Longitudinal changes in circulating biomarkers from baseline to week 48 in treatment-Naïve people living with HIV initiating integrase inhibitor-based antiretroviral therapy
Jose-Ramon Blanco, Miguel Torralba, María Saumoy, Antonia Alcaraz, Mariano Matarranz del Amo, Julian Olalla, Fernando Dronda, Nisa Boukichou-Abdelkader, Enrique Bernal, Sergio Padilla, Joaquim Peraire, Francisca Artigues, María-Jesús Bustinduy-Odriozola, Helena Albendin Iglesias

TL;DR
This study examines how different HIV treatments affect immune and metabolic biomarkers over 48 weeks in people starting treatment for the first time.
Contribution
The study identifies distinct immunomodulatory effects of non-boosted INSTI-based regimens in treatment-naïve HIV patients.
Findings
All INSTI-based regimens achieved viral suppression and increased CD4+ counts, with the greatest increase in the DTG/ABC/3TC group.
The DTG/ABC/3TC regimen showed significant changes in IL-10, CD163, and ICAM-1 levels over 48 weeks.
Distinct longitudinal biomarker patterns suggest potential differences in immune modulation among INSTI-based regimens.
Abstract
Currently, integrase strand-transfer inhibitors (INSTIs) are the cornerstone of antiretroviral therapy (ART), providing potent viral suppression and good tolerability. Emerging evidence suggests that INSTI-based regimens may exert different effects on immune and metabolic pathways, potentially influencing inflammation and comorbidity risk. This study aimed to evaluate the impact of various first-line INSTI-based regimens on a