Regulation of Embryonic Wound Healing by Matrix Metalloproteinases in Xenopus laevis Tailbud Stage
Daniel Kraus, Paulina Kikinderova, Pavel Abaffy, Dominika Kadlcikova, Ravindra Naraine, Radek Sindelka

TL;DR
This study explores how matrix metalloproteinases help in scarless wound healing in Xenopus embryos and shows that this process is similar in mammals.
Contribution
The study identifies specific MMPs and the AP-1 stress response as key regulators of embryonic wound healing in Xenopus.
Findings
MMPs 1, 7, 8, and 9 are expressed during the intermediate phase of wound healing in Xenopus.
MMP expression depends on the stress response mediated by the AP-1 transcription factor.
The findings correlate with mammalian wound healing, suggesting a conserved biological process.
Abstract
Scarless wound healing remains a key goal in regenerative research. However, the gene regulatory networks and mechanisms behind this process are still not fully understood, despite the use of high‐throughput analyses and various research models. In our study, we focus on the intermediate phase and examine the role of remodelling genes called matrix metalloproteinases (MMPs) in the Xenopus laevis model. Through temporal bulk RNA‐Seq analysis, we identified the fos/jun (AP‐1) combination expressed during the early phase and four main mmps (1, 7, 8, and 9) expressed during the intermediate phase of healing. Using specific MMP inhibitors and morpholino‐oligonucleotides targeting fos/jun, we found that mmp expression depends on stress response and that they are crucial for embryonic wound healing. We also analysed other published healing data sets (single cell and bulk RNA‐Seq) and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
