Leptospira prevalence and lineages vary across land-use types due to shifts in small mammal communities
Jeanne A. Rajaonarivelo, Kayla M. Kauffman, Toky M. Randriamoria, James P. Herrera, Natalie Wickenkamp, Magali Turpin, Fiona Baudino, Hillary S. Young, Voahangy Soarimalala, Steven M. Goodman, Charles L. Nunn, Pablo Tortosa

TL;DR
Land-use changes in Madagascar affect small mammal communities and Leptospira infection rates, increasing zoonotic risk in disturbed habitats like rice fields.
Contribution
The study reveals how land-use changes drive shifts in host-pathogen assemblages, increasing Leptospira prevalence in non-native rodents in disturbed habitats.
Findings
Leptospira infection prevalence is higher in bats (37.7%) than in terrestrial small mammals (13.8%).
Non-native mice and rats are infected with cosmopolitan Leptospira species, while native species host distinct lineages.
Flooded rice fields have the highest Leptospira prevalence in mice, exceeding 50% infection rates.
Abstract
Human-induced land-use change can affect the composition of small mammal communities and the ecology of their zoonotic pathogens — yet questions remain on the direction and generality of these changes, which can have opposite effects on disease prevalence depending on the ecological context and pathogen involved. These contrasting patterns highlight the need to investigate how specific host-pathogen assemblages respond to local anthropogenic land-use mosaics. To address this need, we studied terrestrial and bat species composition, Leptospira infection prevalence, and Leptospira species composition across a mosaic of land-use types in northeastern Madagascar. We found differences in host communities between forested, agricultural, and village land-use types for both bat (n = 400) and terrestrial (n = 2,053) small mammal communities. Leptospira infection prevalence was higher in bats…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
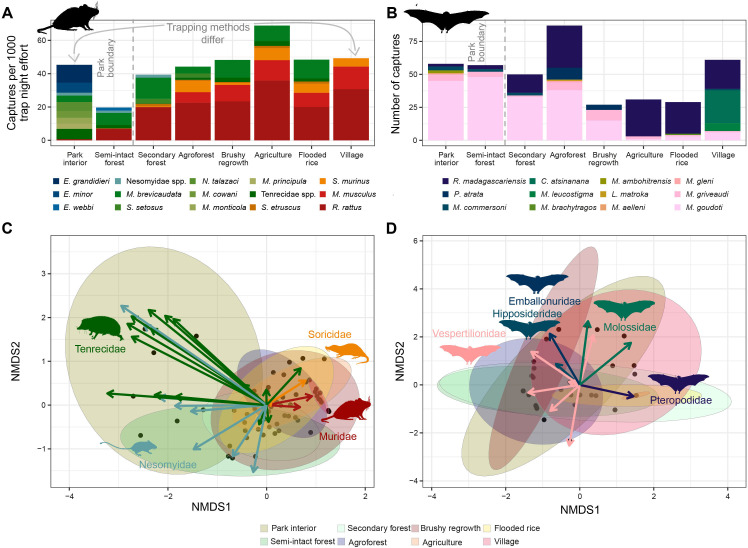 Fig 1
Fig 1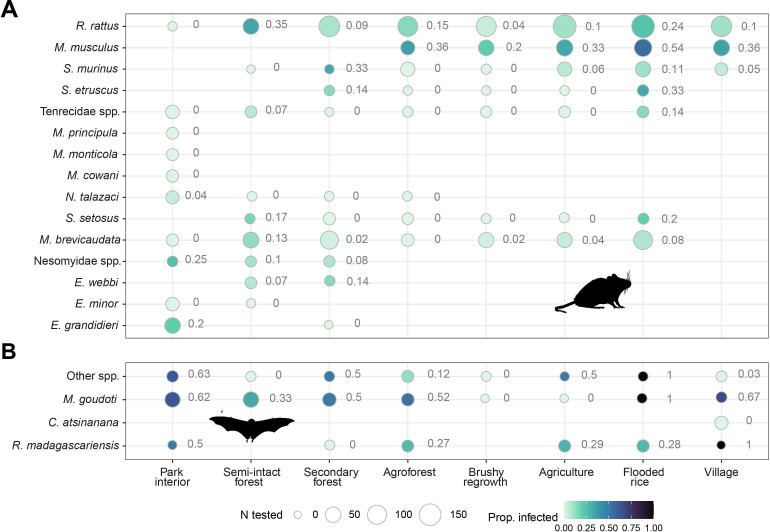 Fig 2
Fig 2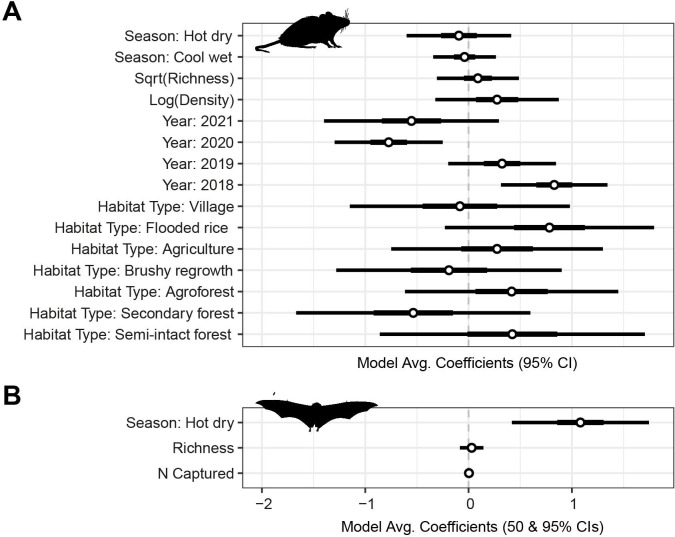 Fig 3
Fig 3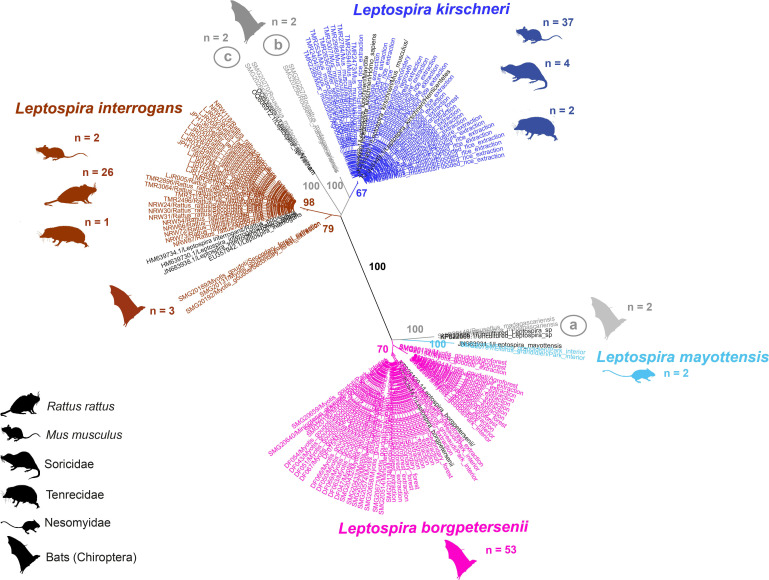 Fig 4
Fig 4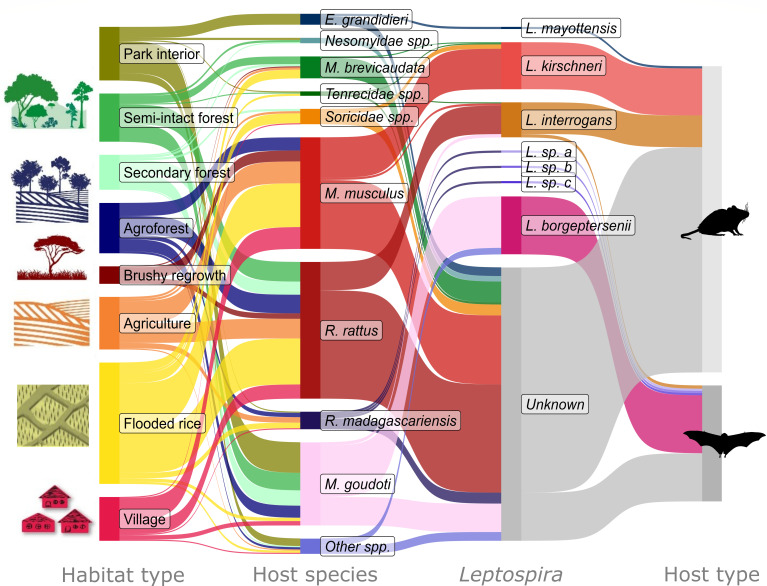 Fig 5
Fig 5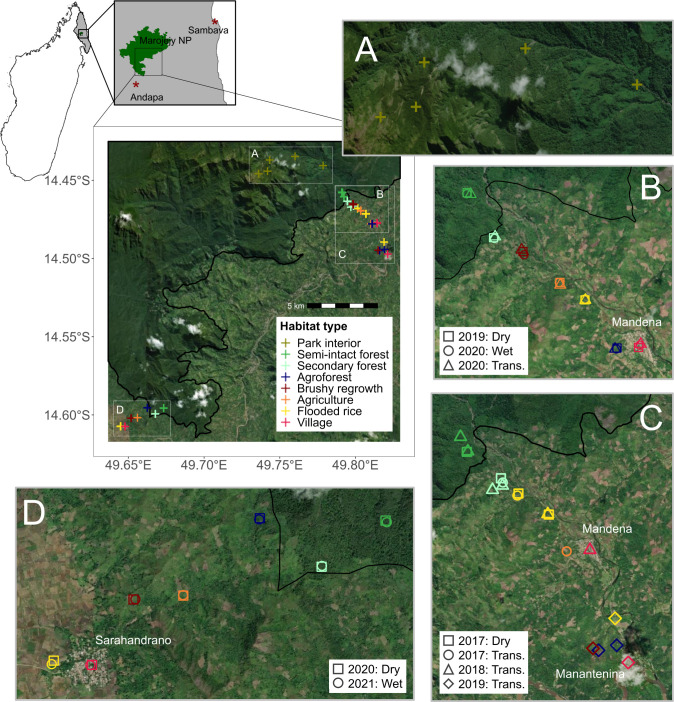 Fig 6
Fig 6- —Fogarty International Centerhttp://dx.doi.org/10.13039/100000061
- —FACE Foundationhttp://dx.doi.org/10.13039/100009879
- —Duke Universityhttp://dx.doi.org/10.13039/100006510
- —Duke Universityhttp://dx.doi.org/10.13039/100006510
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLeptospirosis research and findings · Yersinia bacterium, plague, ectoparasites research · Zoonotic diseases and public health
INTRODUCTION
Land-use change is considered a major driver of disease emergence and re-emergence globally (1–5). As natural habitats are converted into agricultural, anthropogenic systems, the species that rely on those natural habitats tend to decline. In contrast, synanthropic species that thrive in anthropogenic habitats tend to become more abundant (6, 7). The same mechanisms that allow these species to thrive in human-dominated landscapes — fast life histories, high densities, and being both diet and habitat generalists — also make them more likely to carry and transmit a wider range of parasites (8–10). Thus, the habitat-induced shifts in the host community are also altering the parasite community and potentially allowing for novel host-parasite associations (6, 11). Furthermore, synanthropic animals found in more disturbed habitats are more likely to harbor zoonotic parasites and transmit them to people due to their proximity and thus increased direct and indirect contact with people (7, 12, 13). Thus, it is vital to study how land-use change impacts host community composition and, in turn, impacts which parasites are present, their prevalence, and their host associations.
