International Society of Sports Nutrition position stand: effects of dietary antioxidants on exercise and sports performance
Drew E. Gonzalez, Broderick L. Dickerson, Brandon M. Roberts, Jennifer A. Kurtz, Hunter S. Waldman, Adam M. Gonzalez, Matthew J. McAllister, Jeffery L. Heileson, Richard J. Bloomer, Shawn M. Arent, Darren G. Candow, Jeffrey R. Stout, Karen A. Hecht, Bill Campbell

TL;DR
This paper summarizes the International Society of Sports Nutrition's official stance on how dietary antioxidants affect exercise performance and recovery, emphasizing a balanced and individualized approach.
Contribution
The paper provides a comprehensive, evidence-based position on the use of dietary antioxidants in sports, highlighting optimal strategies for performance and recovery.
Findings
Dietary antioxidants can aid recovery but may hinder training adaptations at high doses.
Whole foods are preferred over isolated supplements for antioxidant intake.
Creatine, omega-3s, tart cherry, and astaxanthin show strong evidence for antioxidant benefits.
Abstract
Following a comprehensive review, the International Society of Sports Nutrition (ISSN) has developed an official position on the role of dietary antioxidants in exercise and sport. Antioxidants play a complex, context-dependent role in vivo; they can facilitate recovery from exercise but may also hinder training adaptations when consumed at supraphysiological doses. While endogenous antioxidant systems can effectively maintain redox balance, dietary sources, particularly whole foods, can help mitigate excessive oxidative stress following intense/heavy training or inadequate recovery. The influence of dietary antioxidants depend on timing, dosage, type, and individual factors. The ISSN’s official position encompasses the following: (1) Redox balance exists on a spectrum, with mild oxidative eustress driving beneficial physiological adaptations and excessive oxidative distress impairing…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
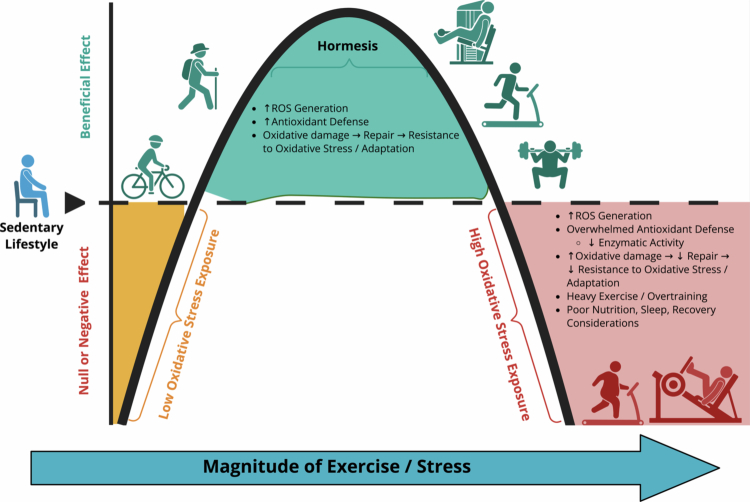 Figure 1
Figure 1| Antioxidant class | Key examples/subclasses | Common food sources | Solubility | Structural characteristics | Antioxidant mechanisms |
|---|---|---|---|---|---|
| Flavonoids | Flavonols, flavones, flavanones, flavanols, isoflavones | Fruits, vegetables, herbs, cereals, nuts, seeds, stems, flowers | Water-soluble | Benzopyran backbone (C6-C3-C6); phenolic or polyphenolic groups |
Hydrogen/electron donation from hydroxyl (–OH) groups to neutralize free radicals (e.g. ROO·, ·OH) Metal chelation (e.g. Fe²⁺) via 3’,4’-dihydroxy B-ring or 3-OH/4-keto on C-ring Induction of Nrf2 signaling |
| Anthocyanins (Flavonoid subclass) | Cyanidin, delphinidin, malvidin | Berries, red/purple grapes, red cabbage, eggplant skin | Water-soluble | Flavylium ion; pH-dependent color shifts; C6-C3-C6 skeleton |
Hydrogen/electron donation from –OH groups Transition metal chelation Support of endogenous defense via Nrf2 |
| Polyphenols (Non-flavonoid) | Phenolic acids, stilbenes (e.g. resveratrol), lignans | Tea, coffee, wine, whole grains, seeds, legumes | Mostly water-soluble | Multiple phenol rings; varied backbones |
Direct ROS/RNS scavenging Modulation of signaling pathways like Nrf2 |
| Tocopherols and Tocotrienols | α-, β-, γ-, δ-tocopherol | Vegetable oils, nuts, seeds, green leafy vegetables | Lipid-soluble | Chromanol ring + phytyl tail (tocopherol); unsaturated tail (tocotrienol) |
Lipid peroxyl radical scavenging (ROO·) in cell membranes Chain-breaking antioxidant in lipid environments |
| Carotenoids | Astaxanthin, β-carotene, lutein, lycopene, zeaxanthin | Carrots, tomatoes, sweet potatoes, spinach, kale | Lipid-soluble | Long conjugated double-bond system (polyene chain) |
Quenching of singlet oxygen (¹O₂) Scavenging peroxyl and other radicals Indirect modulation of gene expression |
| Trace Elements (Cofactor-based antioxidants) | Selenium, Zinc, Copper, Manganese | Brazil nuts, seafood, meat, whole grains, legumes | Varies (bioavailable as ions or chelated forms) | Inorganic elements, often bound in metalloenzymes (e.g. GPx, SOD) |
Cofactors for endogenous antioxidant enzymes (e.g. Glutathione Peroxidase, Superoxide Dismutase) Facilitate neutralization of H₂O₂ and superoxide anion (O₂·–) Indirectly support redox homeostasis |
| Antioxidant | Mechanism of action | Potential benefits | Dosage & duration | Level of evidence to support efficacy | |
|---|---|---|---|---|---|
| Performance | Antioxidant | ||||
| Alpha-lipoic acid | Scavenges ROS, recycles glutathione, metal chelation | May aid recovery, reduce muscle damage and oxidative stress | 100–1800 mg/day for up to 6 months | Weak/Low | Weak/Low |
| Ashwagandha | Scavenges free radicals, modulates Nrf2 and NF-κB pathways | Improves strength, power, recovery; modulates cortisol/testosterone | 240–1250 mg/day for ≥4–12 weeks | Weak-to-moderate | Weak/Low |
| Astaxanthin | Lipid membrane protection, scavenges ROS/RNS | Enhances fat oxidation, reduces muscle damage, improves recovery | 4–12 mg/day for 4–12 weeks | Moderate | Weak-to-moderate |
| Beetroot | Nitrate donor, improves oxygen efficiency (details pending) | Improves oxygen efficiency, may enhance endurance | 250–500 mL/day of beetroot juice (containing 300–600 mg of nitrates) for single, 3–15 days, or up to 4–6 weeks | Moderate-to-high | Weak/Low |
| Blackcurrant | High anthocyanins; antioxidant; NO signaling; mitochondrial protection | Improves endurance and increases fat oxidation | 300 mg/day extract providing 105–210 mg anthocyanins for 7 days; 1–2 h pre-exercise | Moderate | Weak/Low |
| Cocoa flavanols | Flavanol-rich extract, supports NO production, improves endothelial function | May improve vascular function and reduce exercise-induced oxidative stress | 430–700+ mg CF ~2 h pre-exercise or 200–1000 mg/day for 2 weeks–3 months | Weak/Low | Weak/Low |
| Coenzyme Q10 | Supports ETC, scavenges ROS, regenerates vitamins E & C | Reduces oxidative stress, limited performance impact | 30–600 mg/day for 2–12 weeks | Weak/Low | Moderate |
| Curcumin | Modulates NF-κB, direct antioxidant, anti-inflammatory | Reduces pain, inflammation, may improve performance short-term | 180–2000 mg/day for 3 days–3 months | Weak/Low | Moderate |
| Creatine monohydrate | Possible ROS scavenging, anti-inflammatory cytokine effects | Performance enhancement with potential antioxidant/anti-inflammatory support | 20 /day loading or 0.1 g/kg/day for 5–10 weeks | High | Weak–moderate |
| Fucoxanthin | Carotenoid-based ROS scavenger, anti-inflammatory | Improves aerobic capacity, reduces inflammation; cognitive support | 2–21 mg/day for 2–16 weeks | Weak/Low | Weak/Low |
| Glutathione/NAC | Boosts glutathione synthesis; NAC supports redox balance | Increases glutathione, supports aerobic performance in deficient individuals | Glutathione: 200–1000 mg/day, NAC: 1.2–2.0 g/day | Weak-to-moderate | Weak-to-moderate |
| Green Tea Catechins (EGCG) | Potent flavonols; antioxidant; Nrf2 activation; supports redox enzymes | Improves antioxidant capacity, modest fat oxidation, small soreness reductions | 400–800 mg/day catechins or 2–3 cups tea/day for 7–14 days; avoid acute high-dose pre-exercise | Weak-to-moderate | Weak-to-moderate |
| Lutein & Zeaxanthin | Scavenge ROS, support vision/cognition, concentrated in retina | Improves visual acuity, reaction time, possible muscular benefits | 10–34 mg/day for 8–12 weeks | Weak/Low | Weak/Low |
| Omega-3 fatty acids | Reduces ROS via mitochondrial & Nrf2 mechanisms | Reduces inflammation and oxidative stress, supports recovery | 1000–6000 mg/day EPA + DHA for 6–12 weeks | Moderate | Moderate–High |
| Pomegranate polyphenols | Ellagitannins/polyphenols; antioxidant and anti-inflammatory | Improves recovery from eccentric exercise; reduces soreness; beneficial hemodynamics | 500–1000 mL/day juice or extracts providing 500–1000 mg polyphenols for 3–7 days pre-/post-exercise | Weak-to-moderate | Moderate |
| Pycnogenol | Neutralizes ROS, enhances endogenous antioxidants, NOS modulation | Improves endurance, cognitive function, recovery | 60–200 mg/day for 4–12 weeks | Weak/Low | Weak/Low |
| Quercetin | antioxidant/free-radical scavenging; activation/upregulation of Nrf2 antioxidant pathway; modulation of AMPK–SIRT1–PGC-1α signaling promoting mitochondrial biogenesis; anti-inflammatory effects | Reduces soreness, supports mitochondrial health, performance potential | ~500–1000 mg/day (commonly 500 mg twice daily or 1000 mg/day); acute (single-dose or 7 days) to subacute (1–4 weeks) and some chronic trials up to 8 weeks | Weak-to-moderate | Weak-to-moderate |
| Resveratrol | Activates SIRT1, AMPK, Nrf2, affects mitochondrial function | May improve endurance and mitochondrial function, but could blunt training | 250–480 mg/day for 4–8 weeks | Weak/Low | Weak/Low |
| Selenium | Cofactor for GPx, thioredoxin reductase, reduces IL-6 | Supports antioxidant enzyme function, reduces inflammation | 50–250 μg/day for 3–42 weeks | Weak/Low | Weak/Low |
| Spirulina | Increases GSH, SOD, vitamin C, anti-inflammatory | Improves antioxidant status, may reduce oxidative stress | 500–10,000 mg/day for 15–60 days | Weak/Low | Weak/Low |
| Sulforaphane | Activates Nrf2, indirect antioxidant, anti-inflammatory | Enhances recovery, reduces soreness, does not blunt adaptation | 20–40 mg/day for 2–4 weeks | Weak/Low | Weak/Low |
| Tart Cherry | Anthocyanins/polyphenols reduce DOMS, inflammation | Reduces DOMS, supports sleep and inflammation control | 480 mg powder or 60–90 mL juice for 7–14 days | Moderate-to-high | Moderate |
| Urolithin A | Enhances mitophagy and mitochondrial efficiency; reduces inflammation | Improves muscular endurance, fat oxidation, VO2max response, reduces muscle damage | 500–1000 mg/day for 4–8 weeks | Weak/Low | Weak-to-moderate |
| Vitamins E & C | Direct scavengers, regenerate each other, mixed effects | Antioxidant defense, may reduce fatigue; may blunt training at high doses | Vit E: 400–800 IU/day, Vit C: 1000 mg/day | Weak/Low | Moderate–High |
| Zinc | Cofactor for Cu/Zn-SOD, Nrf2 activation, membrane stabilizer | Enhances SOD activity, reduces oxidative/inflammatory biomarkers | 50–100 mg/day for 2–16 weeks | Weak/Low | Weak/Low |
| Compound | Purported mechanism of action | Evidence strength |
|---|---|---|
| Beta-carotene | Scavenges singlet oxygen; precursor to vitamin A | Limited/preclinical |
| Melatonin | Regulates sleep; indirect antioxidant effects | Limited/preclinical |
| Lycopene | Quenches singlet oxygen; lipid peroxidation protection | Limited/preclinical |
| Carnosine | Buffers pH, some antioxidant properties | Limited/preclinical |
| Hesperidin | Citrus flavonoid; modulates antioxidant enzymes | Limited/preclinical |
| Silymarin (Milk thistle) | Flavonoid complex with hepatoprotective, antioxidant action | Limited/preclinical |
| Pterostilbene | Resveratrol analog; SIRT1 activator | Limited/preclinical |
| Gamma oryzanol | Plant sterol; antioxidant and anabolic claims | Limited/preclinical |
| Spinach thylakoid extract | Contains chloroplast-bound antioxidant enzymes (SOD, GPx) | Limited/human |
| GPLC (Glycine propionyl-L-carnitine) | Enhances nitric oxide; reduces lipid peroxidation (MDA) | Limited/human |
| SkQ (Mitochondria-targeted antioxidant) | Plastoquinone derivatives target mitochondrial ROS | Preclinical |
| Ketones | Promotes ketolysis; lowers oxidative stress markers (e.g. MDA) | Preclinical |
| Theaflavins | Polyphenols in black tea; scavenge ROS and modulate inflammatory pathways | Limited/preclinical |
| Apigenin | Flavonoid; Nrf2 activator, anti-inflammatory and neuroprotective | Preclinical |
| Melatonin | Endogenous antioxidant and sleep regulator | Limited/human |
| Lycopene | Carotenoid; quenches singlet oxygen | Limited/human |
| Olive fruit water (Hydroxytyrosol) | Phenolic compound; powerful free radical scavenger | Limited/human |
| PQQ (Pyrroloquinoline quinone) | Redox cofactor; supports mitochondrial biogenesis | Preclinical |
| Pterostilbene | Resveratrol analog; higher bioavailability; activates SIRT1, Nrf2 | Limited/preclinical |
| Sulforaphane | Isothiocyanate; activates Nrf2, enhances endogenous defenses | Limited/human |
| Tyrosol | Phenolic from olive oil; antioxidant and anti-inflammatory | Preclinical |
| Watercress | Rich in isothiocyanates and polyphenols; indirect antioxidant | Limited/human |
| Ergothioneine | Sulfur-containing amino acid derivative; cytoprotective antioxidant | Preclinical |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsExercise and Physiological Responses · Muscle metabolism and nutrition · Vitamin C and Antioxidants Research
Introduction
1
Athletes and human performance professionals (e.g. strength and conditioning coaches, sports dietitians, and tactical facilitators) frequently seek strategies to gain a competitive edge. These individuals often turn to dietary supplements to enhance exercise, athletic, or occupational performance and recovery. Accordingly, dietary or exogenous antioxidants have been proposed as ergogenic aids because of their ability to neutralize free radicals and exhibit anti-inflammatory properties [1–4].
Oxygen consumption increases during exercise [5] and contributes to the production of free radicals, such as reactive oxygen species (ROS) and reactive nitrogen species (RNS), whereas prolonged and/or heavy exercise training acutely augments this production [6]. Exercise-induced oxidative stress (EIOS) appears to exhibit both positive and negative effects, with moderate increases in ROS/RNS levels promoting favorable physiological adaptations, while excessive levels can damage cells and contribute to the development or progression of fatigue and/or various disease states (Figure 1) [7].
