Pathogenic fungus Ustilago maydis exploits the lateral root regulators to induce pluripotency in maize shoots
Mamoona Khan, Nithya Nagarajan, Kathrin Schneewolf, Caroline Marcon, Danning Wang, Frank Hochholdinger, Peng Yu, Armin Djamei

TL;DR
A fungus called Ustilago maydis causes galls in maize by hijacking the plant's lateral root development pathway.
Contribution
The study identifies how the fungus uses specific proteins to manipulate plant signaling for gall formation.
Findings
Tip4 protein activates auxin signaling, leading to pluripotent calli in maize without added hormones.
Genes upregulated during gall formation overlap with those in lateral root development.
Mutations in LBD genes (ra2, rtcs) reduce gall formation, confirming their role in the process.
Abstract
Biotrophic plant–pathogens secrete effector molecules to redirect and exploit endogenous signaling and developmental pathways in their favor. The biotrophic fungus Ustilago maydis causes galls on all aerial parts of maize. However, the responsible gall‐inducing effectors and corresponding plant signaling pathway(s) remain largely unknown.Using molecular and genetic approaches, and transcriptomic comparisons in maize, we identify downstream targets and developmental consequences of the plant TOPLESS (TPL)‐interacting protein (Tip) effectors in gall formation.We demonstrate that Tip4 derepress AtARF7/AtARF19 branch of auxin signaling, leading to the formation of pluripotent calli without the external addition of phytohormones. Comparative transcriptomics in maize further reveals a significant overlap of genes upregulated during U. maydis‐triggered leaf gall formation and the developmental…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Fig. 1
Fig. 1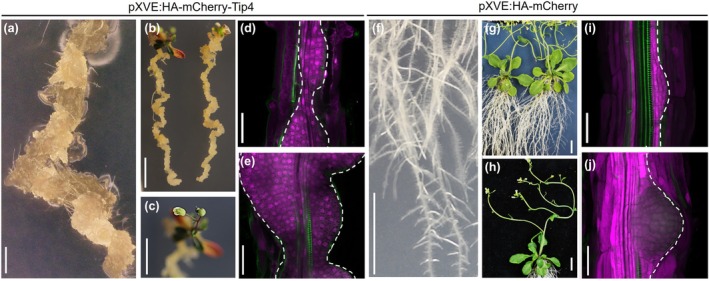 Fig. 2
Fig. 2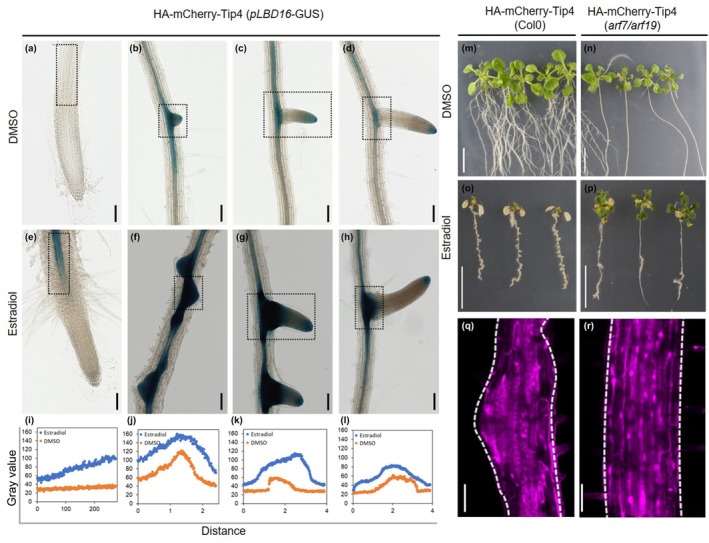 Fig. 3
Fig. 3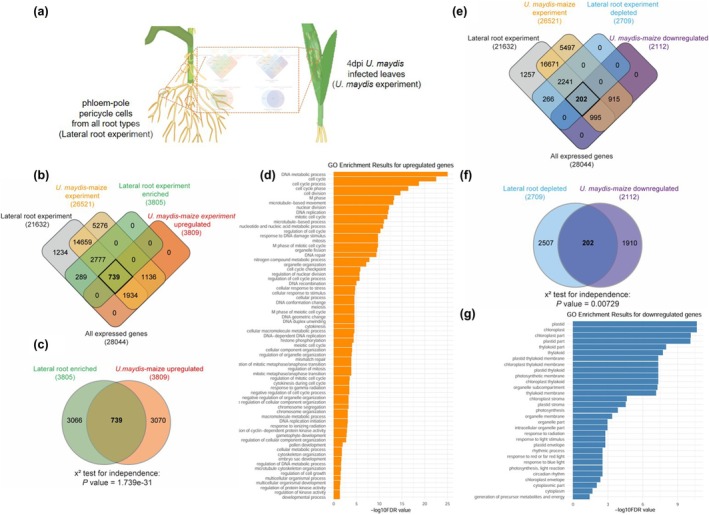 Fig. 4
Fig. 4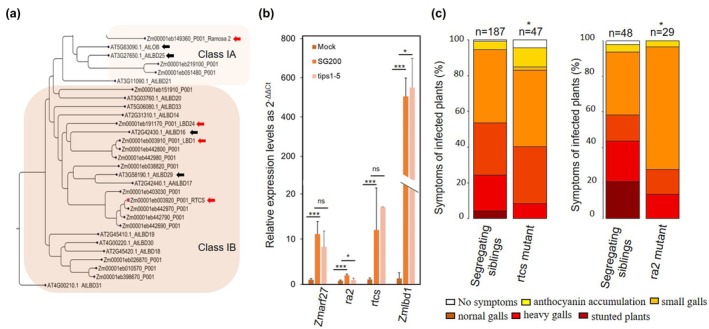 Fig. 5
Fig. 5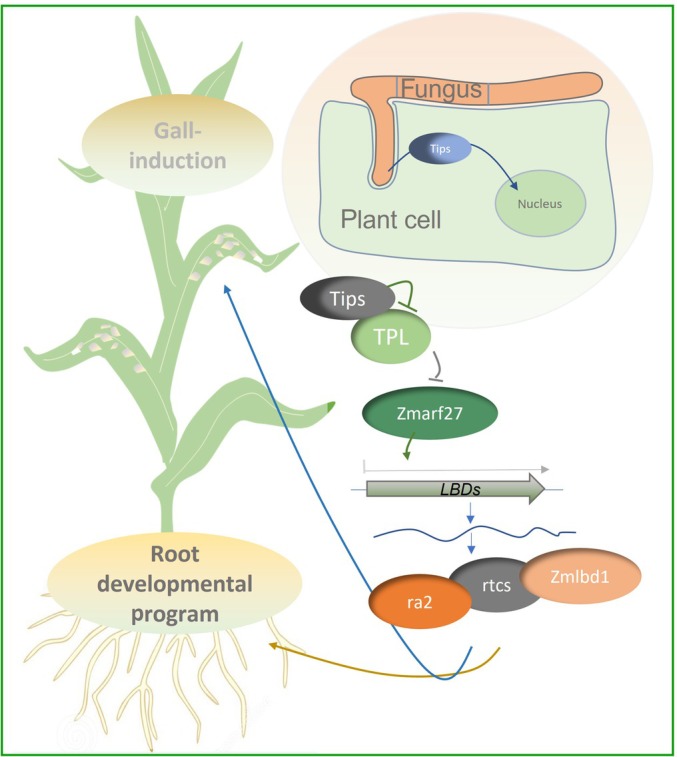 Fig. 6
Fig. 6- —European Research Council10.13039/501100000781
- —Deutsche Forschungsgemeinschaft10.13039/501100001659
- —Austrian Science Fund10.13039/501100002428
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Gene Expression Analysis · Plant Molecular Biology Research · Plant-Microbe Interactions and Immunity
Introduction
Pathogen‐induced plant galls are the morphological outcome of abnormal growth of plant tissue induced by the manipulative activities of the invading organism. These result from the increased proliferation (hyperplasia) and/or increased cell size of a group of cells (hypertrophy). Although more than a century of research, neither the physiological networks nor the exact mechanisms of gall induction and development have been fully elucidated (Dodueva et al., 2020). Ustilago maydis, a basidiomycete fungus, causes common smut disease in maize, infecting all aerial parts of the host, including the stem, leaves, and flowers. Early infection symptoms include local chlorosis and anthocyanin accumulation. A hallmark of U. maydis infection in maize is the formation of galls on aboveground organs, which act as sink tissues that, by the end of the fungal proliferation cycle, become filled with countless black diploid teliospores observable as smut symptoms (Brefort et al., 2009). Ustilago maydisinduced gall formation results from intensive