KIR AA individuals possess strong inhibitory KIR alleles alongside HLA ligands that are protective against leukemia in the Chinese population
Zhihui Deng, Jianxin Zhen, Yunan Li, Shuang Liang, Manru Zhang, Siqi Cai, Renhui Jiang, Zhichao Yang, Qiong Yu, Jinyong Wang, Jie Liu

TL;DR
This study finds that specific KIR and HLA gene combinations protect against leukemia in the Chinese population.
Contribution
High-resolution analysis reveals protective KIR-HLA interactions in KIR AA individuals against leukemia.
Findings
KIR2DL1*00201+C2 interaction reduces ALL risk (p = 0.01).
KIR3DL1*00501+Bw4 80I protects against AML (p = 0.008).
KIR3DL3*001 is protective against AML, while KIR3DL3*009 increases AML risk.
Abstract
The killer-cell immunoglobulin-like receptor (KIR) gene cluster exhibits complicated diversity in haplotype content, copy-number variation (CNV), and allelic polymorphism. To date, 2,286 distinct KIR alleles have been released in the IPD-KIR Database. However, little is known about the impact of high-resolution-level KIR allelic polymorphisms on leukemia. Our previous study showed that the KIR AA genotype carrying more inhibitory genes conferred differential protection against leukemia in the Chinese Southern Han population. Herein, we hypothesized the impact of KIR alleles in the KIR A haplotype and cognate human leukocyte antigen (HLA) ligand on leukemia. The study cohort included 318 ALL patients, 336 AML patients, and 306 unrelated healthy controls. All the study samples were subject to HLA-A, -B, and -C sequencing-based genotyping (PCR-SBT) and high-resolution KIR genotyping for…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
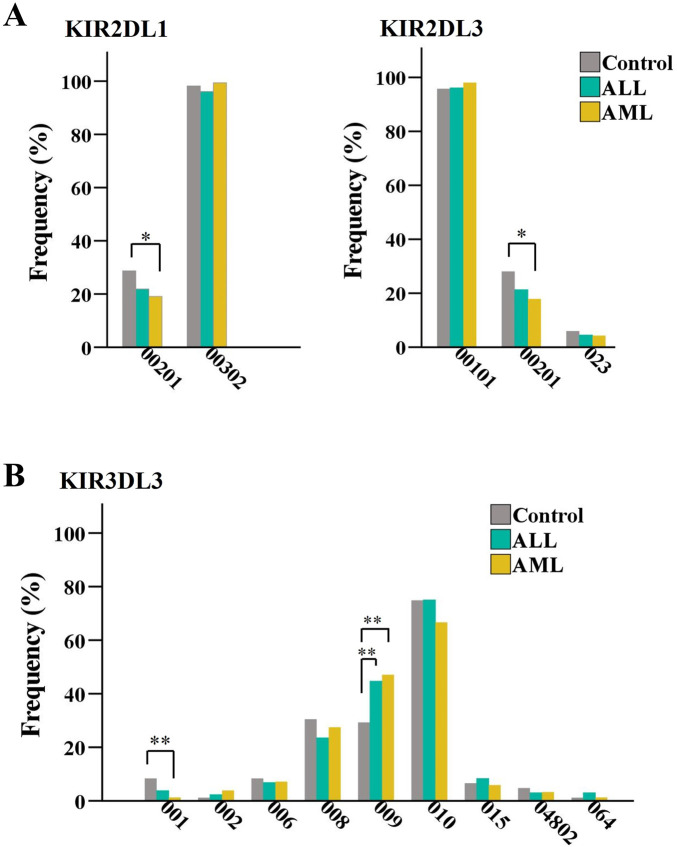 FIGURE 1
FIGURE 1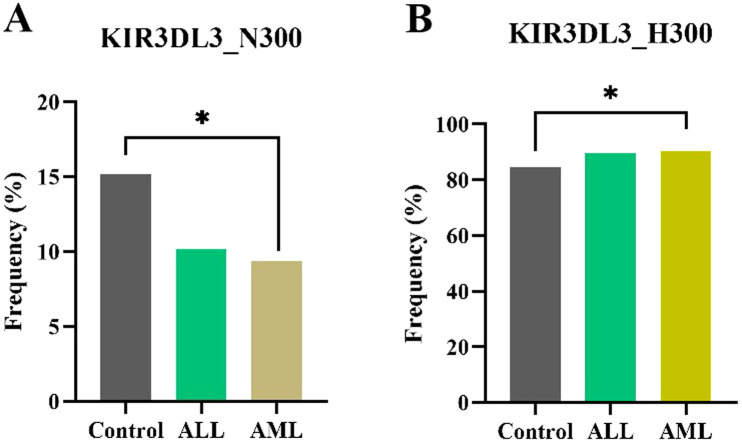 FIGURE 2
FIGURE 2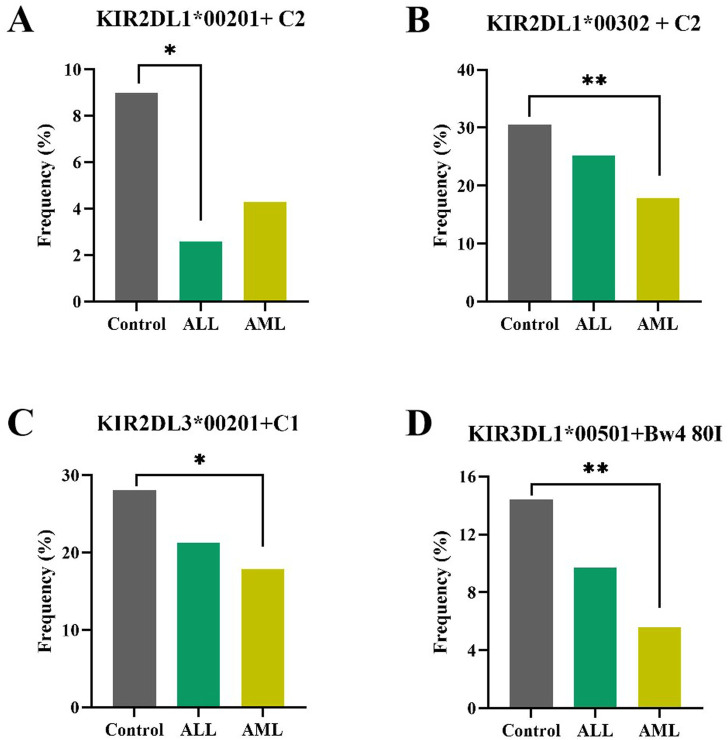 FIGURE 3
FIGURE 3| KIR gene | Healthy controls (N = 306) | ALL patients (N = 318) | AML patients (N = 336) | |||
|---|---|---|---|---|---|---|
| Number of KIR alleles identified | Number of KIR alleles with a frequency of ≥1% | Number of KIR alleles identified | Number of KIR alleles with a frequency of ≥1% | Number of KIR alleles identified | Number of KIR alleles with a frequency of ≥1% | |
|
| 10 | 3 | 14 | 5 | 11 | 4 |
|
| 15 | 4 | 14 | 5 | 15 | 4 |
|
| 8 | 5 | 6 | 5 | 8 | 5 |
|
| 7 | 4 | 5 | 4 | 4 | 4 |
|
| 11 | 6 | 11 | 6 | 12 | 8 |
|
| 18 | 12 | 16 | 13 | 21 | 12 |
|
