B‐GATA factors are required for nitrogen‐responsive growth in Physcomitrium patens and Arabidopsis thaliana
Dario Zappone, Peter Michael Schröder, Ivan Petřík, Xiao Dong, Rudi Schäufele, Korbinian Schneeberger, Ondřej Novák, Claus Schwechheimer

TL;DR
This study shows that B-GATA transcription factors are crucial for plant growth in response to nitrogen, linking them to hormonal and genetic processes.
Contribution
The study provides first experimental evidence for B-GATAs' role in nitrogen-dependent growth in land plants.
Findings
B-GATA mutants show impaired nitrogen-dependent growth and transcriptional regulation in Physcomitrium and Arabidopsis.
Mutants exhibit altered cytokinin homeostasis or signaling, linking hormonal imbalance to growth defects.
B-GATAs influence nitrogen-regulated transcription downstream from cytokinin in an evolutionarily conserved manner.
Abstract
We hypothesized that B‐GATA family transcription factors have important roles in growth regulation in moss.We analyzed B‐GATA family transcription factor mutants from Physcomitrium patens and Arabidopsis thaliana to assess growth, gene expression, and cytokinin‐related processes under varying nitrogen conditions.We found that nitrogen‐dependent growth and transcriptional regulation are strongly impaired in mutants from Physcomitrium and Arabidopsis. We detected altered cytokinin homeostasis or signaling in the mutants, linking hormonal imbalance to growth and transcription defects.We demonstrated a conserved, critical role of B‐GATAs in plant nitrogen‐responsive growth. Results suggest that B‐GATAs influence nitrogen‐regulated transcription downstream from cytokinin, supporting an ancient, evolutionarily conserved mechanism connecting nutrient signaling to growth. We provided…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
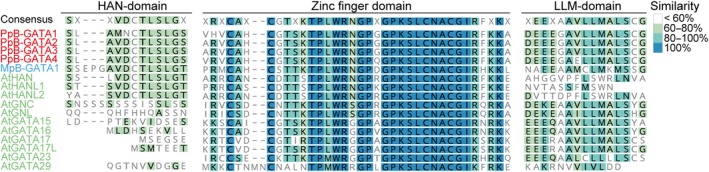 Fig. 1
Fig. 1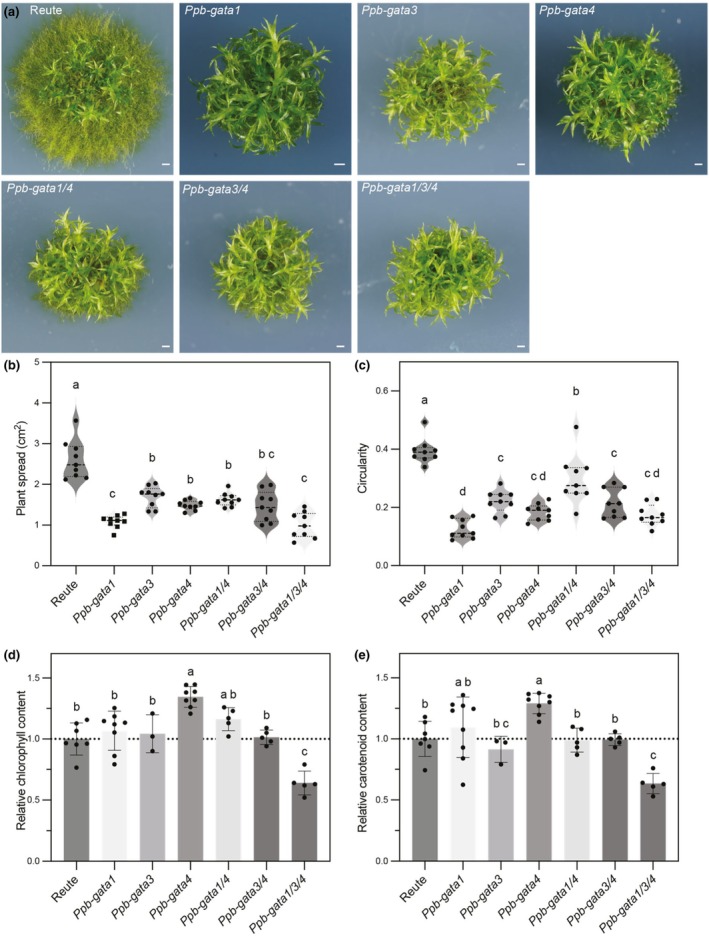 Fig. 2
Fig. 2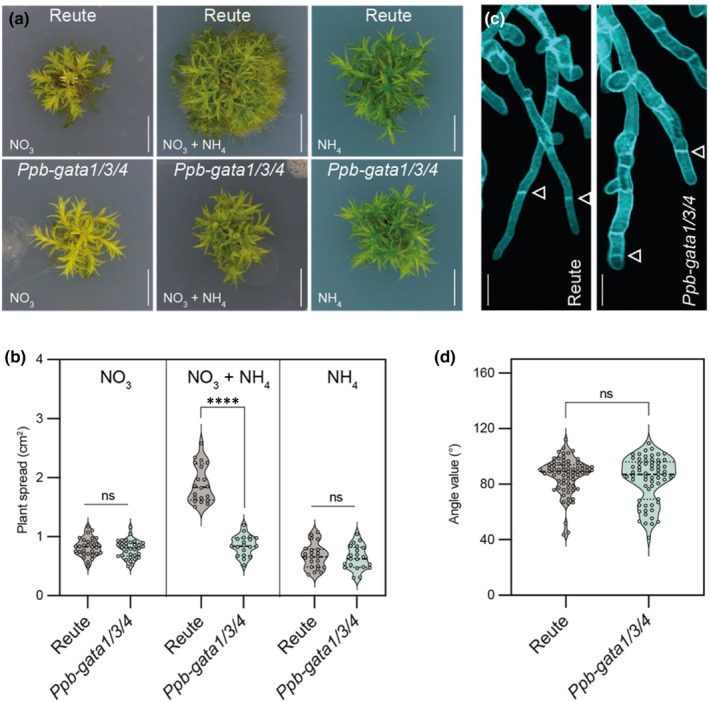 Fig. 3
Fig. 3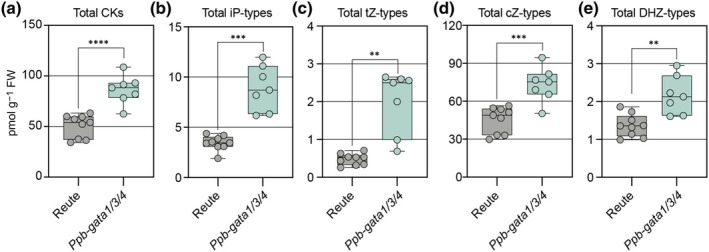 Fig. 4
Fig. 4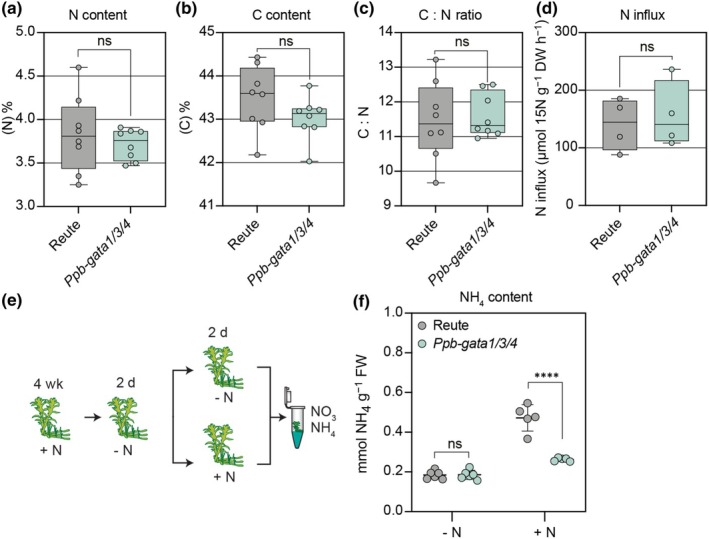 Fig. 5
Fig. 5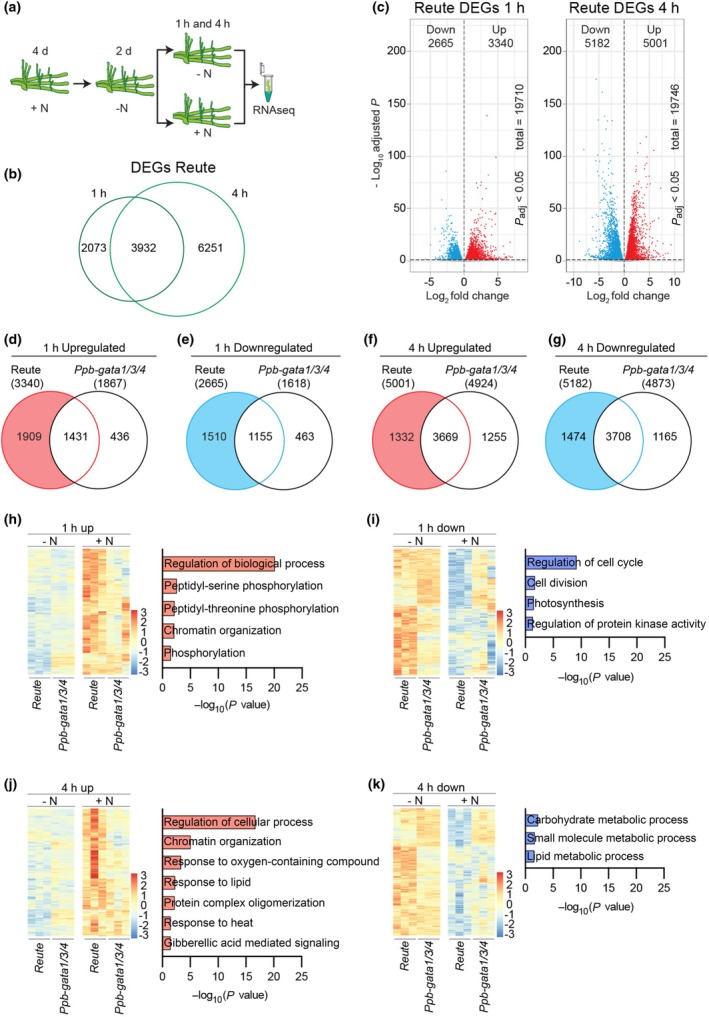 Fig. 6
Fig. 6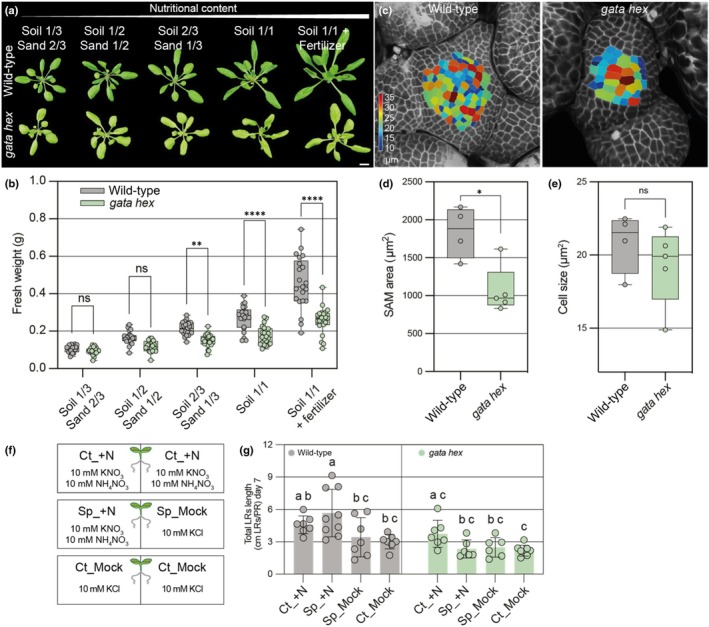 Fig. 7
Fig. 7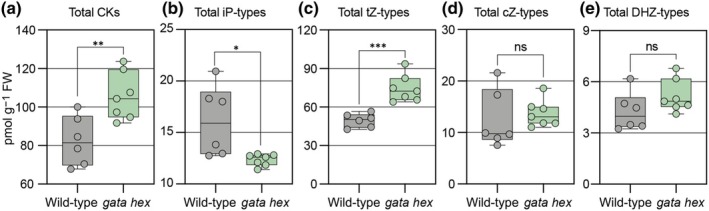 Fig. 8
Fig. 8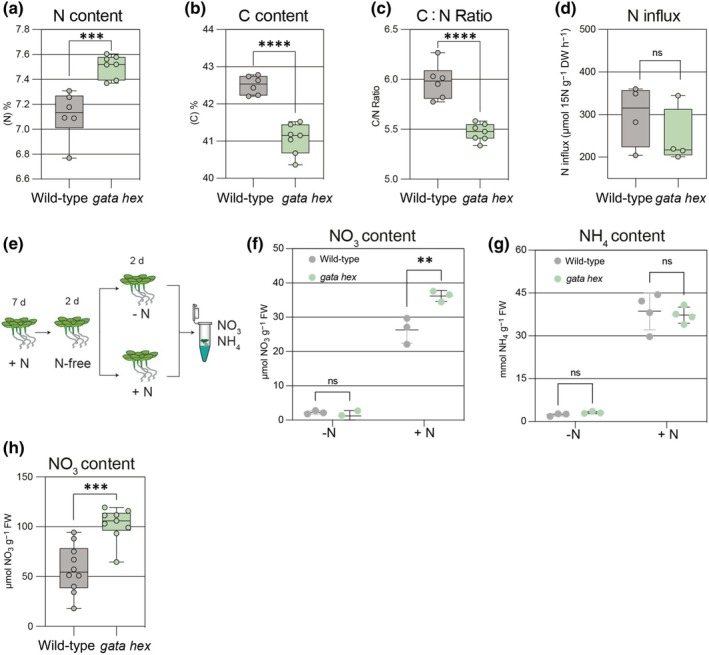 Fig. 9
Fig. 9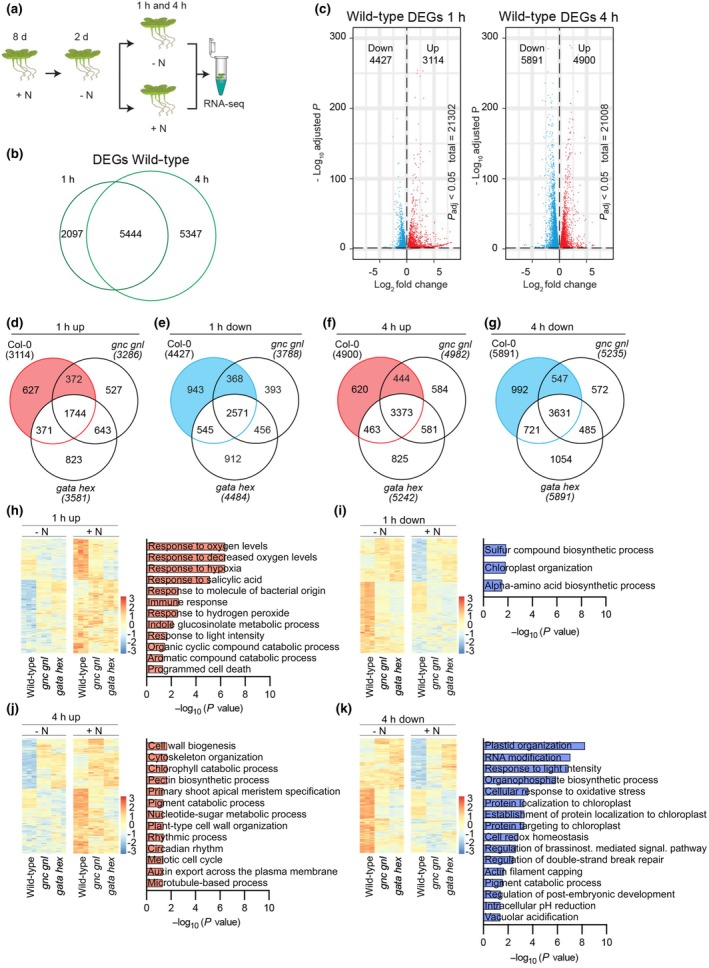 Fig. 10
Fig. 10- —Deutsche Forschungsgemeinschaft10.13039/501100001659
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Plant nutrient uptake and metabolism · Plant Gene Expression Analysis
Introduction
GATA transcription factors are evolutionarily conserved zinc finger proteins that preferentially bind to the DNA sequence G‐A‐T‐W (Lowry & Atchley, 2000). While animal and fungal genomes typically encode only a few GATA factors, for example six in humans and four in the yeast Saccharomyces cerevisiae, the GATA gene family is notably expanded in land plants, with close to 30 members in Arabidopsis thaliana (Schwechheimer et al., 2022). In vascular plants, GATA factors can be classified into four subgroups, classes A, B, C, and D, based on differences in their zinc finger DNA‐binding domains and the presence of additional protein domains at their N‐ or C‐termini (Schwechheimer et al., 2022).
