4D82-5832-AE7F-6DD8C75996A6
861555
Type.
China • Jiangxi Province, Nanchang City, Jiangxi Agricultural University, on diseased leaves of Photinia × fraseri Dress, 26 October 2023, X.X. Luo (HJAUP M1693, holotype), ex-type living culture HJAUP C1693.1 = HJAUP C1693.2 = HJAUP C1693.3 = HJAUP C1693.4.
Etymology.
Named after the host species, Photinia × fraseri, from which the fungus was isolated.
Description.
Regular leaf spots, pale brown in center with brown at margin. Asexual morph on PDA: Conidiomata acervular, globose to clavate, 426–786 μm diam., solitary, black. Conidiophores indistinct and reduced to conidiogenous cells. Conidiogenous cells hyaline, smooth, cylindrical to ampulliform. Conidia fusiform, straight or slightly curved, 19.3–26.4 × 5.2–6.3 μm (x̄ = 23.0 × 5.7 μm, n = 50), 4-septate, slightly constricted at the septa; basal cell conical, 3.4–5.3 μm (x̄ = 4.4 μm), hyaline, smooth, thin-walled, with a single filiform appendage, unbranched, 4.6–10.5 μm (x̄ = 7.9 μm) long; three median cells doliiform to cylindrical, smooth, thick-walled, 12.2–16.2 μm (x̄ = 14.4 μm), concolorous, brown, somewhat constricted at the septa, second cell from the base 3.9–6.1 µm (x̄ = 4.8 μm) long, third cell 4.0–5.4 µm (x̄ = 4.7 μm) long, fourth cell 4.0–5.2 µm (x̄ = 4.7 μm) long); apical cell conical to acute, hyaline, smooth, thin-walled, 3.7–5.0 µm (x̄ = 4.2 μm) long, with 2–3 filiform appendages, arising from the apex of the apical cell each at a different point, unbranched, 12.3–20.8 µm (x̄ = 16.3 μm) long. Sexual morph: not observed.
Pestalotiopsis fraseri (HJAUP C1693.1, ex-type). a, b. Leaf of host plant (front and reverse); c, d. Culture on PDA (front and reverse); e, f. Conidiomata; g–i. Conidiogenous cells and conidia; j–q. Conidia. Scale bars: 200 µm (e, f); 10 µm (g–q).
Culture characteristics.
Colonies on PDA grow fast, filamentous, reaching 85–90 mm diam. after 5–7 days at 25 °C in light and dark conditions (8 h/16 h), white, with flocculent mycelium and entire edge, forming black conidiomata, and reverse white.
Note.
Phylogenetic analyses revealed that the four strains (HJAUP C1693.1^T^, HJAUP C1693.2, HJAUP C1693.3, and HJAUP C1693.4) of P. fraseri form a distinct clade sister to P. koelreuteriae (HJAUP C2757.1^T^ and HJAUP C2757.2) with 100% ML/1.00 BI bootstrap support (Fig. 1). Based on a BLASTn search of P. fraseri (HJAUP C1693.1^T^) and P. koelreuteriae (HJAUP C2757.1^T^) in GenBank, comparisons of nucleotides showed 13 bp (619/632, including three gaps), 1 bp (296/297, no gaps), and 3 bp (479/482, including one gap) nucleotide differences in the ITS, tef1-α, and tub2 regions, respectively. Based on a BLASTn search of P. fraseri (HJAUP C1693.1^T^) and P. trachycarpicola (OP068^T^) in GenBank, comparisons of nucleotides showed 10 bp (530/540, including one gap) and 1 bp (294/295, no gaps) in the ITS and tef1-α regions, respectively. Moreover, P. fraseri differs from P. koelreuteriae (this study) in its conidia with longer basal appendages (4.6–10.5 μm vs. 3.4–6.7 μm) and longer apical appendages (12.3–20.8 µm vs. 8.1–18.5 µm), and from P. trachycarpicola Yan M. Zhang & K.D. Hyde (Zhang et al. 2012b) in its longer basal appendages (4.6–10.5 μm vs. 2.7–5.5 μm). Furthermore, P. fraseri can be distinguished from P. koelreuteriae and P. trachycarpicola by its different host (Photinia × fraseri vs. Koelreuteria paniculata vs. Trachycarpus fortunei).
