Low abundance of phytophagous nematodes under invasive exotic Pinus elliottii – enemy release and plant–soil feedbacks
Lynda S. C. Guerrero, Erika Buscardo, Mario M. Inomoto, Laszlo Nagy

TL;DR
Invasive pine trees reduce nematode enemies in soil, helping them spread more easily in new ecosystems.
Contribution
Shows invasive Pinus elliottii reduces phytophagous nematodes, supporting enemy release and altering soil feedbacks.
Findings
Nematode load on pine is lower than on native species, supporting the enemy release hypothesis.
Pine invasion depletes phytophagous nematodes and reduces overall nematode richness over time.
Pine modifies soil organism composition, influencing plant-soil feedbacks in neotropical ecosystems.
Abstract
According to the enemy release hypothesis (ERH), the fitness of exotic plants and their capacity to become invasive in their area of introduction may partly be attributable to the loss of their natural enemies. Invasive species may also benefit from modifying soil attributes and thereby creating a positive soil–plant feedback.We assessed the relationship between time since the establishment of the invasive Pinus elliottii and enemy release in a montane pine–nematode‐specific context within the Atlantic Forest domain, by comparing soil nematode communities/functional diversity along a virtual chronosequence of invasion.Our findings confirmed the premises of the ERH and suggest that invasion may be facilitated by a lesser nematode load on pine compared to that on native species. The impact of P. elliottii on nematode communities over time was mainly driven by changes in the trophic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
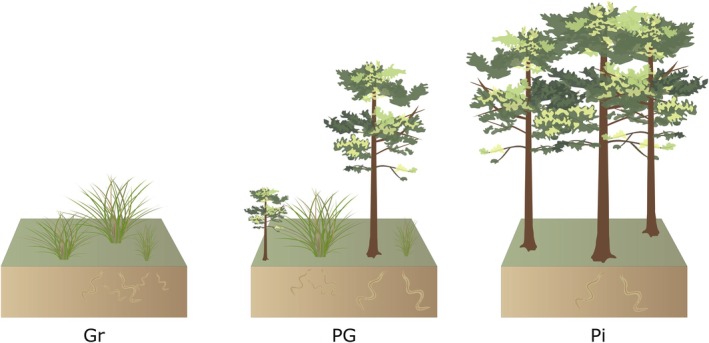 Fig. 1
Fig. 1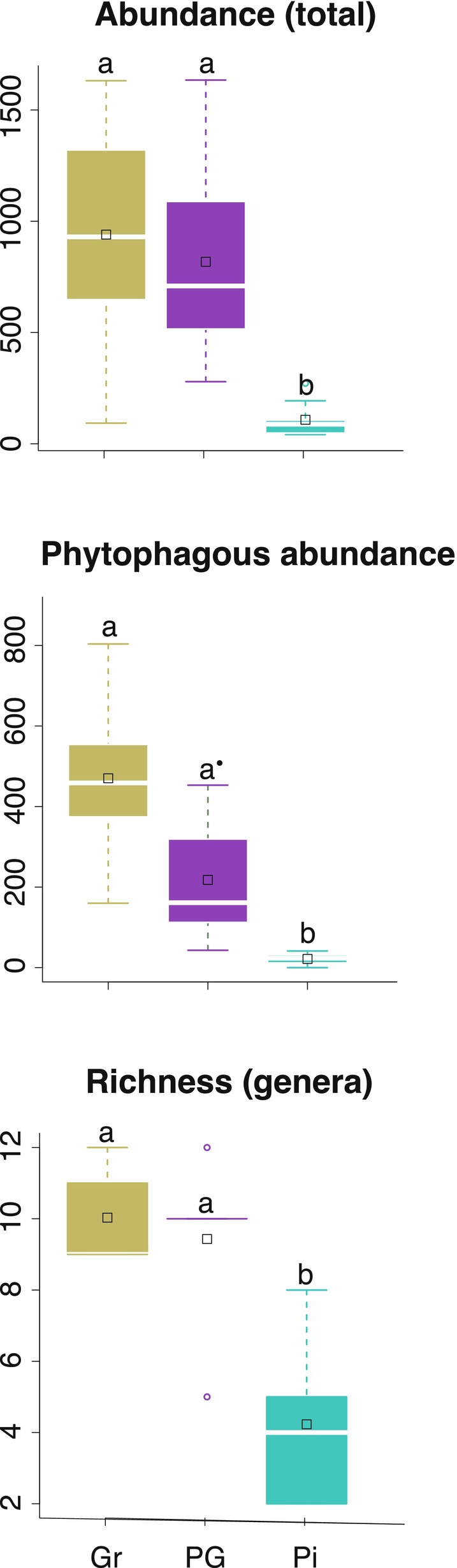 Fig. 2
Fig. 2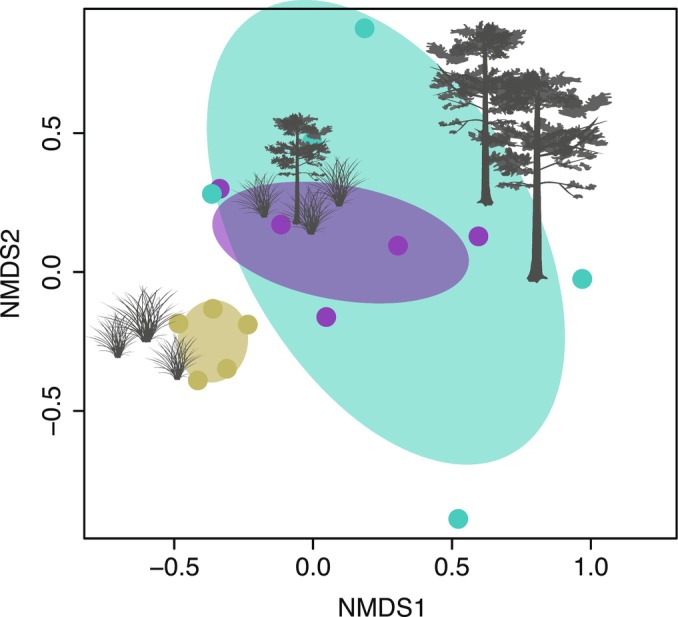 Fig. 3
Fig. 3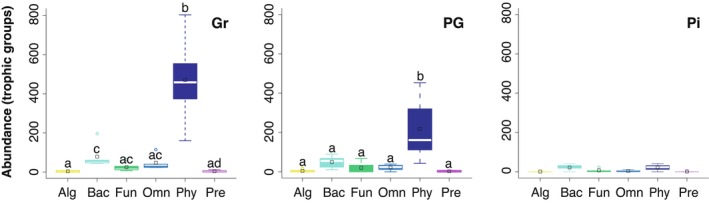 Fig. 4
Fig. 4| Family | Genus | Trophic group | Gr | PG | Pi |
|---|---|---|---|---|---|
| Alloionematidae |
| 3 | − | + | − |
| Anguinidae |
| 2 | + | − | − |
| Aphelenchidae |
| 2 | − | − | + |
| Aphelenchoididae |
| 2 | + | + | + |
| Aphelenchoididae |
| 5 | − | + | − |
| Aulolaimoididae |
| 3 | − | + | − |
| Belondiridae |
| 8 | + | − | − |
| Belonolaimidae |
| 1 | − | − | + |
| Cephalobidae |
| 3 | + | + | + |
| Cephalobidae |
| 3 | − | + | + |
| Cephalobidae |
| 3 | + | − | − |
| Cephalobidae |
| 3 | − | + | − |
| Cephalobidae |
| 3 | + | − | + |
| Criconematidae |
| 1 | + | + | − |
| Criconematidae |
| 1 | + | − | − |
| Criconematidae |
| 1 | − | − | + |
| Criconematidae |
| 1 | − | + | − |
| Criconematidae |
| 1 | − | + | + |
| Diplogastridae |
| 3 | − | + | − |
| Diploscapteridae |
| 3 | + | + | − |
| Dorylaimidae |
| 8 | − | + | + |
| Hemicycliophoridae |
| 1 | + | − | − |
| Hoplolaimidae |
| 1 | + | + | + |
| Hoplolaimidae |
| 1 | + | − | − |
| Leptonchidae |
| 8 | + | − | − |
| Longidoridae |
| 1 | − | − | + |
| Longidoridae |
| 1 | + | + | + |
| Meloidogynidae |