In the angiosperm Arabidopsis, class B GATA factors are further subdivided into HAN‐ and LLM‐domain B‐GATAs (R. Richter et al., 2010; Behringer & Schwechheimer, 2015; Schwechheimer et al., 2022). The LLM‐domain is defined by a conserved C‐terminal leucine–leucine–methionine motif of unknown function that is essential for full GATA activity (Behringer et al., 2014; Schwechheimer et al., 2022). Among the six Arabidopsis LLM‐domain B‐GATAs, GNC (GATA, NITRATE‐INDUCIBLE, CARBON‐METABOLISM‐RELATED) and its paralog GNL (GNC‐LIKE) are the best studied. Their expression is regulated by the phytohormones gibberellin, cytokinin, and auxin, as well as by light (Naito et al., 2007; R. Richter et al., 2010, 2013). GNC and GNL, together with GATA15, GATA16, GATA17, and GATA17L, contribute to key physiological processes including chloroplast development, Chl and starch biosynthesis, as well as stomatal patterning (R. Richter et al., 2010; Chiang et al., 2012; Klermund et al., 2016; Ranftl et al., 2016; Bastakis et al., 2018; Zubo et al., 2018). HAN‐domain B‐GATAs contain a short, conserved domain of unknown function, initially characterized in the han (hanaba taranu) mutant (Zhao et al., 2004). Mutations in HAN cause floral abnormalities, while double and triple mutants of HAN, HAN‐LIKE1, and HAN‐LIKE2 exhibit severe embryonic defects (Zhao et al., 2004; Nawy et al., 2010; Whipple et al., 2010).
Interestingly, the HAN‐ and LLM‐based subdivision of angiosperm B‐GATAs does not apply to bryophytes. The liverwort Marchantia polymorpha and the moss Physcomitrium patens encode one and four B‐GATA genes, respectively, each harboring LLM‐ and HAN‐domains (Schwechheimer et al., 2022; Schroder et al., 2023). In Marchantia, the single B‐GATA, MpB‐GATA, promotes greening and protects against high‐light stress (Schroder et al., 2023). However, the biological roles of the four B‐GATAs from Physcomitrium remain largely uncharacterized (Schroder et al., 2023).
Nitrogen (N) is an essential macronutrient for plant growth, typically acquired as nitrate (NO_3_ ^−^) or ammonium (NH_4_ ^+^). The molecular and physiological responses to NO_3_ ^−^ have been extensively studied: its uptake is mediated by nitrate transporters (NRTs), primarily from the low‐affinity NRT1, in Arabidopsis NRT1.1/CHL1 and NRT1.2, and high‐affinity NRT2 families, in Arabidopsis NRT2.1 and NRT2.2 (Tsay et al., 1993; Filleur et al., 2001; Wang et al., 2018; Lamig et al., 2022). NRT1.1 functions as a dual‐affinity transporter for high‐ and low‐affinity uptake depending on its phosphorylation status (Liu & Tsay, 2003). NH_4_ ^+^ uptake is facilitated by ammonium transporters (AMTs) (Ninnemann et al., 1994; Williamson et al., 2024).
Within the plant, NO_3_ ^−^ is reduced to NO_2_ ^−^ by cytosolic nitrate reductase (NR), NIA1 and NIA2 in Arabidopsis, and further to NH_4_ ^+^ by chloroplastic nitrite reductase (NiR). The resulting NH_4_ ^+^ is assimilated into amino acids, Chl, and citric acid cycle intermediates via the GS/GOGAT cycle, involving glutamine synthetase (GS), glutamate synthase (GOGAT), and glutamate dehydrogenase (GDH) (x. Liu et al., 2022). N uptake triggers signaling events, including changes in cytoplasmic calcium levels, activation of calcium‐dependent protein kinases, and transcriptional responses, for example coordinated by NIN‐LIKE PROTEIN 7 (NLP7), which has been proposed to act as a nitrate sensor (Castaings et al., 2009; Marchive et al., 2013; Ristova et al., 2016; Ruffel et al., 2016; K. H. Liu et al., 2017, 2022; Varala et al., 2018; Brooks et al., 2019). Cytokinin contributes to N‐responsive growth by modulating these signaling pathways (Takei et al., 2002; Sakakibara et al., 2006; Landrein et al., 2018).