Pestalotiopsis
goeppertiae
Taxon classificationFungiAmphisphaerialesPestalotiopsidaceae
M.G. Liao & Jian Ma sp. nov.
CF296B09-00B2-5B7A-BE1A-CE5610216FC1
861556
Type.
China • Fujian Province, Fuzhou City, Fuzhou Botanical Garden, on diseased leaves of Goeppertia makoyana, 24 October 2023, X.X. Luo (HJAUP M1919, holotype), ex-type living culture HJAUP C1919.1 = HJAUP C1919.2 = HJAUP C1919.3 = HJAUP C1919.4.
Etymology.
Named after the genus, Goeppertia, from which the fungus was isolated.
Description.
Regular leaf spots, grey white in center with brown at margin. Asexual morph on PDA: Conidiomata acervular, globose to clavate, 550–1520 μm diam., superficial, solitary or aggregated in clusters, dark brown. Conidiophores indistinct and reduced to conidiogenous cells. Conidiogenous cells hyaline, smooth, cylindrical to ampulliform. Conidia fusiform, straight or slightly curved, 17.0–23.6 × 5.0–5.9 μm (x̄ = 19.5 × 5.5 μm, n = 50), 4-septate, slightly constricted at the septa; basal cell conical, 2.8–4.5 μm (x̄ = 3.5 μm), hyaline or sometimes pale brown, smooth, thin-walled, with a single filiform appendage, unbranched, 2.3–6.0 μm (x̄ = 3.5 μm) long; three median cells doliiform to cylindrical, smooth, thick-walled, 11.0–14.5 μm (x̄ = 12.4 μm), concolorous, pale brown to brown, somewhat constricted at the septa, second cell from the base 3.4–5.1 µm (x̄ = 4.2 μm) long, third cell 3.7–4.6 µm (x̄ = 4.2 μm) long, fourth cell 3.7–4.9 µm (x̄ = 4.2 μm) long); apical cell conical to acute, hyaline, smooth, thin-walled, 3.2–4.1 µm (x̄ = 3.7 μm) long, with 2–3 filiform appendages, arising from the apex of the apical cell each at a different point, unbranched, 12.6–21.3 µm (x̄ = 15.4 μm) long. Sexual morph: not observed.
Pestalotiopsis goeppertiae (HJAUP C1919.1, ex-type). a, b. Leaf of host plant (front and reverse); c, d. Culture on PDA (front and reverse); e, f. Conidiomata; g–i. Conidiogenous cells and conidia; j–q. Conidia. Scale bars: 200 µm (e, f); 20 µm (g–i); 10 µm (j–q).
Culture characteristics.
Colonies on PDA grow fast, filamentous, reaching 85–90 mm diam. after 5–7 days at 25 °C in light and dark conditions (8 h/16 h), white, with flocculent mycelium and entire edge, forming black, brown conidiomata, and reverse pale orange.
Note.
Phylogenetic analyses revealed that the four strains (HJAUP C1919.1^T^, HJAUP C1919.2, HJAUP C1919.3, and HJAUP C1919.4) of P. goeppertiae form a distinct clade sister to P. neolitseae (NTUCC 17–011^T^ and CFCC 54590) with 97% ML/0.96 BI bootstrap support (Fig. 1). Based on a BLASTn search of P. goeppertiae (HJAUP C1919.1^T^) and P. neolitseae (NTUCC 17–011^T^) in GenBank, comparisons of nucleotides showed 9 bp (248/257, including four gaps) and 1 bp (399/400, no gaps) nucleotide differences in the tef1-α and tub2 regions, respectively. Moreover, P. goeppertiae differs from P. neolitseae H.A. Ariy. & K.D. Hyde (Ariyawansa and Hyde 2018) in its conidia with a shorter fourth cell (3.7–4.9 µm vs. 4–5(–6) µm) from the base and longer apical appendages (12.6–21.3 µm vs. (7–)10–15(–17) μm). Furthermore, P. goeppertiae can be distinguished from P. neolitseae by its different host (Goeppertia makoyana vs. Neolitsea villosa).