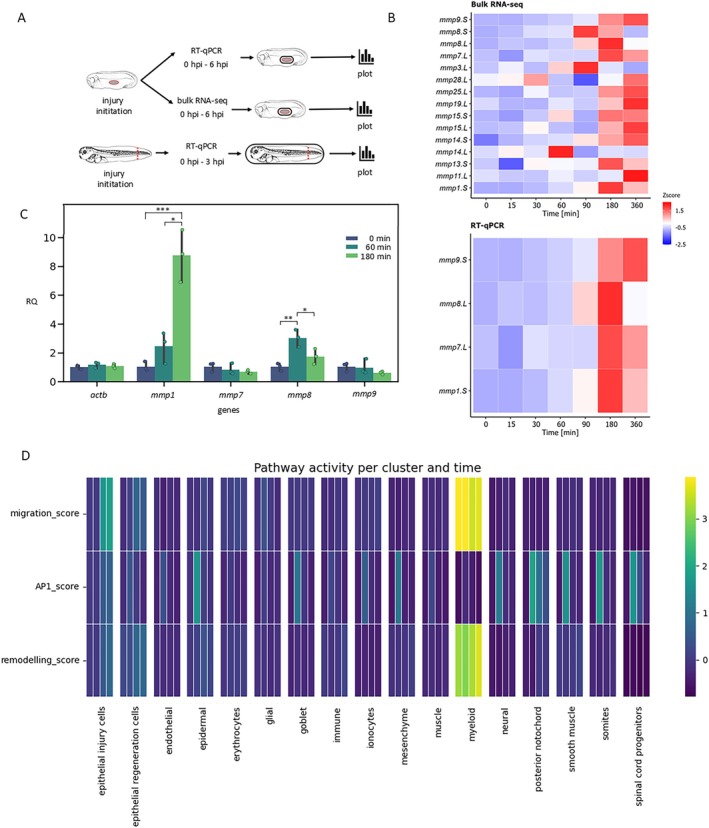 FIGURE 1
FIGURE 1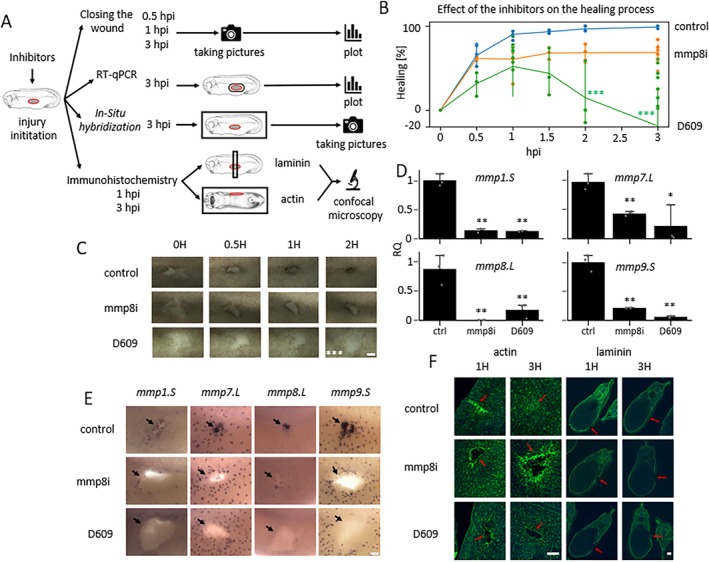 FIGURE 2
FIGURE 2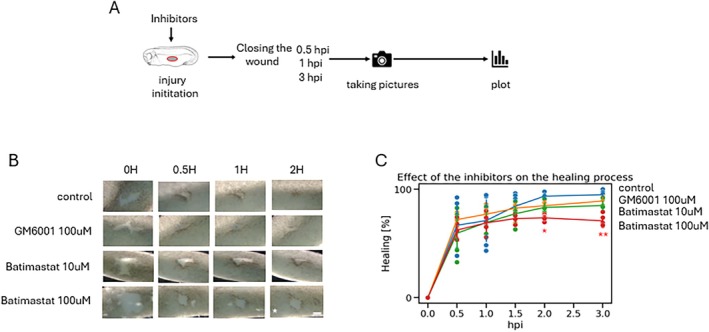 FIGURE 3
FIGURE 3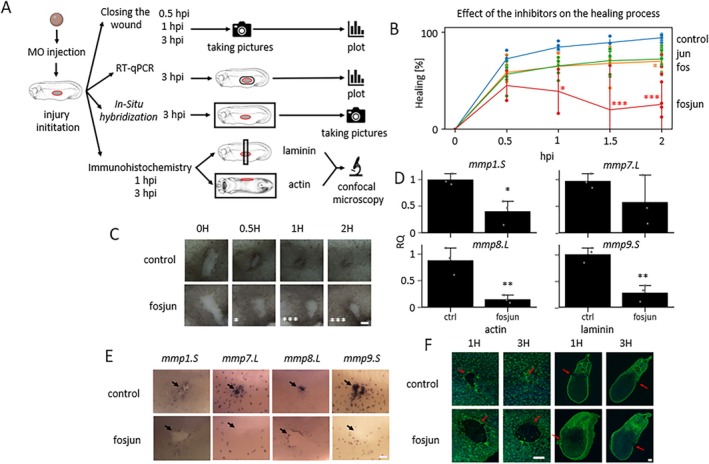 FIGURE 4
FIGURE 4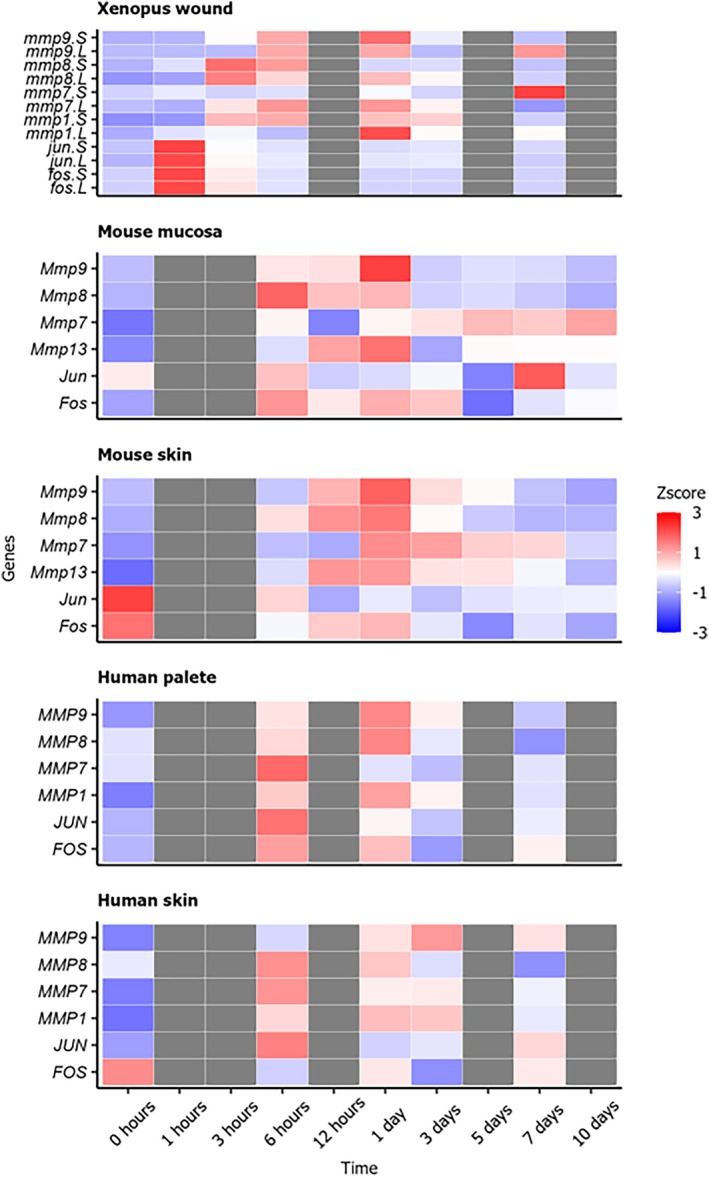 FIGURE 5
FIGURE 5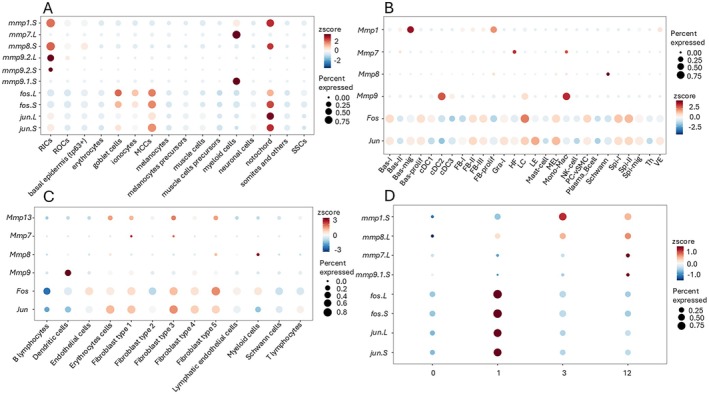 FIGURE 6
FIGURE 6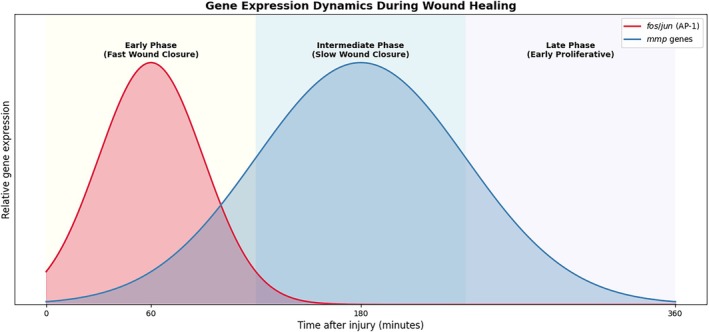 FIGURE 7
FIGURE 7- —Grantová Agentura České Republiky10.13039/501100001824
- —Ministerstvo Zdravotnictví Ceské Republiky10.13039/501100003243
- —Imaging Methods Core Facility at BIOCEV
- —RVO10.13039/100013405
- —MULTIOMICS_CZ (Programme Johannes Amos Comenius, MEYS CR – The European Union
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsWound Healing and Treatments · Protease and Inhibitor Mechanisms · Planarian Biology and Electrostimulation
Introduction
1
Healing processes and their regulation are important areas of study in medical and biological research because they have great potential to improve treatment methods. In this context, healing is defined as the process of restoring skin integrity and homeostasis, which starts immediately after injury [1]. Wound healing is a highly controlled process. We can distinguish two main types: adult and embryonic healing [2]. Understanding these processes is crucial for advancing scientific knowledge and development of new treatments.