In fungi, GATA factors play pivotal roles in N regulation. In Aspergillus nidulans, the GATA factor AreA controls the expression of genes involved in N uptake and catabolism (Arst & Cove, 1973; Chudzicka‐Ormaniec et al., 2019). Under N‐rich conditions, AreA mediates N metabolite repression, suppressing enzymes and transporters for poor N sources (e.g. proline, allantoin, GABA) when superior N sources (e.g. glutamine, asparagine) are available (Caddick, 1994; Hofman‐Bang, 1999; Georis et al., 2009). AreA also regulates genes for N catabolism and NH_4_ ^+^ transport (Caddick, 1994; Monahan et al., 2006). AreB, another GATA factor in A. nidulans, represses formidase (fmdS) and genes involved in arginine catabolism under NH_4_ ^+^‐rich conditions (Chudzicka‐Ormaniec et al., 2019). In Saccharomyces cerevisiae, four GATA factors, Gln3, Nil1, Nil2, and Dal80, modulate N metabolite repression, with Gln3 and Nil1 acting as activators and Nil2 and Dal80 as repressors (Hofman‐Bang, 1999; Cooper, 2002; Magasanik, 2005; Georis et al., 2009).
Inspired by fungal studies, researchers have long sought links between plant GATAs and N metabolism. The Arabidopsis B‐class transcription factor GNC was initially identified as nitrate‐inducible, giving rise to its name GATA NITRATE‐INDUCIBLE CARBON‐METABOLISM‐RELATED (Scheible et al., 2004; Bi et al., 2005). GNC transcript levels increase 2 h after transferring seedlings from glutamine‐containing to NO_3_ ^−^‐containing media (Bi et al., 2005). Furthermore, GATA17L has been implicated in N response as a result of a comprehensive transcriptional network study (Ristova et al., 2016). However, a direct role of B‐GATAs in N‐regulated growth in Arabidopsis has not been conclusively demonstrated.
In Physcomitrium, growth begins with germination of spores into a filamentous protonema, composed of chloronemal and caulonemal cells, which later give rise to leafy gametophores. This stage is essential for nutrient uptake and surface colonization. While the influence of N on Physcomitrium growth is not well understood, prior studies have examined NO_3_ ^−^ transporter expression and genome‐wide transcriptional changes upon NH_4_ ^+^ supplementation, reporting repression of genes involved in NO_3_ ^−^ and NH_4_ ^+^ assimilation and NO_3_ ^−^ metabolism (Tsujimoto et al., 2007; Perroud et al., 2018).
Based on the important role of B‐GATA factors in plant growth and development in the angiosperm Arabidopsis and the bryophyte Marchantia, we speculated that B‐GATA genes may also have an important function in mosses. To examine this hypothesis, we investigated the Physcomitrium B‐GATA genes (PpB‐GATA1–PpB‐GATA4) through mutant analysis and revealed their critical role in N‐responsive growth. We found that mutants of the moss B‐GATA genes are defective in N‐regulated protonema growth and in N‐regulated gene expression. Building on these findings, we extended our analysis to Arabidopsis mutants of the B‐GATAs, where we also uncovered defects in N‐responsive growth and transcription regulation. Taken together, our results highlight an evolutionarily conserved function for B‐GATA transcription factors in plant growth in response to N availability.
Materials and Methods
Biological material
Physcomitrium patens (Hedw.) Mitt. ecotype Reute (Hiss et al., 2017) and Arabidopsis thaliana (L.) Heynh. ecotype Columbia (Col‐0) were used as wild‐type controls in all experiments. The following Arabidopsis mutants were previously characterized: gnc gnl (R. Richter et al., 2010) and gnc gnl gata15 gata16 gata17 gata17l (referred to as gata hex) (Schroder et al., 2023). Physcomitrium mutants were generated as part of this study, as described below.
Methods and primers used in this study are provided as supplemental data (Supporting Information Methods S1; Table S1).
Results
Physcomitrium patens encodes four B‐GATA transcription factors
In angiosperms, B‐class GATAs are divided into those containing either a HAN‐ or LLM‐domain. By contrast, the single B‐GATA from the liverwort Marchantia and all four B‐GATAs from the moss Physcomitrium possess both HAN‐ and LLM‐domains, suggesting that the ancestral land plant B‐GATA may have harbored both domains (Figs 1, S1) (Schwechheimer et al., 2022; Schroder et al., 2023). Phylogenetic analysis shows that the four Physcomitrium B‐GATAs, which we have designated PpB‐GATA1 to PpB‐GATA4, are more closely related to each other than to B‐GATAs from Marchantia or Arabidopsis, indicating that this gene family expanded after the divergence of mosses from the last common ancestor of mosses and liverworts (Fig. S1).
Sequence comparison of B‐GATA factors from Physcomitrium patens (Pp), Marchantia polymorpha (Mp), and Arabidopsis thaliana (At). Alignment of the conserved HAN‐domain, zinc finger domain, and LLM‐domain from selected B‐GATA transcription factors of Physcomitrium patens (Pp), Marchantia polymorpha (Mp), and (At). Residue similarities and identities are color‐coded according to the scheme shown in the figure legend.
Gene expression data from public repositories revealed that all PpB‐GATAs are broadly expressed across diverse tissues. Among them, PpB‐GATA1 and PpB‐GATA3 showed the widest and highest expression, particularly in vegetative tissues such as chloronema, caulonema, gametophores, and rhizoids (Fig. S2) (Winter et al., 2007; Ortiz‐Ramirez et al., 2016). PpB‐GATA2 was most strongly expressed in chloronema and rhizoids, with lower levels in caulonema and gametophores, whereas PpB‐GATA4 exhibited strong expression primarily in mature sporophytes and minimal expression in vegetative tissues (Fig. S2) (Winter et al., 2007; Ortiz‐Ramirez et al., 2016). These data point to PpB‐GATA1 and PpB‐GATA3 as the most broadly and strongly expressed genes within this family.
Generation and molecular characterization of Ppb‐gata mutants
To investigate the biological roles of the PpB‐GATAs, we employed CRISPR/Cas9‐based genome editing to target the four genes (Fig. S3). Following transformation, mutagenesis, and PCR‐based genotyping, we isolated single, double, and triple mutants of PpB‐GATA1, PpB‐GATA3, and PpB‐GATA4. These included single mutant alleles (Ppb‐gata1‐1, Ppb‐gata3‐1, Ppb‐gata4‐1, Ppb‐gata4‐2), the double mutant Ppb‐gata3‐2/4‐3, and the two triple mutants Ppb‐gata1‐1/3‐2/4‐3 and Ppb‐gata1‐3/3‐2/4‐3, all of which harbored frameshift‐inducing deletions or insertions and were consequently classified as loss‐of‐function mutants (Fig. S3). An exception was the Ppb‐gata1‐2 allele in the Ppb‐gata1‐2/4‐2 double mutant, which contained a 15 bp duplication resulting in an in‐frame insertion of six nonnative amino acids (Q‐S‐T‐T‐Q‐C) at the edited site (Fig. S3).
Whole‐genome sequencing confirmed the mutations and allowed assessment of background variation (Table S2). The Ppb‐gata1‐1 mutant exhibited several polymorphisms in noncoding regions, with two missense mutations found in the genes Pp3c6_23550, encoding a pentatricopeptide repeat protein, and Pp3c10_15160, encoding a CORYNE‐related receptor‐like kinase gene. Due to the lack of an alternative Ppb‐gata1 allele, we proceeded with this line despite these background mutations. In the originally analyzed Ppb‐gata4‐1 allele, sequencing also revealed a possible but ultimately unresolvable Ppb‐gata3 mutation, prompting us to use the alternative allele Ppb‐gata4‐2 for further experiments. Among the triple mutants, Ppb‐gata1‐1/3‐2/4‐3 harbored 23 insertions or deletions and 15 single nucleotide polymorphisms, whereas Ppb‐gata1‐3/3‐2/4‐3 had no detectable background mutations and was therefore selected for further analysis (Table S2). For clarity, the allele nomenclature in the remaining manuscript is reduced to the gene mutant names, omitting the allele specifications.
Despite multiple attempts and extensive genotyping, we were unable to obtain Ppb‐gata2 mutants by CRISPR/Cas9 mutagenesis or by homologous recombination.
PpB‐GATAs
are required for protonema growth and pigment biosynthesis
All Ppb‐gata mutants exhibited a pronounced reduction in plant spread when grown on medium containing both NH_4_ ^+^ and NO_3_ ^−^ as N sources (Fig. 2a,b). This phenotype was attributed to impaired protonema growth, which also led to a less circular plant morphology, reflected in a decreased circularity index (Fig. 2).
Characterization of Physcomitium patens Ppb‐gata mutant phenotypes. (a) Representative images of 4‐wk‐old Physcomitrium plants of the indicated genotypes. Bar, 1 mm. (b, c) Quantification of plant morphology based on the images shown in (a). (b) Spread area of the plants. (c) Circularity index as a measure of shape compactness. Each data point represents an independent biological replicate; horizontal black lines indicate the median, and dotted lines mark the first (Q1) and third (Q3) quartiles. Different letters indicate statistically significant differences determined by Brown–Forsythe and Welch ANOVA followed by Dunnett's T3 multiple comparisons test (b) or by one‐way ANOVA with Tukey's multiple comparisons test (c). (d, e) Quantification of photosynthetic pigments in the indicated genotypes: (d) total Chl content (Chla + Chlb); (e) total carotenoid content, both relative to the concentrations detected in the Reute wild‐type. Each data point represents an independent biological replicate; bars show mean ± SD. Different letters indicate statistically significant differences as determined by one‐way ANOVA followed by Tukey's multiple comparisons test.
Given that mutants of B‐GATAs in Arabidopsis and Marchantia show defects in Chl and carotenoid biosynthesis, we measured pigment levels in the Physcomitrium mutants (Ranftl et al., 2016; Bastakis et al., 2018; Schroder et al., 2023). The Ppb‐gata1/3/4 triple mutant showed a significant reduction in both Chl and carotenoids compared with the Reute wild‐type strain, while the other mutants had levels comparable to wild‐type (Fig 2d,e). Our results indicate that the Physcomitrium B‐GATAs defective in the Ppb‐gata1/3/4 triple mutant, like their orthologs from Arabidopsis and Marchantia, are important for proper Chl and carotenoid biosynthesis.