Exercise-induced oxidative stress and the hormetic response. Conceptual model illustrating the dose–response relationship between the magnitude of exercise-induced stress and its physiological effects, with a specific focus on redox balance and adaptation. The x-axis represents increasing magnitude of exercise or stress exposure, while the y-axis reflects the net biological effect, ranging from null or negative to beneficial. At very low levels of stress (e.g. sedentary behavior or minimal physical activity), insufficient reactive oxygen species (ROS) signaling results in limited antioxidant defense activation and minimal adaptive stimulus. Moderate increases in exercise stress fall within the hormetic zone, characterized by transient elevations in ROS that stimulate endogenous antioxidant defenses, enhance cellular repair mechanisms, and promote resistance to oxidative stress, thereby supporting favorable physiological adaptation. In contrast, excessive or prolonged stress exposure – such as high training loads, overreaching or overtraining, inadequate recovery, poor sleep or nutrition, or adverse environmental conditions (e.g. altitude or air pollution) – leads to excessive ROS generation, overwhelmed antioxidant capacity, impaired repair processes, and diminished adaptive responses. Collectively, the figure highlights the nonlinear, inverted-U relationship between exercise stress and biological outcomes, emphasizing the importance of appropriate training loads and recovery to maximize adaptation while minimizing maladaptation. Abbreviation: ROS, reactive oxygen species.
Exogenous antioxidants can influence EIOS, and some evidence suggests that they may attenuate the ROS-mediated pathways necessary for training adaptations [7]. Current evidence supports supplementation, primarily when correcting an insufficiency or deficiency, meeting dietary needs that cannot be met through food, or during periods of elevated stress, such as overreaching, back-to-back competitions, or in austere environments (e.g. smog and altitude) [1,4,8–10]. One’s age, sex, training status, dietary habits, and genetic polymorphisms, as well as factors such as redox biology and exercise type, may influence individual responses to supplementation, making the selection of an appropriate dietary antioxidant challenging [1,11]. Therefore, decisions regarding the use of exogenous antioxidants must be carefully considered.
This document presents the International Society of Sports Nutrition’s (ISSN) position on the effects of dietary antioxidants on exercise and sports performance, with a focus on EIOS and training adaptations, as well as recovery and performance parameters. The key objectives of this ISSN position stand are to (1) provide an overview of the mechanisms of dietary antioxidants, (2) discuss the scientific literature regarding dietary antioxidants and exercise/sports performance and recovery, and (3) discuss whether resistance-trained and endurance athletes should consider supplementing with antioxidants. Based on the available evidence, the dietary antioxidants discussed in this position stand are rated by the authors as having weak, moderate, or high levels of support for improving exercise- and sport-related outcomes.
Methods
2
ISSN position stands are invited papers that the ISSN Editors and Research Committee identify as being of interest to our readers who need position stands to guide evidence-based practice. Editors and/or the Research Committee identify a lead author or team of authors to perform a comprehensive literature review. The draft is then sent to leading scholars for review and comment. The paper is then revised into a consensus statement, reviewed, and approved by the Research Committee and Editors as the official position of the ISSN. This paper also provides a classification of the included dietary antioxidants, ranking them according to the strength of available evidence regarding their effects on exercise performance and antioxidant-related outcomes. Consistent with the 2018 ISSN position stand on research and recommendations [12], the primary writing team classified dietary antioxidants across both the performance and antioxidant domains using the following evidence-based criteria:
- (1)High – Strong evidence to support efficacy and apparently safe: Supplements backed by solid theory and supported by most well-controlled studies in relevant populations, using proper dosing regimens, demonstrating both effectiveness and safety.
- (2)Moderate – Limited or mixed evidence to support efficacy: Supplements with a solid scientific basis but inconsistent results in the existing research. These supplements need more high-quality studies to clarify their effects; however, there is currently no evidence indicating that they are unsafe or harmful when used properly.
- (3)Low – Little to no evidence to support efficacy and/or safety: Supplements that generally lack a solid scientific basis and/or for which the available evidence consistently shows no effectiveness. Supplements with documented safety issues or potential risks are also included in this category.
Notably, these classifications represent a continuum of scientific evidence rather than rigid categories. Accordingly, some supplements may fall between classifications (e.g. low-to-moderate or moderate-to-high), depending on the consistency, quality, and relevance of the available data.
What is an antioxidant?
3
An antioxidant refers to any substance, whether endogenous or exogenous, that reduces or prevents oxidation, defined as the loss of electrons or the addition of oxygen to lipids, carbohydrates, proteins, and deoxyribonucleic acid (DNA) [13]. The antioxidant defense system comprises endogenous enzymatic (i.e. superoxide dismutase [SOD], catalase [CAT], and glutathione peroxidase [GPx]) and non-enzymatic (e.g. glutathione [GSH], uric acid, and coenzyme Q10 [CoQ10]) antioxidants, in addition to exogenous/dietary antioxidants from food and supplemental sources (e.g. tart cherry, blueberry, and vitamin E). To be included in the academic definition of an antioxidant, the compound must exhibit antioxidant activity.
Our definition differs from the FDA 21 Code of Federal Regulations (CFR) § 101.54(g), which permits the “antioxidant” label only for nutrients that (1) demonstrate, after absorption, the ability to inactivate free radicals or prevent free radical-initiated reactions; (2) have an established reference daily intake (RDI); (3) meet defined minimum content levels (e.g. ≥20% of the RDI per serving for a “high” claim); and (4) are specifically identified on the label. According to these requirements, only vitamins C, E, and A (β-carotene), selenium, zinc, copper, and manganese are recognized by the FDA as dietary antioxidants. Other bioactive compounds, such as creatine, CoQ10, N-acetylcysteine, resveratrol, and quercetin, exhibit direct or indirect antioxidant effects but lack an RDI and thus are not FDA-recognized; however, these compounds are included in this analysis.
Mechanisms of action
4
Antioxidants can be derived from a wide variety of sources to mitigate oxidative damage to cellular components [14]. Antioxidants are considered reducing agents that neutralize or scavenge pro-oxidants (i.e. ROS, RNS, and free radicals) by several mechanisms, such as electron donation or quenching singlet oxygen. Within the context of health and disease states, antioxidants have the following mechanisms of action: (1) act as agents to catalytically eliminate ROS (i.e. SOD enzymes); (2) act as agents that minimize the availability of potent catalysts, such as iron ions, copper ions, heme, and heme-containing proteins, thus preventing or reducing the opportunity for oxidative damage to these metal ions; (3) act as singlet oxygen (¹O₂) quenchers, such as carotenoids, which can remove excess energy from singlet oxygen and return this molecule to normal oxygen; (4) act as “sacrificial agents,” which preferentially interact with ROS, halting their ability to damage “more important” biomolecules; and (5) undergo structural sequestration or compartmentalization, wherein the physical containment or breakdown of damaged cellular components prevents further propagation of oxidative injury [15]. In addition to direct mechanisms, many dietary and supplemental compounds function as indirect antioxidants by activating protective pathways, such as the Keap1/Nrf2/ARE system. These pathways can enhance phase II detoxification and the activity of antioxidant enzymes that remain active during redox reactions [16,17]. Taken together, these are the mechanisms of action by which antioxidants mitigate oxidative damage during exercise or sports.
Redox balance and oxidative stress
5
To understand the application of dietary antioxidants, it is essential to understand the concepts of redox balance, the distinction between oxidative eustress and stress, and EIOS. The following sections provide a brief review of these concepts. For further detail, see reviews by Helmut Sies [18–20] and Scott Powers [7,21].
Redox balance and oxidative stress
5.1
Helmut Sies defined “oxidative stress” as a disturbance in the balance of pro-oxidants and antioxidants, favoring pro-oxidants [18]. Since then, Sies’s definition has been adapted to be more comprehensive – “an imbalance between oxidants and antioxidants in favor of the oxidants, leading to a disruption of redox signaling and control, and/or molecular damage [7,19,22].” It is essential to appreciate this definition, given the complex cellular redox dynamics that occur in vivo. As the fields of redox biology and medicine have advanced, our understanding of oxidative stress and redox balance has also evolved. It is proposed that a steady-state redox setpoint exists, and any deviation from this setpoint results in the redox balance being in a state of stress [19]. Sies and colleagues [20] recognized that there are both states of oxidative stress (technically oxidative distress) and oxidative eustress, with the latter referring to an essential aspect of redox control and physiological redox signaling that supports health and adaptation. An important nuance here is the difference between oxidative stress and oxidative damage. Oxidative stress can be reversible, resulting in temporary redox imbalance or perturbed signaling, whereas oxidative damage is typically irreversible, leading to downstream structural alterations in biomolecules [23]. This distinction is essential for understanding biomarker data in exercise and recovery, as studies may report that biomarkers indicate oxidative damage rather than temporary oxidative stress.
Oxidative stress and eustress: ROS, RNS, and free radicals
5.2
Free radicals are molecules with unpaired electrons that are highly reactive in chemical reactions with other molecules [20]. These radicals can be categorized depending on whether they are derived from oxygen or nitrogen (i.e. ROS or RNS). The term “ROS” encompasses the following: singlet oxygen (¹O₂), superoxide (O_2_^·–^; the parent ROS molecule), hydrogen peroxide (H_2_O_2_; formed from the dismutation of O_2_^·–^), the hydroxyl radical (·OH, formed from reactions involving H_2_O_2_ and O_2_^·–^), and peroxyl radicals (ROO·). Notably, ·OH poses a high risk of oxidative damage due to its high reactivity. In terms of RNS, nitric oxide (NO) is considered the parent RNS and can react with O_2_^·–^ to form peroxynitrite (ONOO^−^), which is a reaction that can occur three times faster than the dismutation of O_2_^·–^, suggesting that the formation of ONOO^–^ is highly likely to occur when O_2_^·–^ is present. Consequently, the formation of ONOO^–^ reduces the amount of available NO. Additionally, because ONOO^−^ is a strong oxidizing agent, it can damage thiol groups and nitrate cellular proteins. Ultimately, these ROS/RNS can react with other radicals or cellular components, resulting in oxidative damage. Whether an individual can resolve an oxidative challenge and return to redox balance determines whether the free radicals impart distress or eustress. A state of oxidative distress can negatively impact an individual's health and performance if it remains chronically elevated [7].
The paradoxical nature of EIOS
5.3
Originally, exercise-induced ROS/RNS were thought to be purely detrimental to muscle function and performance [7,21,24]. However, the hormesis model describes a biphasic response in which low to moderate exposure to ROS/RNS stimulates the adaptive upregulation of endogenous antioxidants, thereby improving cellular function. In contrast, chronic or excessive exposure to ROS/RNS causes oxidative damage [25–28]. This bell-shaped response means that too few ROS fails to trigger beneficial adaptations. Moreover, excessive ROS/RNS, which can result from overtraining, inadequate recovery, or under-fueling, can lead to inflammation, muscle damage, and immunosuppression [28]. Although there is no known optimal dose of exercise-induced ROS/RNS for each individual, applying low to moderate training intensity with sufficient recovery generally promotes favorable redox signaling, while excessive stress impairs performance and health [7].
Oxidative stress, exercise, and chronic disease risk
5.4
Numerous diseases are thought to be linked to chronic exposure to oxidative stress, either as a cause or a result of the disease [29]. This phenomenon is interesting because exercise, which is linked to reduced susceptibility to chronic diseases and all-cause mortality [30,31], can also trigger the production of ROS/RNS. However, this highlights the concept of eustress, as exercise promotes favorable adaptations [32] and upregulates the endogenous antioxidant defense system [21,33,34]. Powers and colleagues [7] concluded that EIOS is a “friend” rather than a “foe,” given the substantial body of evidence that demonstrates exercise is linked to a reduction in all-cause mortality. In terms of supplementation with dietary antioxidants, athletes, including tactical athletes and master-level athletes or elderly individuals, may consider leveraging these compounds for health-related outcomes that extend beyond the exercise- and sport-specific contexts. However, dietary antioxidant intake may introduce competing goals. On the one hand, some antioxidants may interfere with exercise-induced adaptations through hormesis. On the other hand, daily antioxidant intake may help maintain redox balance and confer benefits. This raises questions about the considerations at play when working with “special populations,” that are influenced by multitude of factors affecting their health and physical performance (see Section 12 on Special Considerations).
Oxidative stress during exercise
6
Exercise and free radical production
6.1
Exercise causes the production of ROS/RNS. Early work by Dillard and colleagues [35] demonstrated that a 60-min cycle ergometer exercise bout at 50% V̇O_2_max resulted in increased expired pentane, an index of lipid peroxidation. Approximately one decade later, Lovlin et al. [36] and Gohil et al. [37] reported evidence that cycling exercise bouts triggered oxidative stress, as shown by increased concentrations of the lipid peroxidation marker thiobarbituric acid reactive substances (TBARS) and decreased GSH levels accompanied by increased oxidized GSH (GSSG). Several other research teams reported similar effects of acute aerobic exercise on the production of oxidative stress biomarkers [6,7,21,34,38,39]. Then, reports on the impact of anaerobic exercise emerged, as seen in Marzatico et al. [40], who demonstrated that six sets of 150-m sprints increased blood malondialdehyde (MDA) concentrations, another marker of lipid peroxidation, in sprint-trained athletes but not in marathon runners. McBride and colleagues [41] found that performing three sets of eight resistance exercises at a 10-repetition maximum load resulted in increased blood MDA immediately post-exercise. Importantly, these studies [35–37,40,41], among others [42–44], evaluated blood oxidative stress biomarkers, which is the most common method used to analyze EIOS. Other reports have also demonstrated that resistance exercise training can increase oxidative stress biomarkers via muscle biopsies, saliva, and urine [45–47].
Sources of free radical production during exercise
6.2
The discovery of EIOS has led researchers to identify possible sites of ROS/RNS generation within and around skeletal muscle. Interestingly, while it was initially believed that mitochondria were the primary source of ROS generation, it has become increasingly apparent that this is not the case. Boveris and colleagues [48] demonstrated that approximately 2%–5% of the molecular oxygen consumed by mitochondria produces O_2_^·–^, which was thought to increase only with the metabolic demands of exercise (i.e. increased oxidative phosphorylation in contracting skeletal muscle). However, several other sites of ROS/RNS generation contribute more substantially to EIOS, which include phospholipase A_2_ (PLA_2_) and NADPH oxidases [49]. It has been postulated that PLA_2_ contributes to ROS/RNS production through several mechanisms, such as (1) the activation of NADPH oxidases via signaling intermediates that facilitate enzyme assembly, (2) the release of arachidonic acid, which, when metabolized by enzymes, leads to the formation of ROS and RNS byproducts (e.g. O_2_^·–^, H_2_O_2_, ONOO^−^), and (3) the stimulation of mitochondrial ROS production via a calcium (Ca^2+^)-dependent isoform of PLA_2_ (i.e. iPLA_2_ or cPLA_2_). During exercise, when the mitochondrial membrane is disrupted by PLA_2_, leakage of electrons from complexes I and III of the electron transport chain can generate ROS [50]. Powers et al. [7] concluded that more research is warranted to confirm this postulation.
In terms of NADPH oxidase, there are two isoforms within skeletal muscle: NOX1, NOX2, and NOX4 [51]. The NOX2 isoform is found in the sarcolemma and T-tubule, while the NOX4 is found in the sarcoplasmic reticulum and mitochondria [52]. Importantly, NOX2 appears to be the primary site of NADPH-mediated ROS production in exercising muscle, as it is activated by agonists such as cytokines and angiotensin II, as well as by contractile or mechanical stress. Upon mechanical stress or contraction of skeletal muscle, intracellular signaling (e.g. increased calcium or ROS) can promote the translocation of cytosolic subunits to the myocyte membrane, forming an active complex in which NADPH transfers electrons to oxygen molecules to generate O_2_^·–^. Given its mechano- and chemo-sensitivity, NOX2-mediated ROS production increases progressively with the onset and intensity of exercise. Importantly, NOX isoforms have been identified as the primary source of O_2_^·–^ production during rest and exercise [7].
