The complete mitochondrial genome and phylogenetic analysis of scyphozoan jellyfish Chrysaora chinensis (Cnidaria: Scyphozoa: Pelagiidae) in the coastal sea of Fangchenggang, China
Tong Su, Jie Guo, Yutong Xie, Bingfu Tan, Mingben Xu, Liangliang Huang, Junxiang Lai

TL;DR
This paper reports the first complete mitochondrial genome of the jellyfish Chrysaora chinensis and uses it to explore its evolutionary relationships with other jellyfish species.
Contribution
The study provides the first complete mitogenome of Chrysaora chinensis and reveals its close phylogenetic relationship with other Chrysaora species.
Findings
The mitogenome of C. chinensis is 16,783 bp long with 69% AT content and includes 13 PCGs, 2 rRNA genes, 2 tRNA genes, and an ORF.
The gene order is similar to other jellyfish species, and phylogenetic analysis shows C. chinensis is closely related to C. pacifica and C. quinquecirrha.
The findings offer insights into the taxonomy and evolution of scyphozoan jellyfish.
Abstract
The scyphozoan jellyfish Chrysaora chinensis has been frequently observed in the coastal waters of Fangchenggang, China, in recent years. This study assembled and analyzed the first complete mitochondrial genome (mitogenome) of C. chinensis from next-generation sequencing data. The mitogenome was 16,783 bp in length, exhibited an overall AT content of 69%, and encoded 13 typical protein-coding genes (PCGs), 2 ribosomal RNA (rRNA) genes (rrnS and rrnL), 2 transfer RNA (tRNA) genes (trnW-tca and trnM-cat), and an additional open reading frame (ORF), dpo. The mitochondrial gene order was largely consistent with that of previously reported jellyfish species. Furthermore, phylogenetic analysis of 13 mitochondrial PCGs from 19 jellyfish species indicated that Chrysaora chinensis is closely related to Chrysaora pacifica and Chrysaora quinquecirrha. These results provided a valuable reference…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
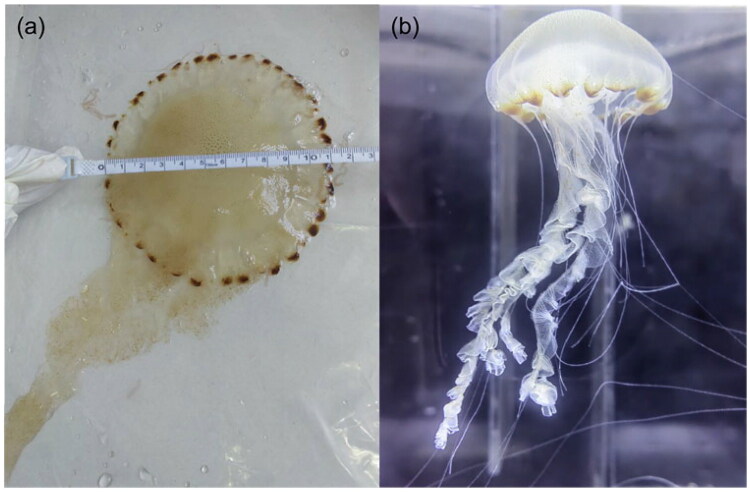 Figure 1
Figure 1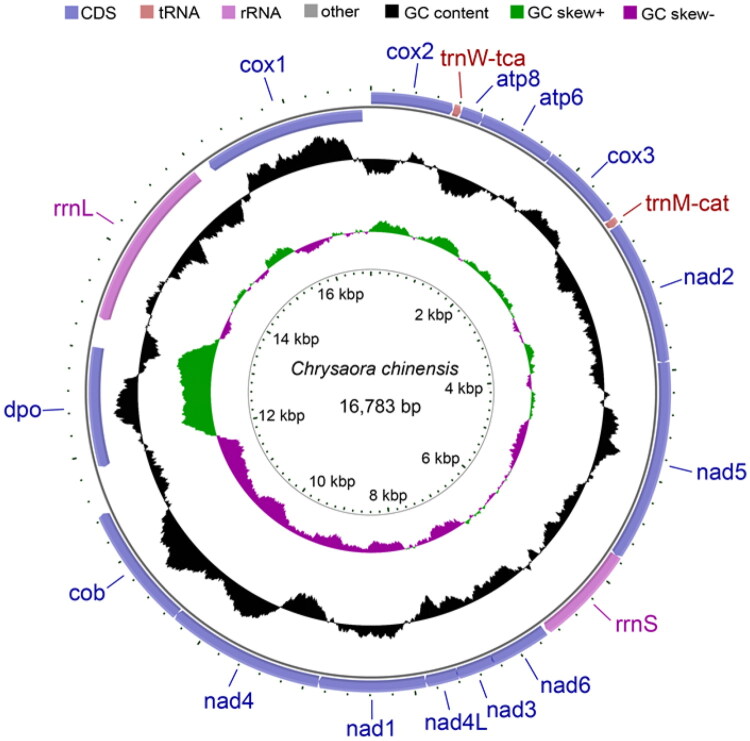 Figure 2
Figure 2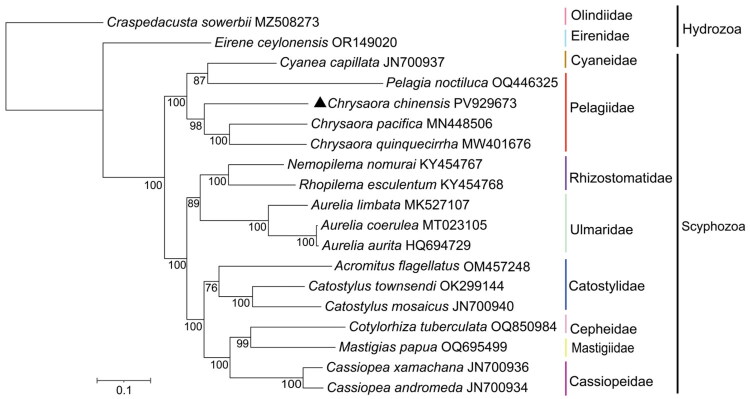 Figure 3
Figure 3| Species name | Accession number | Length | Order | Reference |
|---|---|---|---|---|
|
| 16,783 bp | Semaeostomeae | This study | |
|
| 16,964 bp | Semaeostomeae | (Wang and Yin, | |
|
| 16,875 bp | Semaeostomeae | unpublished | |
|
| 16,390 bp | Semaeostomeae | (Lee and Ki, | |
|
| 16,202 bp | Semaeostomeae | (Kayal et al. | |
|
| 16,971 bp | Semaeostomeae | (Park et al. | |
|
| 16,748 bp | Semaeostomeae | (Seo et al. | |
|
| 16,953 bp | Semaeostomeae | (Karagozlu et al. | |
|
| 17,024 bp | Rhizostomeae | (Wang and Sun, | |
|