Ppb‐gata mutants are defective in protonema formation
The reduced size of Ppb‐gata mutants was primarily due to a strong impairment in protonema formation compared with the wild‐type (Fig. 2a). In Physcomitrium, protonema development is induced by the combined presence of nitrate (NO_3_ ^−^) and ammonium (NH_4_ ^+^) as N sources (Perroud et al., 2018). When grown on media supplemented with NO_3_ ^−^ and NH_4_ ^+^, wild‐type plants exhibited increased plant spread and protonema formation, whereas these responses were absent in the Ppb‐gata1/3/4 triple mutant (Fig. 3a,b). As reported previously, the presence of only NO_3_ ^−^ or NH_4_ ^+^ as a single N source was insufficient to induce protonema development in either wild‐type or mutant plants (Perroud et al., 2018).
*Loss of B‐GATAs influences N‐dependent growth responses in Physcomitrium patens. (a) Representative images of 4‐wk‐old Physcomitrium Reute (wild‐type) and Ppb‐gata1/3/4 triple mutant plants grown on media containing the indicated N sources. Bar, 5 mm. (b) Quantification of plant spread area from the plants shown in (a). Each data point represents an independent biological replicate; horizontal black lines indicate the median, and dotted lines mark the first (Q1) and third (Q3) quartiles. Unpaired t‐test, with Welch's correction applied when variances were significantly different: ***, P < 0.0001; ns, not significant. (c) Representative confocal images of calcofluor white–stained protonemal filaments. Arrows mark the division planes used for angle measurements in (d). Bar, 50 μm. (d) Quantification of cell division plane angles from the cells indicated in (c). Each data point represents an independent biological replicate; horizontal black lines indicate the median, and dotted lines mark the first (Q1) and third (Q3) quartiles. Unpaired t‐test: ns, not significant.
Protonema tissue consists of two cell types: chloronema and caulonema, which can be distinguished by the angles between adjoining cell walls (Coudert et al., 2019). Analysis of cell wall angles revealed no significant differences in chloronema and caulonema proportions between wild‐type and the Ppb‐gata1/3/4 triple mutant (Fig. 3c–d). We therefore conclude that the triple mutant is specifically impaired in N‐induced protonema formation but not in the differentiation of the chloronema and caulonema cell types.
PpB‐GATA1
overexpression produces opposite phenotypes to loss‐of‐function mutants
To further assess the role of PpB‐GATAs, we generated overexpression lines of PpB‐GATA1, one of the highly expressed family members (Fig. S4). In three independent lines, RT‐qPCR confirmed a 2‐ to 7‐fold increase in PpB‐GATA1 expression (Fig. S4a). These overexpression lines exhibited significantly increased levels of Chl and carotenoids, which is in contrast to the reduced pigment content of the Ppb‐gata1/3/4 triple mutant (Fig. S4b,c).
Interestingly, all overexpressors were smaller than the wild‐type, regardless of whether plants were grown on NO_3_ ^−^ and NH_4_ ^+^ or on NO_3_ ^−^ alone. Moreover, the extent of size reduction correlated with transgene expression levels (Fig. S4d,e). Overexpression lines also showed increased circularity under both N regimes, a parameter that correlates with enhanced protonema development (Fig. S4f–i). The increased protonemal circularity observed even in NO_3_ ^−^‐only conditions suggests that PpB‐GATAs can promote protonema development independently of NH_4_ ^+^ availability.
Ppb‐gata mutants accumulate cytokinin
A previous study showed that elevated cytokinin levels suppress plant spread in Physcomitrium (Coudert et al., 2019). To examine whether the Ppb‐gata1/3/4 triple mutant phenotype could potentially be linked to cytokinin biosynthesis, we quantified cytokinin levels in wild‐type and mutant moss. This quantification revealed significantly higher cytokinin levels in the Ppb‐gata1/3/4 triple mutant compared with the wild‐type, and consequently, increased cytokinin levels may contribute to the growth defect observed in the mutants (Fig. 4; Table S3).
*Loss of B‐GATAs influences endogenous cytokinin levels in Physcomitrium patens. (a–e) Quantification of cytokinin (CK) metabolites in 4‐wk‐old Physcomitrium Reute (wild‐type) and Ppb‐gata1/3/4 triple mutant plants. Each graph represents the concentration of a specific cytokinin form or metabolite as indicated. iP, isopentenyladenine; tZ, trans‐zeatin; cZ, cis‐zeatin; DHZ, dihydrozeatin. Each data point represents an independent biological replicate (n = 3–5); the whiskers indicate the full data range (minimum to maximum), boxes show the interquartile range (IQR; spanning the 25th to 75th percentiles), and the central line indicates the median value of the data distribution. Unpaired t‐test, with Welch's correction applied when variances were significantly different: **, P < 0.01; ***, P < 0.001; ***, P < 0.0001.
Nitrogen metabolism and uptake are not impaired in the Ppb‐gata triple mutant
To determine whether the reduced growth in the Ppb‐gata1/3/4 mutant was due to altered N metabolism or a disrupted C : N (carbon‐to‐nitrogen) balance, we conducted elemental analysis on 4‐wk‐old plants grown on standard media (10 mM KNO_3_, 5 mM (NH_4_)2_C_4_H_4_O_6). No significant differences in total N or C content, nor in their ratio, were detected between wild‐type and mutant plants, suggesting that N and C metabolism were not impaired in the mutant tissue analyzed (Fig. 5a–c).
*Loss of B‐GATA transcription factors does not affect endogenous nitrogen (N) and carbon (C) levels in Physcomitrium patens. (a–c) Quantification of total N (a), total C (b), and the C : N ratio (C) in 6‐d‐old Physcomitrium wild‐type and Ppb‐gata1/3/4 mutant protonemata grown on N‐containing medium. Each data point represents an independent biological replicate (n = 3–5 colonies); the whiskers indicate the full data range (minimum to maximum), boxes show the interquartile range (IQR; spanning the 25th to 75th percentiles), and the central line indicates the median value of the data distribution. Unpaired t‐test: ns, not significant. (d) N influx rates measured in 13‐d‐old wild‐type and Ppb‐gata1/3/4 mutant plants following a 12 min exposure with 10 mM 15N‐labeled KNO3 and 15N‐labeled NH4 +. Each data point represents an independent biological replicate (n = 4 colonies); the whiskers indicate the full data range (minimum to maximum), boxes show the interquartile range (IQR; spanning the 25th–75th percentiles), and the central line indicates the median value of the data distribution. Unpaired t‐test: ns, not significant. (e) Schematic overview of the N metabolism experiment. Plants were first N‐starved for 48 h, then transferred to either N‐containing medium (+N; 10 mM KNO3 and 5 mM (NH4)2C4H4O6) or a mock medium lacking nitrogen (‐N; 10 mM KCl). (f) Ammonium (NH4 +) concentrations in tissue extracts measured after N resupply. Each data point represents an independent biological replicate (n = 4 colonies); bars indicate mean ± SD. Two‐way ANOVA followed by Šídák's multiple comparisons test; ***, P < 0.0001; ns, not significant. A corresponding measurement of tissue nitrate (NO3 −) content yielded levels below the detection limit.
We next tested whether N uptake capacity differed between genotypes. Plants were N‐starved for 2 d and then exposed to ^15^N‐labeled NH_4_NO_3_ for 12 min. Isotope analysis revealed no significant differences in ^15^N incorporation between wild‐type and mutant plants, indicating that N uptake was not impaired in the Ppb‐gata1/3/4 mutant in the time frame observed in the study (Fig. 5d).
To examine internal N metabolism, plants starved of N for 2 d were transferred either to N‐containing media (10 mM KNO_3_, 5 mM (NH_4_)2_C_4_H_4_O_6) or to N‐free control media. After 2 d, NO_3_ ^−^ levels were below the detection limit in both genotypes, suggesting rapid NO_3_ ^−^ assimilation. By contrast, NH_4_ ^+^ levels were significantly lower in the Ppb‐gata1/3/4 mutant compared with the wild‐type (Fig. 5e,f). Since our N uptake study had shown that N uptake may not be affected in the mutant, the reduced NH_4_ ^+^ content in the mutant likely reflects increased NH_4_ ^+^ assimilation or altered N metabolism downstream of uptake.
Ppb‐gata mutants display severely impaired transcriptional responses to N supply
To assess whether the Ppb‐gata1/3/4 exhibited altered transcriptional responses to N availability, we performed RNA‐seq analysis on 6‐d‐old wild‐type and Ppb‐gata1/3/4 mutant plants. To this end, plants were first grown on N‐containing medium (10 mM KNO_3_, 5 mM (NH_4_)2_C_4_H_4_O_6) for 2 d, then transferred to N‐free medium for 2 d, followed by re‐exposure to either the N‐containing medium (10 mM KNO_3_, 5 mM (NH_4_)2_C_4_H_4_O_6) or 10 mM KCl mock control medium for 1 or 4 h, taking into account the presence of 10 mM potassium (K) in the N‐containing medium (Fig. 6a).
Transcriptomic characterization of the N response in Physcomitrium patens and the Ppb‐gata1/3/4 mutant. (a) Schematic overview of the RNA‐seq experiment. Wild‐type and Ppb‐gata1/3/4 mutant protonemata were pre‐cultured on N‐containing medium, starved of N for 2 d, and subsequently transferred to either N‐containing (+N; 10 mM KNO3 and 5 mM (NH4)2C4H4O6) or N‐free mock (–N; 10 mM KCl) medium for 1 or 4 h before RNA extraction. (b) Venn diagram showing overlap of differentially expressed genes (DEGs) between +N and –N treatments in wild‐type Physcomitrium (Reute strain) at the 1‐ and 4‐h time points. (c) Volcano plots illustrating the distribution of DEGs in the wild‐type at the 1‐ and 4‐h time points, highlighting genes significantly upregulated or downregulated in response to N. The total number of variables (genes) is provided. (d–g) Venn diagrams comparing DEGs between the wild‐type and the Ppb‐gata1/3/4 mutant under +N vs –N conditions, separated into upregulated (d, f) and downregulated (e, g) gene sets at 1 h (d, e) and 4 h (f, g). Color‐coded sections indicate DEGs exclusive to the wild‐type, which were used for subsequent GO enrichment analysis. (h–k) Heatmaps and corresponding gene ontology (GO) enrichment analyses of DEGs that were regulated in response to N in the wild‐type but not in the Ppb‐gata1/3/4 mutant, potentially representing PpB‐GATA‐dependent transcripts. Panels show upregulated (h, j) and downregulated (i, k) gene sets at 1 h (h, i) and 4 h (j, k).