Pestalotiopsis
koelreuteriae
Taxon classificationFungiAmphisphaerialesPestalotiopsidaceae
M.G. Liao & Jian Ma sp. nov.
DF7962FF-410E-5E7A-9B5D-2F6BF761096F
861557
Type.
China • Hunan Province, Zhangjiajie City, Zhangjiajie National Forest Park, on diseased leaves of Koelreuteria paniculata, 18 October 2024, M.G. Liao (HJAUP M2757, holotype), ex-type living culture HJAUP C2757.1 = HJAUP C2757.2.
Etymology.
Named after the genus, Koelreuteria, from which the fungus was isolated.
Description.
Irregular leaf spots, pale brown in center with brown at margin. Asexual morph on PDA: Conidiomata acervular, globose to clavate, 610–750 μm diam., solitary or aggregated in clusters, black. Conidiophores indistinct and reduced to conidiogenous cells. Conidiogenous cells hyaline, smooth, cylindrical to ampulliform. Conidia fusiform, straight or slightly curved, 18.3–26.9 × 4.1–6.0 μm (x̄ = 22.7 × 5.3 μm, n = 50), 4-septate, slightly constricted at the septa; basal cell conical, 2.7–4.6 μm (x̄ = 3.9 μm), hyaline or sometimes pale brown, smooth, thin-walled, with a single filiform appendage, unbranched, 3.4–6.7 μm (x̄ = 4.9 μm) long; three median cells doliiform to cylindrical, smooth, thick-walled, 12.8–17.3 μm (x̄ = 15.2 μm), concolorous, brown, somewhat constricted at the septa, second cell from the base 3.4–6.6 µm (x̄ = 4.8 μm) long, third cell 3.4–6.5 µm (x̄ = 5.3 μm) long, fourth cell 3.8–6.6 µm (x̄ = 5.2 μm) long); apical cell conical to acute, hyaline, smooth, thin-walled, 2.5–5.1 µm (x̄ = 3.5 μm) long, with 1–3 (mostly 2) filiform appendages, arising from the apex of the apical cell each at a different point, unbranched, 8.1–18.5 µm (x̄ = 13.2 μm) long. Sexual morph: not observed.
Pestalotiopsis koelreuteriae (HJAUP C2757.1, ex-type). a, b. Leaf of host plant (front and reverse); c, d. Culture on PDA (front and reverse); e. Conidiomata; f. Conidiogenous cells and conidia; g–p. Conidia. Scale bars: 200 µm (e); 10 µm (f–p).
Culture characteristics.
Colonies on PDA grow fast, filamentous, reaching 50–60 mm diam. after 5–7 days at 25 °C in light and dark conditions (8 h/16 h), white, with flocculent mycelium and entire edge, forming black conidiomata, and reverse pale orange.
Note.
Phylogenetic analyses revealed that the two strains (HJAUP C2757.1^T^ and HJAUP C2757.2) of P. koelreuteriae form a distinct clade sister to P. fraseri (HJAUP C1693.1^T^, HJAUP C1693.2, HJAUP C1693.3, and HJAUP C1693.4) with 100% ML/1.00 BI bootstrap support (Fig. 1). The nucleotide divergences and morphological differentiations between P. koelreuteriae and P. fraseri have been delineated in the preceding section. Comparisons of nucleotides showed 13 bp (2.1%, including three gaps) in the ITS region. Based on the criteria established by Jeewon and Hyde (2016), a divergence of more than 1.5% in the ITS nucleotide sequence can support the recognition of a novel species. Thus, P. koelreuteriae was proposed as a new species.
Pestalotiopsis
machiliana
Taxon classificationFungiAmphisphaerialesPestalotiopsidaceae
X.X. Luo & Jian Ma, Mycokeys 109: 229 (2024)
DC673386-6F04-529D-AE51-5556306EEC4B
Index Fungorum: IF902325
Type.