panel of circulating biomarkers in treatment-naïve individuals with HIV. We included ART-naïve adults (≥18 years) with confirmed HIV-1 infection initiating a non-boosted INSTI according to the treating physicians’ decisions. The regimen included were bictegravir/emtricitabine/tenofovir alafenamide (BIC/TAF/FTC or Group 1 [G1]), dolutegravir/lamivudine (DTG/3TC or Group 2 [G2]), and dolutegravir/abacavir/lamivudine (DTG/ABC/3TC or Group 3 [G3]). Participants…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
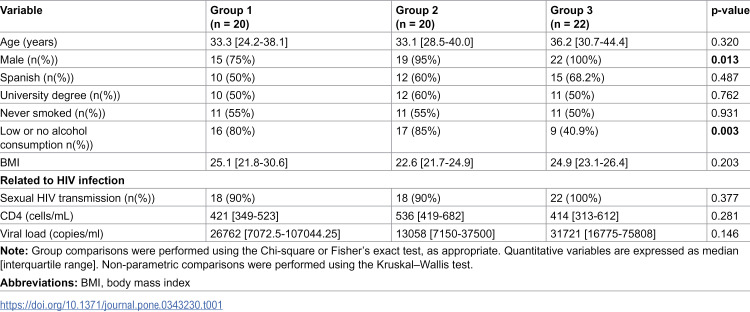 Figure 1
Figure 1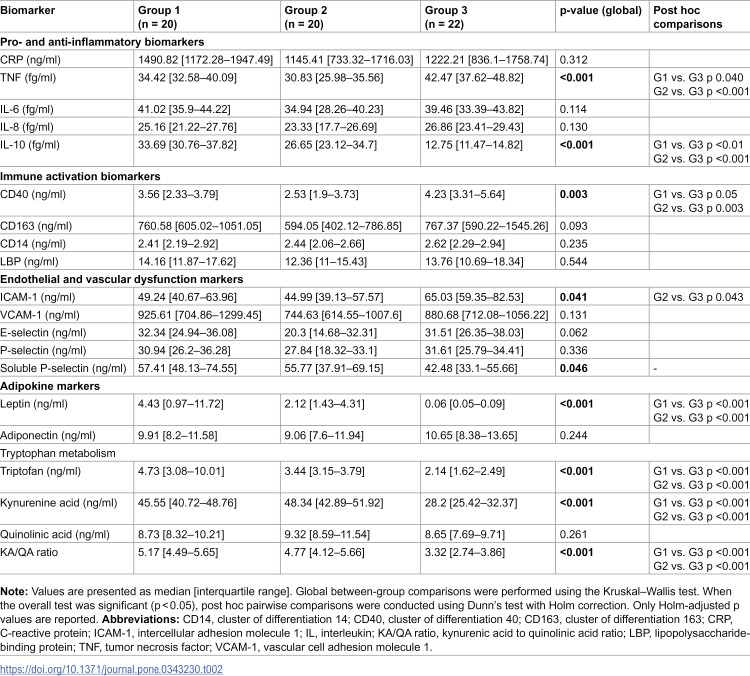 Figure 2
Figure 2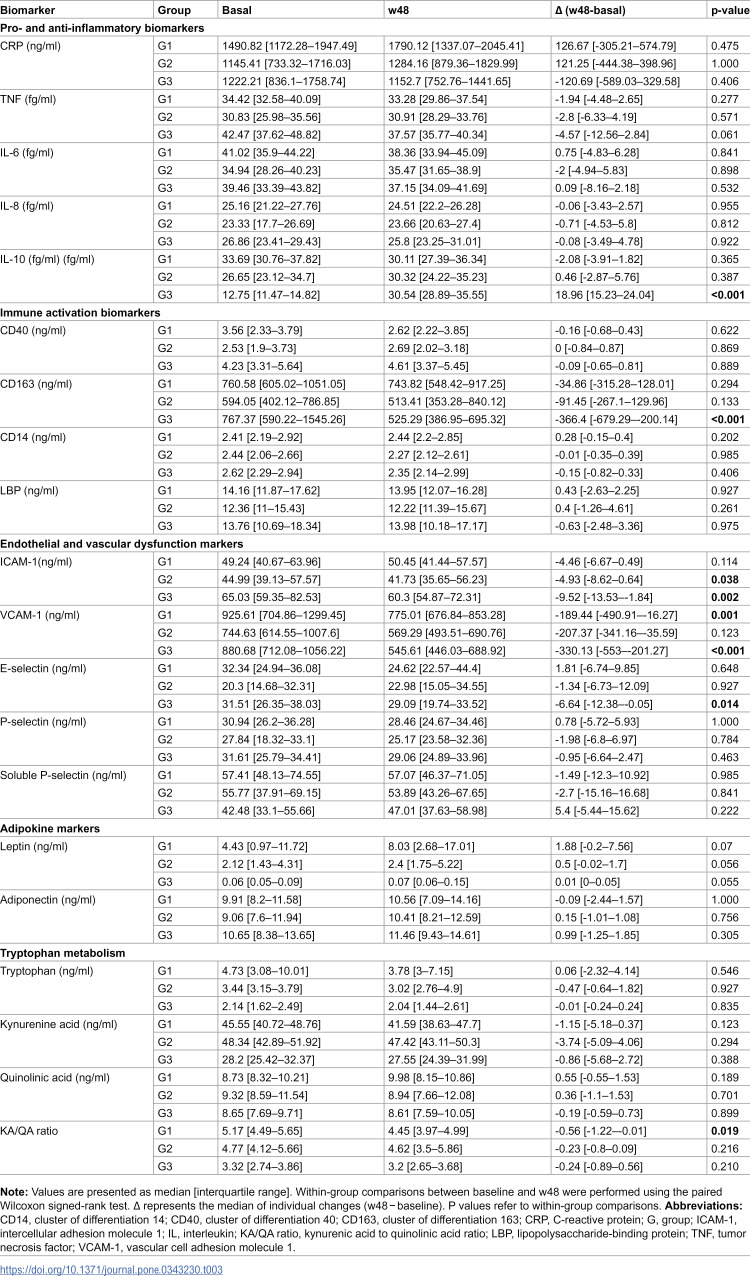 Figure 3
Figure 3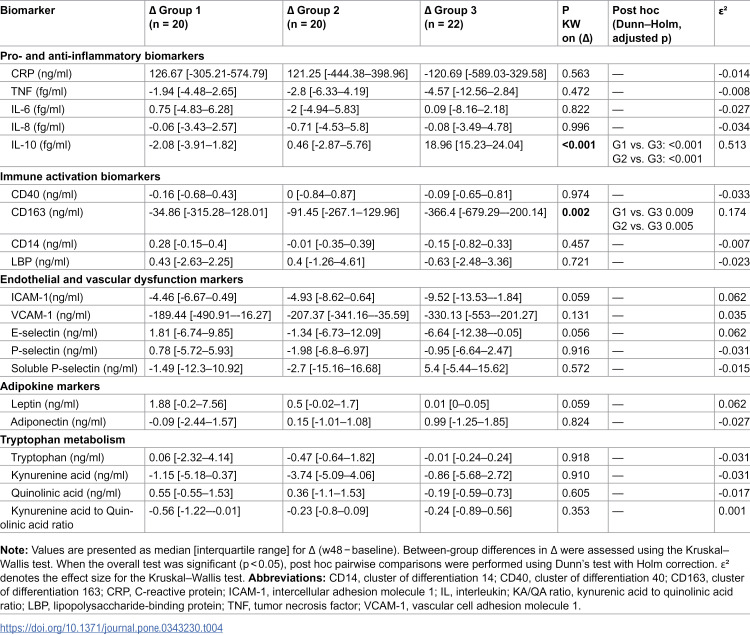 Figure 4
Figure 4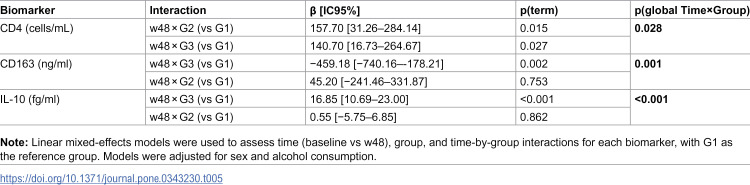 Figure 5
Figure 5- —http://dx.doi.org/10.13039/501100004587Instituto de Salud Carlos III
- —http://dx.doi.org/10.13039/501100004587Instituto de Salud Carlos III
- —http://dx.doi.org/10.13039/501100004587Instituto de Salud Carlos III
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHIV-related health complications and treatments · HIV/AIDS drug development and treatment · HIV/AIDS Research and Interventions
Introduction
The lifelong administration of antiretroviral therapy (ART) requires adherence to treatment strategies that ensure sustained both virological efficacy and long-term safety. However, prolonged ART exposure has been associated with a range of adverse effects [1], underscoring the need for robust evidence to guide optimal regimen selection.