In reality, however, the disease-land use relationship is neither linear nor consistent across disease systems (1, 14). For example, the level of disturbance and habitat preferences of competent hosts result in Lyme prevalence often being highest at intermediate levels of disturbance, while plague prevalence in animals is highest at intermediate to high levels of disturbance (15, 16). The mechanisms for these changes are also varied: land-use change can alter environmental conditions to bring novel host species into contact (e.g., Nipah [17]), favor different vector communities (e.g., leishmaniosis [5, 18]), alter pathogen survival in the environment (e.g., Leptospira [19, 20]), and facilitate parasite invasions (e.g., Trypanosoma [21]). While some of these changes likely have generalizable patterns (e.g., all forms of land-use change likely tend to bring novel host species into contact), others will be idiosyncratic to the pathogen and type of land-use change (e.g., environmental changes to rice might be conducive to a waterborne pathogen while conversion to corn would not increase that pathogen). Predicting the impacts of land-use change on host-parasite-environment interactions often requires disentangling the effects of host community composition and environmental conditions on the transmission of a given parasite across land-use types (3).
The impact of land-use change on disease transmission is highly relevant in Madagascar, an island nation with high levels of species diversity, numerous zoonotic pathogens of public health relevance, extensive poverty, and limited access to healthcare (22–24). As a biodiversity hotspot, many animals native to Madagascar are endemic (i.e., occur nowhere else in the world), and little is known about the pathogens they carry. Native non-primate small mammals include 46 species of bats, 31 endemic species of tenrecs (Tenrecidae), and 28 endemic species of rodents (Nesomyidae, subfamily Nesomyinae) (25). Non-native (introduced) small mammal species in Madagascar include Muridae rodents (synanthropic mice and rats), which favor anthropogenic habitats and are well-known carriers of zoonoses (25, 26). Notable infectious agents with known or inferred zoonotic potential on Madagascar include hantaviruses (27, 28), astroviruses (29, 30), paramyxoviruses (31, 32), Yersinia pestis, the causative agent of plague (16, 33), enterotoxigenic Escherichia coli and Shigella, which are both responsible for diarrheal diseases (34, 35), and Leptospira, which causes leptospirosis (36, 37). These zoonotic pathogens are a public health challenge in many areas of the country, and the majority of these are primarily associated with non-native rodents as compared to native species (32). The transition from communities of predominantly native hosts with their endemic parasites to communities of predominantly non-native hosts and their cosmopolitan parasites makes Madagascar an ideal location for investigating host and parasite community composition shifts and novel host-parasite associations.