| 19 | 15 | 17 | 13 | 18 | 13 |
| Total | 88 | 49 | 83 | 51 | 89 | 50 |
| KIR allele | Control (N = 167) | ALL patients (N = 155) | AML patients (N = 162) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| n (%) | n (%) | OR (95% CI) |
|
| n (%) | OR (95% CI) |
|
| |
|
| 48 (28.7) | 34 (21.9) | 0.70 (0.42–1.16) | 0.16 | | 31 (19.1) | 0.59 (0.35–0.98) |
| 0.41 |
|
| 0 (0) | 5(3.2) | 0.97 (0.94–1.00) | 0.06 | | 6 (3.7) | 0.96 (0.93–0.99) |
| 0.36 |
|
| 47 (28.1) | 33(21.3) | 0.69 (0.41–1.15) | 0.16 | | 29 (17.9) | 0.56 (0.33–0.94) |
| 0.50 |
|
| 14 (8.4) | 5(3.8) | 0.43 (0.15–1.23) | 0.11 | | 2 (1.3) | 0.14 (0.03–0.65) |
| 0.06 |
|
| 49 (29.3) | 59(44.7) | 1.95 (1.21–3.14) |
| 0.08 | 72 (47.1) | 2.14 (1.35–3.39) |
|
|
|
|
| Control | ALL | AML | ||||
|---|---|---|---|---|---|---|---|---|
| n (%) | n (%) | OR (95% CI) |
| n (%) | OR (95% CI) |
| ||
|
|
| 15 (9.0) | 4 (2.6) | 0.27 (0.09-0.83) |
| 7 (4.3) | 0.46 (0.18-1.15) | 0.09 |
| |
| 51 (30.5) | 39 (25.2) | 0.76 (0.47-1.25) | 0.28 | 29 (17.9) | 0.50 (0.30-0.83) |
|
| |
| 47 (28.1) | 33 (21.3) | 0.69 (0.41-1.15) | 0.16 | 29 (17.9) | 0.56 (0.33-0.94) |
|
| |
| 24 (14.4) | 15 (9.7) | 0.64 (0.32-1.27) | 0.2 | 9 (5.6) | 0.35 (0.16-0.78) |
|
|
|
| 0 (0) | 9 (5.5) | 0.94 (0.91-0.98) |
| 6 (3.4) | 0.97 (0.94-0.99) | 0.07 |
- —National Natural Science Foundation of China10.13039/501100001809
- —Science, Technology and Innovation Commission of Shenzhen Municipality10.13039/501100010877
- —Sanming Project of Medicine in Shenzen Municipality10.13039/501100012151
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsImmune Cell Function and Interaction · CAR-T cell therapy research · Lymphoma Diagnosis and Treatment
Introduction
1
Killer-cell immunoglobulin-like receptors (KIRs) are expressed on the surface of natural killer (NK) cells and a subset of cytotoxic T cells. They regulate effector cell activity through interactions with class I human leukocyte antigen (HLA) ligands on target cells, which play a major role in immune surveillance and elimination of tumor and virus-infected cells (Zuo and Zhao, 2021; Deng et al., 2019; Liu et al., 2021; Putz et al., 2024). To perform this function, NK cells express multiple inhibitory and activating receptors; polymorphic inhibitory receptors educate and license NK cells, enabling them to kill tumor or infected cells exhibiting altered or reduced HLA class I expression (Deng et al., 2019).
