How flagellar glycosylation of the phytopathogenic bacterium Pseudomonas amygdali pv. tabaci 6605 affects transport and deposition in saturated sandy porous media
Xin Zheng, Mounia Achak, Edvina Lamy, Yannick Rossez

TL;DR
This study explores how flagellar glycosylation in a plant pathogen affects its movement and retention in sandy soil, offering insights into managing bacterial contamination in agricultural settings.
Contribution
The study reveals how flagellar glycosylation influences bacterial transport and deposition in porous media, providing new insights into microbial behavior in subsurface environments.
Findings
Glycosylated flagella increase bacterial mobility and reduce retention in sandy porous media.
Flagellar hydrophobicity enhances bacterial adhesion and deposition onto sand grains.
Flagellar glycosylation introduces hydrophilic glycans that counteract adhesion effects.
Abstract
To mitigate bacterial contamination in underground farmland, a comprehensive understanding of the transport and adhesion mechanisms of phytopathogenic bacteria in porous media is crucial for safeguarding soil and groundwater. This study aims to elucidate the effects of Pseudomonas amygdali pv. tabaci 6605 flagella (wild type, ΔfliC strain) and their glycosylation (Δfgt1 and Δfgt2 strains) on bacterial transport and deposition in sandy porous media through a combination of experimental observations and numerical simulations. Flagella play a key role in bacterial transport and deposition dynamics through its surface properties. Their intrinsic hydrophobicity enhances bacterial adhesion and promotes deposition onto sandy grains while simultaneously limiting transport through the porous medium. However, glycosylation of flagellin introduces hydrophilic glycans, which counteract this effect…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
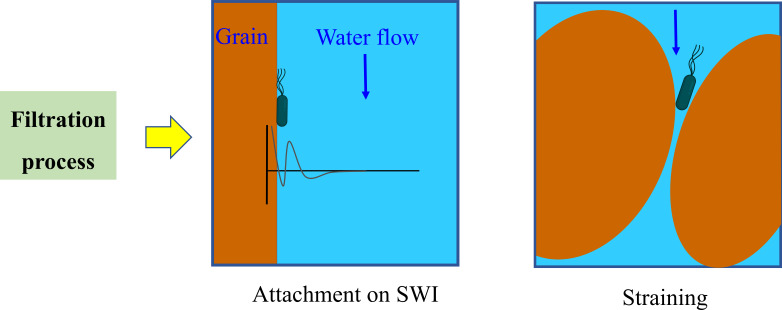 Fig 1
Fig 1 Fig 2
Fig 2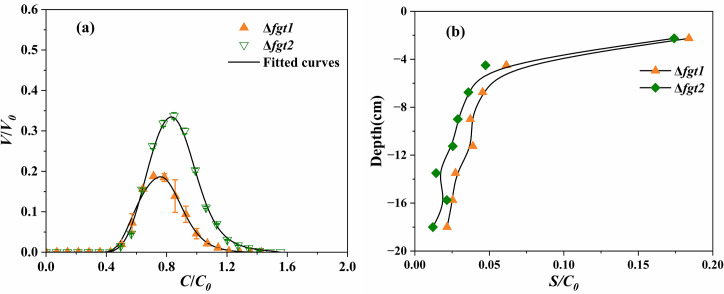 Fig 3
Fig 3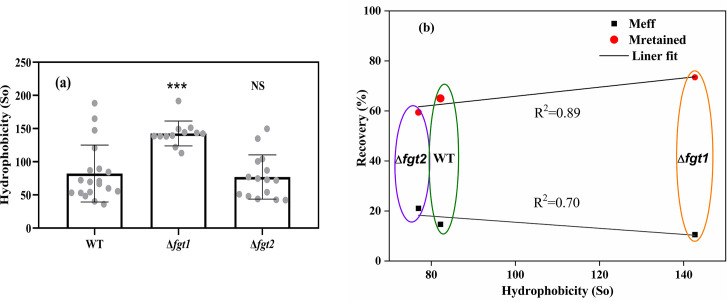 Fig 4
Fig 4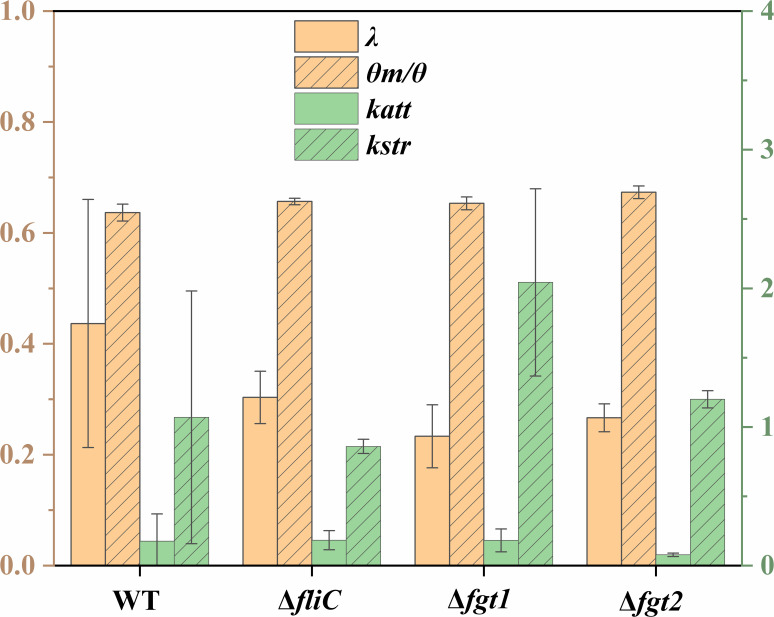 Fig 5
Fig 5| Flagellar phenotype | WT | Δ | Δ | Δ |
|---|---|---|---|---|
| Flagella | On | Off | On | On |
| Glycosylation | On | Off | Off | Partially on |
| Motility | On | Off | On | On |
| Bacterial strain |
| Recovery rate (%) | ||
|---|---|---|---|---|
|
|
|
| ||
| WT (with flagella) | 0.70 ± 0.06 | 14.68 ± 1.95 | 65.04 ± 1.01 | 79.72 ± 1.36 |
| Δ | 0.63 ± 0.01 | 29.24 ± 1.32** | 52.53 ± 0.85*** | 81.77 ± 2.15, NS |