| 1 | + | − | − |
| Mononchidae |
| 5 | + | − | − |
| Mononchidae |
| 5 | + | − | − |
| Mylonchulidae |
| 5 | + | − | − |
| Odontopharyngidae |
| 3 | − | + | − |
| Panagrolaimidae |
| 3 | − | + | − |
| Panagrolaimidae |
| 3 | + | − | − |
| Paratylenchidae |
| 1 | − | + | − |
| Paratylenchidae |
| 1 | + | + | − |
| Plectidae |
| 3 | − | + | − |
| Pratylenchidae |
| 1 | − | + | − |
| Prismatolaimidae |
| 3 | + | − | + |
| Rhabditidae |
| 3 | + | + | − |
| Rhabditidae |
| 3 | − | + | − |
| Teratocephalidae |
| 3 | − | − | + |
| Thornenematidae |
| 8 | + | + | − |
| Trichodoridae |
| 1 | − | + | − |
| Tylenchidae |
| 1 | + | − | + |
| Tylenchidae |
| 2 | + | + | + |
| Tylencholaimellidae |
| 2 | − | − | + |
| Tylencholaimidae |
| 2 | − | + | − |
| Tylencholaimidae |
| 2 | + | + | − |
- —Conselho Nacional de Desenvolvimento Científico e Tecnológico10.13039/501100003593
- —Coordenação de Aperfeiçoamento de Pessoal de Nível Superior10.13039/501100002322
- —São Paulo State Research Foundation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNematode management and characterization studies · Mycorrhizal Fungi and Plant Interactions · Entomopathogenic Microorganisms in Pest Control
Introduction
The success of plant introductions has partly been explained by the presumed escape of introduced plants in their new environment from their natural enemies (e.g. herbivores, pathogens) that occur in their natural range (viz. enemy release hypothesis – ERH; Keane & Crawley, 2002) and which results in a reduced enemy load. The likelihood and strength of enemy release, depends on specific contexts, including phylogenetic relatedness of exotic and native species, time since introduction and resource levels (Keane & Crawley, 2002; Blumenthal, 2006; Mitchell et al., 2006; Hawkes, 2007; Brian & Catford, 2023).
In addition to the reduction in natural enemy pressure, the exotic species may further modify soil attributes/organisms in their rhizospheres (e.g. the composition and structure of soil communities via new biochemistry). According to the ‘novel weapon’ hypothesis (Callaway & Ridenour, 2004), exotic plant species may also produce defense compounds that are not produced by species of the invaded community that could further reduce the abundance and/or richness of potential enemies, including plant‐feeding nematodes (Callaway et al., 2008; Schaffner et al., 2011).
This affects, in turn, the plants via positive soil biota–plant feedback (Klironomos, 2002; Reinhart et al., 2003; van der Putten et al., 2005; Suding et al., 2013; Dickie et al., 2017a; Wilschut et al., 2019). Whether feedbacks are positive or negative depends on the balance between relative positive effects of beneficial soil organisms (e.g. mycorrhizas, nitrogen‐fixing bacteria) and the relative negative effects of plant enemies (e.g. soil‐borne pathogens, herbivores) as well as the indirect effect that plant–soil biota interactions have on plant–plant interactions (Reinhart & Callaway, 2006).