Xanthine oxidation is another implicated source of ROS production in skeletal muscle. The enzyme xanthine oxidase (XO) is located within the capillary endothelial cells that surround skeletal muscle, and muscle contraction can trigger a reaction that activates XO to generate O_2_^·–^, which, when converted to H_2_O_2_ by extracellular SOD, can cross the sarcolemma to enter the myocyte and cause further pro-oxidative effects. XO-mediated O_2_^·–^ production can also interact with NO, produced by skeletal muscle-expressed isoforms of nitric oxide synthase (NOS; nNOS and eNOS) to form ONOO^–^ [53]. The proximity of key sites of ROS/RNS production to skeletal muscle aligns with evidence showing that localized EIOS serves as a proximal signal driving adaptive responses [54].
Effects of exercise-induced oxidative stress on performance parameters
7
Previous reports suggest a connection between oxidative stress, fatigue, and muscle damage, which can impact exercise performance [55–57]. Interestingly, a link exists between increased oxidative stress biomarkers and performance or recovery. For example, Stein and colleagues [58] reported that US Special Forces members who were not selected in the SFAS course had higher levels of metabolites associated with increased oxidative stress. These levels could be due to factors such as poor nutrition, non-functional overreaching or overtraining, or incomplete recovery from prior physical exertion. Furthermore, those selected individuals demonstrated resistance to oxidative stress and exhibited greater physical performance than non-selected individuals [58]. While this study is specific to military selection [58], the concept has been described elsewhere [7,34,59–63]. Additionally, the inflammatory response that occurs during and after exercise mobilizes and activates the immune system to respond to muscle damage. This immune response is further stimulated by stress hormones (i.e. catecholamines, growth hormone, and cortisol), which are released in response to increasing metabolic demands during exercise [64,65]. These physiological responses to exercise can influence subsequent training or competition bouts.
Force production, muscle fatigue, and recovery
7.1
Force production, defined as the capacity of an individual muscle or group of muscles to generate contractile tension, is a critical element of exercise and sports performance. As such, Reid et al. [66] developed a theoretical model describing the association between skeletal muscle redox homeostasis and force production. In short, Reid’s model resembles the exercise-induced hormesis model [26,27,67], suggesting that, under normal physiological conditions, ROS/RNS production plays a crucial role in adapting to exercise [6]. In contrast, excessive production can lead to oxidative damage to the structural integrity of skeletal muscle proteins involved in muscle contraction [39,68]. Specifically, ROS/RNS can influence skeletal muscle contraction through (1) the modulation of Ca^2+^ handling/excitation‒contraction coupling, (2) direct oxidation of contractile proteins, (3) neuromuscular transmission (i.e. impairment of acetylcholine release and receptor sensitivity at the neuromuscular junction site), and (4) nitrosative stress (i.e. excessive NO production leading to ONOO^−^, which in turn can cause protein nitration). Ultimately, these effects can lead to reduced cross-bridge formation, impaired muscle fiber recruitment, and a diminished ability to sustain muscle contractions, thereby inhibiting force output and muscular endurance. Another important aspect of this cascade of events is that ROS/RNS production can promote inflammation via the nuclear factor kappa B (NF-κB) redox-sensitive transcription factor. For example, H_2_O_2_ or ONOO^–^ can phosphorylate and degrade the inhibitor of κB (IκB), thereby releasing NF-κB, which can translocate to the nucleus and activate the transcription of pro-inflammatory cytokines. These cytokines can further reduce force output through similar mechanisms (i.e. impaired excitation‒contraction coupling) or proteolytic signaling, promoting greater fatigue and slower recovery. Under conditions of high-intensity, prolonged, or fatiguing exercise, ROS/RNS production may overwhelm the endogenous antioxidant capacity, leading to oxidative stress, oxidative damage, muscle fatigue and damage, and even delayed onset muscle soreness (DOMS) [69,70]. However, importantly, at low-to-moderate levels of ROS/RNS production, these radicals can facilitate Ca^2+^ release (short-term force production influence) and trigger signaling pathways for adaptation for a more long-term influence on force production (i.e. upregulation of peroxisome proliferator-activated receptor-γ coactivator 1-α [PGC-1α] for mitochondrial biogenesis).
Current evidence suggests that EIOS is likely to affect recovery between training sessions or competitions, primarily due to its association with fatigue, muscle damage, and impaired immune function [49,71]. The post-exercise recovery phase is characterized by oxidative stress stemming from the infiltration and activation of immune cells, such as neutrophils and macrophages, into damaged skeletal muscle tissue [34,72,73]. While these immune cells play a key role in the normal inflammatory and tissue repair response, they may also produce excessive ROS via NOX2 activation. In addition, ROS levels can trigger temporary, reversible, and irreversible oxidative modifications in skeletal muscle, including S-nitrosylation, S-glutathionylation, and disulfide bond formation [74–76]. Specifically, S-glutathionylation of cysteine residues can lead to protein deactivation and inhibition of activity via extracellular vesicles [74]. Therefore, this kind of redox alteration after translation could play a significant role in skeletal muscle signaling during exercise and recovery [77,78]. One additional area that links EIOS to perceived recovery among athletes is DOMS, which often occurs after extreme or unaccustomed exercise [69]. It has been shown that ROS are involved in DOMS, with inflammatory agents in the muscle identified as the source of production [79]. Early studies revealed a connection between ROS, lipid peroxidation, and DOMS after a downhill running protocol was used [79,80]. The timing between ROS production and peak DOMS seems to depend on the context, with some studies showing that ROS levels increase after peak soreness, suggesting that they are out of sync [79]. Conversely, other evidence suggests that ROS production and inflammation may occur closer to the onset of DOMS, especially in untrained people performing unfamiliar eccentric exercise [79,81]. An individual's training history and exercise background probably influence the timing and intensity of oxidative and inflammatory responses after muscle damage [44,69,82–86]. Indeed, adaptations associated with chronic training may attenuate or delay oxidative stress and inflammatory signaling, potentially altering the temporal relationship between ROS production, neutrophil infiltration, and perceived muscle soreness compared with untrained individuals.
Numerous studies have shown that training leads to adaptations in the antioxidant system, thereby reducing oxidative stress. However, evidence suggests that intense exercise can trigger an increase in oxidative stress [87,88], which may contribute to the onset of overtraining and potentially increase oxidative stress [89–91]. Athletes who display overtraining syndrome may be in a state of chronic oxidative stress, which is likely to be indicated by elevated blood biomarkers or reduced plasma antioxidant capacity [92]. These findings suggest that oxidative stress is not only a contributor to overtraining but also a symptom of overtraining. The concept of using an antioxidant to accelerate recovery is intriguing because it could lead to improvements in athletic performance or an increase in the volume of training one can perform. A faster recovery could enable athletes to increase their training frequency or intensity, thereby enhancing their overall performance and competitive edge. However, it is also important to consider this in the context of chronic adaptation.
Muscular adaptation
7.2
It has been clearly demonstrated that the contraction of skeletal muscles will generate ROS/RNS and that low-to-moderate levels of ROS/RNS production can facilitate adaptation in the long term as a means to improve the muscle’s ability to handle future exposure to stressors (i.e. subsequent bouts of training or competition) [7,21]. The type, volume, duration, and intensity of the stressor can dictate the balance between beneficial and detrimental outcomes and whether it creates a state of oxidative distress (commonly referred to as oxidative stress) or eustress. In the context of oxidative eustress (i.e. a modest increase in ROS/RNS), the (1) activation of redox-sensitive transcription factors (i.e. NF-κB) [93], (2) promotion of mitochondrial biogenesis via PGC-1α [94], (3) enhancement of the endogenous antioxidant defense system (e.g. upregulation of endogenous antioxidants, such as catalase) [33,95], and (4) facilitation of muscle remodeling and hypertrophy via the modulation of mitogen-activated protein kinase (MAPK) signaling (i.e. p38 and ERK1/2) [94,96] are all potential ways in which skeletal muscle can adapt to better handle stressors in the future encounter (i.e. subsequent training/competition). Furthermore, preserving redox homeostasis, which involves maintaining ROS/RNS at non-harmful levels while still allowing them to function as signaling molecules, is crucial for a cell to mitigate its toxic effects [97]. As such, mature skeletal muscle cells, along with myogenic stem and progenitor cells, possess antioxidant systems that allow them to adapt to changes in the redox environment [98]. Oxidative stress can also alter the contractility of striated muscle cells under both physiological and pathological conditions, with modifications to the titin protein, directly and indirectly, affecting muscle elasticity and stiffness through changes in protein kinase signaling pathways and protease activation [98]. In contrast, when the stressor creates a state of oxidative stress characterized by excessive ROS/RNS levels, muscle contractile function, inhibition of muscle protein synthesis, mitochondrial damage, and suppression of satellite cell activity are likely impaired, which can ultimately hinder muscle repair and growth, as well as exercise performance [97,99,100].
Endurance performance
7.3
High-intensity or prolonged endurance training increases free radical production and endogenous antioxidant defense. Previous reports have shown that endurance training increases SOD1 and SOD2 activity in skeletal muscle by 20%–112%, while increasing GPx activity by 20%– 177% [6,34,101]. Work by Berzosa et al. [102] and Georgakouli et al. [103] demonstrated that a 30-min submaximal cycle ergometer exercise bout at 70% maximum workload and 50%–60% of heart rate reserve increased plasma total antioxidant capacity (TAC). The upregulation of endogenous antioxidant capacity in response to elevated oxidative stress is widely considered a key mechanism underlying endurance-related adaptations (i.e. mitochondrial biogenesis). However, prolonged or high-volume endurance training may have drawbacks, including temporary decreases in antioxidant capacity [4]. These decreases affect not only endogenous antioxidant enzymes but also circulating dietary antioxidants, such as vitamin E, during intense training periods, further impacting overall antioxidant levels [104]. This increased oxidative stress can lead to immune suppression, potentially impairing recovery and increasing the risk of illness and overtraining [59,92,105–108]. One proposed mechanism is that leukocyte-derived ROS and RNS, which are critical for immune defense, become dysregulated or suppressed with excessive training loads, potentially reducing the body’s ability to fight infections [109]. This effect has been demonstrated in athletes competing in ultra-endurance events, including ultramarathons and Ironman triathlons [110–112]. While concerns about cumulative oxidative damage persist, some researchers argue that current evidence does not definitively demonstrate that high-intensity endurance training causes lasting oxidative damage in well-adapted or trained athletes [113].
Takeaway summary: EIOS can have either beneficial or detrimental effects on force production, recovery, muscle adaptations, and endurance-related performance outcomes, depending on the level of ROS/RNS production from exercise or training bout. Nevertheless, it is recommended to consider the training goals (recovery versus adaptation) and phase of training or competition (i.e. in-season or off-season) concerning these outcome variables to ensure that (1) the level of EIOS is appropriate to generate a favorable impact on the individual and their performance and/or health, and (2) any interventions with dietary antioxidants are not blunting key adaptations and normal physiological responses necessary for improvement through training.
How do antioxidants combat oxidative damage and inflammation
8
Three main categories of antioxidants meet the definition of substances that can slow the oxidation of proteins, lipids, carbohydrates, and DNA. Two key endogenous antioxidant enzymes are SOD and catalase. SOD converts O_2_^·–^ (a highly reactive free radical) to H_2_O_2_ (a less reactive free radical), which can then be further broken down by catalase to yield water (H_2_O) and oxygen (O_2_). This reaction prevents the formation of ·OH, which is considered the most reactive free radical. Second, non-enzymatic antioxidants, such as uric acid and GSH, can scavenge ROS/RNS and donate electrons to highly reactive free radicals, converting them to more stable, less reactive species. For instance, uric acid can react with ·OH to form allantoin and H_2_O or neutralize ONOO^–^ to prevent protein nitration. Similarly, GSH can donate electrons to ·OH or H_2_O_2_, among other free radicals, to convert them to more stable and less reactive species. Finally, functional enzymes (i.e. proteases, lipases, etc.) not only modulate the general metabolism of macromolecules but also exhibit indirect roles in mitigating oxidative damage via multiple mechanisms (i.e. autophagy, blunting misfolded protein translation, mobilizing stored lipids, reducing lipid accumulation, etc.). Dietary antioxidants are generally considered an extension of the non-enzymatic antioxidants mentioned above (Table 1).
The earliest report assessing the impact of a dietary antioxidant, 1200 IU/day of vitamin E for two weeks, on exercise performance was conducted by Dillard and colleagues [35], who found that vitamin E blunted the increase in pentane. This was thought to occur when vitamin E donated a hydrogen atom to lipid ROO·, thereby interrupting lipid peroxidation. Other antioxidants, such as vitamin C, can help “recycle” vitamin E by reducing it back to α-tocopherol, allowing it to continue serving as an antioxidant [114–116]. Vitamin C can also directly neutralize ROS, such as O₂·^–^, H₂O₂, ·OH, and ¹O₂, by donating electrons to render the ROS less reactive [117]. Other antioxidants, such as N-acetylcysteine (NAC) and astaxanthin (AST), can also directly scavenge ROS/RNS and other free radicals while also modulating the redox-sensitive NF-κB and nuclear factor erythroid 2-related factor 2 (Nrf2) pathways [118–124]. Importantly, antioxidants, such as NAC, can work as precursors to other endogenous antioxidants [125]. For example, NAC can provide cysteine, which is the rate-limiting amino acid for GSH synthesis [126]. Another key factor in how dietary antioxidants exert their free-radical neutralizing effects is their molecular structure and solubility. Lipid-soluble antioxidants, such as AST or vitamin E, are especially effective at intercepting radicals within the phospholipid bilayer [127,128]. In contrast, water-soluble antioxidants (e.g. NAC) predominantly operate in aqueous compartments (i.e. blood plasma and the cytosol) to neutralize free radicals [125]. In general, dietary antioxidants can serve as (1) direct free radical scavengers, (2) precursors to other non-enzymatic antioxidants, and (3) modulators of redox-sensitive pathways, which can reduce pro-inflammatory cytokine production and increase endogenous antioxidant enzyme expression [129].
Currently, data suggest that acute dietary antioxidant supplementation can be beneficial if the aim is to mitigate muscular fatigue and recover quickly; however, if the goal is to improve performance or training adaptations, depending on the specific nutrient in question (i.e. vitamin E or vitamin C versus AST), dietary antioxidant supplementation may not be warranted, especially given that chronic, high-dose antioxidant supplementation has been shown to impair performance and exercise adaptations [1,2,4,34,130]. Previous work has demonstrated that there are variations in individual EIOS responses to training, which may offer valuable insights into why some studies report differing results with exercise training and antioxidant treatment [10,131–134]. For instance, Margaritelis and colleagues [131] found that acute, high-intensity eccentric, muscle-damaging exercise led to minimal or no oxidative stress, or even reductive stress, for more than one out of every three individuals among 98 young males. These observations suggest that a person's redox status should be evaluated beforehand when examining the effects of antioxidant intake on EIOS. Margaritelis et al. [132] proposed that if an individual is deficient in a specific nutrient, supplementation with that specific antioxidant may be warranted and could support performance-related outcomes. However, concluding that antioxidant supplementation is necessary solely based on oxidative stress is not ideal, as interindividual differences likely influence how an individual responds to EIOS and subsequent antioxidant intake [135]. There are likely many reasons (e.g. health and/or performance) for an individual to supplement with an antioxidant. Nevertheless, individuals must consider whether they may be deficient in a dietary antioxidant and the strength of the evidence supporting its use.