Madagascar is also experiencing dramatic land-use conversion. Rapid deforestation over the last 60 years has resulted in the island losing around 44% of its native forest (38). Nearly 80% of the Malagasy people live in rural areas, and the majority (82%) live below the poverty line and rely on natural resources and agriculture for subsistence (39, 40). Forests are transformed for the cultivation of rice, cassava, and other subsistence crops, particularly in the eastern portion of the country (41). Madagascar produces cash crops that are in high demand globally, including cloves, coffee, cocoa, and lychee, and is the world’s largest producer of vanilla (42–44), which is a significant source of income for farmers in the northeast (45, 46). Increases in demand for cash crops and soil depletion associated with hillside agricultural practices have accelerated the expansion of cultivation, leading to further forest fragmentation and transformation (47, 48). The combination of increased rates of land-use change, transitioning communities of mammal species and their associated pathogens, and close contact between humans and natural environments makes Madagascar an important location to address questions on how land-use conversion alters host and parasite communities and thus the ecology of infectious diseases (49).
Leptospira is an excellent organism for exploring how land use impacts pathogen dynamics due to environmental and host community composition changes. The different species and serovars of Leptospira tend to be associated with different sets of host species; however, the bacterium is only weakly host specific — one host species can be infected with multiple types of Leptospira, and one type of Leptospira can infect multiple host species. In Madagascar, L. mayottensis, alongside numerous other unnamed lineages, suggests the bacterium has undergone long-term coevolution with native small mammal hosts (50–54) and that L. mayottensis almost certainly originated on Madagascar (55). Three other lineages of Leptospira (interrogans, kirschneri, and borgpetersenii) are introduced and have undergone varying degrees of divergence, particularly in bats (25, 36, 50, 51, 53, 56, 57). These lineages can be broadly grouped into cosmopolitan (i.e., found worldwide) species (L. interrogans and L. kirschneri) and endemic species (L. borgpetersenii and L. mayottensis). The cosmopolitan species are generally found in Muridae rodents (36, 50, 51, 57), while the endemic species are primarily found in bats, tenrecs, and native rodents (53, 57).
In northeast Madagascar, Leptospira prevalence was higher in small mammals trapped in anthropogenic habitats where introduced rodents were more abundant than in native forested habitats (51). Anthropogenic land use promotes the colonization of introduced rats and mice, which harbor the cosmopolitan strains of L. interrogans and L. kirschneri, respectively, and consequently may modify local infection patterns (51, 58). Therefore, this is a model system to explore whether land use induces animal community composition changes that increase parasite prevalence and whether these host community changes result in different host-Leptospira species associations (51, 52, 59).
Leptospira is transmitted indirectly between animals and people through contaminated water and soil (60). Broadly, farmers in tropical regions are at high risk for infection, particularly those who use flooded rice farming techniques, such as those used in Madagascar (61). In Madagascar, Leptospira has been found in people, native and non-native wild animals, bats, cows, and pigs (25, 50, 56, 62–64). However, the epidemiologic data on Leptospira in people are limited; a single study reported a 2.9% (20/678) seroprevalence in people with serovars belonging to L. interrogans and L. noguchii (65). Clinical surveys carried out on neighboring southwestern Indian Ocean islands, as well as experimental studies, have brought insightful information on the virulence of Leptospira isolated from rats, tenrecs, and bats. Indeed, although L. mayottensis and borgpetersenii are zoonotic on Mayotte Island (Comoros Archipelago), their virulence is significantly lower than that of rat-borne L. interrogans (66, 67). Similarly, minimal clinical signs were found in experimental infection of hamsters, a model of acute infections in humans, with L. borgpetersenii isolated from Malagasy (68). Overall, this suggests that endemic species of Leptospira may be less pathogenic in people than cosmopolitan species and that land-use change may favor zoonotic lineages.
Across a mosaic of anthropogenic disturbance in and around Marojejy National Park in northeastern Madagascar (25), we investigated how land use corresponds with the composition of host communities, Leptospira infection prevalence, and Leptospira-host species associations in terrestrial small mammals and bats. Expanding on previous work on Malagasy small mammals at the same site (51), we used samples from additional terrestrial small mammals collected over an additional 3 years (in total 5 years) and bats (only during the added 3 years) across multiple land-use types. From these extensive host surveys, we estimated the composition, diversity, and relative abundance of terrestrial small mammal and bat hosts across the habitat mosaic to assess the differences in Leptospira infection prevalence among land-use types. Additionally, we used extensive sequencing data from animals infected with Leptospira to understand how the composition of Leptospira varied across hosts and habitats.
Using both host and pathogen data, we investigated the following questions: (i) How does the habitat type influence the species composition and density of terrestrial small mammals and bats? We hypothesized that native host species prefer forested or less disturbed habitat types, while synanthropic non-native species would occur at highest abundance in non-native habitats close to human settlements. (ii) How do host composition and environmental factors associated with land-use change impact Leptospira prevalence? Following previous findings (51), we hypothesized that prevalence would be highest in highly disturbed areas favored by non-native small mammal species, such as villages and exposed warm, wet areas such as rice fields, where Leptospira persists in the environment. (iii) Which lineages of Leptospira are found in which hosts and habitat types? This last question allows us to identify habitat types where pathogen switches have likely occurred. Based on previous reports of host-pathogen specificity, we hypothesized that bats and small mammals host different lineages of Leptospira. However, in places where a given host species was less abundant, we anticipated finding lineages typically associated with more abundant host species. Cumulatively, our study provides insights into the mechanisms by which land-use change may alter Leptospira risk by investigating associations between environment, small mammal community composition, and Leptospira lineages across a land-use mosaic.
RESULTS
Small mammal communities across land-use types
Terrestrial small mammals
From 2017 to 2021, 2,053 terrestrial small mammals representing 28 species were captured, including several species endemic to Madagascar from two groups: tenrecs from the Tenrecidae family (Microgale spp., Nesogale spp., Oryzorictes hova, Setifer setosus, and Tenrec ecaudatus) and native Nesomyinae rodents (Eliurus spp., Nesomys spp., and Voalavo gymnocaudus; Fig. 1A; Table S1). However, the majority (n = 1,354; 65.9%) of captured animals were non-native Muridae rodents (Rattus rattus and Mus musculus) and Soricidae shrews (Suncus murinus and S. etruscus). Rattus were captured in all habitat types, including inside the park; they were the most common species captured (n = 898; 43.7%), followed by Microgale brevicaudata (n = 366; 17.8%), introduced mice (n = 273; 13.3%) and shrews (n = 183; 8.9%). Mus were trapped in anthropogenic habitats and absent from forest habitats.