Introduced species may therefore improve their fitness (population growth/reproductive potential) via a reduction in fitness limitation that their natural enemies cause in their home range and via the benefit that they potentially derive from altering soil chemistry and nutrient cycling and modifying plant–soil feedbacks in their exotic range (Keane & Crawley, 2002; Callaway et al., 2004; Wubs et al., 2019; Aldorfová et al., 2020). This, in the absence of dispersal limitation, could increase the spread from the site of introduction to neighbouring habitats, such as has been the case of several species of the genus Pinus introduced in the Southern Hemisphere (Simberloff et al., 2010), where, except for P. merkusii Jungh. & Vriese in south‐east Asia, no native species belonging to Pinaceae exist.
A recent census has shown that plantations of Pinus spp. in Brazil cover c. 1.6 M ha, most of it within the Atlantic Forest biogeographic domain (Industria Brasileira de Árvores, 2020). The plantations have replaced native vegetation, dominated by arbuscular mycorrhizal (AM) symbionts, with monospecific exotic pine species that form ectomycorrhizal (ECM) associations (Wilson & Mayle, 2024). Additionally, plantations have served as source populations of invasion (e.g. P. elliottii Engelm., P. patula Schtdl. & Cham., and P. taeda L.) of montane grasslands (Milani et al., 2020), rich in endemic species, including protected areas in the Mantiqueira mountain range, south‐eastern Brazil. The establishment of pine species has been changing the structure and ecology of these native ecosystems (Taylor et al., 2016; Moyano et al., 2023).
Pine–fungal symbiont co‐invasion can have major effects on ecosystem properties such as soil nutrient cycling (e.g. recalcitrant litter and root exudates; Chapela et al., 2001; Reich et al., 2005), community composition of soil biota and on soil functions, either through modified resource inputs and/or through changes in biotic interactions (Dickie et al., 2011, 2014, 2017b; Peralta et al., 2019; Sapsford et al., 2022). Soil communities mediate plant–soil feedbacks between non‐native and native species, also described for processes during pine invasion (Nuske et al., 2021; Green et al., 2025; Dudenhöffer & Hulme, 2025a,b). Pines depend on ECM fungi for successful establishment and spread and the role of fungal symbionts in pine invasion has received significant attention (Nuñez et al., 2009; Dickie et al., 2010; Policelli et al., 2019, 2023). In contrast to the positive effects of beneficial ECM fungi, the potential role played by soil‐borne enemies on pine invasion and the feedback of enemy release on plant–soil interaction has been less investigated.
Nematodes are important soil‐borne enemies that could directly (e.g. damage to plant tissues) and indirectly (e.g. changes in trophic interactions) affect the invasiveness of exotic pine species in the Southern Hemisphere (Thakur & Geisen, 2019). Nematodes are the most abundant and functionally diverse soil animals (van den Hoogen et al., 2019). They are involved in soil processes including decomposition and nutrient cycling, and affect plant growth (Ingham et al., 1985; Osler & Sommerkorn, 2007; Neher et al., 2012). Based on their feeding habits they have been classified into phytophagous species (henceforth also referred to as plant‐ or root‐feeding nematodes), bacterivores, omnivores, fungivores, algivores and predators (Yeates et al., 1993).
Root‐feeding nematodes cause tissue necrosis, gall formation and inhibition of root growth (Ruehle, 1973) and their presence and/or activity predispose plants to be attacked by opportunistic pathogens such as fungi (Castillo et al., 1998; Back et al., 2002; Jones et al., 2013; Wheeler et al., 2019). The ecological interactions of nematodes with their hosts and with other consumers of the soil food web ultimately affect plant performance and distribution (Neher, 2010; Topalović & Geisen, 2023). Damage caused by phytophagous nematodes to forest trees generally results in nonspecific aboveground symptoms to the plant, making it difficult to quantify nematode effects in the field (Singh et al., 2015; Khan et al., 2020). Phytophagous nematodes reduce seedling growth (e.g. Rotylenchus spp.; Magnusson, 1983) and cause chlorosis and death of pine seedlings in nurseries (e.g. species of Belonolaimus, Hoplolaimus, Pratylenchus, Tylenchorhynchus and Trichodorus; Ruehle, 1966, 1968). Species of some genera, including Criconemoides, Hemicycliophora, and Helicotylenchus parasitise and reproduce on P. elliottii (Khan, 2012).
The diversity and abundance of nematode communities have been shown to be negatively impacted by exotic pine species (Yeates & Saggar, 1998; Scholes & Nowicki, 2000; Dehlin et al., 2008). It remains poorly known, however, if invasive pine species experience enemy release from soil phytophagous nematodes and if soil enemy abundance/diversity changes at different stages of the invasion pathway. Quantifying the relationship between time since invasive pine establishment and the abundance and diversity of phytophagous nematodes in montane ecosystems within the Atlantic Forest domain would contribute to a better understanding of invasion dynamics.
Using as a starting point the community approach where co‐occurring native and exotic species are compared within the same community (Colautti et al., 2004), we hypothesised that, as predicted by the ERH, the abundance of phytophagous nematodes would be less in soils under individuals of P. elliottii colonising native shrubby grasslands (locally called campos de altitude, henceforth open vegetation) compared with open vegetation without pine (H1). We also hypothesised that P. elliottii would exploit the reduction of enemy pressure by root‐feeding nematodes (enemy release) and further reduce their abundance and modify the composition/structure of soil nematode communities in pine plantations (H2a). In alternative to H2a, phytophagous nematodes would increase in pine plantations as generalists become better adapted to the invader and tend to accumulate (H2b; Flory & Clay, 2013; Crous et al., 2016; Dickie et al., 2017a; Brian & Catford, 2023).