Common dietary sources of antioxidants: a food-first framework
9
Exogenous antioxidants include vitamins, minerals, flavonoids, anthocyanins, tocopherols, polyphenols, and carotenoids, which can be obtained from whole foods and/or dietary supplements [136,137]. In general, athletes can obtain optimal amounts of exogenous antioxidants through a well-balanced diet. Nevertheless, athletes and sports performance professionals need to understand which common whole food sources are rich in dietary antioxidants (see Table 1) to (1) ensure optimal intake through diet and (2) identify potential gaps due to personal preferences, dietary restrictions, or food allergies. In such cases, targeted supplementation may be warranted to compensate for insufficient intake from a food-first approach.
The various foods containing these antioxidant compounds (i.e. flavonoids, polyphenols, tocopherols, and carotenoids) can leverage ROS/RNS scavenging abilities largely due to the molecular structure and classifications (i.e. water- or lipid-soluble) of these compounds. For instance, flavonoids – found in fruits, herbs, cereals, nuts, stems, vegetables, seeds, and flowers – are characterized by a benzopyran backbone consisting of two aromatic rings (A and B) and a heterocyclic C ring [138]. This structure confers phenolic or polyphenolic properties that contribute to their antioxidant activity. For example, the hydroxyl (-OH) groups attached to the A, B, or C rings can donate a hydrogen atom or an electron to a free radical (i.e. ROO·), or the 3',4'-dihydroxy groups on the B ring or the 3-OH and 4-keto groups on the C-ring can allow for the chelation of redox-active metals, preventing Fenton reactions to reduce the potential generation of ·OH. Anthocyanins, a subclass of flavonoids, contain a flavylium ion that gives them red, blue, or purple pigmentation (depending on pH). These compounds have the same C6-C3-C6 skeleton as do other flavonoids; therefore, anthocyanins can also donate a hydrogen atom or an electron from their -OH groups to neutralize free radicals (i.e. O₂·^–^, ·OH, ROO·, (¹O₂) or chelate transition metals to prevent the formation of ·OH. Another important function of these compounds is indirect antioxidant effects through the activation of Nrf2 signaling, demonstrating that dietary antioxidants not only directly neutralize ROS/RNS but also support endogenous antioxidant defenses. Table 1 provides a general overview of each dietary antioxidant class and its mechanism of action.
Dietary supplementation: short- and long-term effects in exercise and sport
10
Observational studies have reported that approximately 50%–100% of athletes supplement with vitamins, antioxidants, or other substances to enhance performance and immune function [139–141]. A wide array of dietary antioxidant supplements are marketed for their ability to promote recovery and mitigate exercise-induced oxidative damage and inflammation. Identifying clinically relevant antioxidants can be challenging because of the large number of bioactive compounds with antioxidant activity. Although many of these compounds exhibit antioxidant activity in preclinical studies, they may lack robust clinical evidence to confirm their efficacy or have not been assigned an RDI by the FDA. As such, selecting an appropriate dietary antioxidant is often more nuanced than choosing other ergogenic aids, such as beta-alanine, which has a well-defined mechanism for enhancing high-intensity exercise performance by buffering hydrogen ions.
Several factors can influence an individual’s response to dietary antioxidant supplementation, including (1) training status (e.g. sedentary, recreationally active, resistance- or endurance-trained); (2) baseline endogenous antioxidant capacity and oxidative stress levels; (3) sex and gender differences; (4) age-related changes in redox homeostasis; (5) overall nutritional status and habitual dietary antioxidant intake; (6) the type, dosage, and timing of antioxidant administration; (7) the chemical form and bioavailability of the antioxidant compound; (8) the type, intensity, and duration of the exercise stimulus; and (9) environmental factors or air quality. Therefore, it is crucial for the athlete or the professional working with the athlete or individual to consider these factors in addition to the body of evidence supporting claims made about an antioxidant.
There are arguments for and against the use of dietary antioxidants among athletic and exercise-training populations. Further research warranted to establish whether the dietary antioxidant in question is beneficial or detrimental to training adaptation or performance, given the respective training or sport-specific setting [9]. Current evidence suggests that during periods of intensified training, such as planned overreaching phases or consecutive days of competition, antioxidant supplementation may offer benefits, particularly when the primary goal is to enhance short-term recovery [4]. For athletes undergoing such phases, reductions in subjective or objective markers of muscle soreness or improvements in recovery kinetics may lead to better preparedness for subsequent performance. However, for athletes outside these demanding conditions, it remains unclear whether acute improvements in recovery, attributed to reduced muscle damage, ultimately translate into meaningful performance gains. Table 2 summarizes all the dietary antioxidants discussed subsequently, along with the level of evidence supporting their inclusion in exercise and sport.
Key findings and strategies for dietary antioxidants in exercise and sports
11
Alpha-lipoic acid
11.1
Alpha-lipoic acid (ALA), also known as thioctic acid, serves as a cofactor for various multi-enzyme complexes (e.g. pyruvate dehydrogenase, α-ketoglutarate dehydrogenase, and branched-chain ketoacid dehydrogenase), which play a role in the oxidation of keto acids, energy production, and amino acid metabolism [142]. In terms of its antioxidant capabilities, ALA can recycle endogenous glutathione; scavenge hydroxyl and peroxide radicals, singlet oxygen, and hypochlorous acid; and form chelate complexes with metal ions [143,144]. To date, these antioxidant effects of ALA have primarily been shown in clinical human trials with Alzheimer’s disease [145] and diabetic patients [146–148], as well as overweight/obese adults [149]. Doses ranging from 100 to 1200 mg/day of ALA have been evaluated, with higher doses up to 1800 mg/day for up to 6 months. Nevertheless, only a handful of studies have examined the application of ALA for exercise and sports performance [150–152]. First, Fogarty et al. [152] reported that daily supplementation with 1000 mg/day of ALA for 14 days among 12 healthy males could selectively protect against DNA oxidation, as demonstrated by participants expressing (1) 5.4% shorter DNA tail length (indicating DNA fragmentation and damage within cells) and (2) lower (p < 0.05) plasma 8-OHdG (2.1%), lipid hydroperoxides (7.75%), protein carbonyls (70%), and hydrogen peroxide (0.4) levels following a muscle-damaging exercise protocol. Then, Morawin et al. [151] assessed the effects of ALA (i.e. 1200 mg/day of thiogamma for 10 days) on erythropoietin release following an exercise bout consisting of 90 min of running followed by 15 min of eccentric running at 65% V̇O_2_max at a −1% gradient among 16 healthy young adults. They found that ALA increased erythropoietin release following eccentric running exercise and decreased H_2_O_2_ and other markers of oxidation, but did not affect creatine kinase (CK). Finally, Isenmann and colleagues [150] assessed the effects of both single-dose acute (150 mg dose) and short-term (i.e. 2 × 150 mg/day for 6 days) ALA supplementation on muscle strength recovery and performance among 17 male resistance- and endurance-trained athletes in a randomized, placebo-controlled, crossover manner. A single dose of ALA did not affect 1-RM back squat strength or biomarkers of skeletal muscle damage, oxidative stress, or inflammation. However, the short-term ALA supplementation protocol resulted in participants maintaining their 1-RM back squat strength compared with placebo, which declined and was lower at the final time point. Collectively, reports on ALA suggest that one week of supplementation may reduce biomarkers of muscle damage and oxidation; however, more research and longer studies are needed before definitive recommendations can be made regarding supplementation strategies.
Ashwagandha
11.2
Ashwagandha (ASH; Withania somnifera) is an emerging adaptogenic compound that is considered a fundamental Ayurvedic remedy. ASH has garnered significant attention over the last decade for its potential anti-stress, antioxidant, anti-inflammatory, and immunomodulating properties [153]. ASH is supplemented primarily as a root extract and is known for mediating the neuroendocrine response (i.e. cortisol/testosterone) and various oxidative stress and inflammatory biomarkers, with the potential to impact muscle strength, power, cardiorespiratory fitness, and recovery [153,154]. Recently, ASH has gained increased attention for its antioxidant and anti-inflammatory properties, which may enhance physical performance and recovery [155,156]. ASH contains several bioactive compounds, including withanolides (e.g. withaferin A, withanolide D), alkaloids, sitoindosides, flavonoids, tannins, and polyphenols. It is thought to modulate cellular pathways, such as Nrf2 (promoting endogenous antioxidants such as SOD, CAT, and GPx) and NF-κB (reducing pro-inflammatory cytokines) [155–157]. These mechanisms may aid post-exercise recovery, and some studies have reported positive effects on strength and body composition [158,159].
Few studies have examined the effects of ASH on exercise and sports performance. Bonilla et al. [155] conducted a meta-analysis demonstrating ASH’s effectiveness in improving strength and power outcomes at doses of 240–600 mg/day for ≥6 weeks, though most data were obtained from untrained populations. Furthermore, Wankhede et al. [158] assessed the effect of 300 mg/day of ASH and found improvements in bench press and leg extension strength compared to the placebo group. In addition, the ASH group also experienced greater increases in arm muscle cross-sectional area and chest girth, as well as a greater reduction in body fat percentage [158]. Ziegenfuss et al. [160] found that 12 weeks of ASH supplementation at 500 mg/day, coupled with a resistance training program, resulted in greater squat and bench press strength than the placebo. Additionally, only the ASH group showed improvements in upper- and lower-body power and perceived recovery scores [160]. Finally, Raut et al. [161] reported increased muscular force production during handgrip, quadriceps, and back extensor exercises after 30 days of supplementation with increasing doses of 750–1250 mg/day of ASH in physically active, healthy individuals. Collectively, these findings demonstrate that ASH supplementation can improve performance and recovery; however, more research is warranted.
Astaxanthin
11.3
Astaxanthin, a lipid-soluble, red-orange-colored ketocarotenoid, has attracted attention for its potent antioxidant and anti-inflammatory properties [118–122,162–164]. The effective dosage and duration of AST supplementation (Haematococcus pluvialis microalgae) range from 4 to 12 mg/day for 4–12 weeks to favorably impact cardiometabolic health outcomes [119,165] and reduce inflammatory and oxidative stress biomarkers [166–182]. The unique structure of AST, which has two β-ionone rings connected by a polyene chain, allows it to cross the phospholipid membrane thickness and neutralize ROS/RNS both at the membrane periphery and inside the lipid bilayer [183,184], is naturally found in salmon, lobster, shrimp, and crab. This mechanism may increase fatty acid oxidation during exercise [185] and attenuate or limit EIOS, inflammation, and muscle damage, ultimately promoting quicker recovery [172,186–189]. Owing to its limited dietary availability in whole foods, supplementation with AST is needed for ergogenic benefits. AST is generally recognized as safe (GRAS), and the only notable side effect occurs when it is incompletely absorbed in the gut and is excreted in the stool, imparting a red color [128].
Several studies support the use of AST as potentially beneficial for athletes. For instance, Fleischmann et al. [188] and Sawaki and colleagues [190] found reduced postexercise blood lactate concentrations following supplementation protocols of 12 mg/day AST for 30 days and 6 mg/day AST for 4 weeks, suggesting improved metabolic recovery. Malmsten et al. [191] reported that the use of 4 mg/day AST over six months, when combined with exercise, increased the number of barbell squat repetitions by 55% in paramedic students. With respect to substrate utilization, Brown and colleagues [192] reported that 12 mg/day AST for 7 days increased fat oxidation (≈69%) and reduced respiratory exchange ratios (≈3%) in trained male cyclists. However, most studies have not demonstrated an effect of AST on substrate utilization [189,193,194]. AST has also shown promise in attenuating biomarkers of EIOS, muscle damage, and inflammation following intense exercise [168,172]. For example, Baralic et al. [168] reported increased antioxidant enzyme activity (≈17% –≈42%) following 90 days of 4 mg/day AST supplementation in soccer players, with a follow-up study finding that the increase in high-sensitivity C-reactive protein (CRP) levels with training was attenuated in the AST group [167]. Finally, Djordjevic et al. [172] demonstrated that elite soccer players who ingested 4 mg/day AST for 90 days displayed a blunted increase in postexercise blood creatine kinase levels compared to those receiving placebo (≈21% and ≈29%, respectively). Taken together, these findings demonstrate the ergogenic value and antioxidant and anti-inflammatory benefits of AST in the context of sport and exercise.
Studies using AST have shown either no adverse effects [187] or even a positive effect on performance and training adaptation [191,195]. For example, Barker and colleagues [187] reported that 12 mg/day AST for 4 weeks did not affect resistance training outcomes but reduced muscle soreness (≈57%; p = 0.01) and perceived muscle damage (≈60%; p = 0.02) 24 hours postexercise. This finding demonstrates the ability of ASTs to enhance recovery without impairing training adaptations. While this may seem minimal, one cannot overlook the importance of improving an individual’s perception of soreness or recovery, as this may lead to improved work output in training and competition [196].
Beetroot
11.4
Beetroot (Beta vulgaris) is a nitrate-dense root vegetable that also contains a range of bioactive compounds, including betalains, phenolic acids, flavonoids, ascorbic acid, and carotenoids. These constituents possess antioxidant and anti-inflammatory properties that may offer benefits in both clinical and athletic contexts [197–199]. However, the focus of beetroot supplementation has focused primarily on its nitrate content and NO-mediated effects; emerging evidence suggests that other phytochemicals, particularly betalains, may also contribute to performance and recovery. Betalains have demonstrated the ability to reduce reactive oxygen and nitrogen species (ROS/RNS), modulate inflammatory gene expression, and aid recovery from exercise-induced muscle damage [200]. A recent review even suggested that beetroot juice may outperform nitrate salts alone because of these additional compounds and their combined effects on NO production, mitochondrial function, and vascular health [201]. As such, beetroot supplementation has been explored as a strategy to mitigate oxidative stress, reduce inflammation, and enhance recovery [197,201].
Despite these promising mechanisms, evidence regarding the effects of beetroot on inflammation and oxidative stress is inconsistent, with a series of studies by Clifford et al. [202–205]. demonstrated no impact on markers of EIOS or inflammation following strenuous exercise protocols, including drop jumps, sprint tests, and marathon running; however, some noteworthy ergogenic and recovery effects were observed. First, Clifford et al. [203] examined the effects of single doses of beetroot juice (250 mL or 125 mL) versus placebo on recovery and inflammation after eccentric exercise in 30 recreationally active males. The 250 mL group showed faster recovery in countermovement jump performance at 48 and 72 hours postexercise and reported higher pressure pain thresholds. However, beetroot juice had no effect on markers of exercise-induced oxidative stress, inflammation, or muscle damage. Clifford et al. [204] found that 2 × 250 mL/day for 3 days of beetroot juice supplementation led to quicker recovery for the countermovement jump (7.6%) and relative strength index (13.8%) than did a placebo, with no effect on the biochemical markers among 20 male, team-sports players. Their third study [205] revealed that 3 days of supplementation with beetroot juice (containing ≈210 mg nitrate and ≈405 mg/GAE/L polyphenol content), sodium nitrate (containing ≈210 mg nitrate but no polyphenols), or placebo did not impact any biomarkers following eccentric drop jumps, but the beetroot juice group experienced an attenuation of their pressure pain threshold among 30 recreationally active males. Finally, Clifford and colleagues [202] conducted a marathon field study among 34 experienced runners who consumed either beetroot juice or a placebo during the 3 days post-marathon recovery phase and found no effect on any biomarkers of EIOS or inflammation. Interestingly, Kozłowska et al. [206] demonstrated that 4 weeks of 26 g/day beetroot juice supplementation increased GPx-1 and GPx-3 activity, elevated β-carotene, reduced malondialdehyde (MDA) and advanced oxidation protein products (AOPP), and improved V̇O_2_max. In addition, Daab et al. [207] demonstrated that 2 × 150 mL/day of beetroot juice supplementation for 7 days (3 days pre- and post-exercise) led to an attenuated decrease in countermovement jump performance and maximal voluntary contraction output during the recovery days post-exercise, while no effect was noted for CK, lactate dehydrogenase (LDH), and CRP. Finally, findings from Vilar et al. [208] reported among 32 ultra-endurance runners participating in a 107-km mountain race demonstrated no effect on EIOS or inflammatory biomarkers. Taken together, there appears to be a benefit in terms of recovery, as shown by the attenuated decrease in countermovement jump performance; however, there is little to no effect on EIOS or inflammation.