Principal component analysis (PCA) of samples from both time points revealed that N treatment had a strong effect on global gene expression, whereas the variation between genotypes was comparatively smaller (Fig. S5). One Ppb‐gata1/3/4 replicate from the 1‐h time point showed a distinct clustering pattern compared with the other two replicates. However, neither the 1.5 interquartile range (IQR) rule nor Mahalanobis distance analysis (χ^2^(2), P < 0.01) identified this replicate as a statistical outlier, and it was therefore retained for downstream analyses (Mahalanobis, 1936; Tukey, 1977).
In the wild‐type, DESeq2 identified 6005 differentially expressed genes (DEGs) after 1 h of N treatment and 10 183 DEGs after 4 h (Fig. 6b,c; Table S4). Of these, 3932 DEGs were differentially regulated at both time points, indicating a sustained transcriptional response to N availability. Gene ontology (GO) analysis showed that N‐regulated DEGs were enriched for GO terms related to biological regulation, biosynthetic processes, and cell cycle control, likely reflecting the stimulatory effect of N on cellular metabolism and growth in Physcomitrium (Fig. S6; Table S5).
By contrast, the Ppb‐gata1/3/4 mutant failed to regulate a substantial portion of these N‐responsive DEGs. Specifically, 1909 (57%) of the 3340 upregulated DEGs and 1510 (57%) of the 2665 downregulated DEGs identified in the wild‐type after 1 h of N treatment were not differentially expressed in the mutant (Fig. 6d,e; Table S4c). At the 4‐h time point, 1332 (27%) of the 5001 upregulated DEGs and 1474 (28%) of the 5182 downregulated DEGs in the wild‐type were not responsive in the mutant. These results indicate that N‐responsive transcriptional responses are strongly dependent on the presence of the PpB‐GATA genes.
GO enrichment of the PpB‐GATA–dependent gene sets further revealed enrichment in categories such as regulation of biological processes, cellular function, cell cycle progression, and cell division (Fig. 6h–k; Table S6). Notably, orthologs of the Arabidopsis ethylene receptor genes ETR1 (Pp6c14_10160; orthologous to AT1G66340) and ETR2 (Pp6c9_9400; AT3G23150), both two‐component sensor histidine kinases, were upregulated by N treatment in the wild‐type but unresponsive in the Ppb‐gata1/3/4 mutant. Similarly, 10 Ca^2+^‐dependent protein kinase genes, several AUXIN RESPONSE FACTORs (ARFs), including Pp6c14_9370 (AT2G46530, ARF11), Pp6c4_6680 (AT1G19850, ARF5), Pp6c5_4850 (AT1G59750, ARF1), and Pp6c2_14160 (AT1G19220, ARF19), as well as three SWI/SNF chromatin remodeling factors were differentially expressed in response to N in the wild‐type, but not in the mutant. Finally, the N‐regulation of six XYLOGLUCAN ENDOTRANSGLYCOSYLASE (XET) genes, genes previously linked to protonema differentiation following N treatment (Perroud et al., 2018), was disrupted in the Ppb‐gata1/3/4 mutant, further supporting a critical role for B‐GATA transcription factors in mediating N‐responsive developmental transitions.
To examine the transcription regulation of genes with a proposed role in N uptake and transport, metabolism, and relevant transcriptional responses, we generated a list of Physcomitrium orthologues using a combination of Orthofinder and Diamond BLAST of genes with a role in these processes that had been previously identified in Arabidopsis (Fig. S7; Table S7a) (Vidal et al., 2020). Among the orthologous genes encoding transporter genes, we detected enhanced N regulation of the high‐affinity nitrate transporter orthologue PpNRT2.1 and a loss of N regulation of the ammonium transporter orthologue PpAMT1 and the nitrate transporter orthologues PpNPF6.2, PpNPF8.1/8.4, and PpNPF8.3 (Fig. S7a). Among the N metabolism genes, we detected a loss or weakened N regulation in the Ppb‐gata1/3/4 mutant of the amino acid transporter orthologue PpAAT1 and the glutamate decarboxylase PpGAD3, but stronger N regulation of two orthologues of the nitrate reductase PpNIA1 and of the glutamine synthetases PpGS2 and PpGSR2 (Fig. S7b). Several genes orthologous to transcription factors with a demonstrated or proposed role in N responses showed differential expression between the wild‐type and the mutant, including PpCDF1, PpLBD37/PpLBD38, PpNLP6, PpNLP7, as well as PpTCP20 (Fig. S7c). The MKN2 (KNOX2) gene, Pp6c25_4720, previously associated with a phenotype similar to that observed in Ppb‐gata1/3/4, was not differentially expressed in the mutant, indicating that its misexpression is unlikely to account for the GATA gene mutant phenotype (Coudert et al., 2019).
Finally, we analyzed whether PpB‐GATA gene expression was regulated by N under our experimental conditions (Fig. S7d). Specifically, at the 4 h time point, we found that three PpB‐GATA genes were upregulated in response to the N treatment, while PpB‐GATA4 expression was downregulated. Accordingly, PpB‐GATA1, PpB‐GATA2, and PpB‐GATA3 behave like Arabidopsis GNC, and all four PpB‐GATAs could be classified as N‐regulated B‐GATA genes.
Expression of biosynthesis genes may explain the Chl and cytokinin phenotypes of Ppb‐gata1/3/4
To understand whether the differences in Chl accumulation could be explained through the available RNA‐seq data, we generated a list of Physcomitrium genes orthologous to genes that had been implicated in Chl and cytokinin biosynthesis in Arabidopsis using OrthoFinder and/or Diamond BLAST (Table S7b,c) (Emms & Kelly, 2019).
With regard to the expression of Chl biosynthesis genes, we identified many genes contributing to Chl biosynthesis, for example genes orthologous to the heme biosynthesis genes HEME, HEMEF, and HEMEG, as well as MAGNESIUM CHELATASE SUBUNIT I (CHLI) and PROTOCHLOROPHYLLIDE OXIDOREDUCTASE A (PORA) as being downregulated in Ppb‐gata1/3/4 in comparison with the Reute wild‐type (Fig. S8). The decreased expression of these genes may explain the reduced Chl biosynthesis observed in the mutants.
To establish a link to cytokinin biology, in view of the altered cytokinin levels observed in the mutants, we retrieved a list of genes with a proposed role in cytokinin biosynthesis (trans‐zeatin; tZ) from Physcomitrium from the KEGG database, and identified orthologues of genes related to cytokinin signaling in the literature and using a combination of Orthofinder and/or Diamond BLAST (Table S7c) (Pils & Heyl, 2009; Kieber & Schaller, 2014; Hata et al., 2025). Several genes encoding for isopentenyltransferases (IPT), the LONELY GUY (LOG) cytokinin nucleoside 5′‐monophosphate phosphoribohydrolases, and the catabolic enzyme cytokinin oxidase/dehydrogenase (CKX) were more strongly expressed in Ppb‐gata1/3/4 in comparison with the wild‐type, providing a possible explanation for the abundance of cytokinin in Ppb‐gata1/3/4 (Fig. S9). We further noted the strong upregulation of two genes encoding for cytokinin receptors, PpCHK2 (Pp6c16_5950) and PpCHK3 (Pp6c6_3870).
Arabidopsis mutants of LLM‐domain B‐GATAs exhibit reduced growth responses to nitrogen supply
The observation of a differential N response in Physcomitrium prompted us to investigate whether Arabidopsis mutants of B‐GATA genes also show altered growth responses to N availability. To this end, we compared the wild‐type with two previously characterized mutant lines: gnc gnl, which carries mutations in the LLM‐domain B‐GATA genes GNC and GNL, and gata hex, which is deficient in all six LLM‐domain B‐GATA genes, including GNC and GNL (R. Richter et al., 2010; Schroder et al., 2023). Plants were grown on mixtures of N‐poor sand and N‐rich soil, following established protocols (S. Richter et al., 2010; Landrein et al., 2018; Schroder et al., 2023). To prevent N availability from affecting germination, seeds were first germinated in full‐strength soil and grown for 1 wk before being transferred to the respective soil/sand mixtures for an additional 4 wk. In these experiments, gata hex mutants displayed visibly reduced rosette diameters when grown on soil alone or on fertilized soil, whereas these differences were not apparent on sand‐enriched substrates (Figs 7a, S10a,b). Fresh weight measurements after 4 wk revealed significant growth reduction in gata hex on 2 : 1 soil‐to‐sand mixtures, with a milder reduction also observed in gnc gnl mutants (Figs 7b, S10a,b). The pronounced increase in fresh weight across all genotypes following fertilization suggests that N, or potentially other macro‐ or micronutrients present in the fertilizer, is a limiting factor for growth in our standard soil conditions.
*Loss of LLM‐domain B‐GATAs impairs N‐dependent growth responses in Arabidopsis thaliana. (a) Representative images of 4‐wk‐old Arabidopsis wild‐type (Col‐0) and gata hexuple (gata hex) mutant plants, grown under varying soil/sand mixtures or soil supplemented with fertilizer. Bar, 1 cm. (b) Quantification of fresh weight in Arabidopsis plants as shown in (a). Each data point represents an independent biological replicate (one entire rosette); the whiskers indicate the full data range (minimum to maximum), boxes show the interquartile range (IQR; spanning the 25th–75th percentiles), and the central line indicates the median value of the data distribution. Two‐way ANOVA comparisons test, followed by Šídák's multiple comparisons performed for each soil and sand combination. **, P < 0.01; ****, P < 0.0001, ns, not significant. (c) Representative confocal images of shoot apical meristems (SAMs) from 4‐wk‐old soil‐grown plants, reconstructed using Morphographics X. (d, e) Quantification of SAM surface area (d) and average cell size (e) from the samples shown in (c). Each data point represents an independent biological replicate (one inflorescence meristem); the whiskers indicate the full data range (minimum to maximum), boxes show the interquartile range (IQR; spanning the 25th–75th percentiles), and the central line indicates the median value of the data distribution. Unpaired t‐test: , P < 0.05; ns, not significant. (f) Schematic representation of the split‐root setup used for the analysis shown in (g). Roots were subjected to three conditions: Ct_ +N (control + nitrogen) and Ct_Mock (control mock), where both sides of the root system were exposed to either + N or mock treatment, respectively; and Split_ + N/Sp_Mock, where one side was supplied with N and the other with mock (KCl). (g) Total lateral root (LR) growth in wild‐type Col‐0 and gata hex under the conditions described in (f). Total LR length was normalized to the primary root (PR) length. Each point represents an independent biological replicate; bars indicate mean ± SD. Different letters denote statistically significant differences (two‐way ANOVA followed by Tukey's multiple comparisons test).