To test our hypotheses, we quantified the abundance of phytophagous nematodes in: (1) soils under native open vegetation (control, time 0); (2) under established P. elliottii individuals invading open vegetation (time 1; representing up to c. 30 yr since establishment); and (3) in a monospecific P. elliottii plantation established in the late 1950s, with sparse and species‐poor understorey (time 2; c. 65 yr‐old), thus creating, in a space‐for‐time substitution, a virtual chronosequence of invasion (Fig. 1).
Vegetation types within the Atlantic Forest domain, south‐eastern Brazil. The abundance and functional diversity of soil nematodes was compared among native shrubby grassland locally known as campos de altitude and henceforth referred to as open vegetation (Gr; reference point in time, time 0), 20‐ to 30‐yr‐old exotic Pinus elliottii individuals invading open vegetation (PG; time 1), and P. elliottii plantation (Pi; time 2) established in the late 1950s. Symbols are courtesy of the Integration and Application Network (ian.umces.edu/symbols/), University of Maryland Center for Environmental Science.
Materials and Methods
Study area
The study was carried out at a tropical montane forest site (22°39′S and 45°26′W–45°30′W; 1550 m above sea level), in a conservation area in the state of São Paulo, Brazil. The climate is humid subtropical – oceanic characterised by warm and wet summers and temperate dry winters (Cfb by Köppen classification), with no negative hydrological balance. The average annual temperature between 1973 and 2000 was 14.7°C, and the mean annual rainfall was 1813 mm, with most precipitation falling between October and March.
The soils (latosols, podzols and entisols) have formed over metamorphic rocks with various types of orthogneiss (e.g. gneissified granite, gneiss, migmatite, aplite, granulite; Seibert et al., 1975; Modenesi‐Gauttieri & Hiruma, 2004). They vary in texture and thickness and are characterised by a low pH, low cation exchange capacity and base saturation and high values of exchangeable aluminium (Oliveira et al., 1975). The vegetation consists of a mosaic of secondary forest patches in various stages of regeneration, open vegetation and plantations of exotic conifers and of eucalyptus (c. 20%). The native forest is characterised by two native conifers, Araucaria angustifolia (Bertol.) Kuntze (an endemic species threatened of extinction; IUCN, 2011) and Podocarpus lambertii Klotzsch ex Endl., and various angiosperms, with the emergent layer being dominated by A. angustifolia. Open vegetation is characterised by highly diverse herbaceous vegetation, shrubs and scattered small trees (Seibert et al., 1975).
The site history includes livestock husbandry, subsistence farming and timber extraction over c. 150 yr, up to the late 1970s. The Forestry Institute of São Paulo undertook a reforestation programme of deforested areas with exotic conifers in the late 1950s and early 1960s (Anonymous, 2015). Some of the species, such as P. elliottii, P. taeda or P. patula have become invasive and have been colonising open vegetation.
Pinus elliottii is a species native to North America and is among the known invasive species in the Southern Hemisphere (Simberloff et al., 2010). Species of the genus Pinus are generally tolerant of dry conditions, nutrient‐poor soils and fire regimes (Zanchetta et al., 2007). Pinus elliottii has been reported to invade savanna (de Abreu & Durigan, 2011; Brewer et al., 2018) as well as coastal sandy plains (Bechara et al., 2013), open A. angustifolia secondary forests (Zenni & Simberloff, 2013) and open montane vegetation (Milani et al., 2020), such as that at our long‐term ecological research site in the Mantiqueira range, south‐eastern Brazil. It produces a large number of winged seeds (2–3.6 M seeds ha^−1^yr^−1^; Bechara et al., 2013; Miashike, 2015) that are efficiently dispersed by wind (de Abreu & Durigan, 2011). The density of individuals in invaded Brazilian savanna can reach c. 3500 individuals ha^−1^ (de Abreu & Durigan, 2011) with pulses of invasion (5–7 yr) likely to be linked to founder trees reaching maturity and to favourable climatic events (Brandes et al., 2020). Pine canopy cover/density for similarly aged pine stands has been found to be significantly greater in the introduced (Brazilian savanna) than in the native range (savannas in Mississippi; Brewer et al., 2018).
Sampling design
Soil samples were collected under open native vegetation (control, time 0), under 20‐ to 30‐yr‐old pine individuals invading native open vegetation (time 1), and under pine individuals in a pine plantation established in 1958–1959 c. 65 yr ago, thinned twice (last time in 1984), with the removal of 30% of standing timber volume, followed by natural regeneration (São Paulo State Forest Institute archives, w/o date; time 2; Fig. 1, Supporting Information Fig. S1). This allowed us to quantify the relationship between time since invasive pine establishment and the abundance and diversity of phytophagous nematodes.
Eight sample points were randomly selected in the plantation using the ‘Sample’ module in TerrSet (https://clarklabs.org/terrset/). The selection of target trees in the open vegetation, was determined by their available position. Within the open vegetation, we sampled soil under invasive pines and under native species not affected by the presence of pine. A minimum distance of 50‐m between samples (in both open vegetation and plantation) was observed to avoid spatial autocorrelation and guarantee the independence between samples (Buscardo et al., 2018).
The soil samples were collected in the austral winter (August 2019) using a soil corer with an internal diameter of 5.7 cm, equipped with a plastic liner tube (Giddings Machine Co., Fort Collins, CO, USA). For each sampling point, four soil cores, separated by at least 1.5 m, were collected around a target tree in the plantation and in invaded open vegetation and around a reference point under native species not affected by the presence of pine. The mineral layer (top 10 cm) and the organic horizon (lacking in open vegetation) were separated and the samples per sampling point were bulked to form composite samples. A total of 40 composite soil samples (16 organic horizon, 24 mineral layer) were collected and stored in a cool box for transporting within 12–24 h to the laboratory, where they were refrigerated at 4°C before processing.