The function of KIR on NK cells is dependent on the normal expression of class I HLA ligands on the target cell. Specifically, HLA-C alleles form two ligand groups, C1 and C2, based on key amino acids at positions 77 and 80. These groups interact with different inhibitory KIRs: HLA-C1 binds to KIR2DL2/3, while HLA-C2 engages the KIR2DL1 receptor. HLA-B alleles are divided into Bw4 and Bw6, but only Bw4 allotypes serve as ligands for KIR3DL1, with binding strength influenced by residue 80 (Ile80 > Thr80). The Bw4 motif also appears in some HLA-A alleles (e.g., A23/24/32*), and other HLA-A allotypes such as A03/11* can interact with KIR3DL2.
The KIR gene cluster on chromosome 19q13.4 consists of 14 functional genes and 2 pseudogenes, exhibiting complicated diversity in haplotype content, copy-number variation (CNV), and allelic polymorphism. Based on individual gene content, KIR genes are classified into two broad haplotypes termed A and B. The KIR A haplotype is defined by a fixed set of nine genes with only KIR2DS4 as activating genes, whereas KIR B haplotypes are more diverse and characterized by the presence of more than one activating KIR gene and the absence of KIR2DS4. Four framework genes (KIR3DL3, KIR3DP1, KIR2DL4, and KIR3DL2) are present on virtually all haplotypes, and every KIR haplotype is a combination of a centromeric and a telomeric KIR gene motif. Individuals can be grouped into KIR AA or KIR Bx (including KIR BB and KIR AB) genotype. To date, 2,286 KIR alleles have been recorded in the IPD-KIR Database (Release 2.15) (Robinson et al., 2013).
KIR allelic polymorphism is known to dramatically alter the characteristics of the transcribed glycoprotein, influencing the binding affinity, specificity, surface expression, and signaling capacity of the expressed receptor (Bari et al., 2009; Saunders et al., 2016; Nemat-Gorgani et al., 2018; Le Luduec et al., 2019; Wright et al., 2024). The significance of KIR allele polymorphisms in clinical transplantation, diseases association, population genetics, and evolution studies has drawn extensive interest recently (Bari et al., 2013; Boudreau et al., 2017; Deng et al., 2021; Guethlein et al., 2021; Palmer et al., 2023).
The existing literature exploring the influence of KIR allelic polymorphisms on recipient outcomes after allogeneic hematopoietic stem cell transplantation (HSCT) mainly pointed to donor centromeric (cen) AA KIR2DL (2DL1–3) (Dubreuil et al., 2020), KIR3DL1 (Boudreau et al., 2017), and KIR2DL1 allelic/allotype polymorphisms (Bari et al., 2013; Wright et al., 2024). Centromeric (cen) AA individuals carrying more efficient KIR2DL alleles (KIR2DL1003* and KIR2DL3001*) were associated with better responsiveness and reduced relapse rate in patients with myeloid diseases after T-cell-replete haplo-identical HSCT (Dubreuil et al., 2020). Additionally, KIR3DL1 and HLA-B combinations that were predictive of weak inhibition or noninhibition were associated with significantly lower relapse and overall mortality following allogeneic HSCT in AML patients (Boudreau et al., 2017). Furthermore, KIR2DL1 alleles with arginine at amino acid position 245 (KIR2DL1-R ^ 245 ^) are functionally stronger than KIR2DL1 alleles with cysteine at the same position (KIR2DL1-C ^ 245 ^) (Bari et al., 2009). A donor graft containing KIR2DL1-R ^ 245 ^ (KIR2DL1002/001g*, 003, etc.) was associated with better survival and lower cumulative incidence of disease progression in pediatric allogeneic HSCT (Bari et al., 2013). However, the effect of donor KIR2DL1 allelic diversity on the outcomes of HSCT can be influenced by the conditioning regimen. In a more recent study, donor KIR2DL1003 with KIR2DL1-R ^ 245 ^ severely reduced disease-free survival and increased 5-year relapse incidence for AML patients receiving reduced intensity conditioning (RIC) and T-cell-depleted (TcD) transplantation, suggesting that KIR2DL1003-*positive donors should be excluded for TcD and RIC transplantation (Wright et al., 2024).
Taken together, the above studies demonstrated that the KIR alleles and functional allotypes are apparently associated with the license of NK cells, cancer control, and patient survival after HSCT. However, evaluating the influence of distinct KIR allele and HLA ligands on transplantation outcomes is complex as conditioning regimens (conditioning intensity, T-cell-depleted, etc.) may influence the effect of donor KIR allelic diversity on HSCT outcomes (Wright et al., 2024). In contrast, the present study focuses on the genetic factors of KIR–HLA interactions mediating protection against or susceptibility to leukemia, without considering the influences of conditioning regimens in transplantation.