cell division and expansion in specific cell types (Matei et al., 2018). Notably, not all cells in the infected tissue proliferate, indicating that this process is cell‐type or zone‐specific. For instance, in maize flowers, U. maydis colonizes only immature, undifferentiated anther cells with meristematic activity, sustaining cell division beyond normal development (Gao et al., 2013; Lin et al., 2021). In leaf galls, differentiated bundle sheath cells resume cell divisions, and mesophyll cells enlarge (Matei et al., 2018). However, the molecular mechanisms involved in gall formation remain poorly understood.
The U. maydis genome encodes 467 predicted secreted proteins, and several show distinct expression patterns during maize colonization (Lanver et al., 2018); however, only a few have been functionally characterized. Most known effectors are linked to immune suppression (Doehlemann et al., 2009; Navarrete et al., 2021, 2022; Saado et al., 2022), host metabolism, and phytohormone manipulation (Reineke et al., 2008; Djamei et al., 2011; Tanaka et al., 2014; Rabe et al., 2016; Ma et al., 2018) or fungal development in the host (Tanaka et al., 2020; Lin et al., 2023; Weiland et al., 2023). Yet little is known about effectors directly involved in gall formation. For example, See1 reactivates DNA synthesis in leaf galls but not in tassel galls; its deletion inhibits hyperplastic cell division (Redkar et al., 2015). Sts2 promotes hyperplasia by transcriptional dysregulation, and its deletion reduces cell division in bundle sheath cells (Zuo et al., 2023). Another effector, ApB73, plays a cultivar‐specific role in gall formation (Stirnberg & Djamei, 2016). The persistence of galls despite the deletion of these effectors implies functional redundancy or cooperation among multiple pathways in this complex developmental process.
Auxin, a central regulator of plant growth, also plays a role in plant–pathogen interactions (Kunkel & Johnson, 2021; Nagarajan et al., 2023). During U. maydis infection, auxin levels increase, partly due to fungal INDOLE‐3‐ACETIC ACID (IAA) production, although this alone is not essential for gall formation (Reineke et al., 2008). Auxin‐responsive genes are upregulated in infected tissues, and at least 10 U. maydis effectors (Jsi1, Nkd1, and Tip1–8) target TOPLESS (TPL) transcriptional corepressors to modulate auxin signaling (Darino et al., 2021; Bindics et al., 2022; Navarrete et al., 2022; Huang et al., 2023; Khan et al., 2024). A pentuple deletion mutant of these (Δtips1‐5) led to a significant reduction in gall sizes and numbers (Bindics et al., 2022), highlighting the crucial role of TPL proteins in the biotrophic stage of U. maydis. However, the specific downstream targets of TPL‐controlled signaling remain to be explored. TPL corepressors in auxin signaling are recruited by AUXIN (Aux)/IAA proteins through conserved Ethylene‐responsive element binding factor‐associated Amphiphilic Repression (EAR) motifs (Tiwari et al., 2004; Szemenyei et al., 2008). Aux/IAAs do not bind DNA directly but interact with AUXIN RESPONSE FACTORs (ARFs) to suppress transcription. Since the ability of Aux/IAAs to modulate transcription is dependent on ARFs, the presence of different ARF complements in different cells can also affect auxin signaling specificity (Rademacher et al., 2012; Bargmann et al., 2013; Leyser, 2018).