Extraction of nematodes
The samples were sieved and homogenised using a 2‐mm sieve. One hundred grams of the homogenised soil were used for the nematode extraction with the centrifuge flotation method with sugar (EPPO, 2013). The method allows to isolate nematodes from soil by leveraging their lower density to separate them from heavier soil particles. Soil samples were washed and sieved and then centrifuged in a sugar solution. The suspension containing the nematodes was collected in Erlenmeyer flasks and gradually heated to 60°C and subsequently fixed in formaldehyde (0.27 ml of formaldehyde per 10 ml of suspension). The vials containing the suspension of nematodes were left to rest for 24 h, reduced to 10 ml by siphoning off the extra liquid and kept refrigerated at 4°C for subsequent quantification and identification of nematodes.
Nematode quantification and identification
The quantification of nematodes was made by using Peter's microscope slides and an Olympus CL2 light microscope (×100 magnification). Each sample was manually stirred, and an aliquot of 2 ml of the extraction (20% of the total) was taken for quantification. The nematodes on the slide were counted and the number obtained was multiplied by five to estimate the total abundance per sample. Photographic records were taken by using Olympus Cellsens.
The identification of nematodes was carried out in the Department of Phytopathology and Nematology, School of Agriculture Luiz de Queiroz, University of São Paulo, Brazil. The specimens collected from the Peter's slides were mounted on semi‐permanent microscope slides with 2% formaldehyde for identification under a light microscope (Leica DMLB at ×1000 magnification). Of the eight samples extracted five were randomly drawn for each of the three vegetation types, obtaining a total of 15 samples, which were processed for identifying trophic groups (Yeates et al., 1993). Morphological attributes of the buccal region were used for the taxonomic identification of the specimens to the genus level (Coleman & Wall, 2007). Nematode taxonomic richness and Shannon‐Wiener and Simpson diversity indices at the genus level were quantified for each vegetation type. Abundances were expressed as number of individuals per 100 g soil dry weight.
Data analyses
Since the open vegetation lacked almost completely the organic layer, we focussed the analyses on the mineral layer samples. Differences among vegetation types in total nematode abundance, richness and diversity were determined by carrying out one‐way analyses of variance (ANOVA), while differences in abundance among nematode trophic groups per vegetation type were determined by two‐way ANOVA after testing for normality of the data and homogeneity of the variances. Data were square‐root‐transformed to meet the assumptions of ANOVA. Differences among groups were evaluated with the Tukey test. Differences in nematode community structure among vegetation types were visualised by NMDSs (vegan R package; Oksanen et al., 2013) and assessed both on presence/absence and abundance data of nematode genera with the ADONIS function (vegan) with 10 000 permutations.
The relative abundance of trophic groups in the three vegetation types, and the total number of taxa and the taxa of each trophic group shared among, or unique to each vegetation type were calculated (venndiagram R package; Chen & Boutros, 2011). The distribution of all nematode genera and that of phytophagous genera among vegetation types were analysed using the function ecospatC‐score with 10 000 permutations in the ecospat R package (Di Cola et al., 2017; Broennimann, 2018). The function tests by using a standard null model approach (Gotelli, 2000) for non‐random patterns of co‐occurrence of taxa, that is, whether or not genera co‐occur more or less than expected by chance.
Data manipulation and statistical analyses were carried out using tidyverse (Wickham & Wickham, 2017), ggpubr (Kassambara, 2018), rstatix (Kassambara, 2019), dplyr (Wickham et al., 2020), emmeans (Lenth, 2022) and biodiversityr (Kindt & Coe, 2005) R packages.
Results
Total nematode abundance
Overall mean ± SE total nematode abundance per 100 g dry soil was 557 ± 102. For the open vegetation, it was 835 ± 188, for P. elliottii individuals in the open vegetation, it was 727 ± 165 and for the P. elliottii plantation, it was 108 ± 26 (Fig. 2). Total nematode abundance was significantly lower in the P. elliottii plantation than under open vegetation or under pines established in the open vegetation (ANOVA, F (2,24) = 6.56; P = 0.005; Tables S1, S2; Dataset S1).
Abundance of total nematodes and of phytophagous nematodes in soil (100 g dry weight) and nematode richness at the genus level under native vs. exotic vegetation in montane ecosystems of the Atlantic Forest domain, south‐eastern Brazil. Gr, native open vegetation locally known as campos de altitude; PG, individuals of exotic Pinus elliottii invading native open vegetation; Pi, P. elliottii plantation. Significant differences among vegetation types (one‐way ANOVA) are indicated by different letters (see Supporting Information Tables S1, S2, S4). Boxplots: centre line, median; empty square, mean value; box limits, 25th and 75th percentiles; whiskers, 1.5 times interquartile range; empty circles, outliers; •, P = 0.092.
Nematode richness and diversity
A total of 49 genera were identified (26 in the open vegetation; 28 under P. elliottii colonising open vegetation; 17 in the P. elliottii plantation; Table 1; Dataset S1). Some genera, such as Acrobeles, Aphelenchoides, Helicotylenchus, Tylenchus, and Xiphinema were ubiquitous, whereas others were found exclusively in a single vegetation type. Some plant‐feeding taxa, including the genera Hemicycliophora, Meloidogyne, Discocriconemella, and Rotylenchus, were found in native vegetation but not in the soil in the P. elliottii plantation or under P. elliottii in the open vegetation.
Soil in the P. elliottii plantation had significantly lower values of nematode taxonomic richness than the other vegetation types (ANOVA, F 2,12 = 10.17; P = 0.003; Fig. 2). No differences among vegetation types were found, when comparing either the Shannon diversity (ANOVA, F 2,12 = 0.44; P = 0.657; Fig. S2; Table S1), or the Simpson dominance index values (ANOVA, F 2,12 = 0.86; P = 0.448; Fig. S2; Table S1). The nematode communities in the native vegetation were distinctly different from those under pine individuals invading open vegetation (presence/absence: F = 3.4, R ^2^ = 0.30, P = 0.008; abundance: F = 2.6, R ^2^ = 0.24, P = 0.017) and from those in the pine plantation (presence/absence: F = 2.7, R ^2^ = 0.25, P = 0.008; abundance: F = 2.9, R ^2^ = 0.27, P = 0.008; Figs 3, S3). Nematode communities under pine individuals invading open grasslands differed from those in the pine plantation when considering abundance data (abundance: F = 1.6, R ^2^ = 0.17, P = 0.033; presence/absence: F = 1.4, R ^2^ = 0.15, P = 0.164; Figs 3, S3).