| 15,855 bp | Rhizostomeae | (Wang and Sun, | |
|
| 16,779 bp | Rhizostomeae | (Lin et al. | |
|
| 16,308 bp | Rhizostomeae | unpublished | |
|
| 15,949 bp | Rhizostomeae | (Kayal et al. | |
|
| 15,800 bp | Rhizostomeae | (Kayal et al. | |
|
| 16,590 bp | Rhizostomeae | (Jiang et al. | |
|
| 14,847 bp | Rhizostomeae | (Kayal et al. | |
|
| 16,560 bp | Rhizostomeae | (Xia et al. | |
|
| 18,020 bp | Limnomedusae | unpublished | |
|
| 14,997 bp | Leptothecata | (Chen et al. |
- —Guangxi Natural Science Foundation10.13039/501100004607
- —Basic Research Funds Project of Guangxi Academy of Sciences
- —Guangxi Special Fund Project for Local Science and Technology Development Guided by the Central Government
- —Guangxi Young Talents Development Program Guangxi Young Talents Development Program
- —Guangxi Mangrove Coastal Wetland Ecological Protection and Sustainable Utilization Small Highland Talent Support Project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMarine Invertebrate Physiology and Ecology · Aquatic Ecosystems and Phytoplankton Dynamics · Marine Sponges and Natural Products
Introduction
In recent years, jellyfish blooms have become increasingly frequent in coastal waters worldwide (Dong et al. 2010; Kennerley et al. 2022; Suárez et al. 2022). Such blooms repeatedly damage marine ecosystems and coastal economies, and also endanger human health and safety (Purcell et al. 2007; Wang et al. 2023; Osathanunkul, 2024). The scyphozoan jellyfish Chrysaora chinensis Vanhöffen, 1888 (Cnidaria: Scyphozoa: Semaeostomeae: Pelagiidae), first discovered in the South China Sea near Hong Kong, is named for its type locality (Morandini and Marques, 2010). It is widely distributed in the Western Pacific Ocean and has been observed off the coasts of Indonesia, Sumatra, China, and the Philippines (Morandini and Marques, 2010). Notably, this species has been frequently observed in the waters near Fangchenggang, Qinzhou Bay, China, over the past two years. The jellyfish within the genus Chrysaora, commonly referred to as sea nettles, are characterized by umbrella-shaped bells that can reach several tens of centimeters in diameter, as well as numerous elongated tentacles equipped with nematocysts. They exhibit considerable adaptability to a range of water temperatures and salinities (Morandini and Marques, 2010). The safety of the cold source water intake at Fangchenggang Nuclear Power Plant may be compromised by large aggregations of C. chinensis. However, studies on C. chinensis have been primarily morphology-focused, with mitochondrial gene sequencing efforts concentrated on specific genes such as cox1 and 16S rRNA (Daglio and Dawson, 2017; Low et al. 2019).