(A) Terrestrial small mammal composition transitioned from predominantly native Nesomyidae and Tenrecidae communities, shown in blues and greens inside the park interior, to predominantly non-native Muridae communities on the park exterior. For visual clarity, species with less than 10 individuals captured were grouped by family (see Fig. S2 for all species). (B) Bat species composition also shifted from Vespertilionidae communities in forested areas, Pteropodidae (Rousettus madagascariensis) in agricultural areas, rice fields and villages, and Molossidae communities around the homes in the villages. The two-dimensional non-metric multidimensional scaling (NMDS) plots display the pronounced separation of terrestrial mammals (C) between forested and non-forested habitat types and the lack of this contrast among bats (D). The ellipses in the NMDS plots represent the 95% confidence intervals of the total population by habitat type (t-distributions), and the arrows indicate the strength and direction of the linear correlation between each species and the ordination scores. The arrows are colored by family (see Fig. S3 for plots with arrows labeled by species).
Across habitat types, we found significant differences in the proportion of captures of native species (ANOVA: F = 20.25, df = 7, P < 0.001), animal density (number captured/trap effort; ANOVA: F = 5.05, df = 7, P < 0.001), species richness (ANOVA: F = 6.56, df = 7, P < 0.001), and Shannon diversity (ANOVA: F = 5.03, df = 7, P < 0.001), while species evenness did not differ significantly across habitat types (ANOVA: F = 0.784, df = 7, P = 0.604). Animal density tended to be lower in the semi-intact forest (Fig. S1, Table S2). Species richness was significantly higher in the park interior than in all other habitat types (Tukey post hoc test: P < 0.05; Table S2) except agroforest (P = 0.16) and agriculture (P = 0.28), but these differences only resulted in higher Shannon’s diversity in the park compared to the secondary forest (P = 0.01), brushy regrowth (P = 0.03), and village (P < 0.001; Table S2). The proportion of native species was significantly higher in forested habitats inside the park than outside, and lower in the village than in the secondary forest (Fig. S1, Table S2). The trapping methods limited our ability to make comparisons between the village, where traps were placed inside people’s homes, and the park interior, where transects and a greater ratio of pitfall traps to Sherman/National traps were used, and all other habitat types, where uniform trap grids and pitfall lines were used (see Methods). Notably, most tenrecs (94.0%, 691/735) were captured in pitfall traps.
These community differences result in a pronounced separation in two-dimensional NMDS ordination space (Fig. 1C), with tenrec and endemic rodent species indicating trap grids located in the park interior and less disturbed forests, and introduced Muridae and Soricidae species indicating habitat types outside of the park. The semi-intact forest, located just inside the national park, bridged the differences between inside and outside the park due to the presence of two tenrec species (Microgale brevicaudata and Setifer setosus), which were captured both in forests and in cultivated fields outside forests, and by the presence of two endemic rodents (Eliurus webbi and Nesomys audeberti). The animals captured in each habitat varied in both species composition (PERMANOVA: F = 5.06, df = 7, P < 0.001) and variability (PERMDIST: F = 2.92, df = 7, P = 0.012).
Bats
From 2019 to 2021, 400 bats representing 12 native species (Fig. 1B; Table S2) were captured, including Molossidae (Chaerephon atsinanana and Mops leucostigma), Vespertilionidae (Laephotis matroka and Myotis goudoti), Hipposideridae (Macronycteris commersoni), Emballonuridae (Paremballonura atrata), and Pteropodidae (Rousettus madagascariensis). Though native, none of these species is endemic to Madagascar. Myotis goudoti (n = 189; 47.2%) was the most captured species and was found in all habitat types, though more frequently in forested habitats. Rousettus madagascariensis, the second most abundant species (n = 125; 31,2%), was primarily captured in agricultural fields and in the village.
Compared to terrestrial small mammals, the distribution of species composition in bat communities varied less between land-use types (Fig. 1D; Fig. S1). We found no significant variation in the number of animals captured (ANOVA: F = 0.81, df = 7, P = 0.57), species evenness (Kruskal-Wallis: χ^2^ = 1.76, df = 7, P = 0.97), species richness (Kruskal-Wallis: χ^2^ = 3.47, df = 7, P = 0.84), and Shannon’s diversity index (Kruskal-Wallis: χ^2^ = 2.27, df = 7, P = 0.94) between habitat types. However, we did observe significant differences in communities by habitat type in NMDS space (Fig. 1D). These differences were due to compositional differences (PERMANOVA: F = 2.80, df = 7, P < 0.001) and not variability (PERMDIST: F = 1.78, df = 7, P = 0.109).
Leptospira infection prevalence across land-use types
Terrestrial small mammals
Out of 2,053 captured terrestrial small mammals, 2,032 individuals were tested, and 280 were qPCR positive, leading to an overall infection prevalence of 13.8%. Most infected terrestrial animals, 86.1% (n = 241), were introduced species, while native species represented only 13.9% of the positive cases (n = 39; Table S1). The most infected host species was Mus musculus (100 qPCR positive/272 individuals tested; 36.8%), followed by Rattus rattus (125/896; 13.9%), Suncus spp. (14/177; 7.9%), Setifer setosus (3/54; 5.5%), and Microgale brevicaudata (20/333; 6.0%). No infected individuals were found in several endemic species; however, few individuals of these species were captured (Table S1; range 1 to 31). Assuming a population prevalence equal to what we observed in our abundant native species (5.0%), a sample size of 73 individuals per species would be needed to determine that no individuals of a particular species were infected (power 80%, α = 0.05).
The proportion of infected individuals (prevalence herein) was highest in the rice fields, semi-intact forest, and village homes, and lowest in the park interior, secondary forest, and agricultural areas (Fig. 2A; Table S1). The majority of mice captured in flooded rice fields were infected (40/74; 54%). Rattus rattus and M. brevicaudata were more commonly infected in the semi-intact forest, and shrews (Suncus murinus) were more commonly infected in the secondary forest.
The proportion of infected (A) terrestrial small mammals and (B) bats by species and habitat type. The size of the circles represents the number of animals tested, and the adjacent numbers and shades of the circles represent the proportion of those animals that were infected. Species with fewer than 10 individuals tested across all habitat types were grouped by family (terrestrial small mammals) or as “other species” (bats). See Table S1 for the complete results of captures and testing in all species.
Binomial models of the number of infected animals of those tested during each trapping period were used to consider the joint effects of species composition and habitat type while accounting for seasonal and interannual variability. The most important predictors of infection were habitat type and year, followed by log animal density, square-root species richness, and season (AICc sum weights [sw] = 1.0, 1.0, 0.56, 0.22, 0.15; Table S3). In agreement with the habitat-wise estimates of prevalence, the log odds of infection for terrestrial small mammals were highest in flooded rice fields (β = 0.78 ± 0.52) and semi-intact forest (β = 0.42 ± 0.65), and lowest in the secondary forest (β = −0.54 ± 0.58; Fig. 3A; Table S3). Large interannual differences were found between terrestrial small mammals captured in 2018 and 2019 and those captured in 2017, 2020, and 2021. Samples in the 2018-2019 period had higher odds of infection than in the other years. Due to the study design, disentangling the effects of study locality and interannual variability (year) was not possible, as localities were sampled sequentially, not simultaneously. Both species richness (β = 0.09 ± 0.2) and density (log number of captured terrestrial mammals per night’s trap effort [β = 0.28 ± 0.3]) were weakly positively associated with the log odds of infection. Though the effects of seasonality were weak, infection odds were slightly higher in the warm wet season (reference category) than in both other seasons (β_cool,wet_ = −0.04 ± 0.34; β_hot,dry_ = −0.09 ± 0.26).