Similarly, recent systematic reviews have highlighted the inconsistency in the effects of beetroot supplementation on markers of inflammation and oxidation [209–211]. For example, Jones et al. [209] concluded that beetroot consistently improved functional recovery and soreness but did not influence systemic CRP, IL-6, IL-8, tumor necrosis factor-α (TNF-α), or lipid peroxidation products. Based on six studies that measured markers of inflammation, Rojano-Ortega et al. reported no significant differences between beetroot and placebo [210]. Additionally, beetroot supplementation did not attenuate oxidative stress compared with placebo in three of the four included studies [210]. Only one study reported significant changes in GPx-1 and MDA; however, the authors postulated that these changes were most likely reflective of training-related adaptations rather than direct antioxidant effects [206]. Beetroot (as nitrate-rich juice) appears to aid in functional recovery and some performance outcomes; however, its effects on systemic inflammatory and oxidative stress biomarkers are inconsistent. It is important to note that, based on the ISSN 2018 position stand, nitrate supplementation is recommended with either beetroot or sodium nitrate, at amounts ranging from 300 to 600 mg, 2–3 h before exercise (acute/single dose), as well as for daily use for up to a duration of approximately 6 weeks [12].
Blackcurrant anthocyanins
11.5
Blackcurrant is a type of berry, similar to blackberries and blueberries, that has been studied for their health-promoting benefits, primarily because of their high polyphenolic and anthocyanin contents [212]. In general, berries contain high concentrations of flavonoids, particularly anthocyanins, which are natural pigment responsible for the blue, purple, red, and orange colors of many fruits and vegetables, which are suggested to confer significant health benefits [213]. To this end, blackcurrant anthocyanins (delphinidin-3-rutinoside) contain between 130 and 460 mg/100 g of fruit weight of total anthocyanins, while the total daily estimated consumption amounts range from 3 to 215 mg/d [213]. Furthermore, blueberries contain approximately 62–300 mg/100 g of fruit weight of total anthocyanins, suggesting that blackcurrants may be more ideal sources of anthocyanins [213]. Tang et al. [214] demonstrated that blackcurrant anthocyanins have the ability to attenuate inflammation and oxidative stress while preventing the depletion and damage to the mitochondrial content within an animal model, further supporting the potential for blackcurrant as an effective dietary antioxidant, which may be a better source to leverage in the diet or via supplementation in comparison to other sources such as blueberry anthocyanins.
To date, New Zealand (NZ) blackcurrants appear to be the most highly concentrated in terms of anthocyanins and other phytochemicals compared to those from other countries. Schrage and colleagues found that the NZ blackcurrants contain between 336 and 850 mg/100 mL in juice versus the 170 and 310 mg/100 mL found in non-NZ blackcurrants. Considering this, several research groups have investigated the benefits of blackcurrant supplementation for exercise recovery and sports performance [215–227]. For instance, Willems et al. [227] assessed the impact of 1 week of 300 mg/d of blackcurrant extract supplementation in nine male endurance athletes (i.e. trained cyclists or triathletes with more than 3 years of experience). They found that there was an improvement in the 16.1 km (10-mile) time-trial cycling performance (3.6%) [227]. Another research group [225] also demonstrated that supplementation with blackcurrant juice (containing 300 mg of anthocyanins and 15 mg of vitamin C) resulted in a 1.9% improvement in peak running speed among 23 trained female runners. Furthermore, Cook et al. [228] found that NZ blackcurrant extract supplementation (containing 300 mg/d of CurraNZ*™* and 105 mg of anthocyanins) led to a 27% increase in fat oxidation at 65% of participants’ VO_2_max, alongside a 2.4% improvement in their 16.1 km time-trial performance. Recently, Braakhuis et al. [215] conducted a systematic review and meta-analysis on the effects of NZ blackcurrant within the context of sports performance and found that across nine total studies, there was a 0.45 (95% confidence interval [CI] = 0.09–0.81, p = 0.01) mean percent effect on performance. The authors noted that this improvement in sport-specific performance-related outcomes is not only relevant to athletes but also within the magnitude of effect for medal-winning at the Olympic level of competition [215]. Furthermore, the magnitude of the effect is similar to that observed with caffeine (i.e. 0.41, 95% CI = 0.15–0.68, p = 0.002) [229]. Interestingly, this meta-analysis by Braakhuis et al. [215] did not affect the reduction in oxidative stress or inflammatory biomarkers. The authors attributed this negative result to the low number of studies included; therefore, they suggested that meaningful conclusions could not be made with respect to these biomarkers. Nevertheless, the present data suggest that NZ blackcurrants can confer a small but meaningful improvement in athletic performance following 7 days of treatment with 105–210 mg blackcurrant anthocyanins when taken between 1 and 2 h before exercise [215].
Cocoa flavanols
11.6
Flavonoids are polyphenols composed of two phenyl rings and a heterocyclic ring, and include flavanols, flavonols, isoflavones, flavones, and anthocyanidins. Recent interest has been shown in the application of flavonols from various food sources, including cocoa, wine, fruits, vegetables, and teas. Notably, cocoa, which comes from the seeds of the Theobroma cacao tree’s fruit, is a rich source of flavonols [230]. Cocoa flavanols (CF) are of interest to athletic populations because of their ability to stimulate nitric oxide production, thereby improving vasodilation and endothelial function [231]. Endothelial function, measured by flow-mediated dilation, improves, a benefit acknowledged by the European Food Safety Authority (EFSA), which recommends an intake of approximately 200 mg/day of CF to help sustain normal endothelium-dependent vasodilation. This effect peaks approximately 2 h after ingestion [232,233]. A 2018 systematic review by Decroix et al. [231] noted that CF can improve vascular function, potentially alter substrate oxidation rates, and lessen exercise-induced oxidative stress; however, the authors concluded that more research is needed to assess the chronic effects of CF with and without exercise training. Furthermore, much of the research to date has concluded that the results are equivocal regarding the effects of CF on exercise-induced oxidative stress and exercise performance [234–241]. Nevertheless, Decroix et al. [231] concluded that acute CF supplementation of approximately 400–500 mg or higher 2 h before exercise, or even 2–3 months of CF supplementation, may aid in reducing exercise-induced oxidative stress and improving vascular function during exercise bouts. Furthermore, while the ideal dosage for CF consumption remains a topic of debate, higher total flavanol intake (>700 mg, particularly higher epicatechin intake >80 mg) has been shown to confer beneficial effects.
Coenzyme Q10
11.7
Coenzyme Q10 (CoQ10), also known as ubiquinone or ubiquinol, is a naturally occurring, fat-soluble biochemical cofactor that plays a role in energy production and acts as an endogenous antioxidant. Notably, CoQ10 can increase the production of endogenous antioxidant enzymes, such as SOD, or, when reduced to ubiquinol (Q10 H_2_), it can exert a direct antioxidant effect through free radical scavenging and electron donation to neutralize ROS/RNS [242]. CoQ10 has also been shown to regenerate vitamins E and C and reduce the expression of inflammatory biomarkers [243,244]. Given this, researchers have assessed the impact of CoQ10 supplementation on ROS/RNS in disease conditions, such as cardiovascular disease, diabetes, and hypertension [245–247]. Numerous studies have assessed the impact of CoQ10 supplementation on exercise performance [248–267], with doses ranging from 30–300 mg/day in healthy athletes to >600 mg/day in disease populations, for 2–12 weeks. Among these studies, several failed to demonstrate any impact of CoQ10 supplementation on performance and sport-related outcomes [248,249,251,252,255–257,260,262,264,265,267,268]. However, reports have shown favorable outcomes in reducing biomarkers of oxidative stress [253,258–260,264,269] and muscle damage [253,254,259,260,264]. For example, Emami and colleagues [260] found that taking 300 mg daily for 14 days prevents negative effects on myocardial damage and oxidative stress during the swimming competition phase. While CoQ10 has beneficial effects on attenuating biomarkers of inflammation, EIOS, and muscle damage, most studies demonstrate minimal to mixed effects of short- and long-term supplementation on performance-related outcomes [270–272].
Curcumin
11.8
Curcumin (1,7-bis(4-hydroxy-3-methoxyphenyl)-1.6 heptadiene-3,5-diona) is the main phenolic compound found in turmeric (Curcuma longa L.), which is the oriental yellowish-colored spice of the ginger (Zingiberaceae) family. Curcumin has been investigated as an adjunctive treatment for its pain-reducing [273,274] and anti-inflammatory properties [275] and has emerged as a medicinal treatment for chronic/cardiometabolic disease states [276–278]. In the context of exercise and sports performance, several studies have demonstrated a potential beneficial effect of curcumin supplementation, with few to no adverse side effects [279–289]. Suhett and colleagues [290] conducted a systematic review of the literature on the application of curcumin supplementation in sport and physical exercise and found that several studies demonstrated (1) antioxidant and anti-inflammatory effects [279,281–286,288], (2) ergogenic properties following various supplementation protocols (i.e. both short- and long-term) [280,284,287,288], and (3) reductions in pain perception and muscle damage [284,288,289,291]. For example, Delecroix et al. [280] assessed the impact of a single 2 g dose of curcumin co-ingested with 20 mg of piperine among 10 elite rugby players following an exercise-inducing muscle damage protocol and found a lower loss of mean power during sprinting with the curcumin treatment (1.77% loss) compared to the placebo (13.6% loss). Furthermore, 6–10 mg/day of curcumin appears to be associated with reduced pain and muscle damage [280,284,288,289]. In terms of longer-term benefits, data suggest an antioxidant effect (i.e. lower advanced glycation end products and MDA) following 3 months of curcumin supplementation (10 mg/day) [291]. Suhett and colleagues [290] suggested that doses ranging from 180 to 500 mg/day for 3–7 days are efficacious for improving performance and recovery-related parameters following exercise. However, long-term data are limited, and most of the current literature surrounding curcumin suggests that performance and recovery benefits are observed only with short-term supplementation [290]. In summary, despite its poor bioavailability [292], short-term curcumin supplementation (i.e. up to 1 week) at 180–500 mg/day has been shown to improve performance and recovery-related outcomes. Limited data suggest that longer supplementation periods may be beneficial; however, more research is warranted to demonstrate repeatability.
Creatine monohydrate
11.9
Although not considered a traditional antioxidant, creatine monohydrate (CrM), arguably the most researched ergogenic aid [293,294], has been purported to possess indirect antioxidative properties [295]. Studies have demonstrated that CrM can reduce free-radical-induced damage and inflammation following exercise in animal and human models [296–304]. Nevertheless, the mechanism by which CrM acts as an antioxidant is not fully understood, with some reports suggesting that CrM can increase endogenous antioxidant enzyme activity and the capacity to neutralize ROS/RNS [305–307]. Relatively few human studies have investigated the antioxidant impact of CrM, with mixed results reported to date. Kingsley et al. [308] demonstrated that 7 days of CrM loading dose (20 g/day) did not favorably impact the levels of markers of lipid peroxidation or plasma concentrations of non-enzymatic antioxidants. However, Rahimi [304] found that 20 g/day of CrM supplementation for 7 days reduced urinary 8-OHdG and plasma MDA levels in response to resistance training. Then, Amiri and Sheikholeslami-Vatani [297] found that 10 weeks of resistance training and CrM supplementation (0.1 g/kg/day) increased GPx enzyme levels. Despite these favorable findings, one report suggested that CrM could act as a pro-oxidant [309]. In terms of anti-inflammatory effects, reports have shown that CrM supplementation can act on specific immune cells and reduce the concentrations of the pro-inflammatory cytokines TNF-α and prostaglandin E_2_ (PGE_2_), eliciting decreased inflammation post-exercise [310–313]. For instance, Santos et al. [310] reported that CrM supplementation (20 g/day for 5 days) attenuated changes in PGE_2_ (60.9%) and TNF-α (33.7%) among runners, while those receiving the placebo exhibited increases in PGE_2_ (6.6-fold) and TNF-α (2.34-fold). Furthermore, Bassit et al. [299] demonstrated that 20 g/day for 5 days of CrM supplementation before completing an Ironman reduced the increase in IL-1, IL-6, INF-α, and TNF-α. These findings have been similarly supported by another report from Rawson et al. [314]. However, there are still mixed results regarding CrM’s anti-inflammatory activity, warranting further research to better understand its antioxidant benefits as a supplement. Notably, CrM is one of the rare dietary supplement with a substantial body of evidence supporting its efficacy as an ergogenic aid [293,294]. Therefore, while CrM may be considered to have a low level of support for its antioxidant ability, it is one nutrient that should be considered for its primary benefit (i.e. exercise performance enhancement), and the potential added benefits (i.e. antioxidant/anti-inflammatory) likely serve as an “add-on.”
Fucoxanthin
11.10
Fucoxanthin, a naturally occurring carotenoid found in brown seaweeds, macroalgae, and diatoms [315,316], has a unique structure featuring an allenic bond and nine conjugated double bonds that enhance its stability and antioxidant activity [317,318]. Fucoxanthin is metabolized into Fucoxanthinol and Amarouciaxanthin A, which are considered bioactive compounds. Furthermore, although fucoxanthin is sensitive to heat and air, its bioactivity also stems from its conjugated bonds and high molecular polarity [315,319]. Emerging research suggests that microalgae-based supplements, such as *Phaeodactylum tricornutum (*PT), which contain EPA, DHA, and fucoxanthin, may offer anti-inflammatory and performance-supportive benefits for athletes [315,320–323]. For instance, Stievatter and colleagues [324] demonstrated that PT supplementation (i.e. whole PT containing 294 mg of EPA+DHA and 21 mg of fucoxanthin) for 2 weeks reduced IL-6 levels (46%) and improved functional performance (sit-to-stand time) compared to placebo. In addition, Dickerson et al. [325] demonstrated that PT (4.4 mg of fucoxanthin) combined with supervised exercise training over 12 weeks led to improvements in maximal aerobic capacity, resting heart rate, subjective measures of adherence to a training program, bone mineral indices, HDL-c, and LDL-c/HDL-c ratios in women without affecting weight loss. In addition to potential muscle effects, Abidov et al. [320] conducted a study in postmenopausal women. They reported improvements in total energy expenditure after 16 weeks of high-dose (8 mg/day) fucoxanthin supplementation. There have also been reports of fucoxanthin’s ability to improve aspects of cognitive function among E-gamers [322] and elderly populations [323], attributed mainly to its antioxidant and anti-inflammatory properties, which are important outcomes related to sports performance. Taken together, fucoxanthin supplementation, particularly from the carotenoid-rich microalga PT, shows promise for impacting cardiometabolic and exercise-related outcomes. Importantly, fucoidan, a fucose-rich sulfated polysaccharides, derived from either brown seaweed or echinoderms, can improve the inflammatory and immune response after high-intensity exercise [326]. However, further work is needed in sport-specific populations to understand its impact on exercise and sports performance.