Adult wound healing is a complex process that typically takes longer and is generally better understood than embryonic healing. It ends with the formation of a scar, which often has functional and aesthetic limitations. This type of healing consists of four major phases: haemostasis, inflammation, proliferation and remodelling [3]. The first stage, haemostasis, occurs immediately after injury and involves blood vessel constriction along with fibrin clot formation to stop bleeding. During this phase, platelets are important because of their ability to release growth factors that help recruit neutrophils and monocytes [4, 5, 6]. The next stage, inflammation, is characterised by the activity of neutrophils, macrophages, and other immune cells that work together to clear debris and fight infections [2, 3, 4, 5, 6, 7]. During this stage, epithelial cells and fibroblasts become activated [6]. The third stage, proliferation, is characterised by active fibroblasts that promote tissue regeneration. This stage involves the buildup of extracellular matrix components, including fibronectin, proteoglycans and collagen, which replace the initial fibrin matrix. After angiogenesis and epithelialisation, granulation tissue matures during the final stage (remodelling). This last phase results in a scar rich in collagen, which has less strength and functionality compared to uninjured tissue [8].
In contrast to adult wound healing, embryonic wound healing is less complex, faster and scarless [9]. Still, our knowledge about the signalling molecules and regulatory mechanisms involved remains limited. The embryonic healing process that is studied in Xenopus embryos has three phases: early, intermediate and late [10]. The early phase starts immediately after the injury is initiated. It involves several factors such as calcium, nitric oxide (NO) and reactive oxygen species (ROS). These are also crucial for actin‐myosin ring formation as well as resisting infection, cytoprotection and initiating wound healing. The assembly of the actin‐myosin ring at the wound edge assists with wound closure through cell‐specific contractile movement [3, 11]. Healing‐specific gene expression triggers the second, intermediate phase. During this phase, primitive myeloid cells migrate to the healing site and depend on the activity of remodelling enzymes called matrix metalloproteinases (MMPs). The late phase lasts several more hours and involves further remodelling, cell proliferation and differentiation.
Both adult and embryonic healing processes rely on the enzymatic activity of MMPs. Although the functions of these enzymes are well studied in adults, they remain unclear in embryos. Due to several duplication and fusion events in the vertebrates, there are numerous mmp members, which makes it difficult to determine their individual functions (there are 24 mmps in Homo sapiens , 23 in Mus musculus and 20 in Xenopus laevis ) [12, 13]. MMPs/mmps are regulated not only at the level of enzymatic activity but also at the level of their expression [14]. Embryonic expression of mmps is generally low, but their expression increases after injury. In contrast to the relatively late activation of mmps, several signalling pathways such as AP‐1, MAPK, STAT, the Smad family and NF‐κB are activated during early phases, and one of their roles is to stimulate mmp expression [15]. This relationship is encoded in the regulatory sequences of mmps through regulatory elements [16].
A direct relationship between MMPs and AP‐1 in wound healing has already been proposed [17]. AP‐1 is a transcription factor that regulates gene expression in response to various stimuli [18] and controls several cellular processes, including differentiation, proliferation and apoptosis [19]. Jun proteins, which are part of the AP‐1 complex, can form both homo‐ and heterodimers, while Fos proteins only form heterodimers [20, 21]. The Jun/Fos dimer is more stable and has greater transcriptional activity than other combinations [22]. The dimerisation partners of Jun affect its role in gene activation and cell cycle regulation [23].
In this study, we used various tools to reveal the role of mmps in connection with AP‐1 during wound healing in Xenopus laevis embryos. We found that mmp1.S, mmp7.L, mmp8.L and mmp9.S are predominant and necessary for the healing process and play a role in the actin ring formation and ECM remodelling. We used our previously published single cell RNA‐seq and other available mouse and human healing datasets to validate whether these genes are primarily expressed and how they are involved in the wound healing.
Materials and Methods
2
Preparing of Embryos
2.1
Adult Xenopus laevis females were injected with 400 units of HCG hormone to stimulate egg production for the following day. Fertilisation was performed in vitro, and fertilised embryos were cultured in 0.1×MBS solution (Modified Barth's Saline: 8.8 mM NaCl, 0.1 mM KCl, 0.5 mM HEPES pH 7.8, 0.07 mM CaCl_2_, 0.1 mM MgSO_4_, 0.25 mM NaHCO_3_) at 15°C.
Inhibition of MMPs and AP‐1
2.2
We used four chemical inhibitors to target various MMPs with the goal of assessing their role during early healing stages. For each inhibitor, embryos at stage 26 were incubated in the culture medium with the inhibitor at a given concentration for 1 h before injury. A scratch injury measuring 110–130 μm was made on the lateral side of the embryos using fine forceps. The embryos were then incubated in the inhibitory medium at 15°C and fixed at specific time points (1 and 3 h after injury) for immunohistochemistry and RT‐qPCR. Twenty embryos per condition in at least two independent experiments were used in all inhibitory assays.
The first chemical inhibitor D609 (Calbiochem, 251,400) is a selective inhibitor of phosphatidylcholine (PC)‐specific phospholipase C and a broadly expected general MMP inhibitor. It was used at a concentration of 0.02 mM within the culture medium. The second chemical inhibitor, (MMP‐8 Inhibitor I) (Cayman Chemical, 21,852), targets specifically MMP8 activity. It was used at a concentration of 4 μM within the culture medium. The third inhibitor GM6001 (Sigma Aldrich, CC10) was added to the medium at 100 μM. GM6001 is an inhibitor that is a broad‐spectrum MMP inhibitor. It targets the zinc‐binding site in MMP genes. It also effectively blocks MMP activity and it was reported to inhibit MMP 1, 2, 7, 8, 9, 12, 13, 14, 16, 26. The fourth inhibitor Batimastat (Abcam, ab142087) was added to the medium at concentrations ranging from 10 μM to 100 μM. and it is also a broad‐range MMP inhibitor. It inhibits mainly MMP 1, 2, 7, 9 by chelating the zinc ion in the active site of the enzyme.