At the class level, database and literature surveys combined show that only 29 scyphozoan mitogenomes—assigned to 22 nominal species (20 complete, 2 partial)—have been published to date (“Scyphozoa[Organism] AND mitochondrion[Title]”, NCBI, accessed 6 January 2026). As of now, the NCBI Nucleotide database contains two complete mitogenomes of Chrysaora chinensis: PV929673 (this study, assembled and released by us on 20 July 2025) and a later entry (PX261967). We used the first-available mitogenome of C. chinensis to reconstruct its phylogeny and provide genomic resources for Chrysaora systematics and broader insights into jellyfish evolution.
Materials and methods
Chrysaora chinensis was collected from the coastal waters of Fangchenggang (108°36′37.79″E, 21°37′51.96″N) in Qinzhou Bay, China, using a plankton net on 25 December 2024. The specimen was deposited at the Laboratory of Marine Environmental Science, Guangxi Academy of Sciences (Dr. Jie Guo, [email protected]) under voucher number Chrysaora2024-1201 (Figure 1). Species identity was verified by combining morphological analysis (Morandini and Marques, 2010; Low et al. 2019) and sequencing of the commonly used mitochondrial markers cox1 and 16S rRNA; the resulting sequences showed a high degree of similarity to GenBank accessions KY611280 and MF141690, respectively. From C. chinensis umbrella muscle, genomic DNA was extracted using the DNeasy Tissue Kit (Qiagen, Beijing, China). Prior to NovaSeq library construction, 1 µg of high-quality DNA was sheared to 300–500 bp using a Covaris M220 focused-ultrasonicator; short-insert libraries were then prepared and sequenced on the NovaSeq 6000 platform (BIOZERON Co., Ltd., Shanghai, China), yielding 150 bp paired-end reads. GetOrganelle v1.7.5 was used to assemble the mitogenome (Jin et al. 2020). Assembly completeness and coverage uniformity were assessed by read-depth profiling (Supplementary Figure S1). Mitogenome genes were annotated with MITOS2 (Bernt et al. 2013), and tRNA genes were predicted by tRNAscan-SE (Chan and Lowe, 2019). The complete mitogenome map was then visualized using CGView (Stothard et al. 2019).
Reference images of Chrysaora chinensis. (a) Close-up view of the umbrella (diameter approximately 11 cm) and oral arms (length approximately 26 cm). (b) Overall view of the jellyfish. These photographs were taken by the author Tong Su in December 2024 at the Laboratory of the Guangxi Academy of Marine Sciences, China.
A molecular phylogenetic reconstruction of the class Scyphozoa was performed using 19 mitochondrial genomes, comprising 17 scyphozoan species and 2 hydrozoan outgroups (Eirene ceylonensis and Craspedacusta sowerbii) specifically selected to enhance phylogenetic resolution (Table 1). The mitogenomes were retrieved from NCBI GenBank, including the newly sequenced mitogenome of C. chinensis generated in this study. The phylogeny was inferred from a concatenated nucleotide supermatrix of the 13 mitochondrial PCGs shared by most jellyfish species. These sequences were processed using Phylosuite v1.2.3, which included performing multiple sequence alignment with MAFFT, trimming ambiguous regions and individual columns exhibiting excessive gaps or ambiguities using trimAI and Gblocks, followed by concatenating the cleaned alignments (Zhang et al. 2020). A maximum-likelihood (ML) phylogenetic tree was constructed in MEGA 12 with 1,000 bootstrap replicates (Kumar et al. 2024), and subsequently visualized and manually refined using iTOL (https://itol.embl.de/).