Despite the efficacy of ART in suppressing viral replication, people living with HIV (PLWH) continue to exhibit a higher burden of non-AIDS comorbidities compared to uninfected individuals of similar age [2,3]. Multiple mechanisms contribute to the development of non-HIV-related diseases in PLWH, with persistent immune activation and chronic systemic inflammation playing a central role [4–6]. This inflammatory state may be further amplified by bacterial translocation, underlying comorbidities, and concurrent infections [7]. Circulating biomarkers such as C-reactive protein (CRP), interleukin-6 (IL-6), soluble CD14 (sCD14), soluble CD163 (sCD163), and soluble vascular cell adhesion molecule-1 (sVCAM-1) have been independently associated with all-cause mortality in PLWH [6,8,9]. Moreover, dysregulation of adiponectin levels and alterations in kynurenine pathway metabolites have been linked to an increased risk of non-AIDS-related clinical outcomes [10–12]. Nonetheless, there is currently no consensus regarding the most reliable inflammatory biomarkers for clinical prognostication.
Currently, integrase strand-transfer inhibitors (INSTIs) are a cornerstone of ART, providing potent viral suppression, high tolerability, and a favorable safety profile [13]. Their efficacy has led to their widespread recommendation as first-line therapy in treatment-naïve PLWH [14,15]. Among available INSTIs, dolutegravir (DTG) and bictegravir (BIC) are the most commonly prescribed. Emerging evidence suggests that INSTI-based regimens may differentially modulate immunologic and metabolic pathways [16–18], resulting in distinct profiles of systemic inflammation and immune activation. However, these effects are not consistently observed across all studies [19–21]. A deeper understanding of these differences may have important implications for the long-term risk of comorbidity in PLWH.
Understanding the impact of different initial INSTI-based regimens during effective ART is essential to improving our knowledge of the pathogenesis of HIV-related comorbidities. To address this knowledge gap, we assessed a panel of pathophysiological biomarkers in treatment-naive individuals initiating different INSTI-based regimen. Our objective was to determine whether specific regimens were associated with more favorable biomarker profiles after 48 weeks of therapy, thereby identifying potential advantages in reducing long-term comorbidity risk in PLWH.
Methods
Ethical considerations
Initial approval for this study was obtained from the Comité Ético de Investigación de La Rioja (CEImLAR), which acted as the reference ethics committee, and was subsequently obtained from the ethics committees of all participating institutions. All participants provided written informed consent for the collection and storage of blood samples, as well as for the collection of associated personal and clinical data. The CoRIS cohort was approved by the Research Ethics Committee of the Gregorio Marañón Hospital.
Participants
In this study (March 1, 2021 – June 30, 2024), we recruited ART-naive adults (aged ≥18 years) with confirmed HIV-1 infection. Participants were prescribed a non-boosted INST-regimen according to the treating physicians’ decisions in real-world clinical practice. The regimen included were bictegravir/emtricitabine/tenofovir alafenamide (BIC/TAF/FTC or Group 1 [G1]), dolutegravir/lamivudine (DTG/3TC or Group 2 [G2]), and dolutegravir/abacavir/lamivudine (DTG/ABC/3TC or Group 3 [G3]).
Eligible patients were required to have a baseline CD4 + T-cell count ≥200 cells/mL. Exclusion criteria included hepatitis B or C coinfection, new AIDS-defining conditions within 30 days of screening, pregnancy, and known CVD or diabetes mellitus. These criteria ensured a clinically stable, treatment-naïve population and reduced potential confounding from conditions known to influence circulating biomarkers.