China • Hubei Province, Wuhan City, Wuhan Botanical Garden, on diseased leaves of Bambusa multiplex (Lour.) Raeusch. ex Schult. cv. Fernleaf R. A. Young, 24 October 2024, M.G. Liao (HJAUP M2801, living culture HJAUP C2801.1 = HJAUP C2801.2).
Description.
Leaf tip lesions, paler brown spots with brown margin between diseased and healthy tissue. Asexual morph on PDA: Conidiomata acervular, globose to clavate, 1000–1350 μm diam., solitary or aggregated in clusters, black. Conidiophores indistinct and reduced to conidiogenous cells. Conidiogenous cells hyaline, smooth, cylindrical to ampulliform. Conidia fusiform, straight or slightly curved, 18.1–25.1 × 4.9–6.8 μm (x̄ = 21.3 × 5.8 μm, n = 50), 4-septate, slightly constricted at the septa; basal cell conical, 2.9–5.3 μm (x̄ = 3.9 μm), hyaline or sometimes pale brown, smooth, thin-walled, with a single filiform appendage, unbranched, 4.4–7.1 μm (x̄ = 5.9 μm) long; three median cells doliiform to cylindrical, smooth, thick-walled, 11.2–17.3 μm (x̄ = 13.9 μm), concolorous, brown, somewhat constricted at the septa, second cell from the base 3.5–5.7 µm (x̄ = 4.6 μm) long, third cell 3.9–5.9 µm (x̄ = 4.7 μm) long, fourth cell 3.8–5.7 µm (x̄ = 4.6 μm) long); apical cell conical to acute, hyaline, smooth, thin-walled, 2.3–4.8 µm (x̄ = 3.5 μm) long, with 2–3 filiform appendages, arising from the apex of the apical cell each at a different point, unbranched, 9.3–20.8 µm (x̄ = 14.9 μm) long. Sexual morph: not observed.
Pestalotiopsis machiliana (HJAUP C2801.1). a, b. Leaf of host plant (front and reverse); c, d. Culture on PDA (front and reverse); e–f. Conidiomata; g. Conidiogenous cells and conidia; h–o. Conidia. Scale bars: 200 µm (e, f); 10 µm (g–o).
Culture characteristics.
Colonies on PDA grow fast, filamentous, reaching 55–70 mm diam. after 5–7 days at 25 °C in light and dark conditions (8 h /16 h), white, with flocculent mycelium and entire edge, forming black conidiomata, and reverse pale orange.
Notes.
Pestalotiopsis machiliana was introduced by Luo et al. (2024) and isolated from diseased leaves of Machilus pauhoi in Jiangxi Province, China. Phylogenetic analyses showed that our new isolates (HJAUP C2801.1 and HJAUP C2801.2) cluster with the group of P. machiliana (HJAUP C1704.221, HJAUP C1704.222, HJAUP C1704.223, HJAUP C1790.221^T^, and HJAUP C1790.222) with 99% ML/1.00 BI bootstrap support (Fig. 1). A BLASTn search of GenBank revealed that the sequences of our new isolate (HJAUP C2801.1) and P. machiliana (HJAUP C1790.221^T^) share 99% similarity (630/632, including one gap) in ITS, 100% similarity (272/272, no gaps) in tef1-α, and 99% similarity (481/482, including one gap) in tub2. Morphologically, our isolate aligns well with the original description of P. machiliana (Luo et al. 2024). Thus, based on the high morphological similarity and only minor molecular differences, we identified our new isolate as P. machiliana.