Full model-averaged coefficients plot for infection probability in (A) terrestrial small mammals and (B) bats. Bars show the 50% and 95% confidence intervals for the estimated coefficients. Reference variables were set as follows: 2017 (year for terrestrial small mammals), warm wet (season), and park interior (habitat type). See Table S3 for numeric estimates.
Bats
Infection prevalence in bats (37.7%; 106 qPCR positive/281 individuals tested) was significantly higher than in terrestrial small mammals (Kruskal-Wallis: χ^2^ = 100.07, df = 1, P <0.001). The most abundant bat species, Myotis goudoti, displayed the highest prevalence (76/153; 49.7%), which was much greater than that of the second most abundant species, Rousettus madagascariensis (16/58; 27.6%). We identified infected individuals of all but one species of bat (n = 7) in the family Vespertilionidae, and no infected Molossidae (n = 25) or Emballonuridae (n = 13) individuals, despite having sufficient power to do so, if infection prevalence was similar to what we observed in R. madagascariensis (sample size =9, power 80%, α = 0.05).
We identified infected bats captured in all habitat types except brushy regrowth, with the highest proportion of infected animals found in the park interior (36/58; 62.1%), secondary forest (15/35; 42.9%), and rice fields (10/23; 43.5%; Fig. 2B; Table S1). In general, the variability in infection prevalence between habitat types was greater in terrestrial small mammals than in bats. However, similar to terrestrial small mammals, no linear trends with the level of disturbance were observed (Fig. 2B). Binomial models of the number of infected bats of those tested during each trapping period revealed that habitat type was not important to the overall odds of infection in bats; however, season (sw = 1), species richness (sw = 0.33), and the number of animals captured (sw = 0.28) were important (Table S3). Because bats were trapped over fewer periods than terrestrial small mammals, season and year were aliased (trapping only occurred in the hot, dry period of 2019). Thus, only the season was considered in the bat model. Contrary to the terrestrial mammals, we found a strong seasonal effect in infection odds in bats, which were more likely to be infected during the hot dry season than the warm wet season (β = 1.08 ± 0.34; Fig. 3B; Table S3). However, due to the schedule of trapping bats, we cannot distinguish between season and location-level effects. Again, both species richness (β = 0.03 ± 0.06) and density (number of captured bats [β = 0.00 ± 0.01]) were weakly positively associated with the log odds of infection.
Leptospira lineages by host species and land-use type
We successfully sequenced the secY gene of 136 of our 380 positive samples, 62 of which were from bats and 74 from terrestrial small mammals. Phylogenetic analysis identified Leptospira at the species level and showed a division into several distinct genetic clades (Fig. 4) primarily along Leptospira species lines, with a few additional subgroups (a–c). The separation between host species, habitat type, and Leptospira lineage revealed that some Leptospira species were classified as “cosmopolitan” or associated with non-native species and disturbed habitats, and others as “forest-associated” or found in forest habitats and native species (Fig. 5; Fig. S2, Table S4).
Maximum-likelihood (ML) radial phylogenetic tree built with 136 Leptospira secY sequences from small mammal species trapped in northeast Madagascar and 15 reference sequences (in black). The numbers represent the bootstrap percentage from 1,000 replicates and are indicated when above 50%.
Sankey diagram showing the connections between habitat type, animal host species, and Leptospira species. The flows indicate the number of individuals connecting each habitat, host, infection, and host type. Samples that were positive but were not successfully sequenced are indicated as “Unknown.” Only Leptospira-positive animals are included in this figure, and species with fewer than 10 infected individuals are grouped by family (terrestrial mammals) or as “Other species” (bats). All species and all captured animals are shown in interactive Fig. S4. The diagram was created using the “ggsankey” package (69).
The three clades that corresponded with endemic Leptospira lineages were primarily found in bats and native species of terrestrial small mammals. All the sequences clustered with the L. borgpetersenii reference sequences were from Malagasy bats (Fig. 4). The majority of L. borgpetersenii sequences corresponded to the haplotype closely related to secY^3^48 and were found in Myotis (n = 45) and Miniopterus (n = 6) species. The remaining two sequences of L. borgpetersenii were assigned to a haplotype close to secY^3^61 and found in Myotis species. Most of these individuals were trapped in forest habitats, but a few individuals trapped in the rice fields (n = 3) and village (n = 3) were also infected with this clade of Leptospira (Fig. 5). Both of the sequences in the L. mayottensis cluster were from endemic rodents (Eliurus grandidieri) captured inside the national park ([Fig. 4 and 5](#F4 F5)). The haplotype of these samples was closely related to secY^3^56. The final endemic clade (herein referred to as Leptospira species a) was closely related to L. mayottensis. The two specimens forming this clade were obtained from Rousettus madagascariensis.
The four “cosmopolitan” clades consisted of L. interrogans, L. kirschneri, and two novel groups (L. sp. b and L. sp. c). The sequences forming the clade corresponding to reference L. interrogans were from terrestrial mammals (n = 29) and M. goudoti bats (n = 3) (Fig. 4). The haplotype closely related to the secY^3^13 was found in all R. rattus (n = 26) which were mostly trapped in agricultural fields, in one M. musculus trapped in the village and another captured from the flooded rice, and in a single tenrec (Setifer setosus) captured in flooded rice. The bat sequences in this clade were from two M. goudoti captured in the secondary forest and another captured in the agroforest (Fig. 5). These bat-associated sequences were closely related to the secY^3^9 haplotype, forming a distinct subgroup (Fig. 4).
All sequences in the clade associated with L. kirschneri reference sequences were assigned to a single secY^3^22 allele. Most of the samples in this group were M. musculus (n = 37). However, samples from one tenrec (S. setosus) and three shrews (S. murinus) captured in flooded rice, and samples from one shrew (S. etruscus) and one tenrec (M. brevicaudata) captured in the secondary forest were also in this clade (Fig. 5). The remaining two clades, Leptospira species b (n = 2) and c (n = 2), were similar to Leptospira species a in that they were found in Rousettus madagascariensis trapped in agricultural habitats (Fig. 5).
DISCUSSION
Land-use change alters small mammal communities, which have cascading effects on parasite communities and zoonotic risk (70). Through our multi-year, multi-location effort, we found that Leptospira prevalence and which bacterial species are present vary across land-use types corresponding to shifts in host communities. We found that the species composition of bats and terrestrial small mammals varied between habitat types, with notable shifts between forested habitats, agricultural land-use types, and the village homes. Leptospira infecting these animals also changed from predominantly cosmopolitan lineages (i.e., L. interrogans and L. kirschneri) in disturbed habitats to endemic lineages (L. borgpetersenii and L. mayottensis [53, 57, 71]) in more intact, forested habitats. This resulted in community-level infection prevalence varying with species composition between habitats and Leptospira shifting from endemic-associated lineages in forested habitats to cosmopolitan lineages in agricultural fields and village homes, where invasive Muridae rodents were abundant. Overall, Leptospira infection prevalence was much higher in bats (37.7%) than in terrestrial small mammals (13.8%).