| Δ | 0.64 ± 0.05 | 10.56 ± 1.07, NS | 73.46 ± 5.60, NS | 84.03 ± 4.81, NS |
| Δ | 0.69 ± 0.01 | 21.08 ± 0.70* | 59.45 ± 3.52, NS | 80.53 ± 4.12, NS |
- —China Scholarship Councilhttp://dx.doi.org/10.13039/501100004543
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFecal contamination and water quality · Bacterial biofilms and quorum sensing · Microbial Applications in Construction Materials
INTRODUCTION
The dissemination of phytopathogenic bacteria through subsurface environments such as soil and water is a critical step in plant infection but remains poorly understood. Numerous studies have investigated how environmental and bacterial factors influence transport and retention in porous media. These include physical parameters of the medium (1–3), fluid properties (4–6), and cellular characteristics such as morphology, size, and surface charge (7–10). Among these, flagellum-driven motility has emerged as a pivotal attribute influencing bacterial transport and attachment.
Bacterial flagella are complex structures composed of a rotary motor, hook, and a long extracellular filament that functions as a helical propeller (11). In addition to driving motility, flagellar filaments play a critical role in surface attachment, often serving as the first point of contact between the cell and a surface (12, 13). In many bacterial species, the filament is primarily composed of a protein called flagellin, encoded by the fliC gene. In Pseudomonas syringae, a known plant pathogen, flagellar motility and the expression of fliC are essential for virulence and efficient colonization (14–16). Beyond gene presence and expression, post-translational modifications (PTMs) of flagellin, particularly glycosylation and methylation, have been shown to modulate bacterial motility, adhesion, and host interactions. For instance, in Salmonella enterica serovar Typhimurium, methylation of lysine residues on flagellins enhances transport and adhesion to sandy surfaces under flow conditions (17). This finding raises the question of whether flagellar glycosylation, another widespread PTM, similarly influences bacterial behavior in hydrodynamic environments. Flagellin glycosylation, the covalent addition of sugars to amino acid residues, has been identified in numerous bacterial species and can affect protein stability, folding, immune recognition, and function (18–20). However, its role in bacterial transport through abiotic porous media under dynamic conditions remains largely unexplored.