Auxin also regulates pluripotency and organogenesis by regulating fate, division, and differentiation, enabling new organ formation throughout the plant lifecycle (Perianez‐Rodriguez et al., 2014). Lateral root (LR) formation in plants is an example of postembryonic development. In Arabidopsis thaliana, LRs originate exclusively from pericycle founder cells in response to local auxin maxima (Celenza Jr. et al., 1995; Casimiro et al., 2001), which results in the derepression of AtARF7 and AtARF19, and the expression of LATERAL ORGAN BOUNDARY (LOB) DOMAIN (LBD) transcription factors (TFs; Wilmoth et al., 2005; Okushima et al., 2007; Lee et al., 2009). Formation of callus, proliferating masses of pluripotent cells from various differentiated explants, is often the first step in in vitro plant generation and is also initiated by elevated auxin levels in the tissue culture medium (Lardon & Geelen, 2020). Intriguingly, callus formation involves an ectopic activation of the root primordia development program from pericycle or pericycle‐like cells, even when derived from aerial organs, such as cotyledons and petals (Atta et al., 2009; Sugimoto et al., 2010; Xu et al., 2012). Auxin accumulation in the founder cells mediates the degradation of IAA14/SOLITARY ROOT, which releases AtARF7 and AtARF19 that in turn upregulate the expression of AtLBD16, AtLBD18, and AtLBD29 during callus formation (Lardon & Geelen, 2020). Ectopic LBD expression can induce callus without exogenous hormones, while their suppression inhibits auxin‐induced callus formation (Okushima et al., 2005; Fan et al., 2012). The LBD family comprises 43 members in A. thaliana (Fan et al., 2012) and 49 in Zea mays (Zhang et al., 2020); however, only a few have been shown to regulate LR organogenesis and callus formation in A. thaliana (Feng et al., 2012; Goh et al., 2012; Lee et al., 2013; Pandey et al., 2018), and functional knowledge on LBDs in maize is limited. Rootless concerning crown and seminal roots (rtcs) and rtcs‐like (rtcl) are two paralogous genes in maize and orthologues of A. thaliana AtLBD29 (Berardini et al., 2015) that control seminal and postembryonic shoot‐borne root formation (Taramino et al., 2007; Xu et al., 2015). ramosa 2 (ra2) is an orthologue of A. thaliana AtLBD25 and AtLOB genes (Zhang et al., 2020) and controls inflorescence branching (Bortiri et al., 2006). Although ra2 is highest expressed in the roots of maize (Winter et al., 2007; Hoopes et al., 2019; Woodhouse et al., 2021), its direct role in LR formation in maize has not been explored.
In this study, we reveal the role of a set of Tip effectors in U. maydis‐induced cellular dedifferentiation, cell division, and gall formation. We show that a single Tip effector can induce cellular dedifferentiation leading to callus formation and cell division in transgenic A. thaliana plants. Furthermore, we provide genetic evidence that this process relies on the expression of AtARF7 and AtARF19 TFs. It is also demonstrated that U. maydis induces the expression of Zmarf27, an orthologue of AtARF7 and AtARF19, and the LBDs, which are dominant susceptibility factors in maize during biotrophic colonization. Transcriptomic overlap between gall formation and LR development supports the notion that U. maydis exploits parts of the LR formation pathway in maize for leaf‐gall induction. Ultimately specific maize LBD mutants (ra2 and rtcs) show significantly reduced gall formation in comparison to wild type maize upon U. maydis infection.
Materials and Methods
Molecular cloning
Cloning was performed using either Greengate (Lampropoulos et al., 2013) or Golden Gate (Katzen, 2007) cloning systems as described previously (Khan et al., 2024). Mach1 competent cells (Thermo Fisher Scientific, Waltham, MS, USA) were used for all DNA manipulations and were grown in a double Yeast Extract Tryptone medium (dYT) liquid medium or on YT agar plates with the required antibiotic supplements. pXVE‐HA‐mCherry‐(effector lacking signal peptide) was the construct used for generating transgenic effector lines.
Plant material and growth conditions
Arabidopsis thaliana (L.) Heynh. Columbia was a wild‐type used for generating all transgenic lines. pAtLBD16:GUS reporter line and T‐DNA insertional mutant arf7, arf19 lines were obtained from the Nottingham Arabidopsis Stock Centre (NASC) under the ID N68141 and N24629 respectively and ra2‐R maize mutant seeds were received from maize genetic and cooperation stock center (USA). A. thaliana for dipping were grown in controlled short‐day conditions (8 h : 16 h, 21°C : 2°C, light : dark) while maize was grown in distinct conditions (14 h : 10 h, 28°C : 20°C, light : dark). Floral dipping was used to generate two transgenic A. thaliana lines for each plasmid construct with similar protein levels, selected with glufosinate‐ammonium on soil. For phenotyping, the plants were grown on solidified half‐strength MS agar media, supplemented with 1% (w/v) sucrose plates in growth cabinets (Panasonic environmental test chamber, Type: MLR‐352H‐PE) at 21°C ± 2°C and on long days of 16 h : 8 h, light : dark cycles with 80 μmol/m^2^/s intensity. For estradiol treatments, 7‐d‐old plate‐grown seedlings were transferred either to dimethyl sulfoxide (DMSO) or 10 μM β‐estradiol containing ½ MS plates, and pictures were taken 5, 10, 15, and 20 d after transfer to the plates or otherwise stated. Experiments were repeated at least three times.
Maize infection assays
Maize (Zea mays L.) variety Early Golden Bantam, Old Seeds, Madison, WI, USA, was used for all infections until otherwise stated. Ustilago maydis (DC.) Corda progenitor strain SG200 was used to infect seven‐day‐old maize seedlings for quantitative reverse transcription polymerase chain reaction (qRT‐PCRs) and nine‐day‐old maize seedlings for mutant analyses as described in detail previously (Redkar & Doehlemann, 2016). For qRT‐PCRs, leaf samples were collected c. 1 cm below the hole of the syringe in infected leaves 4 days post infection (dpi), whereas symptom scoring was performed at 12 dpi according to Kamper et al. (2006). Symptom scores were assessed using the Fisher's exact test in R, as described previously (Stirnberg & Djamei, 2016).