The changes in species composition between habitat types were primarily due to changes in the relative abundance of a small number of species and, to a lesser extent, species diversity. While the study site and our samples exhibited high species richness (28 species of terrestrial mammal and 12 species of bats were captured), most individuals were of just a few species: Rattus rattus, Microgale brevicaudata, and Mus musculus among the terrestrial animals and Myotis goudoti and Rousettus madagascariensis among bats. In terrestrial small mammals, introduced Muridae rodents were the primary species captured outside of the national park, and many native species were limited to the forest interior. These differences in animal captures resulted in higher terrestrial animal density and lower species richness in the agricultural matrix than in forested habitats, except for richness in flooded rice fields. As hypothesized, and as is typical of many insular terrestrial small mammal systems (32), we found that invasive species replaced natives, particularly in disturbed land-use types.
Bat communities transitioned from being *M. goudoti-*dominated in forested and brushy regrowth habitat types to *R. madagascariensis-*dominated in agricultural land-use types, with agroforest having roughly equal proportions of these two species. The differences in bat communities between forested and agricultural land-use types were primarily due to the abundance of these two species, not metrics of diversity. The lack of a strong relationship between habitat and species composition may reflect that bats, with their greater dispersal capabilities via flight and associated home ranges, were less restricted to the habitat in which they were captured, since trapping location was likely more reflective of where animals feed than where they roost (72, 73). This may also be in part due to some species of bats benefiting from human-modified habitats such as villages, agricultural fields, and agroforests, because these sites can be used for their roosting and foraging activities (74). Therefore, shifts in animal density and which terrestrial small mammal and bat species were most common transitioned between forested and anthropogenic land-use types to favor non-native small mammals and fruit bats (R. madagascariensis).
The most abundant terrestrial mammal host species in the anthropogenic land-use types tended to be more highly infected. The synanthropic Mus musculus was the primary host of Leptospira in this region (36.8% overall prevalence), and the majority (54%) of mice captured in flooded rice fields were infected. Nevertheless, R. rattus remains an important host due to its higher abundance — they accounted for 43.7% of animals captured — despite lower infection prevalence (13.9%). In absolute terms, more individual R. rattus were infected than any other species in our study. This result sharply contrasts with previous findings from the capital city of Antananarivo, where Mus were highly abundant but rarely infected, compared to Rattus norvegicus (56). Interestingly, the infection prevalence in the second most abundant terrestrial mammal species, M. brevicaudata, was quite low (6.0%), and more broadly, prevalence was lower in native species (9.5% ± 22.2%) than in non-native species (18.7% ±11.6%).
Terrestrial small mammals captured in flooded rice fields were the most likely to be infected, followed by those captured in the semi-intact and villages. The semi-permanent water in rice paddies offers optimum conditions for survival and transmission of this primarily water-borne zoonotic pathogen (75–77). However, the dense vegetation under the forest canopy and the largely year-round rain also maintain humidity on the soil surface in forest habitats, which also favors bacterial survival (19). Rice paddies are likely a source of infection for animals and Malagasy rice farmers.
Interannual and seasonal differences were stronger predictors of infection prevalence than richness or density for both terrestrial and volant animals. Prevalence in terrestrial mammals was higher in the earlier years of the study, which corresponds to the animals that were also included in a previous analysis (51). The reason for this large shift is unknown, and because bats were not captured during the earlier phase of the study, we cannot know whether a similar drop in prevalence occurred in bats. The warm, wet season, which is most favorable for Leptospira, corresponded to slightly higher infection prevalence in terrestrial mammals, but interestingly, the opposite trend was observed in bats. However, this unexpected, large effect of bats having a higher probability of infection in the hot, dry season may simply reflect that this was the only season when animals were captured deep in the park interior, where prevalence was highest. Furthermore, some of the regional variability in habitat type was captured in the year and seasonal model terms since trapping locations covaried with season (bats) and year (terrestrial small mammals).
Leptospira lineages infecting small mammals also shifted between land-use types from endemic lineages (L. borgpetersenii and L. mayottensis [53, 57, 71]) in more intact, forested habitats to predominantly cosmopolitan lineages (L. interrogans and L. kirschneri) in disturbed habitats. This change in Leptospira lineages paralleled the changes in host species composition as the endemic Leptospira lineages only infected native small mammals and bats, while the cosmopolitan lineages infected both native and non-native species. The cosmopolitan clades consisted of L. interrogans, L. kirschneri, and closely related L. sp. b and L. sp. c, and endemic clades consisted of L. borgpetersenii, L. mayottensis, and the closely related L. sp. a. Cosmopolitan Leptospira were primarily found in non-native terrestrial mammals, captured in the more disturbed habitat types, but were also found in non-native terrestrial mammals captured in the forested habitat types. Some of these cosmopolitan Leptospira lineages are known to be zoonotic. For example, we found L. kirschneri, haplotype (secY^3^22), which has been previously found in Hemicentetes semispinosus, a native tenrec species trapped in the humid forest of central-east Madagascar (64), in mice from Portugal (78), and in people from Mayotte Island (79). The L. interrogans, closely related to the secY^3^13 haplotype, was found in non-native R. rattus, along with two M. musculus and an endemic species of tenrec, Setifer setosus. This haplotype was also found in dogs in nearby villages (62), R. norvegicus in urban areas of Guangzhou, China (80), and Kuwait, and R. tanezumi in Thailand (81).
All the sequences obtained from animals captured inside the national park corresponded to the endemic clades of Leptospira. However, we did not obtain sequences for half of the positive specimens from inside the park, where infection prevalence was quite high (45.8% ± 16.2%). While this limits our ability to fully assess whether the shift in host assemblies between forested and non-forested habitats coincides with a shift in Leptospira lineages, we do find some evidence that this shift occurred. The forest-associated Myotis goudoti and Miniopterus spp. were mostly infected with L. borgpetersenii closely related to haplotypes secY^3^48 and secY^3^61. Alignments of L. borgpetersenii secY^3^48 identified in bats were similar to sequences identified from endemic small mammals, including Eliurus minor, Microgale principula, and M. majori captured in other areas of Madagascar (64). Leptospira mayottensis was only found in E. grandidieri, a native rodent species restricted to elevational ranges between 1,250 and 1,875 m on the Marojejy Massif (82). The observed haplotype secY^3^56 of Leptospira mayottensis was previously reported in spiny tenrecs, Tenrec ecaudatus (introduced to neighboring islands in the western Indian Ocean, including Mayotte), and shrew tenrecs, Microgale thomasi (55, 71). The restricted geographical distribution of the native host limits the dispersion of their agent pathogens, causing strong host-parasite specificity and co-radiation processes that may lead to endemism of these microorganisms (55, 64). This is the second report of L. mayottensis in native Malagasy rodents (Nesomyinae), which have previously been found to have mixed infections with L. interrogans and/or L. borgpetersenii (52).