In addition to the chemical inhibitors, we also utilised morpholino oligonucleotides (MO) to directly target AP1 genes. Morpholino oligonucleotides (MO) were designed and purchased from Gene Tools LLC (Philomath, OR, USA). We used four MOs against fos and jun to target both expression variants (L/S). Sequences were:
fos.S (TTCGGATAATAGACTTACCTGTGGA),
fos.L (AGGTCAGATAATTACCTGCTCCATT),
jun.S (GTAGTTTCCATCTTTGCGTTCATAC),
jun.L (GCTTATGTCAGTGTGACGACACCAA).
Fertilised embryos at the one cell stage were transferred into 0.1× MBS with 3% Ficoll and microinjected with the MO (17 ng) into the animal pole before the first cell division. Embryos were incubated at 15°C (each contained at least 20 embryos per condition). We prepared equimolar mixtures of fos.S and fos.L MOs or jun.S and jun.L MOs. We tested their inhibitory effect using an injection of either fos.S/L or jun.S/L MOs or a combination of fos: jun. We found that a mixture of MOs had a stronger effect than individual MOs, and the most effective was a combination of fos: jun MOs 3:1 in comparison with 1:1 or 2:1, all at the final amount of 17 ng per embryo.
Pictures of the injury were taken with a binocular light microscope and a Nikon camera at specific time points (0, 1, 3 h after injury) to track its closure. The wound's closing speed was analysed in ImageJ (NIH). The wound area was measured and normalised to time zero. Wound‐healing closure data for both chemical inhibitor‐treated and morpholino oligomer‐treated samples were analysed in R (version 4.4.2). A two‐way ANOVA test was used with treatment (chemical inhibitors or morpholino oligomers) and time points as factors, followed by Dunnet's post hoc test to compare each treatment group with the corresponding control at each time point (p < 0.05 *, p < 0.01 ** and p < 0.001 ***).
Immunofluorescence Staining
2.3
Immunohistochemistry was performed according to Sive et al. [24] Actin was stained using Alexa Fluor 488–conjugated phalloidin (1:1000; Life Technologies, A12379). Laminin was detected using a primary anti‐Laminin antibody (1:150; Sigma‐Aldrich, L9393) and a secondary goat anti‐rabbit IgG antibody conjugated to Alexa Fluor 488 (1:500; Life Technologies, A11008). Images were obtained on the Zeiss confocal microscope Carl Zeiss LSM 880 NLO (Carl Zeiss AG) and analysed in ZEN (blue edition) software (Carl Zeiss AG). 10 embryos per condition in two independent experiments were collected and analysed.
RT‐qPCR
2.4
Two experiments were performed. In the first, we measured the levels of mmps only in the wound area by dissecting the healing tissue surrounding the injury at various time points (stage 26). In the second experiment, we determined the levels of mmps in whole embryos to assess injury‐associated changes or to identify mmps whose levels remained unchanged (stage 40). Embryos were collected and dissected during wound healing, and samples were stored at −80°C. Following the standard protocol, total RNA was extracted using TRI Reagent (Sigma‐Aldrich, T9424). cDNA was synthesised from 100 ng of total RNA using MAXIMA H Minus Reverse Transcriptase (Thermo Scientific, EP0751). RT temperature conditions were: 25°C for 10 min, 50°C for 30 min and 85°C for 5 min. qPCR was performed using iQ SYBR Green Supermix (Bio‐Rad, 1,708,880) and C1000 Thermal Cycler CFX384 Real‐time system (Bio‐Rad) with thermocycling conditions: denaturation for 95°C for 3 min, followed by 50 cycles of 95°C for 15 s, 60°C for 20 s and 72°C for 20 s. Melting curve analysis was performed in all experiments to control for PCR specificity. Ten embryos per condition in two independent experiments were collected and analysed.
Gene expression changes during healing at stage 40 and stage 26 were analysed in R (version 4.4.2). Differences between each time point for each specific mmp genes were assessed using one‐way ANOVA followed by Tukey's multiple comparison test (for stage 40) or Dunnet's test (for stage 26) (p < 0.05 *, p < 0.01 ** and p < 0.001 ***).
Bulk RNA‐Seq
2.5
RNA‐Seq data regarding the temporal aspects of embryonic wound healing in X. laevis has already been described by Abaffy et al. [10] and was used for this research. In brief, wounded tissue samples from embryos at stage 26 were collected at 0, 0.5, 1.5, 3, 6 h after the injury. RNA was isolated using TRI Reagent (Sigma‐Aldrich, T9424) following the standard protocol. Libraries were prepared from 200 ng of total RNA using SureSelect Strand‐Specific RNA Library Prep for Illumina Multiplexed Sequencing (Agilent, G9691) according to the manufacturer's protocol. Final libraries were equimolar pooled and sequenced on NextSeq 500 using 2 × 75bp HighOutput mode. The data are available at NCBI's Gene Expression Omnibus under GEO Series accession number GSE116667. The pre‐processed data were extracted and the expression levels of mmp1.S, mmp7.L, mmp8.L and mmp9.S were further analysed.
Human microarray expression data from Leonardo et al., 2023 [25] was downloaded from GEO (accession number GSE209609). In this study, authors used human palate and skin tissues to uncover gene expression during the first 7 days after the injury. Mouse microarray expression data from Chen et al., 2010 [26] was also downloaded from GEO (accession number GSE23006). In this study, authors focused on comparing the transcriptomes of oral mucosal and skin wound to find the differences between these two sites. The downloaded data was checked for previous normalisation by visually inspecting the expression data distribution per sample as a boxplot. The z‐scores of the normalised gene expression were calculated for each gene of interest across the time points and heatmaps of these data were plotted.