Confocal microscopy
Confocal microscopy was performed with a Leica SP8 confocal microscope. mCherry was excited at 561 nm, and emission was collected between 578 and 648 nm. Images were processed using the LAS‐X software from Leica.
qRT‐PCR and comparative transcriptome analyses
mRNA was extracted from the ground powder of leaves using a New England Biolabs GmbH (Frankfurt am Main, Germany) RNA extraction kit, and cDNA was synthesized using a Thermo Scientific RevertAid First Strand cDNA Synthesis Kit. Quantitative polymerase chain reaction (qPCRs) were performed with GoTaq qPCR mix (NEB, cat. no. A6001) according to the manufacturer's instructions. Relative amounts of amplicons were calculated according to the 2^−ΔΔCt^ method (Livak & Schmittgen, 2001). The results are the average of four biological replicates. Zmcdk (Zm00001eb350890) was used as housekeeping gene (Lin et al., 2014).
Biostatistical analysis of lateral root dataset and U. maydis dataset
To understand the biological pathway during LR initiation and callus formation in maize, a previously published transcriptomic dataset of U. maydis‐infected maize leaf tissues (Lanver et al., 2018), designated here as *U. maydis‐*maize experiment, was downloaded, and only the time point 4 dpi was selected, as it is the time when galls are formed. The LR transcriptomics, which used cell‐type‐specific RNA sequencing of LR mutant vs wild‐type maize plants via laser capture microdissection, designated here as the LR experiment, was generated in Baer et al. (2025) and is available under sequence read archive accession no. PRJNA1366082. Genes preferentially expressed in phloem‐pole pericycle cells were defined as ‘LR‐enriched’, while genes expressed in xylem‐pole pericycle cells were defined as ‘LR‐depleted’. DESeq2 was used to test for gene expression changes. For both datasets, only genes with at least five normalized read counts for at least one time point/cell type from three replicates were considered as expressed. A differential expression threshold of log_2_ fold change > 1 and Benjamini–Hochberg‐adjusted P value <0.05 was used in both datasets. The overlapping and differentially expressed genes between datasets were visualized by Venn diagram (https://bioinformatics.psb.ugent.be). A chi‐squared (χ^2^) test in R (v.1.4.1717) was used to determine whether the two transcriptome datasets share a significant correlation. The overlapping genes between datasets were functionally characterized by Gene Ontology (GO) annotation and enrichment analysis via AgriGO (v.2, http://systemsbiology.cpolar.cn/agriGOv2/).
Results
Induction of Tip expression leads to strong morphological phenotypes in planta
Although U. maydis has at least 10 TPL‐interacting effectors (Jsi1, Nkd1, Tip1‐8) (Darino et al., 2021; Bindics et al., 2022; Navarrete et al., 2022; Huang et al., 2023; Khan et al., 2024), it is not clear why so many are needed and which downstream pathways are activated. As TPL corepressors are central, conserved negative transcriptional regulators in all land plants, we generated transgenic A. thaliana lines expressing each of these Tip effectors to study their individual biological activity. Intriguingly, this resulted in a range of strong morphological phenotypes across two independent transgenic lines tested compared to an mCherry‐expressing control (Supporting Information Figs 1a, S1–S3). More specifically, the induction of Tip1, Tip2, Tip8, Jsi1, and Nkd1 expression in A. thaliana led to chlorophyll (Chl) loss in the cotyledons and leaves, and overall growth arrest already clearly visible at 4 d after transfer (Figs 1b, S1, S2), while expression of Tip3, Tip4, Tip5, Tip6, and Tip7 led to a strong induction of LR formation and inhibition of primary root length (Figs 1c, S3, S4). Noticeably, the TPL‐interacting effectors that lead to overall growth inhibition and complete Chl loss in A. thaliana previously showed a cell death phenotype upon transient overexpression in Nicotiana benthamiana leaves (Darino et al., 2021; Navarrete et al., 2022; Khan et al., 2024). Based on their cell death‐inducing feature vs LR induction abilities, we categorized the 10 known TPL‐interacting effectors into two classes (Table S1), class I and II represented by Tip1 and Tip4, which exhibited the strongest phenotype of their respective class (Fig. 1b,c). Strikingly, in the strongest case of class II effectors (Tip3, Tip4, and Tip6), the LRs did not grow in size; instead, the whole root thickened due to the initiation of undifferentiated (callus‐like) structures that formed without the external addition of any phytohormones on the plate (Figs 1c, 2a–e, S3A,B,D) as compared to control plants (Figs 1a, 2f–j, S1A). Surprisingly, this phenotype was restricted to the roots only; in the shoots, the growth either slowed down or the leaves turned pale. These plants continued to survive, and the shoots even flowered (Fig. 2c). To test whether these undifferentiated (callus‐like) structures are indeed pluripotent, we transferred them (without their shoots) to the shoot induction media used for A. thaliana regeneration. This led to greening and shoot formation (Fig. S5), supporting this conclusion. To summarize, ectopic expression of TPL‐interacting effectors in A. thaliana resulted in extreme morphological phenotypes, which could be grouped into two distinct classes. In this study, we focus on class II Tip effectors whose expression leads to LR and callus formation in A. thaliana for in‐depth investigation.
Expression of TOPLESS‐interacting protein (Tip) effectors induces strong morphological phenotypes in Arabidopsis thaliana. Seven‐day‐old A. thaliana seedlings expressing (a) pXVE: HA‐mCherry, (b) pXVE: HA‐mCherry‐Tip1, or (c) pXVE: HA‐mCherry‐Tip4 were transferred to agar plates containing estradiol, and images were taken at 0, 4, 8, and 12 d after transfer. DAT, d after transfer. Bars, 1 cm.
TOPLESS‐interacting protein (Tip) effector 4 induces callus‐like structures along the primary root. (a, b) Images of plants expressing pXVE:HA‐mCherry‐Tip4 at 12 d after transfer (DAT) (a) and 29 DAT (b) to estradiol induction medium, showing extensive callus formation along the primary root. (c) Flowering observed in a pXVE:HA‐mCherry‐Tip4‐expressing plant at 29 DAT. (d, e) Confocal images of the root elongation zone in plants expressing pXVE:HA‐mCherry‐Tip4 showing morphologies associated with callus formation. (f, g) Images of control plants expressing pXVE:HA‐mCherry at 12 DAT (f) and 29 DAT (g) to estradiol induction medium, showing normal root development. (h) Flowering observed in a pXVE:HA‐mCherry‐expressing plant at 29 DAT. (i, j) Confocal images of the root elongation zone in control plants expressing pXVE:HA‐mCherry showing morphologies associated with lateral root formation. Green fluorescence indicates auramine staining of the root vasculature. Bars, a = 1 mm; b, c, f, g, h = 1 cm; d, e, i, j = 50 μm.