To address this knowledge gap, we focused on Pseudomonas amygdali pv. tabaci 6605, formerly classified as P. syringae pv. tabaci (21), a phytopathogen responsible for wildfire disease in tobacco. This organism produces a single flagellin type (FliC), which is O-glycosylated with rhamnose-rich glycans and a terminal modified 4-amino-4,6 dideoxyglucosyl (Qui4N) residue, β-d-Quip4N(3-hydroxy-1-oxobutyl)2Me, commonly referred to as viosamine (mVio) (22). The biosynthesis of this glycan structure is regulated by two genes, fgt1 and fgt2. Deletion mutants of these genes produce non-glycosylated or partially glycosylated flagellin, respectively (23). Previous work has shown that loss of flagellar glycosylation in this strain reduces swarming motility and virulence (23, 24), suggesting a potential role in modulating surface interactions.
While protein glycosylation has been linked to altered bacterial adhesion to biotic surfaces (25–27), little is known about its role in bacterial transport and deposition onto abiotic surfaces under hydrodynamic conditions. This study aims to address that gap by assessing whether flagellar glycosylation affects bacterial transport and retention in porous media under flow. In this study, we examined how the presence and glycosylation state of flagella affect bacterial transport and retention in saturated flow through homogeneous sandy porous media. Using Pseudomonas amygdali pv. tabaci 6605 as a model, we compared the wild-type (WT) strain with three isogenic mutants: ΔfliC, which lacks flagellin and flagella; Δfgt1, which produces non-glycosylated flagella; and Δfgt2, which expresses partially glycosylated flagella. Column-scale transport experiments were conducted under controlled laboratory conditions, and breakthrough curves (BTCs) were generated to assess bacterial movement through the porous medium. These BTCs were analyzed using the Hydrus-1D code with a mobile–immobile (MIM) model to evaluate how differences in flagellum structure and modification affect bacterial transport and filtration dynamics. In addition to genetic and structural differences, we also investigated the role of flagellar surface hydrophobicity, a property influenced by glycan composition, on bacterial deposition. By integrating experimental observations with hydrodynamic modeling, this study provides new insights into the contribution of flagella and flagellar glycosylation to bacterial motility, adhesion, and environmental dissemination in dynamic soil and water systems.
MATERIALS AND METHODS
Bacterial strains
The P. amygdali pv. tabaci strains used in this study (Table 1) include the WT, a ΔfliC mutant lacking flagella, and two glycosyltransferase mutants: Δfgt1, which produces non-glycosylated flagella, and Δfgt2, which expresses partially glycosylated flagella. In the WT, the flagellin polymer is O-glycosylated with glycans composed of two or three rhamnose residues and a terminal mVio, attached to six serine residues (positions 143, 164, 176, 183, 193, and 201) (22).
Bacterial suspension preparation
P. amygdali pv. tabaci strains were inoculated on a King’s medium B base broth (KB) agar at 28°C. Then, these strains were grown at 28°C at 85 rpm in KB. Twenty grams of proteose peptone, 1.5 g K_2_HPO_4_, 15 mL glycerol, and 1.5 g MgSO_4_·7H_2_O were needed for preparing 1 L of KB; 20 g agar was required additionally to make KB agar. MgSO_4_·7H_2_O (1.5 g) was added after autoclaving. Bacterial suspension harvested from the stationary phase of bacterial culture was used in this work to avoid bacterial growth during experiments. The numbers of initial and final bacteria were determined by the number of colony-forming units (CFU) using the plating method on KB agar as described previously (17).
Flagellum purification and surface hydrophobicity measurement
The flagella were purified (12, 28), and the surface hydrophobicity was measured as described previously (29). In short, P. amygdali pv. tabaci strains were cultured in KB media at 28℃ for 36 h in an orbital shaker incubator at 80 rpm, harvested by centrifugation, then sheared off using a magnetic stirrer for 1h, followed by centrifugation to collect the supernatant containing the flagella. Ammonium sulfate was slowly added with vigorous stirring until two-thirds saturation was achieved. The flagella were harvested by centrifugation at 15,000*× g* for 20 min under 4℃ after 36 h incubation. After that, the precipitation was re-suspended in Tris-buffered saline. The surface hydrophobicity (So) of purified flagella using PRODAN (30). Eight microliters of 1 mM of PRODAN (prepared in DMSO) solution was added to 1 mL flagellum samples, which were diluted in 20 mM HEPES and 150 mM NaCl (pH = 7.4). The samples were incubated for 10 min in the dark, and the fluorescence intensity was measured with a Cary Eclipse spectrofluorometer. The wavelengths of excitation and emission were 365 and 465 nm, respectively, with corresponding 5 and 5 nm slit widths. The surface hydrophobicity values were determined using at least four measures per sample repeated three times, and the mean value was used.