Single Cell RNA‐Seq
2.6
Single cell RNA‐Seq data following the temporal aspects of embryonic healing and regeneration in X. laevis has already been done and described by Sindelka et al., 2024 [27]. In brief, single cell experiment was performed using 0, 1, 3, 12 h post injury of X. leavis tadpole tails at stage 42. We expect that early phase of regeneration following tail amputation reflects healing. Healing tissues from 50 embryos were collected per sample. Then the tissue was dissociated into cell suspensions. Sequencing libraries were prepared according to the manufacturer's manual Chromium Single Cell 3′ Reagent Kits User Guide (v 3.1)” and 2400 cells per sample were loaded into Chromium chip. The sample libraries were then pooled and sequenced on Illumina NovaSeq 2000, targeting 100,000 read pairs per cell. The data is available at NCBI's Gene Expression Omnibus under GEO Series accession number GSE245312. For this study the previously pre‐processed and normalised data were used. The pathway activity scoring was performed to visualise pathways activity in cell types. The score is the average expression of a set of genes after subtracting the average expression of a reference set of genes. Score analysis was calculated using scanpy package 1.10.4 (Python 3.11) with its build‐in functions. Expression of mmp9, itgb1, pxn, vcl and actn1 were used for the score of migration. Expression of fos, jun, junb, atf3 were used for AP1 pathway score and remodelling phase (mmp1, mmp2, mmp3, mmp7, mmp8, mmp9, mmp10, mmp13, mmp14) were used for wound healing. Results were visualised in matplotlib 3.10.0 package. Similarly, the Figure S1 was done to show expression of individual mmp genes in different cell types in different time points. The package scanpy 1.10.4 in Python 3.11 was used to calculate and visualise the results with its build‐in functions. Expression of individual mmp genes was calculated in each cell types and each time points and visualised with heatmap.
Mouse single cell RNA‐seq data from Guerrero‐Juarez et al., 2019 [28] (GEO accession number GSE113854) and human single cell RNA‐seq data from Liu et al., 2025 [29] (GEO accession number GSE241132) were downloaded. The data were processed using Seurat package (5.2.1) in R (4.4.2). For mouse single cell RNA‐seq [28], cells that displayed < 8000 UMI/cell and < 2500 genes/cell, and no more than 8% mitochondrial gene expression were kept. Data were then log normalised and scaled followed by Principal Component Analysis (PCA). Then clusters of cells were identified with the use of top 40 PCs and then the dimensionality reduction was performed (t‐SNE). Annotation of the clusters were done using the author's marker genes. The average expression of the genes of interest for each cell cluster was plotted using a dot plot. For the human single‐cell RNA‐seq [29], the author's already preprocessed data was used, including their determined cell clusters and annotations. The data were log normalised and scaled. The average expression of the genes of interest for each cell cluster was plotted using a dot plot.
In Situ Hybridisation
2.7
Whole‐mount in situ hybridisation was used to determine what cells express mmp1, mmp7, mmp8 and mmp9. It was performed according to the protocol of Sive et al. [24] Embryos per each condition (10 embryos for each condition) were fixed in (paraformaldehyde) PFA overnight, dehydrated in methanol and stored in a −20°C freezer. The proteinase K treatment was not used. Samples were imaged on a stereomicroscope (Nikon SMZ 1500) and processed using Zoner Photo Studio 17.
Results
3
High Expression of Mmps During Intermediate Phase of Healing
3.1
Our analysis of the previously conducted bulk RNA‐seq experiment during Xenopus embryonic wound healing (Figure 1A) showed that numerous mmps were expressed in the phase we called intermediate (Figure 1B). At the wound site, mmp1.S, mmp7.L and mmp8.L showed a clear pattern with a peak around 3 h post‐injury, while mmp9.S reached its highest expression at 6 h after the injury (Figure 1B). We confirmed these bulk RNA‐Seq results with independent RT‐qPCR and observed the same expression profiles (Figure 1B).
*mmps are involved in embryonic wound healing. (A) Scheme of experimental procedure. (B) Bulk RNA‐seq data show the expression profiles of mmps important in wound healing and RT‐qPCR validation show the expression profiles of four selected mmps (the graphs show z score of normalised gene expression). (C) Expression of selected mmps in the whole embryos during healing studied in embryos of stage 40 (n = 3) RQ—relative quantity normalised to time 0 (hpi—hours post injury), one‐way ANOVA followed by Tukey's multiple comparison test, p < 0.05 *, p < 0.01 **, p < 0.001 **. (D) Single cell RNA‐seq data show which pathways are active across cell types in different time points in order 0, 1, 3, 12 h.
We continued with determining whether the increased mmp expression (mmp1.S, mmp7.L, mmp8.L, mmp9.S) at the injury site was potentially caused by a global increase in expression throughout the entire embryo or if it resulted from cell‐specific migration to the injury site, leading to a localised expression change (increased at the wound site but unchanged in the whole embryo). To determine this, we performed RT‐qPCR analysis of the observed mmps in the whole embryo (stage 40, time points: 0, 1 and 3 h after the injury) and normalised their levels to those in control (uninjured) embryos. We observed that mmp1 and mmp8 showed an increased expression in the entire embryo, suggesting injury‐associated upregulation, while mmp7 and mmp9 maintained the same levels per embryo (Figure 1C).
Additionally, we analysed our Xenopus single‐cell RNA‐seq data for pathway activity across different cell types (Figure 1D). The results suggested cell roles during wound healing. We can see that epidermal cells that express mmp1.S and mmp8.L genes are not highly active during migration, and in addition, their expression level is minimal at time 0, suggesting expression activation following injury. We can observe that these cells show an injury‐associated increase in abundance and are enriched at the wound site during wound healing. In contrast, mmp7.L and mmp9.S are highly expressed genes primarily in the myeloid cells. Previous literature suggested their active migration to the injury site [30, 31, 32] and our scoring parameters also show increased migration and remodelling features. Similar results are shown in the Figure S1. This figure shows the expression of mmp1.S, mmp7.L, mmp8.L, and mmp9.S in different cell types across different time points. Our results suggest that mmp1.S and mmp8.L display localised expression in injury‐related epidermal cells, and mmp7.L and mmp9.S are mainly expressed in migratory myeloid cells.
Mmps Are Necessary for Embryonic Wound Healing
3.2
The mmps expression peak at 3–6 h post‐injury suggests their relevance during intermediate and late phases of healing. We performed transient MMPs inhibitory experiments to study their roles using either the global MMP inhibitor D609 or the MMP8 specific inhibitor (mmp8i) (Figure 2). Both inhibitors led to a weaker wound closure relative to the control (Figure 2B,C). Control embryos closed 97.3% of their wounds in 3 h, in contrast to the embryos treated with D609 inhibitor (18.8%) and the embryos with mmp8 inhibitor (68.6%) (Figure 2B).
