Development of a non-invasive diagnostic method for pathogenic RNA viruses using sebum wiped from the cat’s body surface
Yuri V Fukushima, Namiko Saito, Hirohisa Mekata, Akatsuki Saito

TL;DR
Researchers developed a non-invasive method to detect RNA viruses in cats using sebum from their ears, offering safer diagnostics for both animals and humans.
Contribution
The study introduces a novel sebum-based diagnostic method for feline RNA viruses, including SFTSV, with minimal invasiveness and high sensitivity.
Findings
Feline immunodeficiency virus RNA was reliably detected from ear sebum with sensitivity comparable to blood tests.
SFTSV RNA was successfully detected from sebum samples of infected cats.
The ear was identified as the optimal site for sebum collection for RNA virus detection.
Abstract
The development of non-invasive diagnostic methods for zoonotic viral infections is important for animal welfare and public health. Sebum-based diagnostic methods using commercial oil-blotting films have been used for SARS-CoV-2 detection in humans, but similar strategies for veterinary use remain unexplored. Severe fever with thrombocytopenia syndrome (SFTS), caused by the SFTS virus (SFTSV), presents a major health threat in Asia—especially in Japan, where multiple cases of cat-to-veterinarian transmission have been reported. To address this need for safer diagnostics, we sought to establish a sebum-based RNA virus detection method for cats. We designed primers that efficiently detected RNA from feline sebum while distinguishing it from human and feline DNA/RNA. Using this platform, we deemed the ear to be the optimal sebum collection site and confirmed that feline immunodeficiency…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
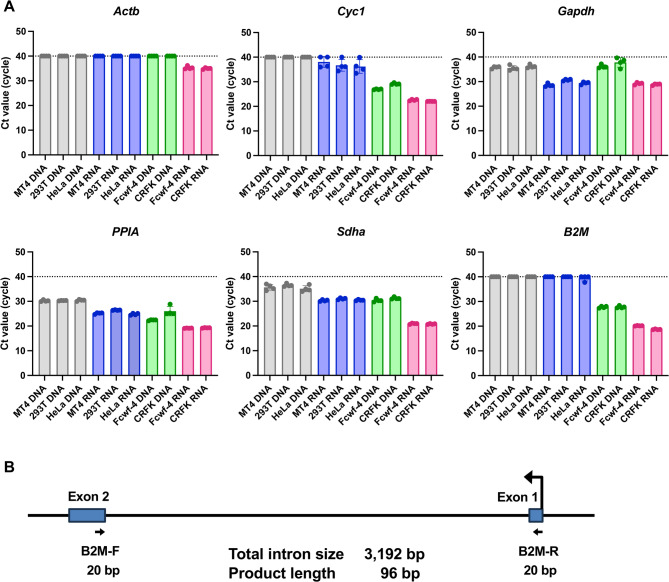 Figure 1
Figure 1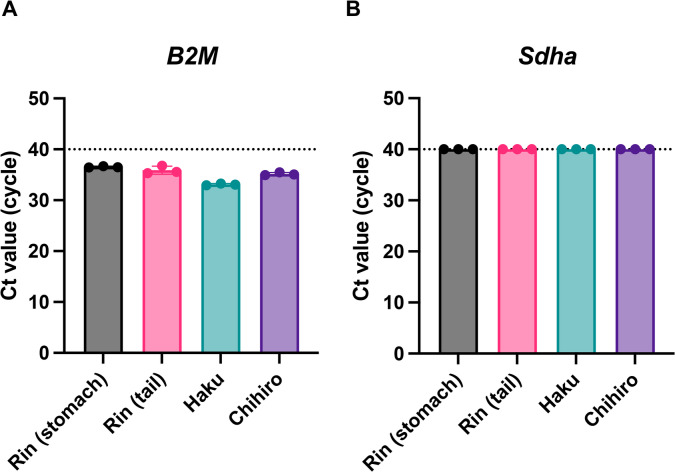 Figure 2
Figure 2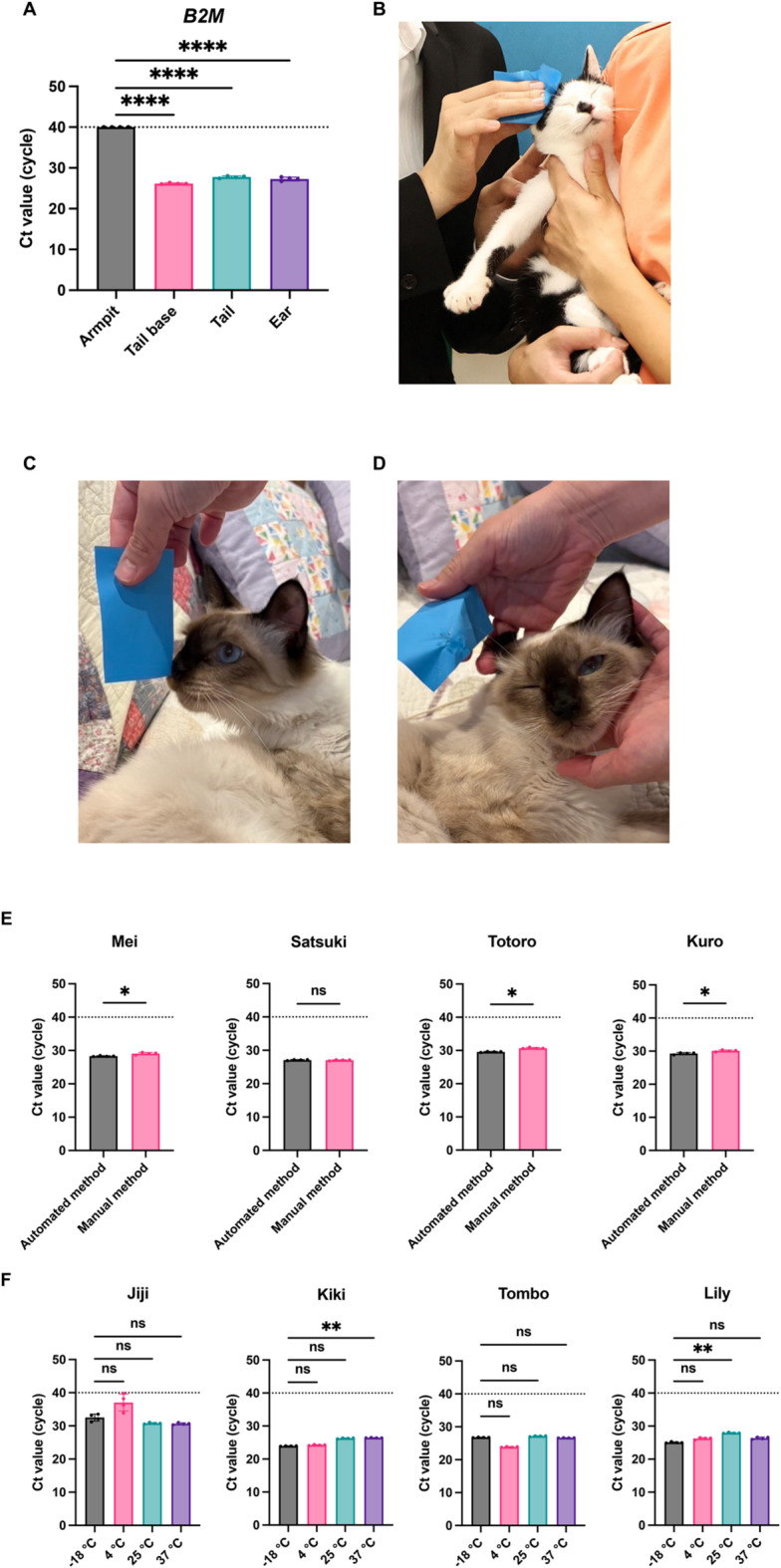 Figure 3
Figure 3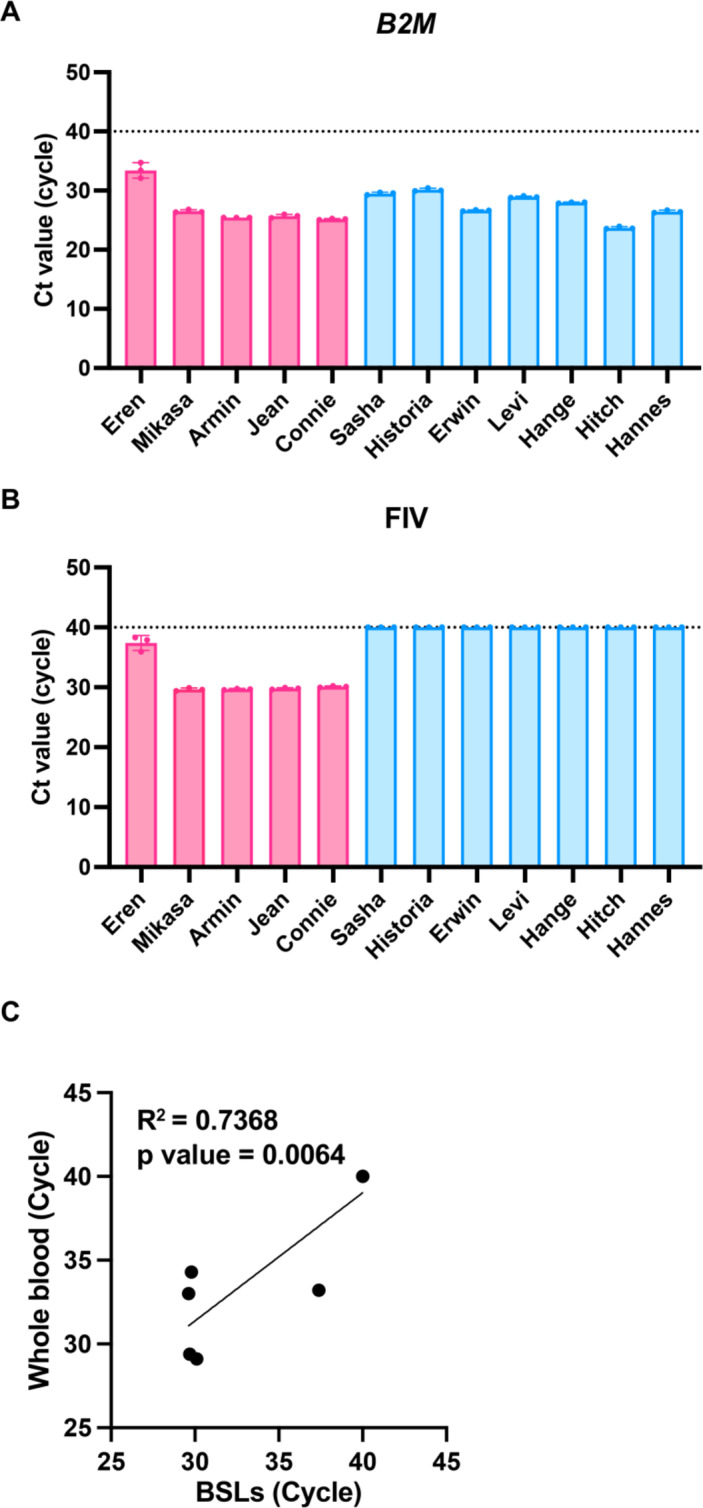 Figure 4
Figure 4 Figure 5
Figure 5- —https://doi.org/10.13039/501100001691Japan Society for the Promotion of Science
- —https://doi.org/10.13039/501100008040Kao Corporation
- —https://doi.org/10.13039/100009619Japan Agency for Medical Research and Development
- —G-7 Grants
- —Ito Foundation Research Grant