Overexpression of class II Tips leads to the formation of calli in A. thaliana roots through the TPL‐AtARF7/AtARF19‐AtLBD16 pathway
Expression of class II U. maydis Tip effectors (Tip3, Tip4, Tip5, Tip6, and Tip7) led to the initiation of LRs and pluripotent callus in the A. thaliana roots. Similar phenotypes have been described previously for LBD TF overexpression in A. thaliana, which act downstream of AtARF7 and AtARF19 during LR formation and for callus formation (Fan et al., 2012; Liu et al., 2018). We therefore hypothesized that class II Tips, following their specific interaction with the TPL class of corepressors (Bindics et al., 2022; Navarrete et al., 2022), interfere with the binding of Aux/IAA corepressors and derepress the auxin signaling cascade associated with LR and callus formation in A. thaliana. To test this, we first crossed pAtLBD16:GUS (the expression of β‐glucuronidase (GUS) reporter is driven under the control of the LBD16 promoter; Bargmann et al., 2014) with the Tip4 expression line pXVE:HA‐mCherry‐Tip4 (Tip4 representative for the class II phenotypes) and observed the GUS‐reporter expression pattern. Microscopic observation revealed that pAtLBD16:GUS expression was strongly induced in a Tip4‐dependent manner in seedling roots after estradiol induction (Figs 3a–l, S6). This implies that Tip‐targeted TPLs are major negative regulators of AtLBD16 in the root, but there may be other factors in Arabidopsis shoots.
TOPLESS‐interacting protein (Tip) 4 effector‐induced root callus formation requires AtARF7 and AtARF19‐mediated AtLBD16 expression. (a–h) Digital microscopy images of seedlings expressing pLBD16:GUS in a pXVE:HA‐mCherry‐Tip4 background. Seven‐day‐old seedlings grown on ½ MS agar were transferred to either dimethyl sulfoxide (DMSO)‐containing medium (a–d) or 10 μM estradiol (e–h) for 4 d before β‐glucuronidase (GUS) staining. Bars, 100 μm. (i ‐l) The GUS staining intensities were evaluated from the images (a–h) in the indicated regions (rectangles) by using fiji (Schindelin et al., 2012) as described previously (Beziat et al., 2017). Orange line in each graph indicate GUS intensity in DMSO‐treated seedling and the blue line in each graph represents GUS intensity in estradiol ‐treated seedling. (m–p) Phenotypes of seedlings expressing pXVE:HA‐mCherry‐Tip4 in wild‐type Columbia (Col‐0; m, o) or arf7/arf19 mutant background (n, p) 10 d after transfer to plates containing DMSO (m, n) or 10 μM estradiol (o, p). Bars, = 1 cm. (q, r) Confocal microscopy of 11‐d‐old Arabidopsis thaliana roots expressing pXVE:HA‐mCherry‐Tip4 in Col‐0 (q) or arf7/arf19 background (r), showing Tip4 effector protein expression 4 d after induction. Bars, 50 μm.
To provide further genetic evidence that morphological phenotypes of class II overexpressing effector lines are a result of the derepression of a branch of the auxin signal cascade that controls LR and callus formation through AtARF7 and AtARF19, we examined the overexpression phenotypes of Tip4 in an arf7/arf19 T‐DNA insertional double mutant background (Okushima et al., 2005), which does not produce LRs. For this purpose, we crossed pXVE:HA‐mCherry‐Tip4 (Fig. 3m) expressing A. thaliana plants with the arf7/arf19 double mutant (Fig. 3n) and observed the phenotypes of homozygous pXVE:HA‐mCherry‐Tip4 arf7/arf19 seedlings in the F2 generation after DMSO (Fig. 3m,n) or estradiol (Fig. 3o–p) treatments. As shown in Fig. 3m–r, whereas the Tip4‐expression‐mediated root length inhibition stayed unaffected (Fig. S4), the Tip4‐expression‐mediated induced callus formation was largely abolished in the arf7/arf19 background (Fig. 3o,p). Furthermore, the effect on leaf chlorosis upon class II Tip expression is strongly reduced in the arf7/arf19 mutant background (Fig. 3o,p). These results suggest a direct role of AtARF7 and AtARF19 in both phenomena, the Tip effector‐mediated callus formation in A. thaliana via the LR pathway, as well as the observed effects of Chl loss in the leaf chloroplasts.
Ustilago maydis induces the expression of genes involved in lateral root formation during biotrophy in maize leaves
To estimate the role of the LR and callus formation pathway during U. maydis biotrophy on maize at the transcriptomic level, we compared previously published transcriptomic data of U. maydis‐infected maize leaf tissues at 4 dpi, the time point of gall formation (Lanver et al., 2018), with transcripts differentially regulated in phloem‐pole pericycle cells, the cell type, which gives rise to LRs in maize (Fig. 4a; Yu et al., 2016). A comparison of these two datasets indicates several genes are commonly regulated during these two biological processes (Fig. 4). In total, 28 044 genes were expressed in sum of the two independent experiments (LR experiment and U. maydis‐maize infection experiment). Among 20 109 commonly expressed genes, we found 3805 genes enriched in phloem‐pole pericycle cells, whereas upon U. maydis infection, 3809 maize genes are significantly (fold change > 2; FDR < 0.05) induced at 4 dpi (Fig. 4b). Between the two datasets of upregulated genes, 739 genes are upregulated in common, which is highly significant in a chi‐squared test for independence (P = 1.739e‐31; Fig. 4c).