*The effect of the MMP inhibition during wound healing. (A) Scheme of the experimental procedure. (B) Quantification of wound closure in time (x axis) calculated as percentage normalised to time 0—Injury (y axis), n = 3–13 per condition, two‐way ANOVA followed by Dunnet's post hoc test, p < 0.05 *, p < 0.01 **, p < 0.001 ***. (C) Brightfield images of wound assays from the representative embryos (scale bar 100 μm). (D) Expression of mmps in wound site studied using RT‐qPCR in 3 h after the injury (control n = 3, mmp8i n = 3, D609 n = 2), one‐way ANOVA followed by Dunnet's post hoc test, p < 0.05 *, p < 0.01 **, p < 0.001 **. (E) Visualisation of selected mmps expression by in situ hybridisation in representative embryos in 3 h after the injury (n = 10, scale bar 50 μm). (F) Actin ring formation defect (phalloidin staining) and problem of basal lamina formation showed with red arrow (Laminin staining) in 3 h after the injury (n = 10, scale bar 100 μm). (mmp8i—Inhibitor of mmp8).
We also determined the expression levels of other mmps in the wound area following inhibition. We detected a significantly decreased expression (Figure 2D). The same decrease of mmps was also observed when using in situ hybridisation. Application of the MMP8 inhibitor led to low expressions of mmp1.S and mmp8.L around the wound and a migratory defect of the myeloid cells, as shown using mmp7.L and mmp9.S staining (Figure 2E). The same pattern was observed after application of the D609 inhibitor (Figure 2E).
Actin‐myosin ring is usually formed around the edges of the wound and helps the wound to close by constricting along the opposite sites. Its formation is crucial for successful healing initiation. We performed immunohistochemistry of actin to visualise its formation. Control embryos showed a clear actin‐myosin ring formed that pulled together at the late healing phase. In contrast, inhibited embryos showed poor actin‐myosin ring formation for both inhibitors and the absence of wound closure after 3 h (Figure 2F). Remodelling of the extracellular matrix (ECM) is also an essential step for healing, and it depends on MMP activity. We performed Laminin immunohistochemistry to visualise basal lamina remodelling following injury. The Laminin layer was reformed at the wound site in control embryos versus no reformation in both the inhibited embryos (Figure 2F).
To validate specificity of mmps for healing, we used two others widely used mmp inhibitors GM6001 and Batimastat. Both inhibitors showed reduced wound closure, especially during the intermediate healing phase (Figure 3B,C). Wound closure for GM6001 remained closed for about 89.2% after 3 h post injury and for Batimastat 84.9% for 10 μM and 70.9% for 100 μM concentrations.
*Inhibition of wound healing with broad range inhibitors. (A) Scheme of the experimental procedure. (B) Brightfield images of wound assays from the representative embryos (n = 5, scale bar 100 μm). (C) Quantification of wound closure in time (x axis) calculated as percentage normalised to time 0—injury (y axis), n = 5 per condition, two‐way ANOVA followed by Dunnet's post hoc test, p < 0.05 *, p < 0.01 **, p < 0.001 **.
AP‐1 Is Required for Mmps Expression
3.3
As discussed above, mmps expression appears between 3–6 h post injury (intermediate phase). However, an earlier wave of gene expression occurs around 1 h. This earlier wave includes genes required for metabolic changes and stress response. Stress is usually accompanied by the expression of AP‐1 components such as jun and fos genes. Previous literature has already suggested that there is a connection between AP‐1 and mmps, and our goal was to determine whether it is also the case during embryonic healing.
We used MOs to inhibit two well‐known AP‐1 genes: fos and jun. We know from bulk RNA‐Seq that both are highly expressed during the early healing phase [10]. We targeted, through injections, a single gene (L/S) or a combination of different genes, followed by wound assay, and found that it decreased the healing capacity. Individual injection of fos or jun MO led to wound closure at the level of 70% and 72.3%, respectively (Figure 4B). However, a combination of fos: jun MOs at a concentration of 3:1 showed a much higher effect of reducing the ability of closure (25.6%) (Figure 4B,C). We performed an RT‐qPCR experiment to quantify the level of mmps expression dependence on AP‐1 activity. AP‐1 loss of function embryos showed significantly decreased expression of mmp1.S, mmp8.L and mmp9.S, while no significant decrease for mmp7.L (Figure 4D). We validated these results by in situ hybridisation and found reduction of staining at the wound site for mmp1 and mmp8 and myeloid cell defect using mmp7 and mmp9 staining (Figure 4E). In addition, AP‐1 loss of function embryos showed also defects in actin ring formation and Laminin layer formation (Figure 4F).
*Importance of AP‐1 during embryonic wound healing. (A) Scheme of the experimental procedure. (B) Quantification of wound closure in AP‐1 loss of function embryos in time (x axis), calculated as percentage normalised to time 0 (n = 2–14), two‐way ANOVA followed by Dunnet's post hoc test, p < 0.05 *, p < 0.01 **, p < 0.001 ***. (C) Brightfield images of wound assays in AP‐1 loss of function. Showed are representative embryos (n = 10, scale bar 100 μm). (D) The effect of AP‐1 loss of function on mmps expression studied by RT‐qPCR at 3 h after the injury (n = 3), one‐way ANOVA followed by Dunnet's post hoc test, p < 0.05 *, p < 0.01 **, p < 0.001 **. (E) Validation of mmps expression using in situ hybridisation (n = 10, scale bar 50 μm). (F) Actin ring formation defect (phalloidin staining) and the problem of basal lamina formation showed with the red arrow (Laminin staining) (n = 10, scale bar 100 μm). (fosjun—Morpholino targeted the combination of fos and jun genes).