Non‐metric multidimensional scaling (NMDS) ordination plot based on abundance data of soil nematode communities (genus level) in native open vegetation and under exotic Pinus elliottii in montane ecosystems of the Atlantic Forest domain, south‐eastern Brazil. Stress 3D, 0.138; sand colour, native open vegetation; purple, individuals of P. elliottii invading native open vegetation; turquoise, P. elliottii plantation. Ellipses denote 95% confidence intervals using the SE of the weighted average sample scores per vegetation type. Symbols are courtesy of the Integration and Application Network (ian.umces.edu/symbols/), University of Maryland Center for Environmental Science.
The most abundant and frequent nematode taxa were reduced in numbers or absent under pine. We recorded an eightfold decrease in nematode abundance and 2.5‐fold decrease in taxonomic richness in the pine plantation compared with those in native open vegetation. Frequently occurring taxa (i.e. those in > 80% of samples) were reduced by 60% under pine invading open vegetation and were absent altogether in the pine plantation. Samples collected under the pine plantation were much dispersed in the ordination plot, especially compared to those of the open vegetation, because communities were less structured and this contributed to the increased distance among samples in the pine plantation. This was more evident when abundance data were considered (Figs 3, S3).
Open vegetation and pine individuals invading open vegetation shared 11 genera, whereas open vegetation and pine plantation and pine plantation and pine individuals invading open vegetation shared eight genera. There were 12 genera unique to open vegetation, 14 to pine individuals invading open vegetation and 6 to pine plantation (Fig. S4). Phytophagous nematodes were represented by nine genera in open vegetation and under pine individuals invading open vegetation and by seven in pine plantation. There were four genera shared between open vegetation and pine individuals invading open vegetation and three between open vegetation and pine plantation as well as between pine individuals invading open vegetation and pine plantation (Fig. S4). There were four phytophagous genera unique to open vegetation and pine individuals invading open vegetation, while pine plantation had three unique phytophagous genera (Fig. S4).
Nematode trophic groups
Phytophagous nematodes represented the most abundant group in all vegetation types (ANOVA, F 2,72 = 37,42; P ≤ 0.001; Tables S1, S3), except in the P. elliottii plantation where there were no significant differences among trophic groups (Fig. 4).
Abundance of nematode trophic groups in soil (100 g dry weight) under native vs. exotic vegetation in montane ecosystems of the Atlantic Forest domain, south‐eastern Brazil. Gr, native open vegetation; PG, individuals of exotic Pinus elliottii invading native open vegetation; Pi, P. elliottii plantation; Alg, algivore; Bac, bacterivore; Fun, fungivore; Omn, omnivore; Phy, phytophage; Pre, predator. Significant differences among trophic groups (two‐way ANOVA) are indicated by different letters (see Supporting Information Tables S1, S3). Boxplots: centre line, median; empty square, mean value; box limits, 25th and 75th percentiles; whiskers, 1.5 times interquartile range; empty circles, outliers.
Open vegetation had a significantly larger abundance of phytophagous nematodes (470 ± 106 per 100 g dry soil) than the pine plantation (22 ± 7) and soil under P. elliottii in the open vegetation (218 ± 74; ANOVA, F 5,72 = 24.08; P ≤ 0.001; Fig. 2; Tables S1, S4). Soil under native open vegetation and P. elliottii in the open vegetation had significantly larger abundance of phytophagous nematodes when compared to that in the pine plantation (Fig. 2; Table S4). Significant differences among vegetation types were also observed for bacterivores and omnivores; both groups showed lower abundance in pine plantation than in the native open vegetation (Table S4).
The relative abundance of phytophagous nematodes decreased when comparing open vegetation (74.7%) with pine invading open vegetation (68.9%) and with pine plantation (38%; Fig. S5). Other trophic groups showed either increasing values or no changes in their relative abundances from open vegetation to pine invading open vegetation and plantation. The groups that contributed most to these changes were bacterivores (open vegetation, 12.5%; pine invading open vegetation, 15.7%; pine plantation, 37.9%) and fungivores (open vegetation, 3.9%; pine invading open vegetation, 6.4%; pine plantation, 12.4%; Fig. S5). Neither the distribution of all genera considered together nor that of phytophagous nematode genera deviated from random expectations within vegetation types. Standardised effect size ([observed value – mean of the null distribution]/standard deviation of the null distribution) values ranged between −1.25 and 1.38, that is, ≤ |2|, indicating neither spatial aggregation nor segregation, that is, random pattern of taxa co‐occurrence (see Table S5).
Discussion
Our study highlights that P. elliottii, an exotic pine species introduced in the Southern Hemisphere from North America that has been invading open vegetation in the Atlantic Forest and in other biogeographic domains, is associated with a reduced root‐feeding soil nematode abundance compared with that in native open vegetation. Phytophagous nematodes were the most abundant trophic group in all vegetation types, but decreasingly less so under pine individuals invading open vegetation (time 1) and remarkably depauperate in the pine plantation established in the late 1950s (time 2) compared with that in native open vegetation (time 0). The results lead us to suggest that P. elliottii might have escaped from phytophagous pressure in its new range and by modifying soil attributes and nematode communities further reduced the abundance of phytophagous nematodes in its rhizosphere (Reinhart & Callaway, 2006). The impact of P. elliottii on nematode communities was mainly driven by changes in the trophic structure with a drastic depletion of abundant and frequent phytophagous species and overall nematode abundance and richness in the pine plantation.