Sample size was determined by the number of eligible participants during the recruitment period, which may limit power to detect modest between-regimen differences. Accordingly, the study was designed as exploratory and hypothesis-generating, with prespecified emphasis on within-person biomarker changes from baseline to w48, and between-regimen comparisons considered secondary.
No participants had documented SARS-CoV-2 infection during the study period.
Blood samples
Blood samples were collected at baseline and again during a second period, approximately 48 weeks after initiating ART. Serum samples were obtained from blood drawn during a study visit and stored at −80° C. For participants receiving DTG/ABC/3TC regimens, data were obtained from the Spanish HIV-positive cohort (CoRIS), and biological samples were provided by the HIV BioBank integrated within the Spanish AIDS Research Network. Briefly, CoRIS is a multicenter cohort that collects a standardized dataset, including sociodemographic, immunological, and clinical information at baseline and through follow-up [22,23]. This cohort is linked to a centralized biorepository, the Spanish HIV Hospital Universitario Gregorio Marañón Biobank (HIV HUGM BioBank) [24]. Participating centers follow standardized protocols to collect an initial blood sample prior to ART initiation and, when possible, additional samples annually during follow-up. All participants provided specific informed consent for the storage and use of their biological samples in the BioBank. To ensure confidentiality and compliance with ethical standards, the authors had no access to any personally identifiable information about the participants.
Circulating biomarkers
We assessed a comprehensive panel of circulating biomarkers, categorized into six categories: a) pro-inflammatory cytokines: C-reactive protein (CRP), tumor necrosis factor (TNF), interleukin (IL)-6, and IL-8; b) anti-inflammatory cytokines, represented by IL-10; c) immune activation markers: (sCD163), and lipopolysaccharide-binding protein (LBP); d) endothelial and vascular dysfunction markers: intracellular adhesion molecule-1 (ICAM-1), vascular cell adhesion molecule-1 (VCAM-1), E-selectin, P-selectin, and soluble P-selectin; e) metabolic and adipokine markers: leptin and adiponectin; and f) tryptophan catabolism markers: tryptophan, kynurenic acid, and quinolinic acid.
CD14, CD40, LBP, TNF, sCD163, ICAM-1, IL-6, IL-8, IL-10, E-selectin, P-selectin, VCAM-1 and adiponectin were measured using human magnetic Luminex assays (BioTechne, Minneapolis, USA; catalogue LXSAHM), following the manufacturer´s instructions. CRP, leptin, adiponectin, tryptophan, kynurenic acid, and quinolinic acid were quantified by an enzyme-linked immunosorbent assay kit (ELISA) according to standard protocols. Assay sensitivity and minimum detectable dose (MDD) are detailed in S1 Table.
This biomarkers panel was deliberately selected to capture complementary and interrelated pathways –chronic inflammation, immune activation, microbial translocation, endothelial dysfunction, metabolic dysregulation, and altered tryptophan metabolism– implicated in HIV pathogenesis and cardiometabolic comorbidities [6,8–12,25–30].
Statistical analysis
Descriptive statistics for baseline quantitative variables were summarized using medians and interquartile ranges (IQRs) due to non-normal distributions assessed by the Shapiro-Wilk test. Categorical variables were described using counts and percentages. For baseline comparisons between treatment groups, the Kruskal-Wallis test was used for continuous variables and the chi-square or Fisher’s exact test for categorical variables, as appropriate.
Missing data for “smoker”, “alcohol consumption frequency”, and “body mass index (BMI)” were imputed using multiple imputation by chained equations (MICE) [31], treating variables as ordered categorical (ordinal logistic regression; method = “polr”). Five imputed datasets were generated (m = 5, seed = 123) and estimates were pooled across imputations following Rubin’s rules.
Absolute changes from baseline to week 48 (w48) (Δ = w48 − baseline) were summarized using medians and IQRs and compared across groups using Kruskal-Wallis tests; when the overall test was significant (p < 0.05), post hoc pairwise comparisons used Dunn’s test with Holm adjustment. Effect sizes for Kruskal-Wallis tests were summarized using epsilon-squared (ε²). Within-group changes from baseline to w48 were additionally assessed using paired Wilcoxon signed-rank tests. Within-group changes from baseline to w48 were described using paired Wilcoxon signed-rank tests, whereas formal longitudinal inference relied on the main time effect and time-by-group interaction terms in the linear mixed-effects models.
Longitudinal changes were analyzed using linear mixed-effects models with random intercepts for each participant. Fixed effects included time (baseline vs w48), treatment group (with G1 as reference), and a time-by-group interaction, adjusting for sex and alcohol consumption, which differed at baseline between groups. Results are reported as regression coefficients (β) with 95% confidence intervals.
All statistical tests were two-sided. For post hoc comparisons, Holm-adjusted p values are reported; otherwise, p values are nominal. Given the exploratory nature and multiple biomarkers, results were interpreted with emphasis on effect sizes and confidence intervals rather than statistical significance alone. Analyses were conducted using R software [32] and relevant packages for data manipulation and modeling [33–36].