Studying KIR polymorphism is critical for understanding its anti-leukemic effects and facilitating subsequent application in optimized donor selection in HSCT and immunotherapy (Deng et al., 2019). Based on the low-resolution level diversity of KIR genes, numerous studies analyzing potential KIR/HLA associations with leukemia have yielded inconsistent results (Babor et al., 2014; de Smith et al., 2014; Vejbaesya et al., 2014; Augusto, 2016; Misra et al., 2016; Shen et al., 2016; Li et al., 2023). The disparate results might be partly due to genetic variation across populations. For instance, de Smith et al. (2014) demonstrated that haplotype A was associated with an increased risk of ALL in Hispanics but not in non-Hispanic whites, while homozygosity for HLA-Bw4 was associated with an increased risk in non-Hispanic whites but not in Hispanics, which demonstrate that the examination of specific well-defined human populations is critical for understanding the role of KIR in leukemia control.
AML is a hematological malignancy originating in the bone marrow characterized by a disruption in normal hematopoietic differentiation (Estey and Dohner, 2006), which accounts for approximately 1.3% of all cancer cases and is responsible for approximately 62% of deaths caused by leukemia (Paparo et al., 2025). ALL is a hematological malignancy characterized by the uncontrolled proliferation of immature lymphoid cells, which is an leading cause of cancer-related death in adults (Siegel et al., 2023; Zhang et al., 2023). China had the highest burden of ALL and AML worldwide, accounting for 38,570.94 incidences and 20,612.91 deaths in 2021 (Ni et al., 2025). Our previous study revealed that the frequency of individuals with the KIR AA genotype among healthy Chinese Hans is significantly greater than that in other populations representing major world groups (including Amerindian, European, Oceanian, and sub-Saharan African). Moreover, the frequency of KIR AA among healthy Chinese Southern Hans (N = 745) is significantly higher than that in adult leukemic patients (N = 836) and pediatric leukemic patients (N = 225), which implies that the KIR AA genotype carrying more inhibitory KIR genes protects the Chinese Han individuals from leukemia and plays a critical role in human adaptive evolution (Deng et al., 2019). These genetically determined distinctions prompted us to explore the association of allelic polymorphisms in all the seven functional KIR genes on the KIR A haplotype with acute leukemia occurrence in the Chinese population, aiming at identifying the risk or protective KIR alleles or KIR–HLA combinations and elucidating potential mechanisms. These findings may provide valuable insights into the pathogenesis of leukemia and facilitate potential application in donor selection for transplantation and NK-cell immunotherapy.
Materials and methods
2
Samples from patients and healthy donors
2.1
The study cohort included 318 ALL patients, 336 AML patients, and 306 age-matched, unrelated, healthy blood donors with the average age of 31.2, 34.3, and 32.7 years, respectively, as detailed in Supplementary Table S1. The studied adult ALL/AML patients who underwent chemotherapy treatments were recruited from the HSCT program at Shenzhen Blood Center from August 1999 through June 2015. The presence or absence of KIR genes had been identified for these patients in our previous study (Deng et al., 2019), but high-resolution KIR genotyping was not performed. Individuals with negative results for hepatitis B virus (HBV), hepatitis C virus (HCV), human immunodeficiency virus (HIV), and syphilis were enrolled as healthy controls. Healthy control samples were randomly collected at Shenzhen Blood Center during the period of June 2013 through May 2014. The high-resolution genotypes of the 14 functional KIRs had been elucidated for healthy controls, as described previously (Deng et al., 2021). Particularly, the healthy control individuals had no family relationships with the leukemic patients. Written informed consent was obtained from all subjects, and the study was approved by the Ethics Review Board of the Shenzhen Blood Center, Shenzhen, Guangdong, China.
High-resolution HLA-A, -B, and -C genotyping
2.2
HLA-A, -B, and -C genotyping was performed using the AlleleSEQR HLA sequencing-based genotyping (PCR-SBT) commercial kit (Atria Genetics, San Francisco, United States). According to the manufacturer’s instructions, exons 2–4 for HLA-A, -B, and -C were sequenced in both directions using an ABI 3730XL DNA sequencer (Applied Biosystems, Foster City, CA, United States). HLA genotypes with four-digit resolution were assigned using Assign 4.7.1 software (Conexio Genomics, Fremantle, Australia).
High-resolution-level KIR genotyping for AML or ALL patients
2.3
We performed high-resolution KIR genotyping for all the seven functional KIR genes (KIR2DL1, KIR2DL3, KIR2DL4, KIR3DL1, KIR3DL2, KIR3DL3, and KIR2DS4) on the KIR A haplotype. KIR genes that tested positive using PCR-SSP were then subjected to sequencing-based genotyping of all exons for each functional KIR gene, as described previously (Deng et al., 2021). KIR alleles with at least three-digit resolution were assigned using Assign 4.7.1 software (Conexio Genomics, Fremantle, Australia) with reference to the KIR alleles Release 2.13 of the IPD (Robinson et al., 2013). When the sequencing results showed ambiguous allele combinations, we further used group-specific PCR primer pairs to amplify and sequence the target KIR alleles separately (Zhang and Deng, 2016). Novel KIR alleles were confirmed via TA cloning and sequencing of KIR transcripts, as described previously (Deng et al., 2021; Yu et al., 2024).