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRabies epidemiology and control · Viral Infections and Outbreaks Research · Viral Infections and Vectors
Introduction
Zoonotic diseases remain a pressing global health challenge, with numerous ribonucleic acid (RNA) virus infections—including severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), severe fever with thrombocytopenia syndrome (SFTS), Nipah virus infection, and rabies—continuing to pose serious worldwide threats^1–4^. Prompt and accurate diagnosis is essential for controlling these infections. Currently, the standard diagnostic approach relies on detecting viral RNA from blood samples. However, this method places a considerable burden on patients due to the pain and anxiety of venipuncture, as well as the risk of bleeding in individuals with hemorrhagic disorders. In addition, handling blood samples exposes healthcare personnel to hazards of contracting the target virus and other blood-borne pathogens, including human immunodeficiency virus (HIV)^5,6^ and hepatitis B virus (HBV)^7^, which represent serious occupational concerns.
Although less invasive diagnostic methods, such as saliva-based testing for SARS-CoV-2 infection, have been introduced^8^, their use is still confined to a limited set of infectious diseases. Consequently, there is an urgent need for novel approaches that are both less invasive for patients and safer for healthcare providers. In response, efforts have been made to develop blood-free testing methods for RNA virus detection. A recent study successfully demonstrated the presence of SARS-CoV-2 RNA in facial sebum using RNA monitoring technology^9^. This method collects sebum with commercially available oil-blotting films and analyzes the extracted mRNA. Sebum-based diagnostics thus represent a promising and minimally invasive strategy, with considerable potential for refinement and broader application.
SFTS, caused by the SFTS virus (SFTSV), is a tick-borne zoonotic disease with a high fatality rate. It is currently endemic in several Asian countries, including Japan, China, South Korea, and Thailand^10^. While tick bites represent the main route of infection^11^, increasing evidence has revealed that human-to-human and^12–14^ animal-to-human^15–17^ transmission also occur as infection routes. SFTSV has been detected at high concentrations in multiple body fluids, including blood, saliva, and urine^18^.