Results
The complete mitogenome of C. chinensis spanned 16,783 bp and encoded 13 typical PCGs (cox1, cox2, atp8, atp6, cox3, nad2, nad5, nad6, nad3, nad4L, nad1, nad4, cob), 2 rRNA genes (rrnL and rrnS), 2 tRNA genes (trnW-tca and trnM-cat) and 1 additional ORF (dpo) (Figure 2). Immediately downstream of rrnL, the 1,161-bp dpo was predicted to encode a 386-amino-acid protein with extensive similarity to family B DNA polymerases. All PCGs were located on the H-strand, and nearly all were in the positive direction, with the exception of rrnL, cox1 and dpo, which were in the negative direction. Regarding the start and stop codons, the PCGs initiated with ATA (nad3, dpo), GTG (nad4), TTA (cox1), or ATG (all remaining genes), and terminated with TAG (cox3, nad5, nad6, dpo) or TAA (the rest). The mitogenome showed a pronounced AT bias, with an overall base composition of 69.00% AT (A: 31.67%, T: 37.33%) and 31.00% GC (G: 15.99%, C: 15.01%). The 13 typical PCGs occupied 11,976 bp (71.36% of the mitogenome); the predicted rrnS (928 bp) and rrnL (1,686 bp) were located between nad5–nad6 and dpo–cox1, respectively.
Circular mitogenome map of Chrysaora chinensis from Fangchenggang coastal waters.
Before phylogenetic inference, we updated the NCBI search (6 January 2026). Subsequent retrieval of a newly deposited mitogenome (PX261967, submitted 7 weeks after ours) revealed 99.77% identity across the full length relative to our assembly, consistent with intraspecific sequence variation; it was therefore excluded from phylogenetic analyses to avoid redundancy. Maximum likelihood analysis revealed two robust clades within Scyphozoa (Figure 3). One clade included six families (Rhizostomatidae, Ulmaridae, Catostylidae, Cepheidae, Mastigiidae, and Cassiopeidae), while the other consisted of Cyaneidae and Pelagiidae. The mitogenome of C. chinensis was sister to the clade formed by C. pacifica and C. quinquecirrha, together constituting a distinct Chrysaora lineage.
Maximum-likelihood phylogenetic tree of 19 jellyfish species based on the 13 protein-coding genes.
Discussion and conclusions
In this study, we sequenced and characterized the complete 16,783-bp mitogenome of Chrysaora chinensis using high-throughput Illumina sequencing, providing the first comprehensive account of its full-length sequence, gene content and base composition. Gene order and content were highly similar to those reported for other scyphozoan jellyfish (Seo et al. 2020; Wang and Yin, 2020; Jiang et al. 2024). The distribution of genes, predominantly on the H-strand and in the positive direction except for rrnL, cox1 and dpo, may influence the essential replication and transcription processes fundamental to mitochondrial function (Tzagoloff and Dieckmann, 2021). In this study, we predicted a 1,161-bp ORF (dpo) immediately downstream of rrnL in the C. chinensis mitogenome; the deduced amino-acid sequence displayed extensive similarity to family B DNA polymerases. An ORF of identical length at the syntenic position is present in the previously published mitogenome of Craspedacusta sowerbyi (Zou et al. 2012), and comparable ORFs have been reported in the scyphozoans Mastigias papua and Cotylorhiza tuberculata (Xia et al. 2023; Jiang et al. 2024). Collectively, the occurrence of these dpo-like ORFs is thought to reflect linear-plasmid integration events (Kayal et al. 2012). Hydroidolinan hydrozoans, however, appear to have lost this ORF, suggesting lineage-specific evolutionary trajectories or alternative functional compensation mechanisms that warrant further experimental investigation (Kayal et al. 2012; Ladoukakis and Zouros, 2017). Nevertheless, within Cnidaria, the absence of dpo may also stem from annotation practices that tend to recognize only the typical set of 13 PCGs. The high sequence variability of dpo-like ORFs, together with limited scrutiny of non-core regions, could lead to their being overlooked.