Column experiments and numerical simulations
A plexiglass column with an inner diameter of 3.4 cm and a length of 18 cm was used for the transport experiments. Homogeneous Fontainebleau sand, with particle sizes ranging from 0.16 to 0.79 mm (D50 = 0.36 mm), was used as the porous medium in this study. The bulk density, porosity, and degree of saturation of the columns were determined by gravimetric analysis (see Material S1). A 20 mL bacterial suspension was injected into the columns, and BTCs were generated by plotting the time-dependent bacterial concentration in the effluent. Moment analysis was used for estimating the retardation factor (R) and mass balance from the effluent (Meff) (31):
Here, is the zero-order moment of BTCs; is the first-order moment of BTCs; C(t) is the time-dependent concentration of the effluent; and C0 is the initial concentration of bacteria. Meff is calculated by the ratio of the bacterial mass recovered at the column outlet to their mass injected at the column inlet (32):
Here, is the time for injection of bacterial suspension into the column. R was determined by the ratio of resident time for bacteria to the theoretical solution resident time:
Here, is the column length (cm); is the total water content; and is the Darcy velocity (cm/min).
After the transport experiments, the porous medium inside the column was sectioned into eight successive layers of 2.25 cm to analyze the spatial distribution of bacteria. Each sandy layer was transferred into a flask containing 0.1 mmol/L NaCl solution and shaken on a rotary shaker to detach bacteria from the sand grains. The plating method was used to quantify the bacterial biomass recovered from the retention profiles (RPs), denoted as Mretained. The total recovery of bacteria (Mtotal) is the sum of Meff and Mretained.
The modified two-region MIM model, incorporating two kinetic deposition sites as previously described (2, 17), was employed in this study. In the MIM model, the liquid phase is conceptualized as two distinct regions: a mobile (flowing) region and an immobile (stagnant) region. The model assumes that bacteria are excluded from the immobile regions and that no exchange occurs between the mobile and immobile zones. Column transport experiments in this study were conducted under saturated water conditions. Under these conditions, the two deposition sites were interpreted as bacterial attachment to solid–water interfaces and irreversible straining, both considered key mechanisms of bacterial retention in porous media (Fig. 1) (33). A reversible detachment process was also considered. The mass transfer between the liquid and solid phases is described by equation 6 (34):
Mechanisms of bacterial deposition in columns under saturated conditions include two primary patterns: physicochemical attachment to solid–water interfaces (SWIs), which allows for potential reversible detachment, and physical straining, which leads to irreversible retention.
Here, ρ is the bulk density of the porous media (g/cm^3^); S is the bacterial concentration in solid phase (CFU/g); s1 and s2 are the bacterial concentrations in solid phase accounting for attachment or irreversible straining (CFU/g), respectively; θ_m_ is the mobile water content; C is the bacterial concentration in the liquid phase (CFU/cm^3^); t is time (min); x is the distance (L); katt is the first-order attachment coefficient (1/min); k_d_ is the first-order detachment coefficient (1/min); kstr is the first-order straining coefficient (1/min); Ψ_t_ and Ψ_x_ describe the deposition of bacteria under time- and depth-dependent depositions, respectively.
Statistical analysis
The statistics were analyzed for significance using Student’s t-test by SPSS 26.0. Origin 8.1 was used for graphic drawing.
RESULTS
Flagellin expression reduces bacterial mobility and promotes retention within porous media
P. amygdali pv. tabaci WT (with flagella) and ΔfliC strains (without flagella) were used in this study to investigate the effects of flagella on bacterial transport and deposition in a sandy medium. Column experiments were conducted in triplicate, and average BTCs (Fig. 2a) as well as RPs (Fig. 2b) displayed a good repeatability for each strain. The experimental parameters are listed in Table 2. Experimental conditions for all replicates are shown in Material S2.
Observed (symbols) and fitted (curves) BTCs (a) and RPs (b) of P. amygdali pv. tabaci WT (with flagella) and ΔfliC (without flagella) strains. V is the volume occupied by water in the column; V0 is the total pore volume of the column; and C, C0, and S are the time-dependent, initial, and solid phase concentrations of bacteria, respectively.
Both WT and ΔfliC strains showed a symmetrical shape on BTCs (Fig. 2a), although the BTCs of the ΔfliC strain exhibited a more symmetrical shape compared to the BTCs of WT. The peak of BTCs for both strains occurred before 1 V/V0 (pore volume), indicating a preferential transport of bacteria.