To further assess the impact of B‐GATA deficiency on N‐responsive growth, we examined shoot apical meristem (SAM) morphology in soil‐grown wild‐type and gata hex plants. SAM size, but not cell size, was significantly reduced in the mutant, supporting the notion that N supply cannot be effectively translated into enhanced plant vigor in the absence of functional B‐GATA transcription factors (Fig. 7c–e).
To investigate the role of B‐GATAs in N‐related systemic responses, we performed a split‐root assay (Figs 7f, S10c) (Ruffel et al., 2011). This approach enables the two root systems of a single plant to be exposed to distinct nutritional regimes, thereby generating heterogeneous nitrogen conditions. Root architecture was analyzed 7 d after transfer under three conditions: (Ct_ +N) a homogeneous N‐rich environment with both sides containing 10 mM KNO_3_ + 10 mM NH_4_NO_3_, (Ct_Mock) a homogeneous N‐depleted environment with both sides containing 10 mM KCl, and (Sp_ +N/Sp_Mock) a heterogeneous split environment with one side containing 10 mM KNO_3_ + 10 mM NH_4_NO_3_ and the other side containing 10 mM KCl. In the wild‐type plants, we observed a compensatory lateral root growth in the heterogeneous condition, with preferential lateral root elongation on the nitrogen‐supplemented side (Sp_ +N) compared with the nitrogen‐free side (Sp_Mock) (Figs 7g, S10d). This compensatory response was severely compromised in the gata hex mutant but still preserved in gnc gnl (Figs 7g, S10d,e). In contrast to previous reports, lateral root growth in N‐depleted conditions (Ct_Mock) did not surpass growth under N‐sufficient conditions (Ct_ +N) in the wild‐type in our conditions, which may be explained by the fact that we used different N treatments for our experiment than those used in the previous report to stay in the context of the other experimental setups used for our studies (Fig. 7g).
Arabidopsis gata hex mutants have elevated cytokinin levels
Vegetative growth in response to N is mediated by cytokinin biosynthesis and signaling (Landrein et al., 2018). We therefore also quantified cytokinin levels in 4‐wk‐old rosettes of wild‐type and gata hex mutants. The analysis revealed a significant increase in trans‐zeatin (tZ)‐type cytokinins in gata hex, along with comparatively minor decreases in isopentenyladenine (iP)‐type cytokinins (Fig. 8a–e). As tZ‐type cytokinins contribute more substantially to the total cytokinin content, this shift resulted in an overall increase of total cytokinins in gata hex when compared with the wild‐type (Fig. 8a). Despite these elevated cytokinin levels, the gata hex mutants had reduced shoot growth and a smaller shoot apical meristem (SAM) area (Fig. 7). This indicates that the growth defect is not due to a deficiency in cytokinin synthesis but rather suggests that the lack of the B‐GATA may disrupt the plant's ability to respond to cytokinin signaling. Further, since the systemic N response in the split‐root assay is cytokinin‐dependent, but gata hex exhibits reduced lateral root proliferation despite increased cytokinin levels (Fig. 7g), the Arabidopsis B‐GATAs may function as mediators downstream from cytokinins in systemic N signaling.
*Loss of LLM‐domain B‐GATA transcription factors alters endogenous cytokinin levels in Arabidopsis thaliana. (a–e) Quantification of cytokinin (CK) metabolites in 4‐wk‐old Arabidopsis wild‐type (Col‐0) and gata hexuple (gata hex) mutant plants. iP, isopentenyladenine; tZ, trans‐zeatin; cZ, cis‐zeatin; DHZ, dihydrozeatin. Each data point represents an independent biological replicate (n = 2/3 rosettes); the whiskers indicate the full data range (minimum to maximum), boxes show the interquartile range (IQR; spanning the 25th–75th percentiles), and the central line indicates the median value of the data distribution. Unpaired t‐test, with Welch's correction applied when variances were significantly different: *, P < 0.05; **, P < 0.01; **, P < 0.001; ns, not significant.
Arabidopsis gata hex mutants have increased nitrogen levels
To examine whether the Arabidopsis B‐GATA genes contributed to N metabolism, we quantified the overall N and C content in Arabidopsis seedlings following a two‐day period of N starvation after having been grown for 3 d in plates containing 10 mM NH_4_NO_3_ and 10 mM KNO_3_. In this experiment, we found that the gata hex mutant exhibited higher N content than the wild‐type, coupled with a reduced carbon content, resulting in a reduced C/N (carbon/nitrogen) ratio (Fig. 9a–c). To examine whether gata hex mutants are defective in N uptake, we performed an N influx assay with gata hex and the wild‐type (David et al., 2019). To this end, we transferred 2‐wk‐old plants to liquid medium lacking N for 2 d before transferring them to a medium containing N (10 mM NH_4_NO_3_ and 10 mM KNO_3_) for 4 h, followed by an additional transfer for a 10‐min treatment to ^15^N‐labeled N (10 mM NH_4_NO_3_ and 10 mM KNO_3_). Isotope analysis following this treatment did, however, not reveal statistically significant changes in N influx under these conditions (Fig. 9d).
*Loss of LLM‐domain B‐GATAs affects endogenous N and C levels as well as NO3 uptake in Arabidopsis thaliana. (a–c) Total nitrogen (N), carbon (C) content, and C : N ratio in 11‐d‐old Arabidopsis wild‐type and gata hexuple (gata hex) mutant seedlings. Each data point represents an independent biological replicate (n = 9/10 seedlings); the whiskers indicate the full data range (minimum to maximum), boxes show the interquartile range (IQR; spanning the 25th–75th percentiles), and the central line indicates the median value of the data distribution. Unpaired t‐test; ***, P < 0.001; ****, P < 0.0001. (d) Nitrate influx in 13‐d‐old seedlings after a 10 min exposure to 10 mM K15NO3 + 10 mM 15NO3 15NH4. Each data point represents an independent biological replicate (n = 12/15 seedlings); the whiskers indicate the full data range (minimum to maximum), boxes show the interquartile range (IQR; spanning the 25th–75th percentiles), and the central line indicates the median value of the data distribution. Unpaired t‐test: ns, not significant. (e) Schematic overview of the experimental setup used to determine NO3 and NH4 as shown in (f) and (g). (f, g) NO3 − (f) and NH4
- (g) content in seedlings 2 d after transfer from N‐free (–N) to N‐containing (+N; 10 mM KNO3 + 10 mM NO3NH4) medium, or a corresponding mock treatment (–N; 10 mM KCl). Each data point represents an independent biological replicate (n = 10/12 seedlings); bars indicate mean ± SD. Two‐way ANOVA followed by Šídák's multiple comparisons test; **, P < 0.01; ns, not significant. (h) Nitrate content in 4‐wk‐old rosettes of the indicated genotypes. Each data point represents an independent biological replicate (n = 2 rosettes); the whiskers indicate the full data range (minimum to maximum), boxes show the interquartile range (IQR; spanning the 25th–75th percentiles), and the central line indicates the median value of the data distribution. Unpaired t‐test: **, P < 0.001.
We next assayed for the NO_3_ ^−^ and NH_4_ ^+^ contents in seedlings transferred for two days to 10 mM NH_4_NO_3_ and 10 mM KNO_3_ following a two‐day N starvation. In this experiment, we measured increased concentrations of NO_3_ ^−^ in N‐treated gata hex seedlings when compared with the wild‐type, while NH_4_ concentrations were comparable between the two genotypes (Fig. 9e–g). Likewise, we measured comparative increases in NO_3_ ^−^ in the rosettes of 4‐wk‐old gata hex plants (Fig. 9h). We concluded that gata hex mutants have increased N content and increased NO_3_ ^−^ levels when compared with the wild‐type, which may not be explained by increased NO_3_ ^−^ uptake. Since gata hex has a decreased N‐responsive growth, these growth differences can thus not be easily explained by decreased N availability in the mutants.
Arabidopsis gata hex mutants are severely impaired in nitrogen‐responsive transcription
To examine gene expression changes in response to N, we exposed 10‐d‐old gnc gnl, gata hex, and wild‐type seedlings that had been N‐starved for 2 d to N (10 mM NH_4_NO_3_ and 10 mM KNO_3_) (Fig. 10a). A principal component analysis revealed a clear separation of all three replicates of the respective treatments (Fig. S11). After 1 and 4 h of N treatment, we detected 7541 and 10 791 DEGs, respectively, in the wild‐type, which included a comparable number of down‐ and upregulated DEGs at both time points (Fig. 10b,c; Table S8). Gene ontology (GO) analysis showed that N‐regulated DEGs in the wild‐type were enriched for terms such as RNA metabolic process, heat response, photosynthesis, or plastid organization, reflecting a broad response of the plant's physiology to N addition (Fig. S12; Table S9). Among the N‐regulated DEGs in the wild‐type, 627 up‐ and 943 downregulated DEGs from the 1‐h time point, as well as 620 up‐ and 992 downregulated DEGs from the 4‐h time point, were not N‐regulated in gnc gnl or gata hex (Fig. 10d–g; Table S8c).
Transcriptomic characterization of the N response in the Arabidopsis thaliana wild‐type and the gnc gnl and gata hex mutants. (a) Schematic overview of the RNA‐seq experiment. Wild‐type and mutant plants were pre‐cultured on N‐containing medium, starved of N for 2 d, and subsequently transferred to either N‐containing (+N; 10 mM KNO3 + 10 mM NO3NH4) or N‐free mock (–N; 10 mM KCl) medium for 1 or 4 h before RNA extraction. (b) Venn diagram showing overlap of differentially expressed genes (DEGs) between +N and –N treatments in Arabidopsis wild‐type at the 1‐ h and 4‐h time points. (c) Volcano plots illustrating the distribution of DEGs in the wild‐type at the 1‐ h and 4‐h time points, highlighting genes significantly upregulated or downregulated in response to N. The total number of variables (genes) is provided. (d–g) Venn diagrams comparing DEGs between the wild‐type and the gnc gnl and the gata hex mutants under +N vs –N conditions, separated into upregulated (d, f) and downregulated (e, g) gene sets at 1 h (d, e) and 4 h (f, g). Color‐coded sections indicate DEGs exclusive to the wild‐type but not in the gata hex mutant, which were used for subsequent GO enrichment analysis. (h–k) Heatmaps and corresponding gene ontology (GO) enrichment analyses of DEGs that were regulated in response to N in the wild‐type but not in the gata hex mutant, potentially representing AtB‐GATA‐dependent transcripts. Panels show upregulated (h, j) and downregulated (i, k) gene sets at 1 h (h, i) and 4 h (j, k).