Similar to previous reports of Leptospira in Malagasy bats, we found considerable diversity, including lineages distinct from other Leptospira species and clades (53, 57). The bats infected with distinct lineages of Leptospira most closely related to L. kirschneri were captured in more disturbed areas. Despite L. sp. a being most closely related to L. mayottensis, which was only found in terrestrial mammals in the park, it was found in Rousettus madagascariensis in agriculture and agroforestry. Leptospira sp. b was only found in bats (n = 2) captured in the flooded rice, where L. kirschneri, its most closely related species, was most commonly found. However, L. sp. c may be a *Rousettus-*specific strain of Leptospira since it was closely related to samples obtained from R. leschenaultii in Vietnam (83). Finally, Myotis goudoti were infected with a L. interrogans haplotype closely related to secY^3^9, which was distinct from rat-associated L. interrogans and did not have any clear habitat-type associations. Conclusions regarding the host specificity of these bat-associated Leptospira are limited because we were unable to obtain sequences for most R. madagascariensis samples. Next steps for describing Leptospira in this region are to isolate Leptospira from these animal hosts so they can be included in a serologic microscopic agglutination test (MAT). With a MAT test, human and animal exposures can be used to assess which animals are the hosts of zoonotic Leptospira, quantify the public health impacts of leptospirosis, and identify serovar-matched bacteriocin vaccines (84).
In conclusion, our study provides a new example of how the conversion of forests into agricultural fields and other anthropogenic habitats increases the prevalence of infection with potentially zoonotic organisms (85, 86). In this case, the primary mechanism involved seems to be that invasive rats and mice are the dominant terrestrial small mammal species in anthropogenic habitats. These rodents tend to have higher infection prevalence and be infected with potentially zoonotic lineages of Leptospira, and thus their occurrence corresponds to elevated landscape-level risk (6, 7). At our study site, these introduced rodents host globally distributed haplotypes of L. interrogans and L. kirschneri, thus suggesting that these cosmopolitan Leptospira were introduced to Madagascar alongside their synanthropic hosts and have altered the parasite community in the agricultural habitats where they are highly abundant.
Notably, we found little to no effect of land-use change on Leptospira prevalence and lineages in bats. This is likely due to differences in the ecology of bats, which are highly mobile, have large home ranges, and were captured where they fed instead of where they roost. Anthropogenic change may well be changing landscape-level infection prevalence of Leptospira in bats, but at a larger spatial scale than encompassed in our study. Altogether, our work demonstrates that converting forests to agricultural fields results in the replacement of native species with invasive species and their parasites when investigated at a spatial scale relevant to the host community.
MATERIALS AND METHODS
Sample collection
Data collection was approved by Institutional Animal Care and Use Committee at Duke University (protocol number A002-17-01 2017-2019, A262-19-12 2019-2021), and by Malagasy authorities (No. 289/17, 146/18, 280/19, 57/20, 191/20, 307/21, 357/21—MEEF/SG/DGF/DSAP/SCB). Samples from small mammals were collected between 2017 and 2021 inside Marojejy National Park and around three adjacent villages: Mandena (−14.477049, 49.8147), Manantenina (−14.497213, 49.821347), and Sarahandrano (−14.607567, 49.647759). The protected area is located in the SAVA region of northeastern Madagascar and has an area of 60,150 ha and an elevational range from 75 to 2,133 m.
Animals were captured in eight different habitat types: (i) primary forest in the park’s interior, (ii) semi-intact forest situated just inside the park, (iii) secondary forest fragments located outside the park that have experienced human or natural perturbation, (iv) agroforest containing agricultural vanilla and cultivated trees, (v) brushy regrowth areas containing shrubs and bushes occurring after swidden cultivation practice, (vi) open mixed agricultural fields with less tree cover, (vii) flooded rice fields, and (viii) in homes and other village structures. Sampling was performed during three field sessions: May 2017 to July 2019 in Mandena and Manantenina; September 2019 to September 2021 in Mandena and Sarahandrano; and October to November 2021 inside Marojejy National Park (see Fig. 6 for more details). Sherman and National traps were placed at 10 m intervals in 90 × 90 m (before September 2019) or 100 × 100 m trap grids (after September 2019). Two pitfall lines of 100 m long (all dates) were installed 20–50 m distance from the trap grid, with 11 buckets (pitfall traps) of 15 L placed every 10 m along each line. Additionally, traps were placed inside people’s homes in the villages. Bats were captured using mist nets (12 × 2.6 m or 6 × 2.6 m) and harp traps (1.0 × 2.6 m) installed along trails and paths adjacent to the terrestrial mammal trap grids. Traps placed in the park interior (not semi-intact forest) were installed along an elevational transect at five sites ranging from 480 to 1,880 m encompassing the swath of the Marojejy Massif. At each of these five sites, two trap lines of 50 traps and three pitfall lines (11 buckets spaced 10 m apart with vertical drift fencing between) were installed for seven consecutive nights. For all habitats outside of the elevational transect, the number of nights of trap effort varied between habitat type and study period, ranging from 5 to 10 consecutive nights. In total, 46,506 nights of trapping occurred (6,941 for pitfall buckets and 39,565 for traps). Non-native and a subset of native (less than 10 for rare species) animals were euthanized and necropsied for a range of different zoonotic disease projects. Relevant to this study, kidney tissues were stored in 70% ethanol and transported to the laboratory for molecular analyses, where they were stored at −20°C. The taxonomy of captured small mammals follows Soarimalala and Goodman (2011) (87).
Map of the sampling sites in (A) Marojejy National Park interior and in the three locations (B) Mandena, (C) Mandena & Manantenina, and (D) Sarahandrano where animals were trapped in different land use types and during three distinct seasons: warm wet period (March–May), cool wet period (June–September), and hot dry period (October–December). Maps were made using the sf (88, 89) and basemaps (90) packages. The country and regional insets were made using shapefiles available in the geodata package (91) and from Protected Planet (92). Basemaps are Esri “World Imagery.”
Tissue processing and Leptospira screening
Kidney tissues were rehydrated overnight in 1.5 mL of autoclaved milli-Q water to remove ethanol, then 20 mg of kidney tissue was finely cut with a sterile scalpel and lysed at 56°C for 3–6 h in 180 µL ATL buffer (QIAGEN, Valencia, California, USA) supplemented with 20 µL proteinase K (IndiSpin QIAcube HT Pathogen Kit 480). Total nucleic acids were extracted using a QIACUBE robot and the Cador Pathogen kit (Qiagen, Valencia, California, USA) per the manufacturer’s instructions with a 200 µL elution volume. Reverse transcription was performed on 10 µL of nucleic acids using the ProtoScript II Reverse Transcriptase and Random Primer 6 (New England BioLabs, Ipswich, Massachusetts, USA) under the following thermal conditions: 70°C for 5 min (denaturation 1), 4°C for 2 min, 25°C for 10 min (denaturation 2), 42°C for 50 min (annealing), and 65°C for 20 min (extension). Reverse transcription was used to screen both RNA (viral) and DNA pathogens with the same cDNA preparation.