Cats, as close companion animals to humans, require protection from viral infections for both feline and public health. Infected cats also pose transmission risks to veterinarians and animal healthcare workers. Several studies have shown that cats are highly susceptible to SFTSV^18^, raising concerns not only about cat-to-human transmission but also about occupational exposure during routine veterinary practices such as blood collection. Indeed, fatal cases of cat-to-human SFTSV transmission have been reported in Japan^19^. Therefore, developing safer diagnostic methods for cats is of critical importance. Building on sebum-based detection methods established in humans, we hypothesized that a diagnostic method with oil-blotting films could provide a less invasive option for cats while also reducing infection risks for veterinarians and animal nurses.
In addition to SFTSV, we also examined feline immunodeficiency virus (FIV), a major RNA virus that affects cats worldwide [reviewed in^20^]. FIV is among the most common and clinically consequential feline infectious diseases, particularly in outdoor cat populations^21^. Transmission occurs primarily through bite wounds inflicted by infected cats^22^. Following infection, FIV targets the immune system, rendering cats more susceptible to secondary infections [reviewed in^23^]. Although many FIV-positive cats remain asymptomatic for extended periods^24,25^, immune suppression increases vulnerability to opportunistic infections caused by otherwise harmless microorganisms [reviewed in^23^]. FIV poses particular challenges in animal shelters, where rescued cats are frequently housed in close proximity, thereby increasing the risk of transmission^26^. Beyond direct medical concerns, FIV status also influences adoption outcomes, as households with uninfected cats may be reluctant to adopt FIV-positive animals. Given these considerations, early and efficient detection is essential for prevention, and the sebum-based diagnostic method proposed in this study offers substantial potential to address these needs.