Expression of Mmp Genes Showed Similarities Among Models
3.4
To study the conservation of wound healing regulation between our Xenopus model and mammalian healing datasets, we utilised available microarray studies from mouse [26] and human studies [25]. Expression of AP‐1 genes (fos and jun) showed similar rapid increase during early healing phase and then decrease in later phases of healing. Surprisingly, the maximum level of both genes was observed at time 0 in mouse and human skin samples compared to 1 h post injury in Xenopus. The same peak at 1 h post injury was found in mouse mucosa and human palate samples. Good correlation was observed among mmp genes during intermediate phase of healing. They showed peak expression between 3 h and 1 day post injury followed by decrease of expression in the later phases in all studied tissues and models (Figure 5).
Comparison of mmps and AP‐1 genes expression studied from microarray datasets. Xenopus, mouse and human datasets were reanalysed and showed similarities during early phase (AP‐1 signalling) and intermediate phase (mmps). In the mouse, Mmp13 serves as an equivalent to Mmp1.
Gene Expression of Mmps and AP‐1 Genes Showed Similarity Also at Cellular Level
3.5
We compared the average expression of the genes of interest at the individual cell cluster level to reveal differences and similarities between Xenopus and mammalian healing models. Recently, we published a single cell analysis of regeneration in Xenopus, where we focused on the early regeneration phase, which in many aspects relates to wound healing [27]. In Xenopus, we found mmp1.S and mmp8.L to be expressed in a sub‐group of epidermal cells (called RICs) and notochord cells, while mmp7.L and mmp9.S to be expressed in myeloid cells. Genes coding fos and jun showed broader expression with primary expression in skin cells (goblet, ionocytes and multicilliated cells) and notochord (Figure 6A). Human single cell analysis of wound healing [29] showed broad expression of fos and jun and cell‐specific expression of Mmps (Mmp1 in skin (keratinocyte) population); expression of Mmp7 and Mmp9 in myeloid cells (dendritic, monocytes and macrophage cells) (Figure 6B). Mouse single cell analysis of wound healing [29] showed again expression of fos/jun in many cell types, in contrast to specific expression of Mmps. Mmp13, which is supposed to substitute Mmp1 in mouse [28, 29] and Mmp7 were expressed in fibroblasts, Mmp8 and Mmp9 were predominant in myeloid and dendritic cells, respectively (Figure 6C).
Expression of AP‐1 and mmp genes studied at single cell level. (A) Single cell RNA‐seq from Xenopus healing (0–12 h post injury) showed expression of mmps in epidermal and myeloid cells and broader expression of fos/jun. (RIC—regeneration initiating cells, ROC—regeneration organising cells, MCCs—multicilliated cells, SSCs—secretory cells). (B) Human single cell study (intact skin, 1 day, 7 days and 30 days post injury) of wound healing showed broad expression of AP‐1 and cell specific expression of Mmps. (DC‐ dendritic cells, HF—heart failure cells, LC—langerhans cells, LE—lupus erythematosus cells). (C) Mouse single cell study (12 days post injury) of wound healing showed broad expression of AP‐1 and cell specific expression of Mmps. (D) Temporal average expression of genes of interest from our Xenopus single cell regeneration study (time 0, 1, 3 and 12 h).
We also checked the average expression of the genes of interest across the analysed time points from our Xenopus single cell data to confirm our observation of injury‐induced increase in expression of mmp1 and mmp8 in contrast to stable expression of mmp7 and mmp9 (Figure 6D). Both AP‐1 genes showed increased expression at 1 h post injury, mmp1 and mmp8 showed increase of expression post injury at 3 and 12 h. In contrast, mmp7 and mmp9 expression remained stable during 1 and 3 h, and an increase could be seen at 12 h, corresponding to the potential of myeloid cell migration to the wound area.
Discussion
4
Wound healing is an interesting biological process. Although the embryonic type appears simple, it is complex and involves many phenotypic changes in surrounding cells and broad gene expression activity. Several waves of new gene expression can be observed, with the first early wave occurring 30 to 60 min after injury. This initial wave is characterised by stress response genes [33], including the AP1 pathway, mainly composed of jun and fos genes. These early response genes are essential for cell migration and re‐epithelialisation [34, 35] and help promote the expression of additional genes with AP‐1 binding sites, such as keratins and matrix metalloproteinases [36, 37, 38, 39]. The role of AP1 has also been studied during later developmental stages of Xenopus and was found to be important for healing and regeneration [40, 41]. The early gene expression wave is supported by actin‐myosin ring formation and rapid wound constriction.
The second wave of gene expression occurs 3 to 6 h after injury [10] and is characterised by a defence response, activation of cellular processes such as proliferation or cell death, and remodelling of the ECM. MMPs are likely key enzymes involved in ECM remodelling in Xenopus embryos. This is also observed in Danio rerio , which also uses mmp9 expression to change the ECM. mmp9 may also influence cytokine signalling during healing [42].
Based on our Xenopus bulk RNA‐Seq data, at least 15 mmp genes are involved in wound healing. We selected the four most prominent mmps: mmp1, mmp7, mmp8 and mmp9 for further analysis. We observed an increase of these mmps around the 3 h post injury at the site of the wound. Similarly, in mouse and human healing datasets, these mmps also showed an increase in their expression at the wound site following injury. In Xenopus embryos, mmp7 and mmp9 are expressed in myeloid cells, which migrate to the wound area [43]. In contrast to myeloid cell migration and accumulation of mmp7 and mmp9 after injury, we found that in Xenopus embryonic healing, the mmp1 and mmp8 are expressed in epithelial cells around the wound area. We also observed that mmp genes are responsible for closing the wound and remodelling the Laminin layer.
The cross‐species conservation of these mmps expression after injury may be explained by their functional roles. The role of Mmp1 during wound healing was shown in mammals to accelerate epithelisation, reduce inflammation and increase vascularisation. It also reduces the scar formation [44, 45]. The expression of Mmp1 in the basal keratinocytes in the front part of the wound is triggered by the loss of ECM [46]. It was also recently suggested to be substituted by the activity of Mmp13 in mouse [29]. A strong increase in Mmp8 has also been documented in other healing models [47], and it has been indicated that Mmp8 contributes to inflammation [48]. It has been shown that Mmp7 in mammals is involved in re‐epithelialisation. It also cleaves ECM or ECM‐associated proteins and facilitates keratinocyte migration. Mmp9 is in mammal wound healing crucial for collagen reorganisation and keratinocyte migration [12].