Statistical analysis
2.4
The observed frequencies of KIR alleles were calculated by directly counting the number of allele-positive individuals and then dividing it by the total number of studied samples, as described in previous studies (Chen et al., 2022). To compare the frequencies of the identified KIR alleles, a chi-squared test was applied using IBM SPSS Statistics 20 software, with a p-value of less than 0.05 indicating statistical significance. Where appropriate, all analyses were corrected using the Bonferroni method (Deng et al., 2019; Chen et al., 2022).
Results
3
Characteristics of KIR alleles identified in patients and healthy controls
3.1
We analyzed all the seven functional KIR genes on the KIR A haplotype, and a total of 88 distinct KIR alleles for healthy controls, 83 for ALL patients, and 89 for AML patients were identified, as summarized in Table 1. Additionally, 28 novel KIR alleles were identified and characterized, as summarized in Supplementary Table S2. Ten of these novel alleles having an observed count of ≥3 were identified in leukemic patients.
The strong inhibitory alleles KIR2DL1003* and KIR2DL3001* were common with observed frequencies exceeding 89.3% in healthy controls and ALL/AML patients. KIR3DL1015*, a strong inhibitor upon binding to the Bw4 ligand, is also frequent in the studied subjects. However, the weakly expressed allele KIR3DL1004*, which is present at allele frequencies of 5%–15% in African, European, Oceanic, and South Asian populations (Norman et al., 2007; Deng et al., 2021), is absent in the Chinese Han population. These findings suggest that the Chinese Han population has a strong predisposition to retain a high number of functional inhibitory KIR alleles.
In both ALL and AML patients, the telomeric KIR3DL1 and KIR3DL2 and centromeric KIR3DL3 genes, which encode inhibitory NK-cell receptors, each possess two or three high-frequency alleles alongside multiple less frequent alleles. In contrast, the centromeric KIR2DL1 and KIR2DL3 genes, which also encode inhibitory receptors, are all dominated by a single high-frequency allele (Supplementary Figure S1 for ALL; Supplementary Figure S2 for AML). This observation agreed well with our previous population study on healthy Chinese Hans (Deng et al., 2021), which indicated that directional selection reduced the sequence diversity of the centromeric KIR in the Chinese Han population, whereas the telomeric KIR region maintained high diversity caused by balancing selection.
KIR2DL1 and KIR2DL3 harbor alleles that tend to confer protection against ALL or AML, while KIR3DL3 allelic polymorphisms associate with leukemia occurrence
3.2
To analyze the role of KIR alleles in conferring susceptibility or protection against leukemia, healthy controls and leukemic patients were categorized into KIR AA and KIR Bx groups and then analyzed independently. Comparison of the frequencies of KIR alleles between healthy controls and patients with the KIR AA genotype (Supplementary Table S3) showed that two centromeric strong inhibitory alleles KIR2DL100201* and KIR2DL300201* were associated with protection against AML (p < 0.05), as shown in Figure 1A. The leukemia-specific novel allele KIR2DL1069*, which was most similar to KIR2DL100302* with two amino acid substitutions (308Ile > Thr and 309Ile > Arg) in cytoplasmic regions, showed susceptibility to AML (p = 0.04). However, the significant differences in these alleles were lost after *p-*value correction (Table 2).
*Comparison analysis of the allele frequencies of KIR2DL1, KIR2DL3, (A) and KIR3DL3 (B) genes between healthy controls and ALL or AML patients with the KIR AA genotype. *p < 0.05 and *p < 0.01.
For the first time, we found that the allelic polymorphism of the structure gene KIR3DL3 was associated with leukemia occurrence (Table 2; Figure 1B). Compared to healthy controls, KIR3DL3001* showed a decreased frequency in AML groups (8.4% vs. 1.3%, p = 0.004), suggesting that KIR3DL3001* tends to confer protection against AML, though the difference did not reach statistical significance (Pc = 0.06). In contrast, the frequency of KIR3DL3009* significantly increased in the AML (29.3% vs. 47.1%, p = 0.001) and ALL (29.3% vs. 44.7%, p = 0.01) groups, indicating that KIR3DL3009* conferred susceptibility to AML/ALL (AML: Pc = 0.016; ALL: Pc = 0.08). KIR3DL3001* differs from KIR3DL3009* by a single missense mutation at Codon 300 AAC>CAC in exon 7, which results in an amino acid substitution of non-charged asparagine (N) to charged histidine (H) in its transmembrane domain. Our results indicate that this functional variant site KIR3DL3_N300H plays a critical role in the occurrence of leukemia in the Chinese population.
As for the healthy controls and leukemic patients with the KIR Bx genotype, no KIR alleles showed significant difference (Supplementary Table S4).
KIR3DL3_N300 and KIR3DL3_H300 showed an association with the occurrence of AML in the Chinese population
3.3
Based on the residue in position 300, KIR3DL3 alleles were grouped as KIR3DL3_N300 (KIR3DL3001*, 006, etc.), KIR3DL3_H300 (KIR3DL3002, *008, 009, etc.), and KIR3DL3_Y300 (KIR3DL3003, *004, etc.). Compared to healthy controls, KIR3DL3_N300 showed a decreased frequency in AML (15.2% vs. 9.4%, p = 0.03), whereas KIR3DL3_H300 showed an increased frequency in AML (84.7% vs. 90.5%, p = 0.03), as shown in Figures 2A,B and Supplementary Table S5. Thus, the functional allotype analysis of KIR3DL3_300 agreed well with the data for the association of KIR3DL3 allelic polymorphisms with AML or ALL. Notably, KIR3DL3 alleles with KIR3DL3_Y300, which is common in Caucasians, African Americans, and Māori populations with a frequency of 33.4%, 33.5%, and 32.7%, respectively (Hou et al., 2011; Vierra-Green et al., 2012; Nemat-Gorgani et al., 2014), is absent in Chinese Hans.