Results
Participant characteristics at baseline
A total of 62 naïve PLWH were included. Baseline characteristics by study group are shown in Table 1. Between-group differences were observed in sex distribution, with men predominating across all groups and reaching 100% in G3 (p = 0.013), and in alcohol consumption, with low or no alcohol use less frequent in G3 (p = 0.003). No significant between-group differences were observed in HIV-related parameters.
Table 1: Baseline characteristics by the study groups.
Baseline biomarker differences across regimens
Baseline biomarker levels were compared across groups using non-parametric tests (Table 2). Several inflammatory, immune activation, endothelial, adipokine, and tryptophan metabolism markers differed significantly between groups. Among pro- and anti-inflammatory biomarkers, TNF and IL-10 showed significant between-group differences, with higher TNF levels and markedly lower IL-10 concentrations in G3 compared with G1 and G2. For immune activation markers, CD40 differed significantly across groups, driven by higher levels in G3, whereas CD163, CD14, and LBP showed no significant baseline differences. Regarding endothelial and vascular dysfunction markers, ICAM-1 and soluble P-selectin differed significantly between groups, with higher ICAM-1 and lower soluble P-selectin levels observed in G3. In the adipokine profile, leptin levels were significantly lower in G3 compared with both G1 and G2, while adiponectin did not differ between groups. Markers of tryptophan metabolism showed pronounced baseline differences: tryptophan, kynurenic acid, and the KA/QA ratio were significantly lower in G3, whereas quinolinic acid levels were comparable across groups.
Table 2: Baseline biomarker levels by study group.
Post hoc pairwise comparisons using Dunn’s test with Holm correction indicated that the significant global effects for TNF, IL-10, CD40, ICAM-1, leptin, tryptophan, kynurenic acid, and the kynurenic acid-to-quinolinic acid (KA/QA) ratio were mainly driven by differences involving G3 vs. G1 and/or G2, whereas G1 vs. G2 comparisons were not significant after correction (Table 2). Although soluble P-selectin showed a significant overall Kruskal–Wallis test, none of the pairwise comparisons remained significant after adjustment.
Within-group changes from baseline to w48
Within-group analyses showed a significant increase in CD4 ⁺ T-cell count (cells/mL) from baseline to w48 in all groups (Table 3). Median changes were +143.5 (60–249.5) in G1 (p < 0.001), + 270 (152.5–545) in G2 (p < 0.001), and +331.5 (222–372.75) in G3 (p < 0.001), with the largest median increase observed in G3. All participants achieved an undetectable HIV viral load by w48. Beyond CD4 + recovery, most biomarkers showed limited within-group change over the 48-week follow-up. Among pro- and anti-inflammatory markers, IL-10 increased significantly in G3 (p < 0.001), while CRP, TNF, IL-6 and IL-8 did not change significantly in any group. For immune activation markers, CD163 decreased significantly in G3 (p < 0.001), whereas CD40, CD14 and LBP remained stable. Among endothelial and vascular dysfunction markers, ICAM-1 decreased in G2 (p = 0.038) and G3 (p = 0.002), and VCAM-1 decreased in G1 (p = 0.001) and G3 (p < 0.001). E-selectin decreased in G3 (p = 0.014), while P-selectin and soluble P-selectin showed no significant within-group changes. No significant within-group changes were detected for leptin or adiponectin, and tryptophan metabolism markers were largely stable, except for a modest reduction in the KA/QA ratio in G1 (p = 0.019).
Table 3: Within-group changes in biomarker levels from baseline to w48.
Between-group differences in changes from baseline to w48
Between-group comparisons of CD4 + T-cell recovery (Δ w48 − baseline) showed a significant difference across groups (Kruskal–Wallis p = 0.013; ε² = 0.113). Dunn–Holm post hoc testing indicated greater CD4 + gains in G2 and G3 compared with G1, whereas changes did not differ between G2 and G3. For other biomarkers, significant between-group differences in within-group changes were observed only for IL-10 and CD163 (Table 4). IL-10 exhibited a strong group effect (p < 0.001; ε² = 0.513), driven by a marked increase in G3 relative to both G1 and G2 (Dunn–Holm p < 0.001 for both). Similarly, changes in CD163 differed across groups (p = 0.002; ε² = 0.174), with a greater reduction in G3 compared with G1 and G2. No significant between-group differences in Δ were detected for the remaining pro- and anti-inflammatory markers, endothelial and vascular dysfunction markers, adipokines, or tryptophan metabolism markers (all p ≥ 0.056).
Table 4: Between-group comparison of within-group changes (Δ) from baseline to w48.