Ustilago maydis induces the expression of central regulators of the lateral root programming in maize shoots (a) Schematic overview of the experimental design. Transcriptomic data from U. maydis‐infected maize leaf tissue at 4 d post‐infection (4 dpi), the stage of gall formation (designated here as U. maydis‐maize experiment), were compared with gene expression profiles from phloem‐pole pericycle cells, the cell type responsible for LR initiation in maize, of 6‐wk‐old plants (designated here as lateral root (LR) experiment). (b) 4‐way Venn diagrams illustrating the overlap pattern of four different transcriptional datasets: expressed genes of LR experiment (21632), expressed genes of U. maydis maize infection experiment (26521), LR initiation enriched genes (3805), and U. maydis maize infection upregulated genes (3809) at 4 dpi. (c) Commonly upregulated genes between transcripts enriched in LR initiation cells and transcripts upregulated in 4dpi U. maydis‐infected maize. (d) The top enriched Gene Ontology (GO) terms are shown for biological processes. Bar thickness represents the number of associated genes, while bar height indicates significance (−log10 FDR). Terms with false discovery rate (FDR) < 0.05 were considered significantly enriched. (e) 4‐way Venn diagrams illustrating the overlap pattern of four different transcriptional datasets: expressed genes of LR experiment (21632), expressed genes of U. maydis ‐ maize infection experiment (26521), LR initiation depleted genes (2709), and U. maydis ‐ maize infection downregulated genes (2112) at 4 dpi. (f) Commonly downregulated genes between transcripts depleted in LR initiation cells and transcripts downregulated at 4 dpi U. maydis maize infection. Statistical significance was assessed using the chi‐squared test for independence. (g) The top enriched GO terms are shown for biological processes. Bar thickness represents the number of associated genes, while bar height indicates significance (−log10 FDR). Terms with FDR < 0.05 were considered significantly enriched.
Subsequently, the 739 commonly induced genes between the datasets were functionally classified according to GO terms using agriGOv2. In total, 69 GO terms belonging to different biological processes displayed significant overrepresentation (false discovery rate (FDR) < 0.05) (Dataset S2; Fig. 4d). Of all GO terms, the most enriched are involved in processes of cell cycle and cell division, suggesting the potential linkage of LR initiation and U. maydis‐induced gall formation in leaves. Also, the overlap of 202 commonly downregulated genes (Fig. 4e) between the U. maydis‐maize infection data and the LR initiation shows high significance in a chi‐squared test for independence (P = 0.00729; Fig. 4f). Analysis of the 202 genes (Fig. 4e,f) from 2709 genes transcriptionally underrepresented in phloem‐pole pericycle cells showed enrichment for 31 GOs (FDR < 0.05; Dataset S3; Fig. 4g). In particular, these involved photosynthesis and chloroplast‐related pathways, consistent with sink‐tissue formation and loss of photosynthetic activity at the place of gall induction by U. maydis.
Taken together, comparison of the two independent transcriptomic datasets generated from very different tissues and with different biological questions shows a significant overrepresentation of commonly up‐ and downregulated genes. This supports the notion that the shoot‐infecting fungus U. maydis recruits the LR pathway during the induction of galls on maize shoots.
Ustilago maydis induces the expression of maize Zmarf27, the orthologue of
AtARF7 and AtARF19 , in maize leaves
Next, we hypothesized that class II Tip effectors induce gall formation in maize leaves by employing common signaling components of the LR and callus formation pathway, including AtARF7/AtARF19 and LBDs. Therefore, we searched for maize orthologues of these pathway genes and identified one gene (Zmarf27) as the orthologue of both AtARF7 and AtARF19, and two genes (Zmlbd24 and Zmlbd1) as orthologues of AtLBD16 (Fig. 5a; Berardini et al., 2015). We then examined their expression levels in *U. maydis‐*infected maize leaves compared to mock control by quantitative real‐time PCR (qRT‐PCR). We also included rtcs, and ra2, orthologues of AtLBD29, that is a closest homologue of class‐IB LBD (Fig. 5a) genes of A. thaliana. Ra2 was previously found to be downregulated in the pentuple‐deletion‐mutant (Δtips1‐5)‐infected maize seedlings (Khan et al., 2024). Moreover, ra2‐like binding sites were enriched in differentially expressed genes from Δtip6 mutant‐infected maize leaves (Huang et al., 2023). Strikingly, the expression levels of Zmarf27, ra2, and Zmlbd1 were significantly and specifically induced in the leaves infected with U. maydis (Fig. 5b). At the same time, rtcs was consistently upregulated but not significantly different between U. maydis‐infected leaves compared with mock treatment in our assays. The transcript abundance of Zmlbd24 was below detection limits in both U. maydis‐infected and mock leaves (data not shown). To further investigate the role of class II Tips, we examined the expression of these genes in the Tips pentuple mutant (Δtips1‐5; Bindics et al., 2022), in which three class II Tips (Tip3, Tip4, and Tip5) are absent, and observed a small but significant reduction in ra2 and Zmlbd1 expression.
Zmarf27 and certain ZmLBDs (ra2, rtcs, Zmlbd1) are required for Ustilago maydis‐induced gall formation. (a) The phylogeny of LATERAL ORGAN BOUNDARY (LOB) domains (LBD) proteins of class‐IA and class IB of Arabidopsis thaliana and Zea mays. The phylogenetic tree was reconstructed using aligned amino acid sequences of LOB domain proteins as an unrooted tree using CLC‐Genomics software 20.04 and a bootstrap value of 1000. Red arrows point to maize genes studied in the b,c and black arrows point to their A. thaliana orthologues. (b) The expression levels of Zmarf27, ra2, rtcs, and Zmlbd1 were tested by quantitative real‐time PCR analyses at 4 d post‐infection of U. maydis SG200 solopathogenic strain, Δtips1‐5 mutant strain or mock (water)‐infected maize seedlings. The 2−ΔΔCt values were calculated and plotted as relative expression levels compared with mock‐infected plants as negative control. Error bars indicate SD. Significant differences between mock control and SG200‐infected leaves, and Δtips1‐5 mutant strain were analyzed by Student's t‐test analysis (, P < 0.05; *, P < 0.005, ns, not significant). Data represent an average of four biological replicates. (c) Nine‐day‐old recessive homozygous maize mutants in rtcs or ra2 genes were infected with U. maydis solopathogenic strain SG200 and compared to segregating population controls. Significant differences were analyzed by Fisher's exact test (, P < 0.05).