Glutathione and N-acetylcysteine
11.11
Glutathione is a predominant endogenous antioxidant consisting of the amino acids glutamate, cysteine, and glycine, while N-acetylcysteine (NAC) is a cysteine precursor that can be used to recycle glutathione [327]. To date, the methods of glutathione administration (i.e. oral versus intravenous) may impact the effectiveness of the dose and duration [328,329]. For instance, Schmitt et al. [329] demonstrated that an oral dose of 450 mg/day for three weeks of glutathione improved antioxidant status, while a single dose of 3000 mg delivered intravenously improved TAC [328]. It has been previously reported that exogenous glutathione is poorly stable and may degrade in the extracellular compartment, rendering it less bioavailable [82]. This may explain why most trials involving healthy adults have generally not demonstrated increases in glutathione concentrations from supplementation [330]; however, a 6-month randomized controlled trial involving apparently healthy male and female subjects reported increases in whole blood, plasma, and erythrocyte concentrations of glutathione following supplementation with low-to-high doses of glutathione (250–1000 mg daily) [331]. In the context of exercise performance, Hwang et al. [332] demonstrated that a combination of glutathione (200 mg/day) and L-citrulline (2 g/day) supplementation, coupled with resistance training for 8 weeks, increased lean mass without affecting any blood clinical chemistry parameters. Another study by Aoi and colleagues [333] found that a 1 g/day dose of glutathione for 2 weeks resulted in suppressed blood lactate levels following 60 minutes of cycling exercise. In terms of NAC, which is chemically similar to glutathione and favors glutathione synthesis, especially among individuals who are deficient in glutathione [132,334,335], the evidence is limited but promising. NAC has been shown to improve aspects of aerobic and anaerobic performance (i.e. V̇O_2_max, time trial, and anaerobic power) by 11.4%–15.4% among individuals who demonstrated baseline glutathione deficiencies. In contrast, performance was unchanged in individuals with high baseline glutathione concentrations [132,334,335]. Kerksick and colleagues [84] supplemented healthy, active males with either 1800 mg of NAC or a placebo for 14 days before a damaging bout of eccentric muscle contractions and three days after the exercise bout. Plasma markers of muscle damage and intramuscular markers of oxidative stress (8-isoprostane and SOD), inflammation (TNF-α, cortisol, neutrophils, neutrophils: lymphocytes), and apoptosis (bax, bcl-2, bax:bcl-2 ratio, and caspase-3) were assessed. NAC supplementation did not affect TNF-α, cortisol, 8-isoprostane, or SOD levels. Currently, data on NAC supplementation suggest that doses of 1.2–2.0 g/day, taken acutely or over several weeks, may affect performance outcomes, with minimal benefits [4]. Ultimately, the true ergogenic and antioxidant effects of NAC in the context of exercise and sports performance remain unclear and require further research [336].
Green tea catechins
11.12
Green tea catechins, specifically epigallocatechin gallate (EGCG), are part of the flavonol family, known for their potent antioxidant, anti-inflammatory, and vascular-regulating effects, which are important for exercise and recovery [337–343]. Their structure includes multiple hydroxyl groups that facilitate the neutralization of free radicals, decrease oxidative stress, and modulate nitric oxide bioavailability, potentially enhancing endothelial function and metabolic health [340]. In addition to their direct antioxidant effects, catechins enhance the body's antioxidant defenses by increasing the activity of enzymes, thereby helping the body withstand oxidative stress from exercise [338]. Human studies indicate that consistent pre-exercise catechin supplementation for at least 7–14 days can enhance overall antioxidant capacity and reduce oxidative stress markers following endurance or high-intensity exercise [340]. These effects tend to be more consistent with repeated dosing rather than with a single dose [1]. When performance benefits are observed, they typically involve modest improvements in fat oxidation during submaximal exercise, increased metabolic flexibility, or slight decreases in perceived effort [344]. In a double-blind, prophylactic supplementation trial, Kerksick and colleagues [84] demonstrated that 14 days of NAC (1800 mg/day) or EGCG (1800 mg/day) resulted in a slight reduction in subjective muscle soreness 24 h after eccentric knee extension exercise. However, neither supplement significantly affected strength loss, CK, LDH, inflammatory cell counts, or apoptotic signaling compared with the placebo. Kerksick et al. [85] in a follow-up study revealed that 14 days of NAC or EGCG did not reduce early increases in the MuRF1, UBE3B, or calpain genes after eccentric exercise in young males. Nevertheless, evidence for substantial improvements in time-trial or maximum performance remains inconsistent [342]. Some research suggests that exercise may enhance vascular function and reduce muscle soreness, possibly through the anti-inflammatory properties of catechins [1]; however, the findings are mixed [1]. No effects have been observed with products low in catechins, short-term dosing, or highly trained athletes whose bodies may already be adapted. Taking a high dose of EGCG immediately before resistance training can temporarily reduce blood flow or cause gastrointestinal issues in some individuals, highlighting the importance of proper timing and dosage [1]. In practice, athletes aiming for antioxidant support without impairing training adaptations may consume 400–800 mg/day of catechins (≈300–500 mg EGCG) from standardized extracts, or approximately 2–3 cups of brewed green tea daily [1]. Most protocols begin 7–14 days before major training sessions or competitions to foster the buildup of antioxidant enzymes. Immediate high-dose pre-workout supplements should be avoided, as they can disrupt redox signaling associated with training. Considering the potential risk of increased liver injury at high catechin doses (e.g. exceeding 800 mg EGCG daily) [345], along with an unclear safety threshold and limited evidence of exercise-related benefits, green tea-based catechin supplements may not be the best choice for athletes as antioxidants [345]. Green tea catechins, especially EGCG, can enhance antioxidant defenses, support vascular health, and influence metabolic responses important for endurance and intense training [1,340]. Although the performance improvements are generally modest and vary, regular intake of catechin-rich products can support recovery and maintain redox balance when integrated thoughtfully into an athlete’s diet [1,340].
Lutein and zeaxanthin
11.13
Lutein and zeaxanthin (LZX) are xanthophyll carotenoids primarily found in dark leafy green vegetables (i.e. kale, spinach) and egg yolks. Their hydroxylated β-ionone ring structures confer hydrophilic properties, enabling more efficient reactivity with oxidants. LZX is highly concentrated in the eye, where it acts as an antioxidant and a blue-light filter, particularly in the macula and retina, helping protect against age-related macular degeneration and cataracts [346,347]. Although lutein and zeaxanthin primarily accumulate in the macula and retina, numerous observational and clinical studies have shown that a higher dietary intake of L/Z is associated with a lower risk of cataracts [348,349]. These links seem to result from the systemic antioxidant effects of L/Z rather than their presence in the retina. Several meta-analyses and randomized clinical trials have demonstrated that higher intakes (>20 mg/day) of LZX over extended periods (up to 12 months) improve macular pigment optical density and visual function [350–361]. Furthermore, LZX deficiency may impair visual function in sports such as baseball, tennis, and target sports [362]. Yoshida and researchers [363] found that supplementation with 10 mg of lutein and 2 mg of zeaxanthin for 8 weeks improved hand‒eye coordination. Additional studies have reported enhanced visual processing speed, reaction time, and neural efficiency following LZX supplementation at 26–34 mg/day in healthy young adults [357,358]. These improvements could benefit athletes participating in visually demanding or high-speed environments.
There are also data suggesting that LZX supplementation can aid older athletes. For example, Murphy and colleagues [364] performed a cross-sectional analysis and longitudinal follow-up on an elderly population. They found that those with higher LZX concentrations had lower odds of frailty progression, better timed-up-and-go scores, and a more positive bone stiffness index. Additionally, Sahni and colleagues [365] found very similar findings; however, a positive association was observed directly between the serum LZX and muscle strength in older adults. Though these results are not derived from a supplementation or sports perspective, they still offer exciting potential for LZX to confer direct musculoskeletal effects, potentially improving exercise and sports performance. In summary, LZX are potent antioxidants that have shown efficacy in improving visual performance-related outcomes and may carry over into sports. Athletes may benefit from increased LZX intake to support visual acuity, cognitive processing, and potentially muscular function.
Omega-3 fatty acids
11.14
Polyunsaturated fatty acids (PUFAs), particularly omega-3 (ω-3) fatty acids such as eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), play a central role in redox homeostasis and in cellular protection against ROS/RNS damage. Unlike saturated fats, PUFAs contain multiple (two or more) carbon‒carbon double bonds that strengthen membrane fluidity and stability, improving defenses against radicals [366–369]. Mechanistically, ω-3 s can (1) activate the Nrf2 pathway, enhancing endogenous antioxidant enzyme activity; (2) reduce ROS/RNS production via mitochondria-modulating mechanisms [370,371]; and (3) increase the amount of cardiolipin present, an inner mitochondrial membrane phospholipid that facilitates oxidative phosphorylation [372,373]. In addition, within mitochondria, DHA suppresses cytochrome C oxidase activity while simultaneously increasing the activity of manganese (SOD), a key mitochondrial antioxidant enzyme [374]. The consumption of diets rich in fish oil results in increased levels and activity of SOD, along with reduced lipid peroxidation, as indicated by lower concentrations of thiobarbituric acid-reactive substances (TBARS) [375]. In conclusion, ω-3 intake leads to marked antioxidant responses, primarily by restoring imbalances in endogenous antioxidant systems [374].
Several meta-analyses have demonstrated that supplementation with ω-3s can confer an antioxidant benefit across diseased and generally healthy populations. Heshmati and colleagues [376] assessed 39 trials with over 28,000 participants and found improvements in TAC and GPx activity along with reductions in MDA; however, the effects on nitric oxide, GSH, SOD, and CAT were equivocal. In another meta-analysis, ω-3 supplementation reduced CRP and increased TAC, but did not affect MDA or GSH [377]. Two other data further support improvements in TAC with prolonged ω-3 supplementation alongside mixed findings for nitric oxide, MDA, and endogenous antioxidant enzyme activity [366,376,377].
Evidence from randomized clinical trials has consistently demonstrated an antioxidant and anti-inflammatory benefit. For instance, Ghiasvand and colleagues [378] found that six weeks of 2000 mg/day of EPA, coupled with vitamin E supplementation, increased the production of anti-inflammatory cytokines, attenuated TNF-α release, and had additional effects on MDA. There are also clinical trials showing a decrease in serum and/or plasma MDA concentrations after prolonged ω-3 supplementation [379–384], with at least 1440 mg/day of a combined EPA and DHA formula from fish oil. Similarly, the results from clinical trials have shown marked improvements in TAC after at least 12 weeks of treatment with 1000 mg/day ω-3, including varying amounts of EPA and DHA [385–388]. In the context of exercise, ω-3 also has anti-inflammatory and antioxidative effects and can mitigate post-exercise muscle damage. For example, Barquilha and colleagues [389] found reductions in CRP, IL-6, CK, and LDH, and an increased GSH/GSSG ratio post-resistance training in physically active men and women who were supplemented with 260 mg/day of EPA and 202 mg/day of DHA for six weeks. Lee and colleagues [390] found that the consumption of fish oil (2100 mg/day of EPA and 720 mg/day of DHA) by older adults participating in a 12-week resistance training program decreased the IL-6, CRP, and TNF-α levels. Recently, VanDusseldorp and colleagues [391] demonstrated that fish oil supplementation of 6000 mg/day of ω-3 (2400 mg/day of EPA and 1800 mg/day of DHA) for approximately seven weeks resulted in attenuation of muscle damage, CK, and LDH 72 h post-eccentric exercise alongside improvements in subjective measures of muscle soreness in college-aged resistance-trained males. Although the results from exercise studies offer promise regarding the effects of ω-3s on muscle damage and oxidative stress markers concurrent with an exercise program, some research has yielded equivocal findings [392]. Nevertheless, supplementing with at least 1000 mg/day of ω-3s with a mixed EPA and DHA formula from fish oils or intake of an equivalent dose of ω-3 from food sources (i.e. cold water fish, flaxseed, and walnuts) yields antioxidative effects on a variety of populations (diseased, healthy, and athletic).
Pomegranate polyphenols and urolithin A
11.15
Pomegranate is a berry-like fruit (produced by Punica granatum) with approximately 52% of its total weight being edible, comprising approximately 78% juice and 22% seeds [393]. Typically, supplementation involves juice pressed from the whole fruit; however, pomegranate extracts in liquid or dry powder forms have gained popularity as concentrated sources of bioactive polyphenols [394]. Interest in pomegranate among athletes and active individuals has increased because of its high polyphenol content and associated cardiometabolic and anti-inflammatory benefits [394,395]. The consumption of pomegranate has been shown to decrease oxidative stress, inflammation, blood pressure, and blood lipids, and its polyphenols have been further researched for their effects on exercise performance and recovery [394,395]. Randomized controlled trials indicate that consuming high-polyphenol juice both before and after eccentric or resistance exercise results in faster strength recovery and reduced soreness [396,397]. Systematic reviews highlight that benefits are more consistent when juices contain more than 0.7 g of total polyphenols per 0.5 L, especially during exercise involving large muscle mass, and when juices are consumed at least 60 min prior to activity [394,395,398]. Conversely, studies in less trained groups or with lower-polyphenol products have shown no significant effects [395,399]. Typical supplementation involves 500–1000 mL of polyphenol-rich pomegranate juice (often divided into multiple doses) or standardized extracts, with at least 500–1000 mg administered daily [395,396,399–403]. These are typically initiated 3–7 days before intense training or events and maintained for 2–3 days afterward [395,396,399–403].
Urolithin A (UA) has garnered attention over the last decade for its promising applications in sports performance [404]. Derived from dietary precursors, such as pomegranates, UA is a naturally occurring metabolite that appears to play a crucial role in enhancing mitochondrial function by stimulating mitophagy and mitigating the age-related accumulation of dysfunctional mitochondria [405]. In terms of its health applications, UA has been assessed within the contexts of osteoarthritis [406], immunity [407], and neurodegenerative disease and disorders [408,409]. More recently, randomized controlled trials have shown much promise for UA in the application of sport [410–412]. For instance, Zhao et al. [412] assessed the impact of 8 weeks of 1 g/d of UA supplementation in a randomized, double-blind, placebo-controlled fashion among 20 resistance-trained male athletes. The UA group showed non-statistically significant increases in the 1RM bench press and squat (Δ = 3.00 ± 0.17 kg, p = 0.051; Δ = 1.35 ± 2.73 kg, p = 0.499, respectively). However, there were significant improvements in maximum voluntary isometric contraction and repetitions to failure (Δ = 36.10 ± 0.62 NM, p = 0.000; Δ = 2.00 ± 0.56, p = 0.001, respectively). Compared to the placebo group, the UA group also improved in maximum voluntary isometric contraction and repetitions to failure after 8 weeks (Δ = 43.50 ± 0.77 NM, p = 0.048; Δ = 2.00 ± 1.22, p = 0.011). Additionally, the UA group showed reductions in CRP (Δ = –0.79 ± 0.38 mg/L, p = 0.032) and SOD (Δ = −4.32 ± 0.90 U/mL, p = 0.041) compared with the placebo group. Whitfield et al. [411] demonstrated that 1000 mg/d of UA for 4 weeks during an altitude training camp (≈1700–2200 m) led to a reduction in ratings of perceived exertion during a 3000 m time trial (p = 0.02), as well as reduced indirect markers of post-exercise muscle damage, such as CK (p < 0.001). Furthermore, the UA group experienced a greater within-group increase in V̇O_2_max (5.4 ± 0.9%, 66.4 ± 0.8 to 70.0 ± 1.0 mL·kg^−1^·min^−1^, p = 0.009, d = −0.83) while only small changes were found in the placebo group (3.6 ± 1.3%, 66.4 ± 0.9 to 68.7 ± 1.0 mL·kg^−1^·min^−1^, p = 0.098, d = −0.54) [411]. Finally, UA upregulated mitochondrial pathways, decreased inflammatory pathways, and increased the expression of markers of mitophagy [411]. While UA supplementation is promising, more work is needed to understand the optimal dosing and duration.