Longitudinal biomarker trajectories in mixed-effects models
Longitudinal changes were evaluated using linear mixed-effects models adjusted for sex, smoking status, and alcohol consumption, with G1 as the reference group. Models included fixed effects for time (baseline vs w48), group (G2 and G3 vs G1), and their interaction. Significant overall time × group effects (global interaction test) are summarised in Table 5.
Table 5: Significant time-by-group Interactions in linear mixed-effects models.
A significant time × group interaction was observed for CD4 ⁺ T-cell count (p(global) = 0.028), with both G2 (p(term) = 0.015) and G3 (p(term) = 0.027) differing from G1 in their w48 change. For CD163, the overall interaction was significant (p(global) 0.001), driven by G3 (p(term) 0.002) rather than G2. IL-10 also showed a significant overall interaction (p(global) <0.001), with a marked divergence in G3 (p(term) <0.001) compared with G1, while no interaction was detected for G2.
Discussion
In this prospective, non-randomized study of ART-naive individuals initiating INSTI-based regimens who successfully achieved virologic suppression, we observed differences in serum levels of some biomarkers across treatment groups. These findings suggest that inflammation in PLWH is a complex and multifactorial process, potentially modulated by the specific INSTIs regimen employed. To our knowledge, this is the first study to compare longitudinal inflammatory biomarker profiles across three commonly used INSTI-based first-line regimens in treatment-naïve PLWH. Our analyses focused on biomarkers previously linked to serious clinical outcomes [6,8–12,25–30].
Notably, absolute changes in quantitative biomarkers from baseline and w48 showed no significant differences among treatment groups in markers of endothelial and vascular dysfunction, adipokines, or tryptophan metabolism. These findings could support the hypothesis of a potential class effect, whereby these agents exert similar immunologic and inflammatory modulation. These observations are clinically relevant [37], as it suggests that the choice of INSTI may be guided by other therapeutic considerations, without a substantial differential impact on short-term systemic inflammation. In line with these findings, Bailon et al. [38] reported comparable levels of inflammatory markers (including sCD14, fatty acid–binding protein 2, tumor necrosis factor–related apoptosis inducing ligand, interferon-γ–induced protein 10, IL-6, CRP and D-dimer) and similar frequencies of activated and exhausted in ART-naïve PLWH treated with DTG/3TC or DTG/FTC/TAF. Similarly, a longitudinal analysis of cryopreserved samples from treatment-naïve individuals receiving BIC/FTC/TAF, DTG/ABC/3TC, or DTG/FTC/TAF found no significant differences in inflammatory biomarkers over time [20]; D-dimer, sCD14, and TNFR1 declined, while hsCRP and IL-6 remained stable [20]. At 12 months, Calza et al. [39] also observed similar reductions in hsCRP, IL-8, and TNF between BIC/FTC/TAF and DTG/3TC. In contrast, our study identified significant differences suggesting that biomarker responses may vary according to the INST-based regimen employed, particularly in G3. In comparison, G1 and G2 exhibited comparable outcomes, with no statistically significant differences observed between them.
Evidence from multiple studies suggest comparable CD4 + T-cell recovery, across INSTIs-based regimens. In the prospective CoRIS cohort, Suárez-García et al. [40] found no significant differences at w48 among ART-naïve PLWH treated with BIC/FTC/TAF (245 cells/mL), DTG/3TC (259 cells/mL), ABC/3TC/DTG (265 cells/mL). Bailon et al. [38], reported similar findings for DTG/3TC (241 cells/mL) and DTG/FTC/TAF (196 cells/mL). Gallant et al. [41] also found no difference between BIC/TAF/FTC (233 cells/mL) and DTG/ABC/3TC (229 cells/mL). A retrospective study [42] comparing DTG/3TC and BIC/FTC/TAF showed sustained CD4 + T-cell gains with no differences through w84. Quiros-Roldan et al. [43] observed improvements with different regimens including protease inhibitors and INSTIs (raltegravir [RAL], elvitegravir [EVG], and DTG), with only DTG showing significantly greater CD4 + T-cell count increases over time. Similarly, Gan et al. [17] found no significant differences at w48 between DTG/3TC (117 cells/mL) and BIC/FTC/TAF (102 cells/mL). No differences were observed in the study by Calza et al. [39] either. In our cohort, median CD4 + T-cell gains were unexpectedly low in G1 (143 cells/mL) and higher in G3 (331 cells/mL) compared to literature. Gains were +270 cells/mL in G2, with both G2 and G3 showing statistically significant differences versus G1. We also observed an increase in CD4 + T-cell counts over time, with group-specific trajectories in G2 and G3 relative to G1. Although we did not assess CD4/CD8 ratio dynamics in the present study, prior observational data in treatment-naïve individuals initiating ART have shown no differences in CD4/CD8 ratio normalization at w48 between DTG/3TC dual therapy and INSTI-based triple regimens [44], supporting comparable immunologic recovery. We acknowledge the lack of longitudinal CD8 measurements and CD4/CD8 ratio dynamics as an additional limitation, which should be addressed in future prospective studies. These patterns may reflect differential immune reconstitution dynamics, potentially influenced by treatment response or baseline immune status.