*Association of the KIR3DL3_300 allotypes with the occurrence of leukemia. Compared to healthy controls, (A) KIR3DL3_N300 showed a decreased frequency in AML (15.2% vs. 9.4%, *p = 0.03) and (B) KIR3DL3_H300 showed an increased frequency in AML (84.7% vs. 90.5%, p = 0.03).
Other allotypes including KIR2DL1-P114/KIR2DL1-L114 (Hilton et al., 2015) and KIR2DL1-R245/KIR2DL1-C245 (Bari et al., 2011; Bari et al., 2013; Hilton et al., 2015) showed no significant difference in the frequencies of these allotypes between healthy controls and ALL/AML patients with the KIR AA genotype. Neither the KIR3DL1-H nor the KIR3DL1-L allotype (Gardiner et al., 2001; Boudreau et al., 2014) was associated with the risk of ALL/AML (Supplementary Table S5 ).
HLA-C2 ligand was associated with protection against AML in individuals with the KIR AA genotype
3.4
To evaluate the role of HLA ligands in conferring susceptibility or protective effects, we compared the observed frequencies of HLA ligands between the healthy controls and ALL/AML patients with the KIR AA genotype; the strong inhibitory HLA-C2 ligand was associated with a protective effect against AML (31.1% vs. 17.9%, P = 0.005), and the difference remained statistically significant after *p-*value correction using the Bonferroni method (Pc = 0.01), as shown in Supplementary Table S6.
For the healthy controls and leukemic patients with the KIR Bx genotype, no HLA ligands showed significant difference.
KIR AA individuals possessing strong inhibitory KIR alleles in the presence of cognate ligands show protective effects against ALL or AML, while KIR Bx individuals possessing the functionally weaker KIR2DL1*004+HLA-C2 interaction confer susceptibility to ALL
3.5
In individuals with the KIR AA genotype, specific KIR–HLA interactions were found to be significantly associated with protective effects against leukemia (Table 3; Supplementary Table S7). As shown in Figure 3A, the KIR2DL100201*+C2 interaction showed a significantly decreased frequency in ALL patients compared to healthy controls (2.6% vs. 9.0%, p = 0.01). Similarly, KIR2DL100302*+C2, KIR2DL300201*+C1, and KIR3DL100501*+Bw4 80I interactions all showed decreased frequencies in AML patients compared to healthy controls (2DL100302*+C2: 17.9% vs. 30.5%, p = 0.008; 2DL300201+C1*: 17.9% vs. 28.1%, p = 0.03; 3DL100501*+Bw4 80I: 5.6% vs. 14.4%, p = 0.008), as shown in Figures 3B–D. These results suggested that individuals with the KIR AA genotype possessing strong inhibitory KIR alleles (2DL100201*, 2DL100302, 2DL300201, and 3DL100501*) in the presence of cognate ligands conferred protection against ALL or AML in Chinese Hans.
Specific KIR–HLA interactions were significantly associated with protective effects against leukemia in individuals with the KIR AA genotype. (A) The KIR2DL100201+C2 interaction showed a significantly decreased frequency in ALL patients compared to healthy controls (2.6% vs. 9.0%, p = 0.01). (B) The KIR2DL100302+C2 interaction showed decreased frequencies in AML patients compared to healthy controls (17.9% vs. 30.5%, **p = 0.008). (C) The KIR2DL3*00201+C1 interaction showed decreased frequencies in AML patients compared to healthy controls (17.9% vs. 28.1%, p = 0.03). (D) The KIR3DL100501+Bw4 80I interaction showed decreased frequencies in AML patients compared to healthy controls (5.6% vs. 14.4%, **p = 0.008). *p < 0.05 and *p < 0.01.
In contrast, analysis of the healthy controls and leukemic patients with the KIR Bx genotype showed that the functionally weaker KIR2DL1004+HLA-C2* interaction conferred susceptibility to ALL (p = 0.01, Table 3). No other KIR–HLA interaction was found to be associated with the occurrence of ALL or AML (Supplementary Table S8).
Discussion
4
Our previous study revealed that the KIR AA genotype carrying more inhibitory KIR genes confers protective effects against leukemia in the Chinese Han population (Deng et al., 2019). This observation prompted us to explore the association of allelic polymorphisms in all the seven functional KIR genes on the KIR A haplotype with acute leukemia. As the allelic diversity of the KIR B-haplotype-specific genes (2DL2, 2DL5, 2DS1, 2DS2, 2DS3, 2DS5, and 3DS1) is limited (Deng et al., 2021) and the frequencies of the KIR B-haplotype-specific genes in ALL/AML patients showed no statistically significant difference in the healthy controls (Deng et al., 2019), we did not analyze the association of KIR B-haplotype-specific alleles with the occurrence of AML/ALL.