In this study, we aimed to establish a diagnostic method for detecting feline RNA virus infections using oil-blotting films. We successfully detected FIV RNA in sebum samples and demonstrated that the sensitivity of this sebum-based method was comparable to standard blood-based testing. In addition, we detected SFTSV RNA using sebum collected with oil-blotting films. These results suggest that sebum sampling offers a promising, minimally invasive approach for diagnosing viral infections in cats, improving veterinary safety and lowering the risk of cat-to-human transmission.
Results
Design and evaluation of primers for the specific detection of cat housekeeping gene mRNA from body surface lipids (BSLs)
To confirm that the oil-blotting films collected sufficient BSLs, we designed primers targeting cat housekeeping gene mRNA with high sensitivity and specificity. Because films might be contaminated by examiners handling them without gloves, we also sought to develop primers capable of distinguishing cat RNA from human DNA or RNA. Genomic DNA and total RNA were extracted from human- and cat-derived cell lines, and primers for six housekeeping genes were evaluated by quantitative real-time reverse transcription polymerase chain reaction (qRT-PCR). Among these, primers targeting Sdha and B2M consistently produced threshold cycle (Ct) values at least eight cycles lower for cat RNA than for human DNA/RNA or cat genomic DNA (Fig. 1A). Both Sdha and B2M also showed relatively low overall Ct values, making them suitable for practical use (Supplementary Table 1). Based on these results, Sdha and B2M were selected for further testing.
Fig. 1. Evaluation of primers for selective amplification of the cat housekeeping gene mRNA. (A) Comparison of Ct values obtained from human genomic DNA (gray), human RNA (blue), cat genomic DNA (green), and cat RNA (red). Data show the mean ± standard deviation (SD) of quadruplicate measurements and represent at least four independent experiments. (B) Schematic of primer design. Blue boxes represent exons, arrows indicate primer sites, and F and R mark forward and reverse primers, respectively.
We next examined whether transcripts of Sdha and B2M could be detected from BSLs using the designed primers (Fig. 1A). Four oil-blotting film samples obtained from three cats were analyzed. B2M was successfully detected with an average Ct value of 36.2 (Fig. 2A), whereas Sdha was not detectable from BSLs (Fig. 2B). On this basis, B2M was chosen as the optimal primer, and a probe was subsequently designed for B2M to enable probe-based detection (Fig. 1B, Supplementary Table 2).
Fig. 2. Detection of cat housekeeping gene mRNA from body surface lipids (BSLs). Four BSL samples were collected from three cats. Data show the mean ± SD of triplicate measurements from one assay, representative of at least three independent experiments.
Optimization of sample processing for sensitive RNA detection from cat BSLs
We first determined the optimal sampling site for RNA collection from cat BSLs. Samples were collected from the armpit, tail base, tail, and ear of the same cat. The tail base, tail, and ear all yielded significantly lower Ct values for B2M compared to the armpit (Fig. 3A). Because the subject was an intact adult male, known to secrete higher levels of BSLs^27^, we concluded that the ear represents the most reliable sampling site across sexes and age groups. In addition, by collecting BSLs and observing the color change of the oil-blotting film, which roughly reflects the amount of BSLs, we found that the color change was most pronounced and most consistent for ear-derived BSLs. All subsequent experiments were therefore conducted using ear-derived BSLs (Fig. 3B–D).
For practical application, we further evaluated whether RNA extraction could be fully automated. Samples from four cats were processed using either a manual protocol (QIAzol + Viral RNA Mini Kit) or a fully automated extraction system (All Maxwell). The automated method provided higher B2M detection sensitivity than the manual protocol (Fig. 3E). Owing to its simplicity and high sensitivity, we adopted the automated method for subsequent experiments. Finally, we tested the stability of SSL samples under different storage conditions to simulate clinical and transport scenarios. Oil-blotting films containing BSLs were stored at − 18 °C, 4 °C, 25 °C, and 37 °C for three days, reflecting estimated maximum shipping times in Japan. Samples from four cats showed no significant differences in B2M detection across the tested temperatures (Fig. 3F).
Fig. 3. Optimization of sample handling for sensitive RNA detection from cat body surface lipids (BSLs). (A) Ct values of B2M measured in BSLs from four sampling sites in the same cat. Data show the mean ± SD of quadruplicate measurements. Differences were analyzed by one-way ANOVA using armpit-derived BSLs as the control; ***p < 0.001, **p < 0.01, *p < 0.05, ns = not significant. (**B **) Representative image of sebum collection from a cat’s side. (C) Oil-blotting film before sampling. (D) Oil-blotting film after sampling, with darker areas indicating collected sebum. (E) Comparison of B2M detection sensitivity between manual and automated RNA extraction. Data show the mean ± SD of quadruplicate measurements. Differences were analyzed by one-way ANOVA using armpit-derived BSLs as the control; ***p < 0.001, **p < 0.01, *p < 0.05, and ns = not significant. (F) Effect of storage temperature on B2M detection in BSLs stored on oil-blotting films at − 18 °C, 4 °C, 25 °C, or 37 °C for three days. Data show the mean ± SD of quadruplicate measurements, representative of at least four experiments. Statistical significance was determined by one-way ANOVA; ***p < 0.001, **p < 0.01, *p < 0.05, ns = not significant.