Our community approach supported one of the premises of the ERH, whereby we found a lower load of phytophagous nematodes under pine individuals in open vegetation than under native vegetation not impacted by pine (H1). The abundance of phytophagous nematodes in the pine plantation decreased twofold compared with that in native open vegetation. In terms of the ERH, the lower abundance of phytophagous nematodes in soils under pine individuals in open vegetation than under native vegetation not invaded by pine could be partly explained by highly species‐specific interactions between nematodes and host plants and/or by the lack of co‐introduced phytophagous nematode species with pine from its native range. When exotic species are not phylogenetically closely related with co‐occurring native species they are likely to experience lower specialist diversity and lower impacts from generalists, as enemies in the invaded range are less adapted to target them (Brian & Catford, 2023). Native vegetation at our study site is generally colonised by arbuscular mycorrhizal fungi and is characterised by a highly diverse and endemic‐rich herbaceous vegetation, shrubs and scattered small trees which are phylogenetically very distant from P. elliottii (Robim & Pfeifer, 1988; Dornelles et al., 2021). Processes of coevolution between specialist nematodes and host plants in these natural ecosystems might therefore have been contributing to plant species‐specificity, resulting in lower phytophagous nematode associations with exotic pines. Additionally, the survival of specialist phytophagous nematodes could be compromised in the absence of the host (Perry & Moens, 2011) which could have led to low abundances of native nematodes under the exotic species. This could have been the case at our study site, where native vegetation cover is progressively suppressed as pine individuals invading open grasslands increase their canopy cover. The lower number of phytophagous nematodes observed in the plantation than under pine individuals invading open grasslands suggests also that specialist enemies were either not co‐introduced with pine or that they did not overcome some biotic and/or environmental filters, conversely to what has been observed for some ECM fungal species that were co‐introduced with pine and became co‐invasive.
Our hypothesis (H2a) that P. elliottii could benefit from the initial reduction of the enemy pressure represented by root‐feeding nematodes in its exotic range and then further reduce their abundance and modifying the composition/structure of soil nematode communities, was confirmed by our results. While the relationship between P. elliottii and the abundance of phytophagous nematodes was related to the stage of invasion of natural ecosystems, the abundance of root feeding nematodes was significantly lower (10‐fold decrease) in the pine plantations (closed canopy pine cover sustained for over 65 yr) than under scattered pine individuals invading open vegetation. This contradicts our alternative hypothesis (H2b) that invasive plants accumulate enemies after establishment (either via co‐invasion of non‐native enemies and/or via adaptation or acclimation of resident enemies; Flory & Clay, 2013; Crous et al., 2016; Dickie et al., 2017b).
Community composition was significantly different among vegetation types and was mainly related to differences in nematode trophic structure, with a major depletion of abundant/frequent phytophagous taxa and overall nematode abundance/richness in the pine plantation. The distributions of nematode communities/phytophagous nematodes within vegetation types did not deviate from random expectations suggesting that the nematode communities, even if significantly different from one another, are mainly structured by stochastic processes. A random pattern of species co‐occurrence has been found in previous studies suggesting that neutral processes are the main drivers of the assembly of nematode communities at the landscape scale and could be attributable to dispersal limitation and soil patchiness (Ettema et al., 2000; Zinger et al., 2019). While some soil microorganisms may be dispersed by wind, the dispersal of larger‐sized soil biota requires active movement, and the dispersal capacity/distance of nematodes is estimated to range from 0.1 to 1 m yr^−1^ (Bardgett & van der Putten, 2014). Being dispersal limited, nematodes are therefore susceptible to environmental heterogeneity at the local scale and related spatial processes, that is, being deterministically associated with environmental heterogeneity, which itself can be rather stochastic.
Nematode communities may be affected by the presence of pine in multiple and nonexclusive ways. The negative relationship between the abundance of phytophagous nematodes and the stage of pine invasion of native vegetation at our study site suggest that P. elliottii might benefit from the reduction of the enemy pressure caused by root‐feeding nematodes and further reduce their abundance over time via positive soil biota – plant feedbacks. Previous research comparing nematode communities in soils under native and exotic pine species has found greater nematode abundance/diversity under native plants than under pines and related it to the availability of resources and microhabitat for nematodes, including soil organic matter and microbial biomass (Yeates & Saggar, 1998; Scholes & Nowicki, 2000; Dehlin et al., 2008). A major reduction in taxonomic and functional diversity has been reported to have resulted in the biotic homogenisation of soil invertebrate communities in plantations of exotic pines and under colonising individuals, established in invaded native grasslands. The homogenisation was associated with increased mineralisation of nitrogen and depletion of soil carbon (Chapela et al., 2001; Dickie et al., 2011; Cifuentes‐Croquevielle et al., 2020) driven by the inhibition of carbon turnover (Araujo & Austin, 2020). A recent study conducted in the vicinity of our study site has quantified soil microbial biomass, soil mineral nitrogen and functional genes involved in the nitrogen cycle in soils in a P. taeda plantation (Muñoz, 2021). The high abundances of the nitrogen fixing nifH and ammonia oxidising bacterial and archaeal amoA genes found in the plantation have suggested that the pine plantation may have been nitrogen limited and the potential great nitrogen demand of the fast‐growing pine plantation could have led, over time, to soils depleted in nutrients, possibly associated with a decrease in nematode diversity/abundance. These pieces of evidence, albeit indirect, would support the hypothesis that reduced resource availability and/or the introduction of novel litter and root exudates under exotic pine species has a negative impact on nematode abundance and diversity.