In the present study, we found that individuals with the KIR AA genotype possessed strong inhibitory KIR alleles for 2DL1, 2DL3, and 3DL1 in association with cognate ligands that are protective against leukemia in the Chinese population. High-resolution genetic analysis indicated that the strong inhibitory interaction KIR2DL100201*+C2 conferred protective effects against ALL, while KIR2DL100302*+C2, KIR2DL300201*+C1, and KIR3DL100501*+Bw4 80I interactions exerted protective effects against AML. However, the functionally weaker KIR2DL1004+HLA-C2* interaction was associated with ALL risk (p = 0.01) among individuals with the KIR Bx genotype.
A previous study showed that when both the donor and host expressed HLA-C1/HLA-C2/HLA-Bw4 ligands, NK cells expressing three inhibitory receptors (2DL1/2DL3/3DL1) exhibited maximum responsiveness against K562 cells and contributed to the lowest relapse rates in patients with AML and myelodysplastic syndrome following haplo-HSCT in the Chinese population (Zhao et al., 2019). KIR3DL1 alleles showed differential expression level and inhibitory capacity, and KIR3DL1005* combines the low frequency and level of expression with strong inhibitory function (Yawata et al., 2006). Interestingly, KIR3DL1005* and its associated haplotypes are associated with superior tyrosine kinase inhibitor (TKI) therapeutic effects, and the combinations of these KIR and HLA alleles may correlate with potent NK-cell immunity against CML (Ureshino et al., 2018). These studies further substantiated the reliability of our findings (Ureshino et al., 2018; Zhao et al., 2019). Moreover, numerous studies demonstrated that strongly inhibiting KIR receptors enable education and maturation of NK cells with strong effector functions (Yawata et al., 2006; Kim et al., 2008; Brodin et al., 2009), and higher numbers of inhibitory KIR ligands promote increased numbers of circulating NK cells, stronger killing capacity, and greater NK-cell repertoire diversity (Yawata et al., 2006; Brodin et al., 2009; Deng et al., 2021). These reports may partly explain why strong inhibitory KIR alleles for KIR2DL1, KIR2DL3, and KIR3DL1 in the presence of cognate ligands confer protective effects against leukemia in the Chinese population.
In the context of tumors and chronic infections, NK-cell exhaustion is characterized by the downregulated expression of activating receptors (e.g., NKG2D and FCGR3A), upregulated expression of inhibitory receptors (e.g., KLRC1, PD-1, TIGIT, and Tim-3), decreased production of effector cytokines (e.g., IFNγ and GZMB), and impaired cytolytic activity (Bi and Tian, 2017; Zhang et al., 2018; Jin et al., 2024). Previous reports demonstrated that the absence of inhibitory KIR–HLA interactions is associated with the induction of NK-cell exhaustion (Ardolino et al., 2014; Seo et al., 2017). We speculated that NK cells with the KIR AA genotype, which carries more inhibitory KIRs paired with cognate HLA ligands, may exhibit enhanced protection against exhaustion induced by prolonged activation. However, this hypothesis necessitates further experimental validation to confirm its validity.
KIR2DL1 alleles with KIR2DL1-R ^ 245 ^ (KIR2DL1002/001g*, 003, etc.) are functionally stronger than KIR2DL1 alleles with KIR2DL1-C245 (KIR2DL1004, 007, etc.) (Bari et al., 2009). Both KIR2DL1002 ^+^ NK cells and KIR2DL1003* ^+^ NK cells have a stronger degranulation compared to KIR2DL1004* ^+^ NK cells in C2 ^+^ donors (Dubreuil et al., 2020). The common KIR2DL1003*, less common KIR2DL1002*, and Cen-B-associated KIR2DL1004* are the three dominant KIR2DL1 alleles identified in the Chinese population. In the presence of the HLA-C2 ligand, the functionally stronger inhibitory KIR2DL1002* and 003 alleles carrying KIR2DL1-R ^ 245 ^ exhibit protective effects against ALL or AML in Chinese individuals with the KIR AA genotype, while the functionally weaker inhibitory allele KIR2DL1004 having KIR2DL1-C ^ 245 ^ confers ALL risk in individuals with the KIR Bx genotype. Our findings are consistent with previous studies (Bari et al., 2013), which reported that patients who received a donor graft containing KIR2DL1-R ^ 245 ^ had lower cumulative incidence of disease progression and better survival in allogeneic HSCT.