Discussion
Morphological characteristics have consistently served as a fundamental criterion for the identification of Pestalotiopsis species. However, with the continuous increase in the number of Pestalotiopsis species, the criteria used for species identification have become increasingly ambiguous (Huanaluek et al. 2021). Maharachchikumbura et al. (2014) previously suggested that continued reliance on conidial size for species delineation within Pestalotiopsis is methodologically unsound. In this study, no significant differences were observed in conidial size between P. acericola and P. chiangmaiensis Y.R. Sun & Yong Wang bis (16.1–24.5 × 5.2–6.9 µm vs. 16–26 × 4–7 µm; Sun et al. 2023), but the two species were clustered into different branches, and the nucleotide differences between P. acericola (HJAUP C1856.1^T^) and P. chiangmaiensis (MFLUCC 22–0127^T^) showed 0.8% (560/565, one gap) in ITS, 7.4% (250/270, eight gaps) in tef1-α, and 3.5% (440/456, one gap) in tub2. Sun et al. (2023) proposed that the individual cell length characteristics of the three median cells should not be considered criteria for interspecific differentiation, as individual cell length measurements duplicate the description of the total length of the three median cells. Therefore, it was suggested that individual cell length measurements be removed from morphological descriptions to achieve simplification. Additionally, based on phylogenetic analyses, P. exudata P. Razaghi, F. Liu & L. Cai and P. multiappendiculata P. Razaghi, F. Liu & L. Cai were proposed as novel species (Razaghi et al. 2024), but their conidial septation (4–5-septate) is inconsistent with the generally accepted morphological concept of 5-celled conidia for Pestalotiopsis species. Although phylogenetic analyses together with morphology currently serve as an effective approach for identifying Pestalotiopsis species, ambiguous morphological concepts can render the classification of these taxa questionable.
To date, 457 species of Pestalotiopsis have been recorded in Index Fungorum (Index Fungorum 2025), but molecular data are available in GenBank for only 159 species. The absence of molecular data precludes the resolution of interspecific relationships through phylogenetic analyses and results in substantial ambiguity in the taxonomic placement of morphologically congruent species within the genus. Maharachchikumbura et al. (2012) tested 10 gene regions (ACT, tub2, CAL, GPDH, GS, ITS, LSU, RPB1, SSU, and tef1-α) to resolve cryptic Pestalotiopsis species and ultimately selected ITS, tef1-α, and tub2 as the most effective molecular markers. However, recent phylogenetic analyses based on these three markers revealed that some species, such as P. verruculosa (MFLUCC 12–0274), lacking tub2, and P. chinensis (MFLUCC 12–0273), P. sequoiae (MFLUCC 13–0399), and P. yunnanensis (HMAS 96359), lacking tub2 and tef1-α, showed low support values (Hsu et al. 2024; Li et al. 2024; Yin et al. 2024). The low support values for these taxa may be attributed to the absence of tef1-α and/or tub2 sequence data, which likely resulted in insufficient phylogenetic information. Thus, completing the ITS, tef1-α, and tub2 sequence information for Pestalotiopsis species is crucial for enabling a more robust discussion of their taxonomic status through phylogenetic analyses. Considering this issue, we conducted phylogenetic analyses using ITS, tef1-α, and tub2 sequences, and our newly obtained 21 strains nested within the genus Pestalotiopsis formed six independent lineages with reliable support values and can be proposed as five new phylogenetic species, namely P. acericola, P. corchorifolii, P. fraseri, P. goeppertiae, and P. koelreuteriae, and one known species, P. machiliana.
Appendages are often used as an important basis for the morphological identification of Pestalotiopsis species, but studies have confirmed their critical roles in conidial dispersal and host colonization (Nag Raj 1993; Gareth Jones 2006; Crous et al. 2012). This facilitates the widespread colonization of various plants by Pestalotiopsis as endophytes and pathogens, thereby enhancing host diversity and competitiveness for ecological niches. Pestalotiopsis causes diseases on a variety of host plants as a pathogen, including tea, ericaceous plants, grapes, pomegranates, blueberries, and other crops, resulting in significant economic losses (Horikawa 1986; Xu et al. 1999; McQuilken and Hopkins 2004; Keith et al. 2006; Espinoza et al. 2008; Joshi et al. 2009). Many Pestalotiopsis species, when living as endophytes, have been demonstrated to produce numerous secondary metabolites with diverse structural characteristics (Xu et al. 2010), and previous studies have reported over 160 different compounds isolated from Pestalotiopsis species, which demonstrate significant potential in antifungal and antimicrobial activities and compound synthesis (Xu et al. 2014). Thus, further research is necessary to contribute to the fields of plant pathology and fungal taxonomy and to explore their functional roles.