Recent single cell studies in mammals also showed differences among expression of mmps during healing. Surprisingly, human wound healing reflects a similar wound‐healing process as is in Xenopus, but mouse healing differs. For example, myeloid (immune) cells express mmp7/Mmp7 and mmp9/Mmp9 in both human and Xenopus, but mouse study showed rather the expression of a combination of Mmp8 and Mmp9 in immune cells.
Axolotl has more mmp genes than is typical for most vertebrates (including humans). Some mmp genes are exclusive for the salamander lineage only [49]. As well as Xenopus, they both have more mmp genes, which may lead to better healing ability. The expression levels of some mmps, such as mmp2 and mmp9, persist long after injury and support their important role in matrix remodelling [50]. Other studies focusing on the importance of the healing process showed the involvement of additional mmps which were not enriched in Xenopus embryos. The expression of mouse Mmp3 was expressed in proliferating cells adjacent to the wound edge and was described as important for wound contraction [51]. Also, the expression of Mmp10 was found to colocalise with Mmp1 in healing mammalian epidermis. A decrease of Mmp10 leads to a disorganised migrating epithelium, degradation of newly formed matrix, aberrant cell–cell contacts of the migrating keratinocytes, and an increased rate of cell death of wound edge keratinocytes [52]. We assume that the wide range of remodelling enzymes with likely species‐specific activation will lead to significant challenges in studies trying to understand the individual healing mechanisms.
In summary, our study showed that the combination and dependence of AP‐1 and mmps expression are crucial for embryonic wound healing. Based on recent single cell studies in human [29] and Xenopus [27], we can also speculate about more complex regulation of AP‐1 and downstream mmps. Study of healing in Xenopus revealed potential regulation of wound specific mmps and ECM components controlled by cell co‐expression of junb and fosl1. In human healing study, authors found similar master regulator FOSL1 to be required for efficient wound healing. Taken together, AP‐1 and mmps are responsible for initiating wound closure and tissue remodelling (Figure 7). The disruption of mmps expression causes the incompetence of myeloid cells, which migrate to the wound area and prevent the formation of the actin‐myosin ring in the edge epithelial cells, resulting in a wound closure defect.
Summary of AP1 and mmps roles during embryonic wound healing.
Conclusion
5
We were able to provide the first detailed temporal analysis of specific mmp genes (mmp1.S, mmp7.L, mmp8.L and mmp9.S) in wound healing in Xenopus laevis embryos showing their importance. We also demonstrated that mmp genes functionally depend on the early AP‐1 activation pathway. These results support the connection of early stress responses directly to ECM remodelling enzymes. Our integration of bulk RNA‐seq and single cell RNA‐seq confirms that mmps expression patterns observed in Xenopus laevis are conserved at the cellular level also in mammalian wound healing. This shows potential in therapeutic targeting.
Author Contributions
Daniel Kraus: conceptualisation, methodology, investigation, validation, formal analysis, data curation, data analysis, writing – original draft, visualisation; Paulina Kikinderova: investigation; Pavel Abaffy: investigation, analysis, writing; Dominika Kadlcikova: investigation; Ravindra Naraine: data analysis, writing; Radek Sindelka: conceptualisation, methodology, validation, writing – original draft, visualisation, supervision, funding acquisition.
Funding
This work was supported by Grantová Agentura České Republiky (24‐12027S), Ministerstvo Zdravotnictví Ceské Republiky (NW24‐03‐00459), Imaging Methods Core Facility at BIOCEV (LM2018129, MEYS CR), RVO (86652036), MULTIOMICS_CZ (Programme Johannes Amos Comenius), MEYS CR – The European Union, (CZ.02.01.01/00/23_020/0008540).
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Figure S1: Expression of mmp genes in different cell types. Single cell RNA‐seq data show which mmp genes are expressed in which cell type
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1P. Martin , “Wound Healing–Aiming for Perfect Skin Regeneration,” Science 276, no. 5309 (1997): 75–81.9082989 10.1126/science.276.5309.75 · doi ↗ · pubmed ↗
- 2B. J. Larson , M. T. Longaker , and H. P. Lorenz , “Scarless Fetal Wound Healing: A Basic Science Review,” Plastic and Reconstructive Surgery 126, no. 4 (2010): 1172–1180, 10.1097/PRS.0b 013e 3181 eae 781.20885241 PMC 4229131 · doi ↗ · pubmed ↗
- 3S. Bunman , N. Dumavibhat , W. Chatthanawaree , S. Intalapaporn , T. Thuwachaosuan , and C. Thongchuan , “Burn Wound Healing: Pathophysiology and Current Managementof Burn Injury,” Bangkok Medical Journal 13, no. 2 (2017): 91.
- 4W. G. Owen , C. T. Esmon , and C. M. Jackson , “THE Conversion of Prothrombin to Thrombin: I. CHARACTERIZATION OF THE REACTION PRODUCTS FORMED DURING THE ACTIVATION OF BOVINE PROTHROMBIN,” Journal of Biological Chemistry 249, no. 2 (1974): 594–605.4809530 · pubmed ↗
- 5D. M. Monroe and M. Hoffman , “What Does It Take to Make the Perfect Clot?,” Arteriosclerosis, Thrombosis, and Vascular Biology 26, no. 1 (2006): 41–48.16254201 10.1161/01.ATV.0000193624.28251.83 · doi ↗ · pubmed ↗
- 6G. S. Schultz , G. A. Chin , L. Moldawer , and R. F. Diegelmann , “Principles of Wound Healing,” in Mechanisms of Vascular Disease: A Reference Book for Vascular Specialists [Internet] (University of Adelaide Press, 2011).
- 7P. Cowled and R. Fitridge , “Pathophysiology of Reperfusion Injury,” in Mechanisms of Vascular Disease: A Reference Book for Vascular Specialists [Internet] (University of Adelaide Press, 2011).30484990 · pubmed ↗
- 8N. X. Landén , D. Li , and M. Ståhle , “Transition From Inflammation to Proliferation: A Critical Step During Wound Healing,” Cellular and Molecular Life Sciences 73 (2016): 3861–3885.27180275 10.1007/s 00018-016-2268-0PMC 5021733 · doi ↗ · pubmed ↗