The increase in IL-10, a key immunoregulatory cytokine with potent anti-inflammatory properties and a critical role in modulating immune responses and limiting the development of inflammatory and autoimmune disorders [45], observed in G3 may reflect a compensatory anti-inflammatory response. Jianu et al. [46], reported a non-significant reduction in IL-10 levels in response to DTG-based treatment, highlighting the variability in IL-10 dynamics among different regimens. Similarly, Quiros-Roldan et al. [43] observed that one year of effective ART led to significant changes in multiple inflammatory biomarkers, including TNF, sCD14, hs-CRP, IL-7, D-dimer, sCD163, IL-6, and IL-10, although only TNF, sCD14, hs-CRP, IL-7, and sCD163 showed differential values between treatment groups. At 12 months, significant reductions were reported for TNF with DTG, sCD14 with RAL and DTG, D-dimer with all INSTIs, and sCD163 with RAL. In contrast, IL-6 and IL-7 increased significantly with EVG. In our case, IL-10 levels showed a highly significant time × group interaction in G3, despite the absence of a main time effect. This suggests that anti-inflammatory regulation may follow a distinct trajectory in G3, possibly influenced by specific clinical or immunological conditions unique to this group.
In relation to sCD163, this is a marker of macrophage activation involved in modulating inflammatory responses [47]. It plays a protective role by attenuating monocyte hyperactivation during infectious and inflammatory states, in part through the downregulation of pro-inflammatory cytokines such as TNF, IL-1β, IL-6, and IL-8 [47,48]. Additionally, sCD163 may suppress the production of pro-inflammatory chemokines like MCP-1 promote the release of anti-inflammatory mediators such as IL-10 [49], and limited the expression of pro-apoptotic transcription factors [48]. Clinically, however, has been associated with chronic immune activation and adverse outcomes in various inflammatory and infectious conditions, including HIV [47]. Regarding ART, Quirós-Roldán et al. [43] reported a significant decline in SCD163 with RAL but not with DTG. Similarly, Beltrán et al. [50] observed that sCD163 levels decreased after 48 weeks of ART, although levels remained higher than in HIV-negative controls. Notably, those receiving protease inhibitors had a smaller reduction in sCD163 levels than those on PI-sparing regimens. Importantly, no participants in that cohort received INSTIs, in contrast to our study. In our cohort, sCD163 levels decreased from baseline to w48, with the largest reduction observed in G3. In mixed-effects models, a significant time × group interaction indicated a distinct temporal profile in G3 relative to G1, consistent with group-specific modulation of monocyte/macrophage activation. These findings suggest differential inflammatory resolution across regimens, although mechanistic inferences should be made cautiously.
The simultaneous and significant modulation of IL-10 and sCD163 observed in G3 may reflect two non-mutually exclusive phenomena. This pattern may reflect activation of anti-inflammatory pathways and a shift toward an immunoregulatory phenotype, consistent with M2 macrophage polarization and IL-10-mediated immune modulation [51]. Alternatively, these changes could represent a compensatory response to immune activation or endothelial dysfunction, with IL-10 and sCD163 serving to limit inflammatory damage [52]. The magnitude and clinical relevance of these changes require confirmation in prospective studies with extended follow-up and validated endpoints.
Strengths
A key strength of this study is the prospective, real-world longitudinal profiling of a broad inflammatory biomarker panel from ART initiation to w48 across contemporary INSTI-based regimens under standardized procedures.
Limitations
This study has several limitations. First, the relatively small sample size limited statistical power to detect modest between-regimen differences, particularly across multiple biomarkers. Accordingly, the primary inference pertains to within-person change from baseline to w48, whereas between-regimen contrasts are secondary and exploratory. Second, all regimens evaluated were INSTI-based but differed in both the INSTI agent and the accompanying NRTI backbone; therefore, observed differences cannot be attributed to the INSTI component alone and should be interpreted as comparisons of complete regimens rather than head-to-head INSTI effects. Future studies including NNRTI- or PI-based regimens with similar NRTI components would be needed to more clearly disentangle drug class specific effects on immune and inflammatory biomarkers. Third, the non-randomized design may introduce selection bias and residual confounding from unmeasured factors despite multivariable adjustment. Fourth, baseline differences in several biomarkers indicate underlying heterogeneity at ART initiation, which may affect the interpretability of absolute between-group comparisons; this was addressed by focusing on within-person longitudinal changes and adjusting analyses for covariates that differed at baseline. Fifth, concomitant medications and lifestyle factors were not systematically captured and may have influenced biomarker trajectories; additionally, longer follow-up is needed to assess durability beyond w48. Sixth, sex-based analyses were not feasible due to baseline imbalance across groups, which is relevant given known sex-related differences in soluble mediators and HIV outcomes [53]. Seventh, use of archived biobank samples, although processed under standardized conditions, may introduce pre-analytical variability (e.g., storage duration and freeze-thaw effects). Eighth, missing data were handled using multiple imputation under a missing-at-random assumption, which cannot be formally verified and may introduce bias if violated. Ninth, only two time points were available, limiting the ability to characterize non-linear biomarker trajectories.