Consistent with most cnidarians reported to date, C. chinensis possessed only two 2 rRNA genes (rrnL and rrnS) and 2 tRNA genes (trnW-tca and trnM-cat), The trnW-tca and trnM-cat) were 70 bp and 71 bp, respectively, and showed high conservation across cnidarians (Kayal et al. 2012; Zou et al. 2012). The RNA genes had a slightly higher AT content than the PCGs. Medusozoan mt-tRNAs exhibited greater primary-sequence and secondary-structure variability than their anthozoan and demosponge homologues (Wang and Lavrov, 2008). The consensus secondary structures inferred for Hydrozoa, Scyphozoa and Staurozoa revealed class-specific differences in acceptor-stem length, deletions within the D-arm/D-loop region and nucleotide losses (Kayal et al. 2012). In addition, available partial cubozoan mitochondrial sequences have been reported to lack tRNA genes (Kayal et al. 2012). Accordingly, these mt-tRNA variations provide comparative reference points that merit further testing in future class-level studies of Medusozoa.
Phylogenetic analysis of 13 typical PCGs from 19 jellyfish taxa revealed that C. chinensis clustered with C. pacifica and C. quinquecirrha, forming a clade that was genetically distinct from other taxa (Figure 3). Differences in their PCGs among these species may be related to geographical population isolation and site polymorphisms. The need for expanded mitochondrial genome data within the Pelagiidae family was evident, given the limited information available. In conclusion, this study provided a complete mitochondrial reference for C. chinensis, which will facilitate future phylogenetic, phylogeographic, and evolutionary studies of scyphozoans.
Supplementary Material
Supplemental material.docx
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bernt M et al. 2013. MITOS: improved de novo metazoan mitochondrial genome annotation. Mol Phylogenet Evol. 69(2):313–319. 10.1016/j.ympev.2012.08.02322982435 · doi ↗ · pubmed ↗
- 2Chan PP, Lowe TM. 2019. t RN Ascan-SE: searching for t RNA Genes in Genomic Sequences. Methods Mol Biol. 1962:1–14. 10.1007/978-1-4939-9173-0_131020551 PMC 6768409 · doi ↗ · pubmed ↗
- 3Chen Y et al. 2023. The complete mitochondrial genome and phylogenetic analysis of hydrozoan jellyfish Eirene ceylonensis (Cnidaria, Hydrozoa, Eirenidae) in the coastal sea of Qinhuangdao, China. Mitochondrial DNA B Resour. 8(12):1381–1385. 10.1080/23802359.2023.229084538189024 PMC 10768735 · doi ↗ · pubmed ↗
- 4Daglio LG, Dawson MN. 2017. Species richness of jellyfishes (Scyphozoa: discomedusae) in the Tropical Eastern Pacific: missed taxa, molecules, and morphology match in a biodiversity hotspot. Invertebr Syst. 31(5):635–663. 10.1071/IS 16055 · doi ↗
- 5Dong ZJ, Liu DY, Keesing JK. 2010. Jellyfish blooms in China: dominant species, causes and consequences. Mar Pollut Bull. 60(7):954–963. 10.1016/j.marpolbul.2010.04.02220553695 · doi ↗ · pubmed ↗
- 6Jiang H et al. 2024. Characterization and phylogenetic analysis of the complete mitochondrial genome of Cotylorhiza tuberculata assembled using next-generation sequencing. Mitochondrial DNA B Resour. 9(9):1268–1272. 10.1080/23802359.2024.240692839328354 PMC 11425685 · doi ↗ · pubmed ↗
- 7Jin JJ et al. 2020. Get Organelle: a fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 21(1):241. 10.1186/s 13059-020-02154-532912315 PMC 7488116 · doi ↗ · pubmed ↗
- 8Karagozlu MZ, Seo Y, Ki JS, Kim CB. 2019. The complete mitogenome of brownbranded moon jellyfish Aurelia limbata (Cnidaria, Semaeostomeae, Ulmaridae) with phylogenetic analysis. Mitochondrial DNA Part B Resources. 4(1):1875–1876. 10.1080/23802359.2019.1614494 · doi ↗