Since it can be assumed that the differential regulation of these genes may be the consequence of the loss of the GATA factors, we specifically assessed the functions of the genes that were regulated in the wild‐type but not in gata hex. Among the upregulated gene sets, we found that DEGs related to changes in oxygen supply, cell wall biosynthesis, cytoskeleton, and Chl catabolism were regulated in the wild‐type but not in the mutant at the 1‐ or 4‐h time points (Fig. 10h–k; Table S10). Among the downregulated DEGs, genes related to plastid organization and chloroplast function were N‐regulated in the wild‐type but not in the gata hex mutant, which we noted with interest since the GNC and GNL GATA factors had previously been implicated not only in Chl biosynthesis but also in chloroplast and amyloplast biology (Chiang et al., 2012; Bastakis et al., 2018; Sala et al., 2023).
We next assessed the regulation of genes that had previously been associated with N biology in Arabidopsis, including genes required for N transport, N assimilation, and N‐dependent transcriptional responses (Fig. S13; Table S7a). Among these, several genes related to N transport were differentially regulated in the mutants, as well as genes encoding nitrate reductase (AtNIA), nitrite reductase (AtNIR), as well as glutamine synthetases (AtGLU; AtGS2) and asparagine synthetase (AtASN1), which showed strong differential regulation when compared with the wild‐type (Fig. S13; Table S7a). The latter expression changes could qualify for the alterations in nitrate levels that we had observed in the mutants after N treatment (Fig. 8). Among the transcriptional regulators, we noted, amongst others, differential regulation of AtbZIP1, AtCDF1, AtNLP6, AtNLP7, as well as AtTGA1 and AtTGA2 (Fig. S13c; Table S7a). When we specifically assessed the regulation of the B‐GATA genes in response to the N treatment, we detected an N‐dependent downregulation of GNC and GNL, but at the same time an upregulation of the B‐GATA factors GATA15, GATA17, and GATA17L (Fig. S13d).
Strong differential regulation of cytokinin signaling genes in the gata hex mutant
Mutants of the B‐GATA genes have been associated with reduced Chl biosynthesis (R. Richter et al., 2010; Bastakis et al., 2018; Schroder et al., 2023). When we assessed the RNA‐seq data with regard to the expression of genes required for Chl biosynthesis, we detected the downregulation of a broad set of Chl biosynthesis genes in gata hex when compared with the wild‐type, highly similar to previously published gene expression studies related to the role of B‐GATA genes in greening (Fig. S14; Table S7).
When we explored genes with a role in cytokinin biosynthesis, we did not note gene expression changes that would qualify to explain the increased cytokinin content detected in the gata hex mutant (Fig. S15a–e; Table S7). Strikingly, we found that the expression of the majority of genes with a role in cytokinin signaling was upregulated in the mutant when compared with the wild‐type, suggesting a strong impairment of cytokinin signaling in the mutants, a possible consequence of the elevated cytokinin levels in the mutant or defective feedback regulation (Fig. S15a–e; Table S7).
Discussion
B‐GATA
gene structure
In this study, we characterize the biological function of the four B‐GATA genes from the moss Physcomitrium. Unlike vascular plant B‐GATAs, but similar to the single B‐GATA in the bryophyte Marchantia, moss B‐GATAs contain HAN‐ and LLM‐domains (Schwechheimer et al., 2022). The biochemical activity of either domain is unknown, and mutation analyses of B‐GATA genes in vascular plants showed that both domains are individually required for full function of the respective B‐GATA subtypes (Whipple et al., 2010; Behringer et al., 2014). Given the co‐occurrence of both domains in bryophyte B‐GATAs, it had previously been proposed that the common land plant B‐GATA ancestor may have contained both domains (Schwechheimer et al., 2022). It has also been suggested that the two domains are functionally interdependent and that, consequently, heterodimerization between HAN‐ and LLM‐domain B‐GATAs may be required for full B‐GATA protein activity in vascular plants (Zhang et al., 2013; Schwechheimer et al., 2022). However, the absence of phenotypes typically observed in mutants of HAN‐domain B‐GATAs, for example abnormal flower morphology and embryo lethality, in higher‐order mutants of LLM‐domain B‐GATAs, as in the gata hex mutant used in this study, which carries mutations in all six Arabidopsis LLM‐domain B‐GATA genes and was previously unavailable, now argues against such functional interdependence between the two B‐GATA subfamilies (Zhao et al., 2004; Zhang et al., 2013; Schwechheimer et al., 2022; Schroder et al., 2023).
The four LLM‐domain B‐GATAs from Physcomitrium exhibit very high sequence conservation and are essentially identical in the conserved zinc finger domain, the HAN‐domain, and the LLM‐domain (Fig. 1). In vascular plants, the zinc finger DNA‐binding domain among the B‐GATAs is highly conserved, but its sequence allows predicting whether a B‐GATA factor contains a HAN‐ or an LLM‐domain (Behringer et al., 2014; Schwechheimer et al., 2022). The presence of a basic lysine (K) residue in the zinc finger domain of the four Physcomitrium B‐GATAs, as well as in the single Marchantia B‐GATA, at a position where vascular plant HAN‐domain B‐GATAs have a serine (S), suggests that bryophyte B‐GATAs, from this perspective, are more closely related to LLM‐domain B‐GATAs than to HAN‐domain B‐GATAs of vascular plants (Fig. 1).
Functional redundancy among the
PpB‐GATAs
Although we lack functional evidence from dedicated genetic studies, we assume that the four B‐GATAs are functionally redundant. Expression analyses using publicly available data indicate that their expression has diverged across different cell and tissue types. We generated single, double, and triple mutants with defects in PpB‐GATA1, PpB‐GATA3, and PpB‐GATA4. We unsuccessfully used two alternative approaches to obtain a mutant for the fourth family member, PpB‐GATA2. Whether PpB‐GATA2 harbors a critical biological role that would preclude mutant survival is difficult to assess. The relatively low expression of the gene, along with the presumed functional redundancy among the PpB‐GATAs, would be an argument against such an essential role.
Ppb‐gata1/3/4 mutants have reduced Chl and carotinoid levels
In order to be able to perform the analysis with a reasonable effort and, in part, cost, we have focused our analysis primarily on the study of the moss Ppb‐gata1/3/4 triple mutant. We found that the Ppb‐gata1/3/4 triple mutant has reduced Chl and carotenoid contents, and thereby the moss mutant phenotypically resembles B‐GATA gene mutants from Arabidopsis and Marchantia (Ranftl et al., 2016; Bastakis et al., 2018; Schroder et al., 2023) (Bi et al., 2005; Hudson et al., 2011). The reduced expression of several Chl biosynthesis genes in Ppb‐gata1/3/4, notably the downregulation of multiple HEME gene orthologs, CHLI, and PORA, as observed in our RNA‐seq analysis, may account for the observed Chl reduction. As a note of caution, we would like to emphasize here the fact that the Chl quantifications, as well as the cytokinin quantifications discussed below, were obtained from plants grown on N‐containing medium, whereas the RNA‐seq data refer to plant material from N‐starved plants.
N‐responsive growth is defective in B‐GATA
gene mutants
Importantly, we found that Ppb‐gata1/3/4 mutants are defective in the formation of protonemata typically observed in moss in growth medium containing the N sources NO_3_ and NH_4_. Similarly, we observed reduced growth in response to N supply when growing Arabidopsis gnc gnl double mutants and even more so when growing gata hex hexuple mutants on soil containing increased N contents or supplemented with fertilizer. Further, we observed reduced systemic growth in gata hex when roots were grown on N‐depleted medium in split‐root assays.
Transcriptional responses to N strongly depend on B‐GATA
genes
These observations triggered our interest in studying the role of the B‐GATAs in their transcriptional response to N after N‐starvation, initially in Physcomitrium and subsequently in Arabidopsis. In both species, we found a substantial reduction (Physcomitrium 42%; Arabidopsis 27%) in the number of genes regulated by N‐supply in the wild‐type after 1 and 4 h, suggesting that the B‐GATA genes make a significant contribution to the transcriptional N response.
Functional characterization of the classes of genes required for N uptake, assimilation, and response revealed differential regulation of genes belonging to all functional classes in the moss and Arabidopsis mutants. In combination with physiological analysis, we could conclude, however, that neither mutant was defective in N uptake. Similarly, the expression of genes required for N assimilation, specifically the enhanced N‐induced expression of genes encoding for nitrate reductases (moss Pp6c10_5150, Pp6c10_5180 and Arabidopsis AT1G77760, AT1G37130) and nitrite reductases (Pp6c25_1290, Pp6c16_12770, Pp6c1_8170 and AT2G15620), as well as for the genes encoding for glutamine (Pp6c2_2380, Pp6c20_11610 and AT3G60120) and asparagin synthetases (AT3G47340 and AT5G65010), argues for enhanced N assimilation in the mutants. In line with these observations, our physiological analyses found increased NO_3_ contents in mutants after N supplementation of N‐starved plants, at least in Arabidopsis, whereas NO_3_ contents in moss were below the detection limit. In moss, in turn, NH_4_ contents were reduced when N‐starved Ppb‐gata1/3/4 mutants were supplemented with NH_4_. From this analysis, we can conclude that the transcriptional responses for N assimilation genes are strongly affected in the B‐GATA gene mutants from both species, which may correlate with altered N metabolism in the respective plants.
Transcriptional responses downstream from N supply have been studied extensively, and time‐resolved kinetic analyses have identified transcription factors that respond to N supply. Several of the relevant studies identify GATA motifs as overrepresented promoter elements in N‐regulated genes, arguing from this angle for an important role of GATA factors as regulators of N‐responsive growth. Many of these transcription factors are differentially regulated in the moss Ppb‐gata1/3/4, as well as in the Arabidopsis gnc gnl, and gata hex mutants, for example CDF1 (Pp6c15_10030 and AT5G23040), NLP6 (Pp6c19_1500 and AT1G64530), NLP7 (Pp6c17_1870 and AT4G24020), TCP20 (Pp6c10_10790 and AT3G27010), TGA1 (Pp6c3_4570 and AT5G65210), TGA4 (Pp6c14_2590 and AT5G10030), which invites the conclusion that at least some of the transcriptional responses downstream from N are conserved between moss and Arabidopsis (Ristova et al., 2016; Ruffel et al., 2016; Varala et al., 2018; Brooks et al., 2019).