Efficient detection of FIV virus RNA from BSLs
After optimizing the sampling site, we evaluated the feasibility of detecting B2M and FIV RNA from ear-derived BSLs. Samples were collected from 12 cats. B2M was successfully detected in all samples (Fig. 4A), confirming the ear as the optimal site for SSL collection. FIV RNA was detected only in cats previously diagnosed as FIV-positive by blood testing (Fig. 4B), demonstrating full agreement between sebum-based and conventional diagnostics.
Using values shown in Fig. 4B, we compared the performance of the sebum-based diagnostic method with the blood-based method in terms of FIV RNA sensitivity. Oil-blotting film and whole-blood samples were collected from the same six cats, and Ct values for FIV RNA were analyzed. The oil-blotting film method showed detection sensitivity equivalent to that of conventional whole-blood testing with this group of samples (Fig. 4C).
Fig. 4. Efficient detection of feline immunodeficiency virus (FIV) RNA from body surface lipids (BSLs). (A) Ct values of B2M from BSLs collected from 12 cats. Red symbols indicate FIV-positive cats; blue symbols indicate FIV-negative cats. Data show the mean ± SD of triplicate measurements. (B) Ct values of FIV RNA detected in the same BSL samples. (C) Correlation of Ct values obtained by sebum-based versus conventional blood-based testing for FIV RNA.
Detection of SFTSV RNA from Cat BSLs
As the final experiment, we investigated whether SFTSV RNA could be detected from sebum. BSLs were collected with oil-blotting films from the base of the tail of a cat naturally infected with SFTS. SFTSV RNA was successfully detected only from SSL samples collected at the base of the tail (Fig. 5).
Fig. 5. Detection of severe fever with thrombocytopenia syndrome virus (SFTSV) RNA from cat body surface lipids (BSLs). Ct values of SFTSV measured in BSLs from two cats. Red symbols represent the SFTSV-positive cat (Loid), and blue symbols represent the SFTSV-negative cat (Anya).
Discussion
In this study, we established a sebum-based diagnostic method for detecting RNA virus infections in cats. This approach aimed to minimize invasiveness for animals while enhancing safety for examiners. We developed primers that efficiently amplified viral RNA from feline sebum while distinguishing it from human DNA/RNA and feline genomic DNA. We also identified optimal sampling sites and showed that FIV RNA, a major feline pathogen, could be reliably detected from ear-derived sebum. Notably, the sensitivity of FIV detection using sebum was equivalent to that of conventional blood-based testing. Moreover, we successfully detected SFTSV RNA from sebum collected at the tail base, providing proof of concept for sebum-based diagnostics of SFTS.
The establishment of a sebum-based diagnostic method has important implications for both animal welfare and the One Health framework. By substituting needle-based blood collection with non-invasive sebum sampling, the physical and psychological burden on cats can be substantially reduced. This method aligns with the Five Freedoms of animal welfare, particularly “Freedom from Pain, Injury, or Disease” and “Freedom from Fear and Distress.” In addition, sebum-based testing could improve biosafety for cat owners, local communities, veterinarians, and veterinary nurses by lowering the risk of exposure to infectious agents during sample handling. Furthermore, because we identified B2M as a gene that can be stably detected in BSLs collected from cat ears, approaches developed in the human medical field may be extendable to cats. For example, studies of atopic dermatitis in humans have utilized mRNAs present in skin surface lipids. If such findings are translated into practical clinical applications in humans, their application to cats can be reasonably anticipated, given that a gene stably detectable from ear-derived BSLs has already been identified in cats, thereby broadening the potential use of this approach beyond human medical care.
As described above, several less-invasive diagnostic methods exist in humans, and similar approaches are available for cats. These include saliva-based testing and oral swab examinations in cats. However, the sebum-based method offers clear advantages over other less-invasive sampling strategies for several reasons. First, conjunctival and oral swab sampling often require physical restraint and are difficult for a single veterinarian to perform, particularly in uncooperative cats. In contrast, sebum collection using oil-blotting paper can be readily performed by a single veterinarian or even by cat owners themselves. This advantage is particularly important in animal shelters, where efficiency and minimal staffing are essential. Second, relative to conjunctival or oral swab sampling and urine collection, ear sebum sampling can be performed during natural human–cat interactions. In routine contact with cats, touching or gently handling the ears is common, whereas inserting swabs into the oral cavity or attempting urine collection is not. Accordingly, in cats that tolerate human contact, sebum sampling can be conducted during normal, low-stress behavior, distinguishing this approach from other less-invasive examinations. Third, oral swab sampling requires specialized consumables, which increase costs. In addition, urine collection in cats often involves invasive cystocentesis or housing cats in specialized cages to collect voided urine. Cats placed in unfamiliar environments may not urinate as expected, making timely sample collection difficult.