To elucidate the role of this pathway during U. maydis biotrophy, we next looked for the maize mutants of the respective LOB‐domain TFs and identified previously published mutants in rtcs and ra2 genes. We tested the ability of the recessive rtcs and ra2 loss‐of‐function mutants (Fig. S7) to form galls upon U. maydis infection. For this purpose, 9‐d‐old maize seedlings of segregating rtcs‐1 and ra2‐R alleles were infected with the solopathogenic U. maydis strain SG200, and symptom scoring was performed 12 dpi. The results of this experiment show a slight but significant reduction in virulence in homozygous recessive mutants compared with the segregating population (Fig. 5c). Taken together, U. maydis induces the expression of Zmarf27 and certain LBDs during its biotrophy in maize leaves, and single rtcs and ra2 mutants show virulence defects, suggesting a role of this branch of auxin signaling during gall formation (Fig. 6). Moreover, the weaker virulence defects of recessive single mutants of LBD genes indicate their redundant roles in this process.
Working model for class II Topless (TPL)‐interacting protein effectors in maize shoot reprogramming. Class II TPL‐interacting protein (Tip3, Tip4, Tip5, Tip6, and Tip7) effectors translocate to the plant nucleus and interact with maize TPL proteins during Ustilago maydis biotrophy, leading to de‐repression of maize auxin response factor 27 (Zmarf27). Zmarf27 can then bind the promoters of specific LATERAL ORGAN BOUNDARIES DOMAIN transcription factors ra2, rtcs, and Zmlbd1, driving transcriptional reprogramming toward cellular dedifferentiation and gall formation in the leaves where they do not express under normal developmental conditions. Blunt ended arrow represents inhibition while a pointed arrow indicates upregulaton.
Discussion
More than a century of U. maydis research has been in part driven also by the curiosity of how this fascinating biotrophic fungus can induce prominent galls on all aerial parts of its host plant maize (Küster, 1911). However, the underlying mechanisms remained elusive. Since the U. maydis genome has been published (Kamper et al., 2006), many effector genes have been placed into the context of gall formation based on studies of their deletion strains, which were impaired in pathogenicity and hence altered gall formation upon infection (Brefort et al., 2014; Redkar et al., 2015; Lanver et al., 2017; Djamei et al., 2023; Khan & Djamei, 2024). Nevertheless, it was unknown how many factors would be needed to induce galls in planta in the absence of fungal infection. Here, we provide significant insights through in planta by Tip effector overexpression, in which major phenotypic changes were observed and could be classified in two distinct classes. For the strongest representative of the class II Tip effectors, Tip 4, we provide genetic evidence that it derepresses AtARF7 and AtARF19 TFs, leading to activation of LBD genes involved in LR and callus formation. In contrast to the previously reported effectors, the class II Tip effectors act dominantly and independently of the presence of U. maydis infection, in which overexpression of a single effector is sufficient to induce an endogenously encoded cellular dedifferentiation and proliferation program. In the strongest phenotypes (Tip3, Tip4, and Tip6), this results in pluripotent callus formation at positions in which LRs would normally emerge, whereas Tip5 and Tip7 primarily alter LR frequency and length. All class II Tips also induce leaf chlorosis, although the severity varies among effectors. The phenotypic variation within this group suggests that, although all class II Tips act as negative regulators of TPL/TPR proteins, their activities display a degree of specificity. We further show an overlap in transcriptional programming between *U. maydis‐*induced gall formation in maize leaves and LR emergence. These results are completely novel in the U. maydis–Zea mays pathosystem and in line with recent findings in A. thaliana that demonstrate auxin‐induced callus formation occurs from the pericycle (or pericycle‐like cells) within multiple organs through a root development pathway, during which the ectopic activation of root meristematic genes is required for subsequent regeneration programs (Che et al., 2007; Sugimoto et al., 2010).
The discrepancy that U. maydis causes galls occurs solely on the aerial organs of its host plant maize, while class II Tips overexpressed in A. thaliana solely induce calli in the roots, which requires some consideration. One explanation could be that during U. maydis infection, the complex background manipulation by a whole effectome creates metabolic and transcriptional preconditioning, which cannot be recapitulated by expression of a single U. maydis effector. The transcriptional derepression of AtLBD16 homologs (Fig. 5b) is therefore important but not sufficient to trigger callus formation in the shoot. Furthermore, this capacity appears to be restricted, as U. maydis induces cell division (hyperplastic leaf gall tissue) only in specific cell types, namely bundle sheath cells, rather than throughout the entire leaf (Matei et al., 2018). Nevertheless, class II Tips show clear effects also in the shoot of transgenic plants, that is chlorosis. This situation also occurs in U. maydis galls, which are sink tissues that show a loss of chloroplasts and turn pale yellow or red due to anthocyanin accumulation.
It has been demonstrated previously in A. thaliana that the formation of LRs, as well as the development of callus, both require several common regulatory components of the LR developmental pathways, that is AtARF7, AtARF19, AtLBD16, AtLBD18, AtLBD29, and AtLBD33 (Sugimoto et al., 2010; Fan et al., 2012; Perianez‐Rodriguez et al., 2014). The formation of callus upon Tip4 overexpression is almost abolished in the arf7/arf19 double mutant background, placing AtARF7 and AtARF19 downstream of Tip4 and demonstrating their importance in Tip4‐induced callus formation in A. thaliana. The fact that there were still a few callus‐like structures appearing upon Tip4 overexpression in the arf7/arf19 mutant background could be due to the involvement of other ARFs, for example AtARF5, in this process (Vangheluwe & Beeckman, 2021). Leaves of A. thaliana plants expressing class II Tips turn chlorotic, and this phenomenon seems also to be dependent on AtARF7 and AtARF19 as the leaf phenotypes of Tip4 are widely rescued in the arf7/arf19 mutant plants (Fig. 3o,p). We also observed this in western blots targeting class II Tips overexpression after already 5 d induction. This correlates with the yellowing of the leaf, and the Rubisco band in the ponceau‐staining of western blots is very faint (Fig. S8), indicating that not only Chl but also chloroplast function in total is negatively regulated by AtARF7 and AtARF19 activities. Consistent with this, GO‐enrichment analysis of the commonly downregulated genes between *U. maydis‐*infected maize leaves and LR initiation‐repressed genes revealed an overrepresentation of photosynthesis and chloroplast‐related components (Fig. 4g).