Supplementary Material
XML Treatment for Pestalotiopsis acericola
XML Treatment for Pestalotiopsis corchorifolii
XML Treatment for Pestalotiopsis fraseri
XML Treatment for Pestalotiopsis goeppertiae
XML Treatment for Pestalotiopsis koelreuteriae
XML Treatment for Pestalotiopsis machiliana
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Akinsanmi OA, Nisa S, Jeff-Ego OS, Shivas RG, Drenth A (2017) Dry flower disease of macadamia in Australia caused by Neopestalotiopsis macadamiae sp. nov. and Pestalotiopsis macadamiae sp. nov. Plant Disease 101(1): 45–53. 10.1094/PDIS-05-16-0630-RE 30682313 · doi ↗ · pubmed ↗
- 2Ariyawansa HA, Hyde KD (2018) Additions to Pestalotiopsis in Taiwan. Mycosphere: Journal of Fungal Biology 9(5): 999–1013. 10.5943/mycosphere/9/5/4 · doi ↗
- 3Ariyawansa HA, Hyde KD, Jayasiri SC, Buyck B, Chethana KWT, Dai DQ, Dai YC, Daranagama DA, Jayawardena RS, Lücking R, Ghobad-Nejhad M, Niskanen T, Thambugala KM, Voigt K, Zhao RL, Li G-J, Doilom M, Boonmee S, Yang ZL, Cai Q, Cui Y-Y, Bahkali AH, Chen J, Cui BK, Chen JJ, Dayarathne MC, Dissanayake AJ, Ekanayaka AH, Hashimoto A, Hongsanan S, Jones EBG, Larsson E, Li WJ, Li Q-R, Liu JK, Luo ZL, Maharachchikumbura SSN, Mapook A, Mc Kenzie EHC, Norphanphoun C, Konta S, Pang KL, Perera RH, Phookamsak R · doi ↗
- 4Bate-Smith EC, Metcalfe CR (1957) Leuco–anthocyanins. 3. The nature and systematic distribution of tannin in dicotyledonous plants. Journal of the Linnean Society of London, Botany 55(362): 669–705. 10.1111/j.1095-8339.1957.tb 00030.x · doi ↗
- 5Cao B, Phurbu D, Ralaiveloarisoa A, Liimatainen K, Niskanen T, Ramírez-Cruz V, Bradshaw AJ, Dentinger BTM, Ramírez-Guillén F, Cortés-Pérez A, Guzmán-Dávalos L, Villalobos-Arámbula AR, Castro-Jauregui O, da Silva PS, Day R, Davoodian N, Lebel T, Castellano M, May TW, Merje Vasey J, Hofer K, Thangavel R, Braithwaite M, Braithwaite L, Dutta AK, Chattopadhyay P, Roy N, Tanti B, Biswas PR, Arumugam E, Kezo K, Kaliyaperumal M, Murugadoss R, Ji JX, Kakishima M, Cooper J, Nuytinck J, Luo ZL, Wang WP, Zh · doi ↗
- 6Chaiwan N, Wanasinghe DN, Mapook A, Jayawardena RS, Norphanphoun C, Hyde KD (2020) Novel species of Pestalotiopsis fungi on Dracaena from Thailand. Mycology 11(4): 306–315. 10.1080/21501203.2020.1801873 PMC 772302133329926 · doi ↗ · pubmed ↗
- 7Chen YY, Maharachchikumbura SSN, Liu JK, Hyde KD, Nanayakkara RR, Zhu GS, Liu ZY (2017) Fungi from Asian Karst formations I. Pestalotiopsis photinicola sp. nov., causing leaf spots of Photinia serrulata. Mycosphere: Journal of Fungal Biology 8(1): 103–110. 10.5943/mycosphere/8/1/9 · doi ↗
- 8Crous PW, Verkley GJM, Christensen M, Castañeda-Ruiz RF, Groenewald JZ (2012) How important are conidial appendages? Persoonia 28: 126–137. 10.3767/003158512 X 652624 PMC 340940923105158 · doi ↗ · pubmed ↗