Although no formal adjustment for multiple comparisons was applied and p values are therefore nominal-raising the risk of type I error, particularly for between-regimen contrasts across the biomarker panel-these results should be interpreted cautiously and considered hypothesis-generating. In this context, the heterogeneity reported across studies of inflammatory biomarkers, likely driven by differences in populations, regimens and follow-up, further underscores the need for harmonized biomarker evaluation strategies [7].
Conclusion
While all INSTIs-based regimens were associated with immunologic improvement, modest differences in immune activation and regulatory markers suggest each regimen may have a distinct immunomodulatory profile. The clinical relevance of these differences remains uncertain and warrants investigation in longer-term studies with robust clinical endpoints. Future research should examine the relationship between biomarker dynamics and the incidence of comorbidities to assess whether these immunologic variations translate into meaningful health outcomes for PLWH. Long-term monitoring of these biomarkers may help personalize ART and identify patients who could benefit from tailored strategies to reduce comorbidity risk.
Supporting information
S1 TableLimits of detection for each biomarker.(PDF)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chawla A, Wang C, Patton C, Murray M, Punekar Y, de Ruiter A, et al. A Review of Long-Term Toxicity of Antiretroviral Treatment Regimens and Implications for an Aging Population. Infect Dis Ther. 2018;7(2):183–95. doi: 10.1007/s 40121-018-0201-6 29761330 PMC 5986685 · doi ↗ · pubmed ↗
- 2Teeraananchai S, Kerr SJ, Amin J, Ruxrungtham K, Law MG. Life expectancy of HIV-positive people after starting combination antiretroviral therapy: a meta-analysis. HIV Med. 2017;18(4):256–66. doi: 10.1111/hiv.12421 27578404 · doi ↗ · pubmed ↗
- 3Gueler A, Moser A, Calmy A, Günthard HF, Bernasconi E, Furrer H, et al. Life expectancy in HIV-positive persons in Switzerland: matched comparison with general population. AIDS. 2017;31(3):427–36. doi: 10.1097/QAD.0000000000001335 27831953 PMC 5302412 · doi ↗ · pubmed ↗
- 4Pandrea I, Landay A, Wilson C, Stock J, Tracy R, Apetrei C. Using the pathogenic and nonpathogenic nonhuman primate model for studying non-AIDS comorbidities. Curr HIV/AIDS Rep. 2015;12(1):54–67. doi: 10.1007/s 11904-014-0245-5 25604236 PMC 4369284 · doi ↗ · pubmed ↗
- 5Hsue PY, Deeks SG, Hunt PW. Immunologic basis of cardiovascular disease in HIV-infected adults. J Infect Dis. 2012;205 Suppl 3(Suppl 3):S 375-82. doi: 10.1093/infdis/jis 200 22577211 PMC 3349295 · doi ↗ · pubmed ↗
- 6Deeks SG, Tracy R, Douek DC. Systemic effects of inflammation on health during chronic HIV infection. Immunity. 2013;39(4):633–45. doi: 10.1016/j.immuni.2013.10.001 24138880 PMC 4012895 · doi ↗ · pubmed ↗
- 7Llibre JM, Cahn PE, Lo J, Barber TJ, Mussini C, van Welzen BJ, et al. Changes in Inflammatory and Atherogenesis Biomarkers With the 2-Drug Regimen Dolutegravir Plus Lamivudine in Antiretroviral Therapy-Experienced, Virologically Suppressed People With HIV-1: A Systematic Literature Review. Open Forum Infect Dis. 2022;9(4):ofac 068. doi: 10.1093/ofid/ofac 068 35265729 PMC 8900931 · doi ↗ · pubmed ↗
- 8Furman D, Campisi J, Verdin E, Carrera-Bastos P, Targ S, Franceschi C, et al. Chronic inflammation in the etiology of disease across the life span. Nat Med. 2019;25(12):1822–32. doi: 10.1038/s 41591-019-0675-0 31806905 PMC 7147972 · doi ↗ · pubmed ↗