Gall formation is a widely occurring phenomenon caused by various pathogens such as gall wasps, bacteria such as Phytoplasmas, Pantoea agglomerans, Pseudomonas savastanoi, Xanthomonas citri, Rhodococcus fascians, the root‐knot and cyst nematodes, and certain rust fungi and smuts (Harris & Pitzschke, 2020). Gall formation might have several advantages for the colonization and proliferation in the host, including immune suppression (Navarrete et al., 2022) and efficient nutrient acquirement (Horst et al., 2010; Sosso et al., 2019). Furthermore, considering the biotrophic lifestyle of these pathogens, there is possibly also a reduction in interference with essential plant functionalities during massive proliferation in a separated tissue irrelevant for plant survival. We provide here evidence that U. maydis employs the postembryonic organogenesis pathway to induce galls. Unlike galls and giant cells formed by root‐knot nematodes, in which direct overexpression of AtLBD16 is induced for callus formation, the class II Tip effectors of U. maydis induce callus formation upstream of AtARF7 and AtARF19 by distinct suppression of TPL functions. This highlights an example of convergent evolution of the pathway comprising TPL as a negative regulator of AtARF7 and AtARF19.
Competing interests
None declared.
Author contributions
MK and AD were involved in conceptualization, methodology. MK, NN and KS were involved in investigation. DW and PY were involved in transcriptomic data generation and analysis. MK was involved in project administration. MK, NN, CM, FH and AD were involved in resources. MK and AD were involved in original draft. AD and MK were involved in funding acquisition and supervision.
Disclaimer
The New Phytologist Foundation remains neutral with regard to jurisdictional claims in maps and in any institutional affiliations.
Supporting information
Dataset S1. Transcriptomic comparison of U. maydis‐infected maize leaves at 4 d post infection with gene expression in phloem‐pole pericycle cells from 6‐wk‐old maize, the site of lateral root initiation. Dataset S2. List of 739 genes commonly induced in both datasets, functionally classified by Gene Ontology terms. Dataset S3. List of 202 genes commonly downregulated in both datasets, functionally classified by Gene Ontology terms.
Fig. S1 A. thaliana plants expressing Topless interacting protein effector 1 (Tip 1) show chlorophyll loss and inhibition of overall growth phenotypes. Fig. S2 A. thaliana plants expressing Topless interacting protein (Tip) effectors of class I showing chlorophyll loss and inhibition of overall growth phenotypes. Fig. S3 A. thaliana plants expressing Topless interacting protein (Tip) effectors of class II showing phenotypes of increased lateral roots/callus‐like structures and inhibition of root lengths. Fig. S4 The primary root lengths of A. thaliana seedlings expressing either pXVE:HAmCherry, or pXVE:HA‐mCherry‐Tip4. Fig. S5 The root explants of pXVE:HA‐mCherry‐Tip4 pre‐incubated with 10 μM estradiol for the induction of Tip4 expression were transferred to shoot‐inducing medium (SIM) to induce de novo shoot regeneration. Fig. S6 TOPLESS interacting protein (Tip)‐4 effector‐induced root callus formation requires AtLBD16 expression. Fig. S7 Characterization of ra2‐R and rtcs‐1 recessive mutants. Fig. S8 Arabidopsis. thaliana plants expressing Topless interacting protein effector 4 (Tip 4) show chlorophyll loss and absence of rubisco. Table S1 Summary of cell – death and other morphological phenotypes observed for Topless interacting protein (Tip) effectors of Ustilago maydis across different studies.Please note: Wiley is not responsible for the content or functionality of any Supporting Information supplied by the authors. Any queries (other than missing material) should be directed to the New Phytologist Central Office.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Atta R , Laurens L , Boucheron‐Dubuisson E , Guivarc'h A , Carnero E , Giraudat‐Pautot V , Rech P , Chriqui D . 2009. Pluripotency of Arabidopsis xylem pericycle underlies shoot regeneration from root and hypocotyl explants grown in vitro . The Plant Journal 57: 626–644.18980654 10.1111/j.1365-313X.2008.03715.x · doi ↗ · pubmed ↗
- 2Baer M , Zhong Y , Yu B , Tian T , He X , Gu L , Huang X , Gallina E , Metzen I , Bucher M et al. 2025. Ethylene signal‐driven plant‐multitrophic synergy boosts crop performance. bio Rxiv . doi: 10.1101/2025.11.28.690471. · doi ↗
- 3Bargmann BO , Birnbaum KD , Brenner ED . 2014. An undergraduate study of two transcription factors that promote lateral root formation. Biochemistry and Molecular Biology Education 42: 237–245.24615800 10.1002/bmb.20780 · doi ↗ · pubmed ↗
- 4Bargmann BO , Vanneste S , Krouk G , Nawy T , Efroni I , Shani E , Choe G , Friml J , Bergmann DC , Estelle M et al. 2013. A map of cell type‐specific auxin responses. Molecular Systems Biology 9: 688.24022006 10.1038/msb.2013.40PMC 3792342 · doi ↗ · pubmed ↗
- 5Berardini TZ , Reiser L , Li D , Mezheritsky Y , Muller R , Strait E , Huala E . 2015. The Arabidopsis information resource: Making and mining the “gold standard” annotated reference plant genome. Genesis 53: 474–485.26201819 10.1002/dvg.22877 PMC 4545719 · doi ↗ · pubmed ↗
- 6Beziat C , Kleine‐Vehn J , Feraru E . 2017. Histochemical staining of beta‐glucuronidase and its spatial quantification. Methods in Molecular Biology 1497: 73–80.27864759 10.1007/978-1-4939-6469-7_8 · doi ↗ · pubmed ↗
- 7Bindics J , Khan M , Uhse S , Kogelmann B , Baggely L , Reumann D , Ingole KD , Stirnberg A , Rybecky A , Darino M et al. 2022. Many ways to TOPLESS ‐ manipulation of plant auxin signalling by a cluster of fungal effectors. New Phytologist 236: 1455–1470.35944559 10.1111/nph.18315 · doi ↗ · pubmed ↗
- 8Bortiri E , Chuck G , Vollbrecht E , Rocheford T , Martienssen R , Hake S . 2006. ramosa 2 encodes a LATERAL ORGAN BOUNDARY domain protein that determines the fate of stem cells in branch meristems of maize. Plant Cell 18: 574–585.16399802 10.1105/tpc.105.039032 PMC 1383634 · doi ↗ · pubmed ↗