B‐GATA gene functions can be linked to cytokinin biology
Intriguingly, the growth defect of the Physcomitrium Ppb‐gata1/3/4 mutant resembles that of moss lines with impaired cytokinin‐regulated development caused by overexpression of the KNOTTED family transcription factor MKN2 or by misexpression of an ISOPENTENYL TRANSFERASE gene in the mkn2 mutant (Coudert et al., 2019). In Arabidopsis, GNL (GNC‐LIKE) was previously identified as a strongly cytokinin‐responsive gene, which led to its alternative name CGA1 (CYTOKININ‐RESPONSIVE GATA1). We have previously shown that the transcription of all six Arabidopsis LLM‐domain B‐GATA genes can be induced by cytokinin, albeit less strongly than that of GNL/CGA1 (Naito et al., 2007; Ranftl et al., 2016). In the same study, we found that a mutant lacking five of the six Arabidopsis LLM‐domain B‐GATAs exhibits defects in phyllotaxy, floral organ initiation, accessory meristem formation, and senescence, phenotypes attributable to impaired cytokinin signaling (Ranftl et al., 2016). Similarly, Mpb‐gata1 mutants from the liverwort Marchantia display gemma cup formation defects that phenocopy those of mutants in the MYB transcription factor GCAM1 (GEMMA CUP‐ASSOCIATED MYB1) (Schroder et al., 2023). GCAM1 is a master regulator of vegetative reproduction in Marchantia and has recently been linked to the cytokinin pathway in this species (Komatsu et al., 2025). Here, we show that moss Ppb‐gata1/3/4 and Arabidopsis gata hex mutants accumulate higher cytokinin levels than their respective wild‐type counterparts. In mutants from both species, numerous genes associated with cytokinin biosynthesis (Physcomitrium) or signal transduction (Arabidopsis) are differentially expressed, indicating that cytokinin‐responsive transcription or feedback regulation in the cytokinin pathway are misregulated. The reduced accumulation of Chl, as well as the reduced transcription of Chl biosynthesis genes and other aspects of chloroplast biology, may, at least in Arabidopsis, also be explained by a reduction of cytokinin signaling (Chiang et al., 2012; Ranftl et al., 2016). Collectively, these findings suggest that B‐GATAs contribute to cytokinin‐regulated growth in mosses, bryophytes, and angiosperms, pointing to an evolutionary conservation of this role for B‐GATA factors in land plants.
In summary, our study identifies an important role for the B‐GATA genes in mediating N‐dependent growth in Physcomitrium patens and Arabidopsis thaliana. The increases in cytokinin levels in mutants from both species and the strongly impaired transcriptional responses, in combination with the impairment of N‐responsive growth, suggest that the B‐GATA genes may translate the role that has been attributed to cytokinin as a systemic regulator of N‐mediated plant growth. Through our studies, we provide solid and strong proof for the anticipated but as yet unproven role of plant GATAs in N‐dependent plant growth regulation. Dedicated studies are needed to examine the functional and potentially direct relationship between the GATA factors and the reported N‐dependent gene expression changes.
Competing interests
None declared.
Author contributions
DZ and CS conceived all biological experiments. PMS generated the *Physcomitrium Ppb‐*gata gene mutants. IP and ON performed and analyzed the cytokinin quantifications. XD and KS analyzed the Ppb‐gata mutant genome sequences. RS supported and analyzed the ^15^N uptake studies.
Disclaimer
The New Phytologist Foundation remains neutral with regard to jurisdictional claims in maps and in any institutional affiliations.
Supporting information
Fig. S1 Phylogenetic tree of B‐GATA proteins from Physcomitrium patens (Pp), Marchantia polymorpha (Mp), and Arabidopsis thaliana (At). Fig. S2 PpB‐GATA gene expression levels across different tissues in Physcomitrium patens. Fig. S3 Genotypes of Physcomitrium patens Ppb‐gata mutant alleles obtained by CRISPR/Cas9‐based mutagenesis. Fig. S4 PpB‐GATA1 overexpression suppresses growth and enhances Chl and carotenoid accumulation in Physcomitrium patens. Fig. S5 Principal component analysis (PCA) of transcriptomic profiles from Physcomitrium patens Reute and the Ppb‐gata1/3/4 triple mutant. Fig. S6 Gene ontology (GO) enrichment analysis of differentially expressed genes (DEGs) in response to nitrogen in Physcomitrium patens Reute. Fig. S7 Differential regulation of nitrogen transport, metabolism, and transcription regulation genes may underlie the reduced N response in Physcomitrium patens Ppb‐gata1/3/4 mutants. Fig. S8 Altered expression of Chl biosynthesis genes may contribute to reduced Chl accumulation in Physcomitrium patens Ppb‐gata1/3/4 mutants. Fig. S9 Expression profiles of cytokinin biosynthesis genes in Physcomitrium patens Ppb‐gata1/3/4 mutants. Fig. S10 Loss of GNC and GNL leads to a partial suppression of N‐dependent growth responses in Arabidopsis thaliana. Fig. S11 Principal component analysis (PCA) of transcriptomic profiles from Arabidopsis thaliana and b‐gata mutants. Fig. S12 Gene ontology (GO) enrichment analysis of differentially expressed genes (DEGs) in response to N in Arabidopsis thaliana. Fig. S13 Differential regulation of nitrogen transport, metabolism, and transcription regulation genes may underlie the reduced N response in gnc gnl and gata hex mutants from Arabidopsis thaliana. Fig. S14 Altered expression of Chl biosynthesis genes in Arabidopsis thaliana gnc gnl and gata hex mutants. Fig. S15 Expression profiles of cytokinin biosynthesis genes in Arabidopsis thaliana gnc gnl and gata hex mutants. Methods S1 Supplemental materials and methods.
Table S1 List of primers used in the study.
Table S2 Results of the whole genome sequencing of Physcomitrium patens Ppb‐gata mutant lines.
Table S3 Results of the cytokinin quantifications in Physcomitrium patens and Arabidopsis thaliana wild‐type and gata mutants.
Table S4 Results of the RNAseq analysis conducted in Physcomitrium patens.
Table S5 Results from a gene ontology (GO) enrichment analysis results from g:Profiler of differentially expressed genes (DEGs) in nitrogen (10 mM KNO_3_ + 5 mM (NH_4_)2_C_4_H_4_O_6) vs mock (10 mM KCl) conditions in Physcomitrium patens Reute.
Table S6 Results from a gene ontology (GO) enrichment analysis results from g:Profiler of differentially expressed genes (DEGs) in nitrogen (10 mM KNO_3_ + 5 mM (NH_4_)2_C_4_H_4_O_6) vs mock (10 mM KCl) conditions in Physcomitrium patens Reute but not in Ppb‐gata1/3/4, grouped according to up‐ or downregulation and time point as shown in Fig. 6.
Table S7 Lists of genes from Arabidopsis thaliana and their orthologues from Physcomitrium patens involved in nitrogen response, Chl biosynthesis, and cytokinin biosynthesis and response, as analyzed for this study.
Table S8 Results of the RNAseq analysis conducted in Arabidopsis thaliana.
Table S9 Results from a gene ontology (GO) enrichment analysis results from ClusterProfiler of differentially expressed genes (DEGs) in nitrogen (10 mM KNO_3_ + 10 mM NH_4_NO_3_) vs mock (10 mM KCl) conditions in Arabidopsis thaliana (Col‐0) wild‐type, grouped according to up‐ or downregulation and time point as shown in Fig. S12.
Table S10 Results from a gene ontology (GO) enrichment analysis from ClusterProfiler of differentially expressed genes (DEGs) in nitrogen (10 mM KNO_3_ + 10 mM NH_4_NO_3_) vs mock (10 mM KCl) conditions in Arabidopsis thaliana (Col‐0) that are not nitrogen‐responsive in the gata hex mutant, grouped according to up‐ or downregulation and time point as shown in Fig. 10h–k.Please note: Wiley is not responsible for the content or functionality of any Supporting Information supplied by the authors. Any queries (other than missing material) should be directed to the New Phytologist Central Office.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Arst HN Jr , Cove DJ . 1973. Nitrogen metabolite repression in Aspergillus nidulans . Molecular & General Genetics 126: 111–141.4591376 10.1007/BF 00330988 · doi ↗ · pubmed ↗
- 2Bastakis E , Hedtke B , Klermund C , Grimm B , Schwechheimer C . 2018. LLM‐domain B‐GATA transcription factors play multifaceted roles in controlling greening in Arabidopsis. Plant Cell 30: 582–599.29453227 10.1105/tpc.17.00947 PMC 5894840 · doi ↗ · pubmed ↗
- 3Behringer C , Bastakis E , Ranftl QL , Mayer KF , Schwechheimer C . 2014. Functional diversification within the family of B‐GATA transcription factors through the leucine‐leucine‐methionine domain. Plant Physiology 166: 293–305.25077795 10.1104/pp.114.246660 PMC 4149714 · doi ↗ · pubmed ↗
- 4Behringer C , Schwechheimer C . 2015. B‐GATA transcription factors – insights into their structure, regulation, and role in plant development. Frontiers in Plant Science 6: 90.25755661 10.3389/fpls.2015.00090 PMC 4337238 · doi ↗ · pubmed ↗
- 5Bi YM , Zhang Y , Signorelli T , Zhao R , Zhu T , Rothstein S . 2005. Genetic analysis of Arabidopsis GATA transcription factor gene family reveals a nitrate‐inducible member important for chlorophyll synthesis and glucose sensitivity. The Plant Journal 44: 680–692.16262716 10.1111/j.1365-313X.2005.02568.x · doi ↗ · pubmed ↗
- 6Brooks MD , Cirrone J , Pasquino AV , Alvarez JM , Swift J , Mittal S , Juang CL , Varala K , Gutierrez RA , Krouk G et al. 2019. Network Walking charts transcriptional dynamics of nitrogen signaling by integrating validated and predicted genome‐wide interactions. Nature Communications 10: 1569.10.1038/s 41467-019-09522-1PMC 645103230952851 · doi ↗ · pubmed ↗
- 7Caddick MX . 1994. Nitrogen metabolite repression. Progress in Industrial Microbiology 29: 323–353.7765131 · pubmed ↗
- 8Castaings L , Camargo A , Pocholle D , Gaudon V , Texier Y , Boutet‐Mercey S , Taconnat L , Renou JP , Daniel‐Vedele F , Fernandez E et al. 2009. The nodule inception‐like protein 7 modulates nitrate sensing and metabolism in Arabidopsis. The Plant Journal 57: 426–435.18826430 10.1111/j.1365-313X.2008.03695.x · doi ↗ · pubmed ↗