The presence of Leptospira in the samples was tested using a previously published probe-specific real-time PCR targeting bacterial 16S (93), and samples with a cycle threshold (Ct) value < 40 were considered positive. Five microliters of cDNA was amplified in a reaction mixture composed of 10 µL of Quanti Nova Probe PCR (Qiagen), 0.5 µL (10 µM) of each primer (reverse and forward), 0.4 µL (10 µM) of the probe, and 3.6 µL of RNAse-free water. Thermal conditions consisted of an initial denaturation step (95°C for 2 min) followed by 45 cycles of annealing (95°C for 5 s) and extension (60°C for 5 s). For all positive samples (Ct value < 40), Leptospira were characterized at the species level using the secY gene, a housekeeping gene commonly used for species identification of pathogenic Leptospira infecting small mammals (64). For this, 2 µL of cDNA from each RT-PCR-positive sample was used as PCR template in a reaction mixture composed of 12.5 µL of GoTaq G2 Hot Start Green Master Mix (Promega, Madison, Wisconsin, USA), 0.5 µL (10 µM) of each primer, and 9.5 µL of RNAse-free water. PCR conditions were as follows: a first GoTaq activation step (95°C for 2 min), then 45 cycles of denaturation (94°C for 30 s), annealing (58°C for 30 s), and extension (72°C for 1 min), then a final extension step (72°C for 5 min). When we failed to obtain sequences with standard secY primers (94), further analysis using alternative degenerate (64) and secYinner primers (95) was performed. Sanger sequencing was carried out on both strands (Genoscreen, Lille, France), and the obtained chromatograms were manually aligned and edited using Geneious software v.9.1.8.
Statistical analysis
All statistical analysis was performed in R software v.4.4.1 (96). We visualized patterns in small mammal communities in association with land use type by using NMDS analysis. Next, we used a PERMANOVA to test whether the community composition of small mammals and a PERMDIST to test whether community variability (dispersion) differed across habitat types. For each habitat type, we calculated species richness (the number of species of small mammals or bats), Shannon diversity (accounts for the abundance and evenness of the species present), and evenness using the diversity function in the “vegan” package (97). An ANOVA followed by a Tukey’s post hoc test for pairwise comparisons was used to compare species richness and diversity between habitat types.
To investigate the effect of environmental variables and species composition on Leptospira prevalence, we used binomial models of the number of infected animals given the number tested in each trapping period (i.e., the 5- to 10-day period when traps were installed in each habitat type in each season). In the models of the terrestrial mammal infections, the environmental variables were the type of habitat (park interior, semi-intact forest, secondary forest, agroforest, brushy regrowth, agriculture, flooded rice, and village), season (warm wet, cool wet, and hot dry), year (2017, 2018, 2019, 2020, and 2021), and locality (Mandena, Manantenina, Sarahandrano, and park interior). The species composition variables included Shannon diversity, species richness, evenness, animal density (number of small mammals trapped per night of trap effort), and the proportion of captured animals that were native species. In the models of bat infections, the environmental variables included the same habitat types, but year (2019 and 2021), season (hot dry and cool wet), and locality (Mandena, Sarahandrano, and park interior) were modified to include just the periods in which bats were captured. The species composition variables in the models of bat infections were Shannon diversity, richness, evenness, and the number of animals captured at each location. Due to issues with multicollinearity and singularity, Shannon diversity and locality were dropped from both sets of models.
To identify which factors best predict infection with Leptospira, all combinations of predictors were performed using the “dredge” function with the “MuMIn” package (98). The best model subset, containing models with differences less than two of the corrected Akaike Information Criterion (AIC), was then used to find variable importance (sum AIC weight for all models in the best subset containing the variable). The full model-averaged coefficients were used to describe the effect of each variable on the odds of detecting animals infected with Leptospira during each trapping period.
Phylogenetic analysis
Phylogenetic analysis was performed with all secY sequences obtained and 15 reference sequences that were aligned with Seaview software v.5. The phylogenetic tree was inferred using ML with default settings, including a General Time Reversible model for base pair substitutions and 1000 bootstrap replicates, and then edited with FigTree software v.1.4.4. Allelic numbers for the secY gene were assigned using the Public PubMLST database (scheme#3). Leptospira species assignment was carried out using the Leptospira isolates database available on PUBMLST, and 783 fully or partly genotyped strains were used with secY^3^ allele determination.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Carlson CJ, Brookson CB, Becker DJ, Cummings CA, Gibb R, Halliday FW, Heckley AM, Huang ZYX, Lavelle T, Robertson H, Vicente-Santos A, Weets CM, Poisot T. 2025. Pathogens and planetary change. Nat Rev Biodivers 1:32–49. doi:10.1038/s 44358-024-00005-w · doi ↗
- 2Faust CL, Mc Callum HI, Bloomfield LSP, Gottdenker NL, Gillespie TR, Torney CJ, Dobson AP, Plowright RK. 2018. Pathogen spillover during land conversion. Ecol Lett 21:471–483. doi:10.1111/ele.1290429466832 · doi ↗ · pubmed ↗
- 3Lambin EF, Tran A, Vanwambeke SO, Linard C, Soti V. 2010. Pathogenic landscapes: interactions between land, people, disease vectors, and their animal hosts. Int J Health Geogr 9:54. doi:10.1186/1476-072X-9-5420979609 PMC 2984574 · doi ↗ · pubmed ↗
- 4Johnson PTJ, Preston DL, Hoverman JT, Richgels KLD. 2013. Biodiversity decreases disease through predictable changes in host community competence. Nature 494:230–233. doi:10.1038/nature 1188323407539 · doi ↗ · pubmed ↗
- 5Glidden CK, Nova N, Kain MP, Lagerstrom KM, Skinner EB, Mandle L, Sokolow SH, Plowright RK, Dirzo R, De Leo GA, Mordecai EA. 2021. Human-mediated impacts on biodiversity and the consequences for zoonotic disease spillover. Curr Biol 31:R 1342–R 1361. doi:10.1016/j.cub.2021.08.07034637744 PMC 9255562 · doi ↗ · pubmed ↗
- 6Mc Farlane R, Sleigh A, Mc Michael T. 2012. Synanthropy of wild mammals as a determinant of emerging infectious diseases in the Asian-Australasian region. Ecohealth 9:24–35. doi:10.1007/s 10393-012-0763-922526750 PMC 7088064 · doi ↗ · pubmed ↗
- 7Gibb R, Redding DW, Chin KQ, Donnelly CA, Blackburn TM, Newbold T, Jones KE. 2020. Zoonotic host diversity increases in human-dominated ecosystems. Nature 584:398–402. doi:10.1038/s 41586-020-2562-832759999 · doi ↗ · pubmed ↗
- 8Becker DJ, Streicker DG, Altizer S. 2018. Using host species traits to understand the consequences of resource provisioning for host-parasite interactions. J Anim Ecol 87:511–525. doi:10.1111/1365-2656.1276529023699 PMC 5836909 · doi ↗ · pubmed ↗