However, sebum collection requires no specialized equipment beyond inexpensive oil-blotting film and can be completed immediately by wiping the inner ear. Thus, the sebum-based method is cost-effective, rapid, and does not require professional consumables or waiting time. Taken together, these practical advantages support our conclusion that sebum testing represents an equivalent or superior non-invasive alternative to existing sampling methods.
Previous studies have demonstrated that sebum contains diverse biomolecules, including mRNA, as a result of holocrine secretion in humans^28^. On this basis, we hypothesized that feline sebum would also harbor mRNA. Supporting this, we successfully detected B2M-specific mRNA from feline sebum (Fig. 2), establishing a biological rationale for RNA detection from this sample type. To determine optimal sampling sites, we compared B2M Ct values across multiple body regions and found that ear-derived sebum consistently provided one of the strongest detection signals (Fig. 3A–D). Considering potential variation by sex and age, we focused subsequent experiments on ear sampling, which yielded reproducible RNA detection outcomes.
To enable practical application, we incorporated automated RNA extraction (Fig. 3E), which markedly shortened processing time without compromising sensitivity. We further evaluated sample stability across diverse storage conditions. B2M detection sensitivity remained consistent after three days at temperatures ranging from − 18 °C to 37 °C (Fig. 3F). These results support the feasibility of nationwide sample shipment, potentially expanding diagnostic access and mitigating healthcare disparities, particularly in geographically remote regions.
Regarding FIV diagnostics, B2M mRNA was identified in all samples, ensuring ear-derived sebum as a reliable source for sebum-based testing (Fig. 4A). Sebum- and blood-based diagnostics results showed complete concordance (Fig. 4B). Moreover, FIV detection with the sebum-based method achieved sensitivity equivalent to conventional blood testing (Fig. 4C), underscoring its possibility for clinical adoption. Unlike antibody-based kits, which cannot distinguish maternal antibodies from true infection^29^, this method directly targets viral RNA, enabling accurate diagnosis in kittens. Collectively, these findings indicate that sebum-based diagnostics could augment or strengthen current testing strategies.
Importantly, this work presents the first evidence that SFTSV RNA is detectable in feline sebum (Fig. 5), providing proof-of-principle for sebum-based SFTS diagnostics. Subsequent studies will compare detection sensitivity between sebum and blood samples and assess clinical utility across broader populations.
Despite encouraging outcomes, blood testing remains indispensable for a comprehensive health assessment. Hematological measures, such as platelet and white blood cell counts in SFTS^30^ and CD4 T-cell counts in FIV^31^, are critical for diagnosis. We therefore propose an integrated framework in which sebum- and blood-based tests are applied complementarily, depending on clinical objectives. Sebum testing is especially useful when venipuncture is difficult—for example, in fearful or aggressive cats, large-scale screening of community cats, or suspected zoonotic risk cases. It also provides a safer option for kittens and clinically fragile animals.
For cats prone to severe stress during veterinary visits, blood collection can worsen anxiety and deter future care [reviewed in^32^]. By contrast, sebum collection from the ear requires only gentle restraint and produces substantially less stress. Owners could also be trained to collect sebum at home, enhancing access to diagnostic testing for cats that are hard to manage in clinics.
In this study, numerous samples were collected from cats housed at the Miyazaki City shelter. Owing to its simplicity, safety, and low training threshold, sebum-based testing is particularly suited to high-throughput shelter screening. Unlike blood collection, which generally requires two trained professionals, ear-derived sebum sampling can be performed by one individual with limited instruction. Considering the global importance of FIV and FeLV screening in shelters^26^, this method offers considerable value for preventing viral spread before adoption. Ongoing validation studies combining sebum- and blood-based diagnostics are underway in collaboration with Miyazaki City shelters, aiming to establish standardized protocols.
This study has several limitations. First, sebum-based testing focuses on pathogen detection and cannot substitute for blood tests in evaluating hematological or biochemical markers. Second, in multi-cat environments, behaviors such as ear-licking may introduce saliva contamination, potentially yielding false positives, especially for viruses present in body fluids such as SFTSV^18^. This limitation may be alleviated by gathering behavioral information from owners and integrating targeted screening questions. Third, the number of samples remains limited and should be examined in a larger sample pool. Finally, the relatively complex RNA extraction workflow remains a practical barrier. To address this, we developed a fully automated extraction platform (Fig. 3E), facilitating wider clinical and epidemiological applications.
In conclusion, the sebum-based diagnostic method developed in this study offers a minimally invasive and reliable approach for detecting RNA viral infections in cats. Applying this technique in practice may also enhance examiner safety by lowering risks linked to blood handling and reducing direct contact with cats potentially carrying zoonotic pathogens. Together, these advantages emphasize the broad utility of sebum-based diagnostics for clinical practice, shelter management, and epidemiological surveillance, thereby supporting the One Health framework. Future work will focus on refining sampling protocols, validating diagnostic performance across diverse populations, and establishing standardized guidelines for veterinary and public health professionals. Collectively, these efforts aim to strengthen feline viral infection control, improve animal welfare, and promote wider adoption of minimally invasive and safe diagnostic tools for RNA virus detection.