Pycnogenol
11.16
Pycnogenol^®^ (PYC) is a procyanidin-rich extract derived from pine bark (Pinus pinaster) known for its potent antioxidant activity. PYC can neutralize ROS/RNS [413], upregulate endogenous enzyme activity [414], modulate NOS [415], inhibit lipid peroxidation [416], and exhibit metal-chelating properties [417]. Clinical studies have shown that daily PYC supplementation (100–200 mg) over 12 weeks can improve cardiometabolic health outcomes, including blood pressure, lipids, glucose, hemoglobin A1c, and biomarkers of oxidative stress and inflammation [418–423]. In addition to its metabolic health effects, PYC has functional benefits, such as reduced osteoarthritic pain and improved physical performance (WOMAC index) following supplementation at 150 mg/day for 90 days [424]. In a more athletic context, PYC has shown potential ergogenic effects, with studies reporting improvement in maximal aerobic capacity, aerobic power, time to fatigue, and recovery parameters in cyclists, triathletes, and military trainees at doses ranging from 60 to 200 mg/day for 4–8 weeks [425–429]. For instance, Hara and associates [426] found that 28 days of 60 mg/day of PYC supplementation led to enhanced maximal aerobic capacity, aerobic power, and caloric expenditure during cycling exercise. Furthermore, supplementation with PYC has also resulted in reductions in EIOS, improved performance on standardized physical ability tests (e.g. sit-ups and push-ups), and quicker total race times in triathlons [426,428]. Reports have also demonstrated PYC’s ability to mitigate exercise-induced cramps and muscle pain at doses ≥100 mg/day for 3–5 weeks [427,429,430]. Finally, PYC has demonstrated positive effects on cognitive function at doses ≥150 mg/day [431–435] in addition to enhanced working memory, pattern recognition, and mood, and reducing serum oxidative markers in both young and older adults, who typically use 100–150 mg/day for 12 weeks or longer [432,433,435–438]. In summary, PYC appears to support aspects of cardiometabolic health, cognitive and physical performance, and recovery in clinical and exercise settings. While data on PYC are limited, athletes may benefit from doses ≥100 mg/day across short- and long-term durations, and future studies should explore optimal supplement strategies.
Quercetin
11.17
Quercetin is a flavonoid found predominantly in citrus fruits and green leafy vegetables, with exceptionally high concentrations in capers, red onion, berries, red wine, buckwheat tea, and broccoli [439,440]. Its wide availability in whole foods makes regular intake a practical dietary strategy, with evidence suggesting benefits for metabolic health, including reductions in inflammation [441–443] and free radicals [439,444–447], as well as potential improvements in performance markers such as neuromuscular function and reduced soreness [258,259]. Mechanistically, quercetin is thought to activate the Nrf2 signaling pathway, thereby indirectly enhancing the expression of endogenous antioxidant enzymes [260]. However, findings on its ergogenic potential in athletic populations are limited and inconsistent. Some studies report improvements in outcomes, such as cycling performance, mitochondrial biogenesis, and body composition [448,449], while others report no significant differences compared to controls [450]. Most research has used doses of 400–600 mg, which are administered once to three times daily between meals for 7 days to 8 weeks [450]. Owing to quercetin's the poor bioavailability, its absorption can be enhanced by co-ingesting it with vitamin C or other vitamins and minerals, particularly when it is consumed after meals [450]. After absorption, quercetin undergoes extensive metabolism in the gut and liver, resulting in low levels of active quercetin in the bloodstream [451]. Differences in bioavailability – due to the form of quercetin in foods and individual enzyme variability – can further reduce its effectiveness. These metabolites may exhibit relatively weak antioxidant activity, potentially limiting the overall impact of quercetin on the body. While some evidence supports quercetin’s ability to reduce exercise-induced ROS and improve performance, further research in athletes is needed to confirm its efficacy.
Resveratrol
11.18
Resveratrol (3,5,4′-trihydroxystilbene) is a naturally occurring polyphenolic compound found in grapes, red wine, peanuts, and berries [452,453]. It possesses potent antioxidant and anti-inflammatory properties, scavenging ROS/RNS directly from the body. Resveratrol can also activate cellular pathways, such as SIRT1, AMPK, and Nrf2, which regulate mitochondrial biogenesis, energy metabolism, and redox balance [454,455]. Preclinical trials have consistently demonstrated that resveratrol can enhance exercise-induced adaptations, including improvements in mitochondrial function, endurance performance, and resistance to oxidative stress [456–458]. However, existing human data remain less supportive, reporting mixed or limited benefits. Some trials suggest that resveratrol may blunt training adaptations by interfering with redox-sensitive signaling [459–461]. For instance, Gliemann et al. [459] found that 250 mg/day of resveratrol in older men undergoing 8 weeks of high-intensity training attenuated gains in VO₂max and cardiovascular improvements. Similarly, Tsao et al. [462] found that although 480 mg/day resveratrol supplementation for 4 days reduced IL-6 levels in young athletes, it had no effect on other oxidative or performance markers. Because of these inconsistencies, resveratrol is not currently considered a strong candidate for use as a performance or recovery aid for healthy, active individuals or athletes, and further research is needed to determine the optimal dose. The heterogeneity in human responses underscores the need for carefully designed, randomized controlled trials, particularly among populations that stand to benefit most, such as older adults or individuals with physical limitations. For example, a pilot randomized controlled trial was conducted by Anton et al. [463] explored the efficacy of resveratrol combined with exercise in older adults, highlighting the importance of dose-dependent effects and potential improvements in functional outcomes. Future research should clarify optimal dosing, timing, and bioavailability issues, as well as interactions with physical training, to better define the role of resveratrol in supporting physical performance and health.
Selenium
11.19
Selenium is an essential trace mineral found in foods such as Brazil nuts, seafood, organ meats, and whole grains [464,465] that serves as a cofactor for various selenoproteins (e.g. GPx) involved in endogenous antioxidant defense and immune regulation. Selenium exerts many of its biological effects through its incorporation into GPxs, thioredoxin reductases, and selenoprotein P – key enzymes that mitigate oxidative stress by neutralizing hydrogen peroxide and lipid hydroperoxides [466]. Several studies have investigated the impact of selenium on redox balance and inflammation in physically active populations. For instance, trained cyclists who were supplemented with 240 µg/day of selenium-enriched yeast for 10 weeks showed increased GPx activity and reduced MDA levels compared with those in the placebo group [467]. In another study among active females, selenium supplementation (250 µg/day for 3 weeks) reduced oxidative stress and increased antioxidant levels, with the selenium-only group showing greater changes than the selenium + HIIT group [468]. In addition to its antioxidant effects, selenium supplementation has been shown to reduce serum IL-6 levels, though it does not affect CRP or TNF-α levels [469]. In addition, a selenium-containing supplement providing 50 µg/day of selenium over 21 days of intensive training supported physical performance and heart rate regulation in highly skilled tracks and field athletes [470]. Furthermore, in a recent study, individuals who were supplemented with selenium experienced faster decreases in IL-6 and CRP levels after high-intensity exercise, suggesting that selenium may enhance post-exercise recovery through its anti-inflammatory effects [471]. While selenium does not consistently enhance performance [472], supplementation can prevent deficiencies, reduce EIOS and mitochondrial changes, and may aid recovery – though evidence in trained athletes remains limited.
Spirulina
11.20
Spirulina is a blue‒green algae-derived, photosynthetic, gram-negative bacterium (Arthrospira platensis) that has been shown to have antioxidant, anti-inflammatory, anti-diabetic, anti-hypertensive, and lipid-lowering effects, with a safe dosage range of 3–10 g/day [473]. Currently, in the field of sport and exercise nutrition, spirulina supplementation is not well established for its positive ergogenic effects; however, several human trials may provide insight into its potential applications [474]. Koite and colleagues [475] found that supplementation with 2 g/L of Arthrospira extract (containing 1 g/L of phycocyanin and 0.5 g/L of Spirulysat, which includes proteins, amino acids, enzymes, vitamins, and minerals) for 12 weeks led to a reduction in urinary isoprostane levels. Bohorquez-Mordai et al. [476] conducted a systematic review and meta-analysis. They found that spirulina supplementation led to reductions in body mass, body fat percentage, and waist circumference among obese adults. Furthermore, Ismail and colleagues [477] found that spirulina (i.e. [500 × 2] or [500 × 4] mg for 60 days) resulted in reductions in the serum concentrations of MDA, lipid hydroperoxide, and total cholesterol, with increased GSH, vitamin C, SOD, glutathione-s-transferase activity, and glutathione. Recently, a study by Krokidas et al. [478] demonstrated that 42 mg/kg/day of spirulina for 15 days did not affect isometric muscle performance or exercise-induced muscle damage. More research is needed to better understand the impact of spirulina supplementation in athletic and trained populations.
Sulforaphane
11.21
Sulforaphane (SFN) is a naturally occurring isothiocyanate derived from cruciferous vegetables (i.e. broccoli sprouts), which contain high concentrations of its precursor, glucoraphanin. Upon chewing or processing, the enzyme myrosinase converts glucoraphanin to SFN (allowing it to be bioavailable) [479,480], which has drawn interest for its potent indirect antioxidant, anti-inflammatory, and cytoprotective properties [16,481–483]. Importantly, cooking deactivates myrosinase, and many broccoli supplements contain only glucoraphanin, reducing its efficacy unless it is paired with active myrosinase [484,485]. SFN primarily exerts its antioxidant effects [17,486,487], enabling a hormetic effect. This makes it distinct from traditional antioxidants (e.g. vitamins C and E), which can blunt training adaptations when used excessively [488]. SFN has also been shown to suppress pro-inflammatory cytokines (e.g. IL-6 and TNF-α), reduce NF-κB activation, and improve mitochondrial health [489–491]. These characteristics make SFN a compelling candidate for supporting recovery from intense training, reducing inflammation, and promoting metabolic adaptations in athletic populations [492–494]. In general, supplementation protocols have involved participants ingesting ≈20–40 mg/day of SFN derived from broccoli sprout extracts or supplements for 4–12 weeks [495,496]. Recent studies [492–494] suggest that SFN supplementation reduces post-exercise CK levels and shortens perceived recovery time without hindering hypertrophy or strength development [497]. Limited but promising data are available on the application of SFN in the context of exercise and recovery. For instance, Sato et al. [494] reported that 28 days of SFN supplementation (30 mg/day) in healthy men reduced plasma creatine kinase levels following an upper-body exercise-induced muscle-damage protocol compared with placebo. Similarly, Komine et al. [497] demonstrated that, compared with a control diet, 14 days of SFN supplementation (30 mg/day) before a muscle-damaging protocol reduced the expression of markers of soreness and oxidative stress. While more large-scale human trials are needed, the current findings support SFN’s use as a non-blunting, recovery-promoting antioxidant, especially when derived from myrosinase-active broccoli sprout extracts.
Tart cherry
11.22
Montmorency tart cherries (Prunus cerasus L.) are rich in phytonutrients, particularly flavonoids and other phenolic compounds, which have potent antioxidant and anti-inflammatory properties [498]. Emerging evidence suggests that the consumption of tart cherry juice or a powdered form may help lower cholesterol and triglyceride levels, improve blood pressure regulation and blood glucose control, enhance sleep quality and cognitive function, and support exercise recovery [498–501]. Given their high content of anthocyanins and polyphenols, tart cherries have garnered widespread interest among athletes and researchers for their potential to reduce muscle soreness, accelerate recovery, and support performance following high-intensity training [501,502].
To date, numerous studies have assessed the impact of tart cherry supplementation on resistance training [399,503–509], long-distance running [503,510–512], cycling [513,514], repeated sprints [513], and sport-specific training [515]. In terms of resistance training, as with some of the results seen with AST supplementation, the data demonstrate no unfavorable impact on key aspects of training adaptation [507]. For example, Jackman et al. [507] found that supplementation with 60 mL/day of Montmorency tart cherry concentrate for two weeks did not affect the anabolic response to resistance training. Notably, dietary antioxidants can negatively influence the natural training adaptation response [4]. Nevertheless, this does not appear to be the case with short-term tart cherry supplementation, at least at present.
Additionally, a 2020 meta-analysis of 10 studies revealed that consuming tart cherries for 7 days before and up to 1.5 h before an endurance exercise bout improved performance outcomes, such as time trials, time to exhaustion, and total work performed [516]. Wangdi and colleagues [517] later demonstrated that acute supplementation with tart cherry providing over 800 mg of polyphenols improved performance in a 15-km cycling time trial. The greatest improvements occurred when supplementation occurred 90 min before exercise (compared to 30 or 150 min), aligning with the greatest phenolic metabolite exposure during exercise. Notably, the performance benefits were more pronounced in trained individuals than in recreationally active individuals.
Tart cherry supplementation has also been shown to support post-exercise recovery [502]. Preloading tart cherry juice (i.e. two daily 30 mL servings) for several days before exercise can help preserve muscle function across various forms of physical activity [502,518]. A 2021 meta-analysis of 14 studies reported improvements in strength, reduced DOMS, and lower CRP and IL-6 levels, but no effect on CK or TNF-α [502]. Moreover, tart cherry supplementation has been shown to enhance the recovery of muscle function, reduce inflammation and oxidative stress markers, and alleviate pain following high-intensity running and cycling [510,511,519,520], with similar benefits observed after repeated sprint exercise [513,521,522]. Some studies have shown that tart cherry juice can reduce pain and promote strength recovery following heavy eccentric resistance exercise [505,508]. Supplementation with tart cherry powder (480 mg/day) has also been shown to reduce markers of muscle damage and inflammation and to decrease perceived soreness following intense strength and endurance exercise in trained individuals [504,512]. Finally, tart cherry supplementation may also increase circulating melatonin levels, thereby improving sleep duration and quality, further supporting recovery [523].
Given its potent antioxidant and anti-inflammatory properties, tart cherry supplementation appears to be a viable strategy for athletic populations to enhance exercise and sports performance while attenuating EIOS and inflammation, thereby promoting quicker recovery following high-intensity training [501–505,508,510–513,515,516,521,524]. Several tested supplementation dose and duration protocols have been reported in the literature, utilizing both powdered and juice tart cherry and varying timing strategies [502,516,525]. Generally, athletes considering tart cherry supplementation should aim for one of the following protocols: (1) a 200–500 mg capsule of tart cherry powder (containing 66–257 mg of anthocyanin), (2) 60–90 mL of cherry juice concentrate diluted with water (containing ≈550–820 mg of anthocyanin), or (3) 300–400 mL of cherry juice (containing ≈80 mg of anthocyanin) [502,516,525].
Vitamin E and vitamin C
11.23
Although the FDA recognizes Vitamin E (α-tocopherol) and Vitamin C (ascorbic acid) as antioxidants, the data overwhelmingly indicate mixed or limited performance and recovery-enhancing benefits when taken alone or in combination within the context of exercise and sport [3]. Vitamin E comprises lipid-soluble compounds that contain four tocopherols and four tocotrienols, with α-tocopherol being the most biologically active form. Tocopherols and tocotrienols are potent free radical scavengers. Once vitamin E neutralizes radicals (e.g. fatty acid peroxyl or tocopheroxyl radicals), vitamin C can regenerate it, allowing it to continue scavenging ROS/RNS [526]. Vitamin C is hydro-soluble and can also play a role in direct free radical scavenging. While vitamins E and C can be taken alone, Higgins and colleagues [3] conducted a systematic review of studies up to October 2020, concluding that vitamin E is most effective when combined with vitamin C, while vitamin E alone lacks strong support for enhancing sports performance.
Currently, the recommended daily allowance (RDA) for vitamin E is 15 mg (α-tocopherol) for both men and women, while the RDA for vitamin C is 90 mg for men and 75 mg for women. Notably, most athletes who are supplemented with dietary antioxidants are likely to meet or exceed these recommended intakes. However, it is important to note that the RDA may not necessarily be considered optimal for athletic populations [527]. Therefore, before considering dietary supplementation, it is recommended that individuals evaluate their diet and aim for a food-first approach, in which they consume foods rich in vitamins E and C to reap the benefits of these key antioxidants [3], as some data suggest that when one consumes substantially more than the RDA for these nutrients, the national training adaption response may be blunted [528]. For example, Ristow and associates [528] found that administering 400 IU/day of vitamin E and 1000 mg/day of vitamin C for 4 weeks to 39 untrained, healthy young men blunted PGC1-α induction and subsequent mitochondrial biogenesis, as well as key endogenous antioxidant enzymes. Therefore, if dietary supplementation is warranted, it is essential to consider the sport in which an individual is engaged and their training status.