The abundance of phytophagous nematodes could also be reduced by the presence of defence compounds produced by P. elliottii (‘novel weapon’ hypothesis; Callaway & Ridenour, 2004). Pine species exert different defence responses against the nematode Bursaphelenchus xylophilus (Steiner & Bührer) Nickle in the xylem including phytohormone signalling, secondary metabolism (e.g. terpenoids, phenylpropanoids), oxidative stress, plant defence response and resistance (Modesto et al., 2022). Similarly, one may speculate in the absence of evidence that pine trees might produce defence products against root‐feeding nematodes akin to that reported by Modesto et al. and which might not be produced by native species.
The alteration of native plant communities by exotic pine species is accompanied by changes in belowground diversity (Sapsford et al., 2022) and these processes are likely to affect biogeochemical cycles and ecosystem functioning. The fact that nematode communities were least structured under the pine plantation further supports the positive feedback hypothesis by putting in evidence the disruption of nematode communities by P. elliottii. The reduced taxonomic and functional belowground diversity in ecosystems dominated by exotic pine is likely to reduce the number of trophic interactions both aboveground and belowground, with losses of diversity and multifunctionality (Keith et al., 2009; Zhang et al., 2019).
Our study has two notable limitations. First, nematodes were assessed exclusively in the austral winter, which coincides with low precipitation when compared with the wet summer in south‐eastern Brazil. Seasonality has previously been observed to affect the composition and structure of nematode communities (Girgan et al., 2020) as well as nematode resources including bacteria and fungi (Buscardo et al., 2018, 2024). By taking a one‐time snapshot, we were unable to morphologically identify most juvenile individuals and thus it may be that our results underestimate nematode richness and abundance within trophic groups. Second, nematodes were isolated exclusively from the mineral horizon (the organic layer was almost absent in the open vegetation without pine), and it is therefore likely that overall nematode abundance was underestimated. Nonetheless, our mean total nematode abundance data (4.18 M m^−2^) falls within the range reported by Yeates (2007) for Pinus species (3.25 M to 6.95 M m^−2^).
In conclusion, our study shows that over time, established pine in its exotic range in the Atlantic forest domain, is associated with a low abundance of root‐feeding nematodes, which is likely to be related to: an initial reduced enemy pressure on pine in the early stages of establishment and supporting therefore the premises of the ERH; and a change towards potential positive plant–soil feedbacks in the longer‐term. The findings suggest that both enemy release from phytophagous nematodes and plant–soil feedbacks are likely to contribute, together with pine dispersal capacity and morphological/physiological traits, to the invasiveness of pine species in montane ecosystems in the Southern Hemisphere. Nonetheless, a deeper understanding of plant–soil interactions in response to exotic plant populations to novel belowground communities in the introduced range is needed to be able to project exotic species invasions shaped by interacting global change drivers and their implications for ecosystem processes.
Competing interests
None declared.
Author contributions
LN and EB planned the research; LN and LSCG undertook the fieldwork; LSCG extracted and counted soil nematodes; MMI and LSCG identified nematode genera and/or species using the morphology of adult and juvenile forms; LSCG and EB made the statistical analyses; EB led the writing of the manuscript with input from LN, LSCG and MMI. All authors have read and approved the final version of the manuscript.
Disclaimer
The New Phytologist Foundation remains neutral with regard to jurisdictional claims in maps and in any institutional affiliations.
Supporting information
Dataset S1 Taxonomic identity and abundance of nematodes.
Fig. S1 Sampling area in the Atlantic Forest domain. Fig. S2 Nematode diversity. Fig. S3 Nonmetric multidimensional scaling ordination plot of soil nematode communities. Fig. S4 Nematode genera and trophic groups shared among different vegetation types. Fig. S5 Relative abundance of nematode trophic groups in soils under different vegetation types. Table S1 Results of ANOVA on soil nematode abundance, richness and diversity and on the abundance of nematode trophic groups among and within different vegetation types. Table S2 Results of the Tukey test comparisons of soil nematode abundance and richness among vegetation types. Table S3 Results of the Tukey test comparisons of the abundance of different soil nematode trophic groups within different vegetation types. Table S4 Results of the Tukey test comparisons of the abundance of soil nematode trophic groups among vegetation types. Table S5 Results of a null model analysis to determine the co‐occurrence of total nematode genera and of phytophagous genera among vegetation types.Please note: Wiley is not responsible for the content or functionality of any Supporting Information supplied by the authors. Any queries (other than missing material) should be directed to the New Phytologist Central Office.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1de Abreu RCR , Durigan G . 2011. Changes in the plant community of a Brazilian grassland savannah after 22 years of invasion by Pinus elliottii Engelm. Plant Ecology and Diversity 4: 269–278.
- 2Aldorfová A , Knobová P , Münzbergová Z . 2020. Plant–soil feedback contributes to predicting plant invasiveness of 68 alien plant species differing in invasive status. Oikos 129: 1257–1270.
- 3Anonymous . 2015. Plano de manejo do Parque Estadual de Campos do Jordão, São Paulo, Brasil.
- 4Araujo PI , Austin AT . 2020. Exotic pine forestation shifts carbon accumulation to litter detritus and wood along a broad precipitation gradient in Patagonia, Argentina. Forest Ecology and Management 460: 117902.
- 5Back M , Haydock P , Jenkinson P . 2002. Disease complexes involving plant parasitic nematodes and soilborne pathogens. Plant Pathology 51: 683–697.
- 6Bardgett RD , van der Putten WH . 2014. Belowground biodiversity and ecosystem functioning. Nature 515: 505–511.25428498 10.1038/nature 13855 · doi ↗ · pubmed ↗
- 7Bechara FC , Reis A , Bourscheid K , Vieira NK , Trentin BE . 2013. Reproductive biology and early establishment of Pinus elliottii var. elliottii in Brazilian sandy coastal plain vegetation: implications for biological invasion. Scientia Agricola 70: 88–92.
- 8Blumenthal DM . 2006. Interactions between resource availability and enemy release in plant invasion. Ecology Letters 9: 887–895.16796578 10.1111/j.1461-0248.2006.00934.x · doi ↗ · pubmed ↗