Methods
Ethical approval
All procedures involving the collection of feline samples were approved by the Animal Care and Use Committee of the University of Miyazaki (approval No. 2025-024) and conducted in accordance with the University of Miyazaki Experimentation regulations and relevant institutional guidelines. We have obtained written consent from the animal shelter for the use of cats in this study.
ARRIVE statement (non-invasive animal study)
This study is reported in accordance with the ARRIVE guidelines. All procedures involving animals were non-invasive and conducted in a manner that minimized animal distress.
BSL sample collection
BSLs were collected from animal hospitals and the Miyazaki Animal Protection Center between March and August 2025 using STF Oil Clear Film (Hakugen Earth Co., Ltd., Tokyo, Japan; 75 mm × 160 mm × 3 mm; JAN code: 4902407040930). The total sample number was 81, and sampling was performed by gently wiping designated areas, without anesthesia or physical restraint. Each used film was sealed individually in a plastic bag and stored at − 30 °C until processing.
Clinical stages of FIV-infected cats
As all BSL samples from FIV-infected cats were obtained from an animal shelter and most cats had been rescued from the streets, the exact timing or duration of FIV infection could not be determined. However, all cats included in this study were asymptomatic carriers, exhibiting no clinical signs of immunosuppression and appearing clinically healthy at the time of sampling.
Cell culture
Lenti-X 293T cells (Homo sapiens; TaKaRa Bio Inc., Kusatsu, Japan; Cat# Z2180N), HeLa cells (Homo sapiens; ATCC, Manassas, VA, USA; Cat# CCL-2), Fcwf-4 cells (Felis catus; ATCC; Cat# CRL-2787), and Crandell-Rees feline kidney cells (Felis catus; JCRB, Ibaraki, Japan; Cat# JCRB9035) were maintained in Dulbecco’s modified Eagle’s medium (Nacalai Tesque, Kyoto, Japan; Cat# 08458-16) supplemented with 10% fetal bovine serum (FBS) and 1× penicillin–streptomycin (Nacalai Tesque; Cat# 09367-34). MT4 cells (Homo sapiens; JCRB; Cat# JCRB0135) were maintained in Roswell Park Memorial Institute medium (RPMI-1640; Nacalai Tesque; Cat# 30264-56) with 10% FBS and 1× penicillin–streptomycin (Nacalai Tesque; Cat# 09367-34).
Extraction of RNA from Cat BSLs
Sebum samples collected on oil-blotting films were cut into small pieces and placed in 2.0 mL microcentrifuge tubes. QIAzol reagent (1,450 µL; QIAGEN, Hilden, Germany; Cat# 79306) was added, and tubes with samples were incubated in a constant-temperature shaker at 200 rpm for 5 min at room temperature. After brief vortexing, 1,300 µL of the supernatant was transferred to fresh tubes. Chloroform (260 µL; Nacalai Tesque, Kyoto, Japan; Cat# 08402-84) was added, followed by vortexing and centrifugation at 15,000 × g for 15 min at 4 °C. The aqueous phase (600 µL) was carefully collected, and RNA was purified with the RNeasy Mini QIAcube Kit (QIAGEN; Cat# 74116), including on-column DNase treatment under a custom protocol. In brief, 600 µL of 85% ethanol was added, mixed thoroughly, and loaded onto RNeasy Mini spin columns. The columns were washed with RW1 buffer, treated with DNase I, washed again with RW1, and finally washed with RPE buffer. RNA was eluted twice with 50 µL RNase-free water, yielding a final volume of 100 µL.
RNA cleanup was performed using the Monarch^®^ Spin RNA Cleanup Kit (10 µg; New England Biolabs, Ipswich, MA, USA; Cat# T2030S). In brief, 200 µL Buffer BX and 300 µL ethanol were added to 100 µL RNA solution and mixed by pipetting. The mixture was loaded onto the spin column and centrifuged at maximum speed for 1 min. The column was then washed twice with 500 µL Buffer WX, each step followed by centrifugation for 1 min. After a final dry spin to remove residual buffer, the column was placed onto a new RNase-free 1.5 mL microcentrifuge tube, and RNA was eluted with 8 µL RNase-free water.
For automated RNA extraction, RNA from cat BSLs was processed using the Maxwell^®^ RSC 48 Instrument (Promega, Madison, WI, USA; Cat# AS8500) in combination with the Maxwell^®^ RSC miRNA Plasma and Serum Kit (Promega; Cat# AS1680), according to the manufacturer’s instructions.