Among the 14 functional KIR genes, the framework gene KIR3DL3 is highly polymorphic with 235 alleles, which has been shaped by natural selection (Leaton et al., 2019). However, KIR3DL3 expression is inhibited by methylation of the promoter and is restored to the cell surface at similar levels to other KIR genes upon demethylation (Trompeter et al., 2005; Trundley et al., 2006). In the NK cells extracted from the blood of healthy human adults, KIR3DL3 expression is detected at a low level (Torkar et al., 1998; Long et al., 2001), but it is more common in the lungs and digestive tract (Palmer et al., 2023). KIR3DL3 interacts with the B7 family protein HERV-H LTR-associating 2 (HHLA2), which mediates inhibition in T cells and NK cells and is implicated for immune checkpoint targeting (Verschueren et al., 2020; Wojtowicz et al., 2020; Bhatt et al., 2021). However, little is known about the biological function of the KIR3DL3 molecule and the significance of its allelic polymorphisms. Our study, for the first time, found an association of KIR3DL3 allelic diversity with leukemia. KIR3DL3001* tended to confer protection against AML (P = 0.004, Pc = 0.06). In contrast, KIR3DL3009* conferred susceptibility to AML (Pc = 0.016). KIR3DL3001* differs from KIR3DL3009* by a single missense mutation at Codon 300 AAC>CAC, resulting in an amino acid substitution of non-charged asparagine (N) to charged histidine (H) in its transmembrane domain, suggesting that the functional variant KIR3DL3_N300H plays a critical role in protection against or susceptibility to ALL or AML in Chinese Hans. The further comparative analysis for KIR3DL3_300 allotypes also indicated that KIR3DL3_N300 alleles conferred protection against AML, whereas KIR3DL3_H300 alleles were associated with an increased risk of AML. Our findings may provide new insights into the role of KIR3DL3 polymorphisms in leukemia pathogenesis.
One limitation of this study is that the key aspects of the KIR3DL3 mechanism and the functional impact of its polymorphisms on leukemia remain to be elucidated. For leukemic patients, we failed to determine the expression of KIR3DL3 on NK cells and HHLA2 expression on leukemic blast, though HHLA2 is expressed and upregulated in tumors of the lung, gastrointestinal tract, kidney, and liver (Chen et al., 2019; Jing et al., 2019). Our future work will include the construction of NK cells with homozygous KIR3DL3001* ^+^ or KIR3DL3009* ^+^ and NK-cell cytotoxicity assay against HHLA2-positive target cells. Another limitation is that the relatively low sample numbers we obtained for the NK-cell cytotoxicity tests did not allow for a thorough analysis and validation of interactions between KIR allele and cognate ligand that conferred protection or susceptibility to AML/ALL among the randomly selected NK-cell donors and target cells. Such experiments require pre-genotyping of a large number of NK-cell donors.
In summary, the present study investigated the KIR allelic polymorphisms of all the seven functional KIR genes on the KIR A haplotype in Chinese AML/ALL patients and healthy controls. We found that individuals with the KIR AA genotype possess strong inhibitory KIR alleles in association with cognate ligands that are protective against acute leukemia. Notably, the allelic polymorphisms of the structure gene KIR3DL3 were associated with leukemia occurrence. Our findings may provide valuable insights into leukemia pathogenesis and better understanding of the immune mechanisms.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ardolino M. Azimi C. S. Iannello A. Trevino T. N. Horan L. Zhang L. (2014). Cytokine therapy reverses NK cell anergy in MHC-deficient tumors. J. Clin. Invest 124, 4781–4794. 10.1172/JCI 74337 25329698 PMC 4347250 · doi ↗ · pubmed ↗
- 2Augusto D. G. (2016). The impact of KIR polymorphism on the risk of developing cancer: not as strong as imagined? Front. Genet. 7, 121. 10.3389/fgene.2016.00121 27446203 PMC 4923111 · doi ↗ · pubmed ↗
- 3Babor F. Manser A. R. Fischer J. C. Scherenschlich N. Enczmann J. Chazara O. (2014). KIR ligand C 2 is associated with increased susceptibility to childhood ALL and confers an elevated risk for late relapse. Blood 124, 2248–2251. 10.1182/blood-2014-05-572065 25163702 · doi ↗ · pubmed ↗
- 4Bari R. Bell T. Leung W. H. Vong Q. P. Chan W. K. Das Gupta N. (2009). Significant functional heterogeneity among KIR 2DL 1 alleles and a pivotal role of arginine 245. Blood 114, 5182–5190. 10.1182/blood-2009-07-231977 19828694 PMC 2792213 · doi ↗ · pubmed ↗
- 5Bari R. Leung M. Turner V. E. Embrey C. Rooney B. Holladay M. (2011). Molecular determinant-based typing of KIR alleles and KIR ligands. Clin. Immunol. 138, 274–281. 10.1016/j.clim.2010.12.002 21239231 PMC 4733522 · doi ↗ · pubmed ↗
- 6Bari R. Rujkijyanont P. Sullivan E. Kang G. Turner V. Gan K. (2013). Effect of donor KIR 2DL 1 allelic polymorphism on the outcome of pediatric allogeneic hematopoietic stem-cell transplantation. J. Clin. Oncol. 31, 3782–3790. 10.1200/JCO.2012.47.4007 24043749 PMC 3795888 · doi ↗ · pubmed ↗
- 7Bhatt R. S. Berjis A. Konge J. C. Mahoney K. M. Klee A. N. Freeman S. S. (2021). KIR 3DL 3 is an inhibitory receptor for HHLA 2 that mediates an alternative immunoinhibitory pathway to PD 1. Cancer Immunol. Res. 9, 156–169. 10.1158/2326-6066.CIR-20-0315 33229411 PMC 8284010 · doi ↗ · pubmed ↗
- 8Bi J. Tian Z. (2017). NK cell exhaustion. Front. Immunol. 8, 760. 10.3389/fimmu.2017.00760 28702032 PMC 5487399 · doi ↗ · pubmed ↗