To date, numerous studies [488,528–547] have investigated the effects of vitamin E, with or without vitamin C, on exercise and sports performance, as have studies [548–556] examining vitamin C alone. Following various supplementation protocols, most studies have reported no effect [488,529,531,533,534,536,540,542,543,545,549,553,554] on performance outcomes, with some reporting unfavorable outcomes (i.e. blunted training response) [528,530,551]. Furthermore, Nikolaidis et al. [557] conducted a review of 10 investigations on the ergogenic benefits of vitamin E and/or C supplementation during chronic exercise and subsequent adaptations, noting that only 2 of the 10 studies showed an ergogenic effect, while another 2 showed an ergolytic effect. In contrast, the remaining six studies reported no effect. Notably, in some cases, reports have also demonstrated no negative impact on the training response. For example, Zoppi et al. [546] assessed the effects of vitamin E (800 mg) and C (1000 mg) supplementation (divided into four daily doses) on performance, inflammation, and oxidative stress among male soccer players during a three-month pre-season training period. These findings demonstrated that the group supplementing with Vitamins E and C, compared to the placebo group, had reduced lipid peroxidation and muscle damage without hindering training adaptations. The most important consideration for individuals in regard to vitamins E and C is a well-balanced diet. For athletes following a standard American diet, supplementation with vitamins E or C may be appropriate, as dietary factors (e.g. a Western diet) can lead to relatively low vitamin stores, which can increase muscular fatigue and soreness. Therefore, supplementation can be promising in this context [558].
Zinc
11.24
Zinc, an essential trace element, plays a crucial role in endogenous antioxidant defense and redox homeostasis through both direct and indirect mechanisms, despite not being redox-active itself. One of the most well-established roles of zinc is serving as a cofactor for antioxidant enzymes, particularly copper–zinc-SOD, which is responsible for catalyzing the dismutation of O_2_^·–^ [559]. A study by Ribeiro and colleagues [560] revealed that 70 mg/day of zinc over 16 weeks in patients undergoing chemotherapy for colorectal cancer led to higher SOD activity during chemotherapy compared with placebo, demonstrating the indirect antioxidant role of zinc. Zinc is also a membrane stabilizer that protects cellular structures by maintaining sulfhydryl groups and displacing redox-active metals such as iron and copper, thereby reducing their pro-oxidant activity [561]. Zinc has been shown to induce the expression of metallothionein, a zinc-binding protein with potent free radical-scavenging capacity [562]. At the molecular level, zinc can activate the Nrf2 pathway, thereby enhancing the expression of endogenous antioxidant genes [563]. In the context of exercise and sports performance, zinc supplementation at 5 mg/kg/day for 4–6 days/week increases the levels of GSH, GPx, and SOD while reducing the level of MDA [564]. A meta-analysis of 21 RCTs involving 1321 participants further supports zinc’s ability to reduce biomarker concentrations of oxidative stress and inflammation, such as CRP, TNF-α, and MDA [565]. Collectively, these findings suggest that zinc can support endogenous antioxidant defense; however, more work is warranted among athletic populations, with exercise and sports performance outcomes at the forefront of assessment to better understand the application of zinc supplementation in sports.
Purported antioxidants: emerging or very limited evidence to support efficacy
11.25
Several nutrients and compounds are claimed to have antioxidant properties. However, it is important to recognize that most of these studies lack extensive pre-clinical data and have not yet been evaluated in randomized human clinical trials. Therefore, while acknowledging that these nutrients may offer some benefits to consumers, it is also clear that, like most dietary antioxidants with currently low efficacy evidence, these claimed antioxidants are not generally recommended and need further research. Table 3 lists these purported antioxidants and their respective mechanisms of action.
Section summary
11.26
There are numerous dietary antioxidants that athletes or performance professionals may consider leveraging in the context of exercise and sports; however, it is paramount that these individuals evaluate the data supporting these nutrients. There are a few “top-tier” dietary antioxidants with strong evidence supporting their efficacy for performance and recovery-related outcomes.
Special considerations
12
Elderly and masters athletes
12.1
Master’s athletes are generally defined as individuals over 35 or 40 years of age, but categories and age cutoffs vary by sport. Recently, increasing attention has been given to master’s athletics because this unique subgroup of adults is interested in optimizing their athletic performance while reducing the risk of disease burden, as they face physiological challenges associated with aging, including increased oxidative stress, inflammation, and anabolic resistance. While regular exercise can help maintain redox homeostasis [566–568], aging can independently increase ROS/RNS production [569,570], increasing the susceptibility of individuals to oxidative damage and, thus, poor health, performance, and recovery outcomes [568,571]. Given this, dietary antioxidant supplementation offers a potential solution to support these individuals in their respective training settings while also helping combat some of the natural processes of aging. Currently, research indicates that master’s athletes may not have adequate dietary antioxidant intake to support muscle function, immune health, and recovery [8]. Notably, although numerous studies have reported that redox and performance benefits from antioxidant supplementation, the overall evidence remains mixed. Some high-dose antioxidant studies have shown no effect, ergolytic effects, impaired performance, blunted training adaptations, or reduced endogenous antioxidant signaling [488]. As such, an antioxidant-rich food-first approach should be employed. Any complementary course of antioxidant supplementation should be tailored to specific scientific evidence, such as that presented herein. Interestingly, Guo and colleagues [572] recently conducted a systematic review of 26 studies encompassing 2819 master’s athletes and found that these individuals tend to consume more micronutrients than the general public. However, they may benefit from certain dietary antioxidant supplementation strategies, such as ω-3 fatty acids and polyphenols.
Tactical and occupational athletes
12.2
Tactical and occupational athletes are negatively affected by occupation-specific stressors, which can increase exposure to oxidative stress, inflammation, and increased disease risk [573]. There are several reasons for occupation-induced oxidative stress and inflammation, including intense physical exertion, psychological stress, shift work, poor dietary habits, traumatic experience, and other occupation-specific pressures (i.e. deployment and family separation) [574]. These findings highlight the key reasons why tactical or occupational athletes may consider supplementing with dietary antioxidants. However, to date, data are limited, and the results are mixed. For instance, Gonzalez and colleagues [189] assessed the impact of four weeks of AST (12 mg/day) supplementation among 15 career male firefighters and found that participants increased their ventilatory anaerobic threshold by 8.8%. In addition, they found that the firefighters showed lower physiological, oxidative, and inflammatory biomarker responses to simulated firefighting tasks after supplementation with AST. Another study by McAllister et al. [575] found that an acute dose of curcumin (i.e. a single dose of 1.5 g of curcumin/69 mg of curcuminoids) did not impact the oxidative stress response or physical performance during a simulated live burn, victim search, and rescue exercise. Regardless, tactical and occupational athletes need to identify pragmatic strategies to improve their performance, health, and recovery. While dietary antioxidants could benefit these populations, the data are limited.
Sex differences
12.3
It has been suggested that more than half of collegiate female athletes use traditional and non-traditional dietary supplements at least once a month to gain a competitive edge [576,577]. Previous reports have demonstrated sex-based differences in antioxidant status and responses to dietary antioxidant intake [577,578], with biological differences in hormonal regulation, body composition, and metabolism also likely affecting how men and women respond to increased free radical production and antioxidant intake [577,578]. Estrogen, for instance, exhibits intrinsic antioxidant properties and may enhance endogenous defense systems by modulating the expression and activity of key antioxidant enzymes, such as SOD and GPx [579]. It seems that females may produce less ROS and have higher antioxidant enzyme activity than males; however, this is not conclusive, as few studies have shown a uniform consensus or differences in several key antioxidant enzymes [578]. There is some evidence that men may derive greater protective benefits from dietary antioxidants, particularly as measured by the Composite Dietary Antioxidant Index (CDAI), compared to women [580]. However, it remains unclear whether these sex-based differences are consistent across women with different reproductive or menopausal statuses, as fluctuations in estrogen may substantially influence antioxidant defenses. Nevertheless, Duan et al. [580] assessed data from adults in the National Health and Nutrition Examination Survey (NHANES) cycles spanning 2001–2018 and found that the CDAI (calculated by summing the standardized intake values of six antioxidants: vitamin A, vitamin C, vitamin E, zinc, selenium, and total carotenoids) was negatively associated with all-cause mortality exclusively in men. The authors noted that estrogen may play a role by activating antioxidant pathways, thereby reducing the benefits of exogenous antioxidants in this context [578,579,581–583]. A 2023 ISSN position stand [584] highlights several dietary supplementation strategies for female athletes and women engaged in exercise (i.e. resistance or aerobic) training, including several dietary antioxidants, such as nitrates and CrM. These dietary supplements were found to be efficacious in the available literature for female populations, based on physiological theory and sex physiology [584].
Mitigating traumatic brain injury
12.4
Traumatic brain injuries (TBIs) are a major public health issue that can result in disability or premature mortality [585–588]. Daneshvar et al. [589] have suggested that an estimated 1.7 million individuals have sustained a TBI annually, with other reports indicating that over 69,000 TBI-related deaths occurred in the US in 2012 [585]. It is important to note that concerns about TBIs in sport have persisted for decades [589,590], with reports by Powell and Barber-Foss [591] suggesting that as many as 1219 mild TBIs were reported across various high school sports in 1999. Nowadays, the Centers for Disease Control have estimated that between 1.6 and 3.8 million concussions occur in sports and recreational activities annually [589]. This is also a major concern for tactical and occupational populations. For instance, approximately 500,000 US service members have sustained TBIs within the past 20 years [587]. Strack et al. [592] also noted that, in a sample of 1112 firefighters, 66% had reported at least one head injury in their lifetime, which was associated with higher levels of post-traumatic stress disorder and depression symptoms when compared to those who had not suffered a head injury. It is clear that TBIs are an issue, and these reports do not account for those individuals who suffer from TBI but do not seek medical attention [589]. Given that TBIs pose challenges and that sustaining a TBI induces an increased state of oxidative stress/redox imbalance, identifying avenues for the prevention and treatment of these injuries has led to interest in TBI-specific supplementation with select nutraceuticals and antioxidant compounds [586]. A 2024 review by Conti and colleagues [586] noted the specific supplementation recommendations for CrM (i.e. 4 × 5 g/day), ω-3 s (2–4 g/day with 2 g from DHA), choline (1–2 g/day), magnesium (400 md/day), NAC (4 g/day for 7 days followed by 3 g/day thereafter) and berry anthocyanins (250–400 mg/day), alongside food sources that should be considered for an individual needing to follow a nutritional and/or supplementation protocol for prevention and/or treatment for TBIs.
Conclusion and position of the International Society of Sports nutrition
13
Dietary antioxidants play a complex, context-dependent role in exercise and sport, striking a delicate balance between supporting recovery and potentially blunting physiological adaptations (e.g. with high-dose supplementation). While endogenous antioxidant systems are critical for maintaining redox balance, strategically increasing dietary antioxidants, preferably from whole foods, can enhance defense against excessive oxidative stress, especially during periods of high training load or inadequate recovery. However, the timing, dosage, and type of antioxidant compound must be carefully considered, as their effects vary according to training goals, individual physiology, and environmental demands. Ultimately, a personalized, evidence-based approach that acknowledges both the hormetic nature of exercise-induced oxidative stress and the complex biology of antioxidant action offers the greatest potential to optimize health and performance outcomes. Based on a comprehensive review and critical analysis of the literature regarding antioxidants conducted by experts in the field and selected members of the International Society of Sports Nutrition (ISSN), the following conclusions represent the official position of ISSN:
- Redox balance exists on a spectrum, with mild oxidative eustress driving beneficial adaptations, while excessive distress impairing recovery and performance.
- Exercise-induced ROS can support adaptation at moderate levels but may cause muscle damage, inflammation, and reduced endurance when excessive.
- Antioxidants from endogenous systems and dietary sources protect cells by neutralizing free radicals and limiting oxidative damage to key biomolecules.
- FDA labeling for “antioxidant” claims applies to nutrients with established RDIs and demonstrated antioxidant activity; this typically includes vitamins C and E, β-carotene (a source of vitamin A), selenium, zinc, copper, and manganese.
- Several dietary compounds exhibit antioxidant activity, showing potential for both direct and indirect antioxidant effects. However, the strength of evidence varies, and their use should be tailored to align with specific performance or recovery goals.
- Regular exercise improves endogenous antioxidant defenses and should be the primary strategy for enhancing redox capacity before considering supplementation.
- Whole foods rich in flavonoids, polyphenols, carotenoids, vitamins, and minerals are the preferred antioxidant sources, with supplements used to fill dietary gaps.
- Supplementation is best reserved for nutrient insufficiencies and deficiencies, inadequate dietary intake, or periods of high stress, while chronic high-dose use may blunt training adaptations.
- Responses to supplementation vary by individual factors such as training status, baseline antioxidant capacity, demographics, diet, and injury risk, with some compounds offering cognitive, behavioral, or trauma-related benefits in specific populations.
- Supplements such as creatine (i.e. 0.1 g/kg/day), omega-3 fatty acids (1000–6000 mg/day EPA+DHA for 6–12 weeks), tart cherry (480 mg powder or 60–90 mL juice/day for 7–14 days), and astaxanthin (4–12 mg/day for 4–12 weeks) rank among the top nutrients for their antioxidant effects, with moderate- to high-quality evidence supporting their use in recovery or performance without interfering with training adaptations. Most others exhibit weak or mixed data; therefore, selection should be tailored to training goals, biology, and the strength of the evidence.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Mason SA, Trewin AJ, Parker L, et al. Antioxidant supplements and endurance exercise: current evidence and mechanistic insights. Redox Biol. 2020 Aug;35:101471. doi: 10.1016/j.redox.2020.10147132127289 PMC 7284926 · doi ↗ · pubmed ↗
- 2Martinez-Ferran M, Sanchis-Gomar F, Lavie CJ, et al. Do antioxidant vitamins prevent exercise-induced muscle damage? A systematic review. Antioxidants. 2020;9(5):372. doi: 10.3390/antiox 905037232365669 PMC 7278664 · doi ↗ · pubmed ↗
- 3Higgins MR, Izadi A, Kaviani M. Antioxidants and exercise performance: with a focus on vitamin E and C supplementation. Int J Environ Res Public Health. 2020 Nov 15;17(22):8452. doi: 10.3390/ijerph 1722845233203106 PMC 7697466 · doi ↗ · pubmed ↗
- 4Clemente-Suárez VJ, Bustamante-Sanchez Á, Mielgo-Ayuso J, et al. Antioxidants and sports performance. Nutrients. 2023 May 18;15(10):2371. doi: 10.3390/nu 1510237137242253 PMC 10220679 · doi ↗ · pubmed ↗
- 5Sen CK. Oxidants and antioxidants in exercise. J Appl Physiol. 1995;79(3):675–686. doi: 10.1152/jappl.1995.79.3.6758567503 · doi ↗ · pubmed ↗
- 6Powers SK, Jackson MJ. Exercise-induced oxidative stress: cellular mechanisms and impact on muscle force production. Physiol Rev. 2008 Oct;88(4):1243–1276. doi: 10.1152/physrev.00031.200718923182 PMC 2909187 · doi ↗ · pubmed ↗
- 7Powers SK, Deminice R, Ozdemir M, et al. Exercise-induced oxidative stress: friend or foe? J Sport Health Sci. 2020 Sep;9(5):415–425. doi: 10.1016/j.jshs.2020.04.00132380253 PMC 7498668 · doi ↗ · pubmed ↗
- 8Maughan RJ, Burke LM, Dvorak J, et al. IOC consensus statement: dietary supplements and the high-performance athlete. Br J Sports Med. 2018 Apr;52(7):439–455. doi: 10.1136/bjsports-2018-09902729540367 PMC 5867441 · doi ↗ · pubmed ↗