Primers
All primers were designed with reference to earlier studies^33^ and the QIAGEN database of housekeeping genes^34^. Sequences were generated using Primer-BLAST (https://www.ncbi.nlm.nih.gov/tools/primer-blast/; accessed May 5, 2025) based on the Felis catus genomic sequence (accession no. JAFEKA010000007.1). The following criteria were applied for primer design: (1) expected PCR product size was set between 70 and 150 bp, (2) primers separated by at least 1,000 bp of intronic sequence, and (3) primers were required to span an exon–exon junction (Supplementary Table 2). All primers were synthesized as standard desalted DNA oligonucleotides by Eurofins Genomics (Tokyo, Japan) and stored at 4 °C until use.
Real-time quantitative polymerase chain reaction (RT-qPCR)
Total DNA was extracted from cultured cells using the DNeasy Blood & Tissue Kit (QIAGEN, Hilden, Germany; Cat# 69506) with RNase A (17,500 U; QIAGEN, Cat# 19101) and 100% ethanol, following the manufacturer’s protocol. Total RNA was extracted from cultured cells using the RNeasy Mini Kit (QIAGEN; Cat# 74104) with QIAshredder columns (QIAGEN; Cat# 79656), the RNase-Free DNase Set (QIAGEN; Cat# 79254), and 2-mercaptoethanol (Bio-Rad, Hercules, CA, USA; Cat# 1610710). RNA from sebum samples on oil-blotting films was purified with the Monarch^®^ Spin RNA Cleanup Kit (10 µg; New England Biolabs; Cat# T2030S) with 85%–100% ethanol. DNA and RNA concentrations were measured using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA).
The mRNA expression of six housekeeping genes—Actin Beta (Actb), Cytochrome C1 (Cyc1), glyceraldehyde-3-phosphate dehydrogenase (Gapdh), peptidylprolyl isomerase A (PPIA), succinate dehydrogenase complex flavoprotein subunit A (Sdha), and Beta-2-microglobulin (B2M)—was quantified using the One Step TB Green^®^ PrimeScript™ PLUS RT-PCR Kit (Perfect Real Time) (TaKaRa Bio, Shiga, Japan; Cat# RR096A). RT-qPCR was carried out under the following cycling conditions: reverse transcription at 42 °C for 5 min, initial denaturation at 95 °C for 10 s, followed by 40 cycles of 95 °C for 5 s and 60 °C for 34 s.
To quantify FIV RNA (Supplementary Table 3), SFTSV RNA (Supplementary Table 4), and B2M mRNA, RT-qPCR was performed using the PrimeDirect^®^ Probe RT-qPCR Mix (TaKaRa; Cat# RR650A). Conditions were reverse transcription at 90 °C for 3 min, annealing at 60 °C for 5 min, and 40 cycles of 95 °C for 5 s and 60 °C for 30 s. For all reactions, melting-curve analysis was conducted to verify amplification specificity, and relative expression levels were determined using the 2^^ΔCt^ method.
Statistical analysis
The differences in Ct values across treatments were assessed using one-way analysis of variance (ANOVA), followed by Tukey’s post hoc test, or by the Kruskal–Wallis test with Dunn’s post hoc test where appropriate. A p-value ≤ 0.05 was considered statistically significant. All analyses were conducted in GraphPad Prism (ver. 10.2.3; GraphPad Software, San Diego, CA, USA).
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary Material 1
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Fehlner-Gardiner, C. et al. Rabies in cats-An emerging public health issue. Viruses 16(10) (2024).10.3390/v 16101635 PMC 1151239539459967 · doi ↗ · pubmed ↗
- 2Seo, J. W. et al. Clinical update of severe fever with thrombocytopenia syndrome. Viruses 13(7) (2021).10.3390/v 13071213 PMC 831001834201811 · doi ↗ · pubmed ↗
- 3Ando, T. et al. Severe fever with thrombocytopenia syndrome in cats and its prevalence among veterinarian staff members in Nagasaki, Japan. Viruses 13(6) (2021).10.3390/v 13061142 PMC 823225734198717 · doi ↗ · pubmed ↗
- 4三重県内におけるネコからヒト(獣医師 )への感染が疑われた重症熱性血小板減少症候群༈SFTS༉の死亡例. Japan Institute for Health Security: Infectious Agents Surveillance Report. 165-167 (2025).
- 5Murphy, B. G. et al. The late asymptomatic and terminal immunodeficiency phases in experimentally FIV-infected cats-A long-term study. Viruses 15(8) (2023).10.3390/v 15081775 PMC 1045790637632117 · doi ↗ · pubmed ↗
- 6Kristin Mac Donald, M. et al. Effects of Passive Transfer of Immunity on Results of Diagnostic Tests for Antibodies Against Feline Immunodeficiency Virus in Kittens Born to Vaccinated Queens. 1554–1557 (JAVMA, 2004).10.2460/javma.2004.225.155415568387 · doi ↗ · pubmed ↗
- 7Osako, H. et al. Clinical factors associated with SFTS diagnosis and severity in cats. Viruses 16(6) (2024).10.3390/v 16060874 PMC 1120930538932167 · doi ↗ · pubmed ↗
- 8Riemer, S. et al. A review on mitigating fear and aggression in dogs and cats in a veterinary setting. Animal (Basel)11(1) (2021).10.3390/ani 11010158 PMC 782656633445559 · doi ↗ · pubmed ↗