The ΔfliC strain exhibited faster transport than the WT strain, as indicated by its lower retardation factor (0.63 compared to 0.70 for WT, Table 2) and earlier breakthrough times. Different recovery rates from effluent and retention profiles were obtained for WT and ΔfliC mutants (Table 2). The average recovery of effluent for WT was 14.68% and 29.24% for ΔfliC strains, in agreement with the BTC behavior, with a higher peak for ΔfliC strains. Logically, WT was more retained in the sand (65.04%) than the ΔfliC strain (52.53%). A similar total recovery (effluent + profiles) was obtained for both strains, with lower total recovery for the WT strain (79.72%), while the ΔfliC strain reached 81.77%. The RP replicates of WT and ΔfliC strains (Fig. 2b) displayed a non-monotonic distribution behavior with a high number of attached bacteria at the column inlet (layer 0–2.25 cm). WT exhibited higher dispersion in deeper layers (12–18 cm) than ΔfliC strains.
Glycosylation of flagella promotes bacterial transport through the sand, resulting in higher effluent recovery and decreased retention
To explore the influence of flagellar glycosylation on bacterial transport and deposition, Δfgt1 (without glycosylation) and Δfgt2 (partially glycosylated) strains were observed and compared in regard to BTCs and RPs, together with WT. BTCs and RPs of all strains showed a good repeatability (Fig. 3).
Observed (symbols) and fitted (curves) BTCs (a) and RPs (b) of P. amygdali pv. tabaci Δfgt1 (flagella without glycosylation) and Δfgt2 (flagella partially glycosylated) strains.
All replicates showed a symmetrical shape on BTCs (Fig. 3a). The Δfgt1 strain lacking flagellar glycosylation exhibited a faster transport than the WT and Δfgt2 (partially glycosylated) strain. This was confirmed by the same start of BTCs but earlier end of BTCs. The same result was also illustrated by a lower retardation factor of the Δfgt1 strain (0.64, Table 2) compared to the WT (0.7) and Δfgt2 strain (0.69). Although no obvious difference was observed for the BTCs between WT and Δfgt2 strain, which displayed almost the same starts and ends of BTCs, the peak of the Δfgt2 strain was higher than the WT, suggesting higher recovery of this strain in the effluent, confirmed by higher Meff (Table 2).
Even though the Δfgt1 exhibited a faster transport, this strain had the lowest recovery in the effluent (10.56%, Table 2) in comparison to the WT (14.68%) and the Δfgt2 strain (21.08%), a result in agreement with the lowest BTC peaks of replicates. The Δfgt1 strain was mostly retained in the sand (73.46%) compared to the WT and Δfgt2 strains presenting 65.04% and 59.49% retention rates, respectively. Furthermore, the Δfgt1 strain showed the highest total recovery (84.03%) compared to the WT (79.72%) and the Δfgt2 strain (80.53%).
The RPs of WT (Fig. 2b), Δfgt1, and Δfgt2 (Fig. 3b) strains had a non-monotonic distribution among replicates with the highest number of bacteria retained (S/C0) in the column inlet (0–2.25 cm). The dispersion of the Δfgt2 strain was the lowest among layers 2.25–18.0 cm, which showed a better repeatability than the WT and the Δfgt1 strain. The Δfgt1 strain was more dispersed among replicates in layer 2.25–18.0 cm. Contrary to the Δfgt1 and Δfgt2 strains, WT had a more dispersed number of bacteria in layer 12–18 cm.
Flagellar hydrophobicity reduces bacterial transport but promotes deposition
The surface hydrophobicity of the purified flagella from WT, Δfgt1, and Δfgt2 (Fig. 4a) was higher for the Δfgt1 mutant than that of WT and showed a significant difference (P < 0.001). No significant difference was observed between the WT and the Δfgt2 strain.
*(a) The flagellar hydrophobicity of P. amygdali pv. tabaci WT, Δfgt1 (without glycosylation), and Δfgt2 (partially glycosylated) strains. Replicates are shown as mean values; error bars represent standard deviations, and statistical significances were determined by Student’s t-test with WT (**P < 0.001). NS, not significant. (b) Relationship between bacterial flagellar hydrophobicity, recovery rate from effluent (Meff), and retention rate in the sand (Mretained).
A linear relationship was observed between flagellar hydrophobicity and both the effluent recovery rate (Meff) and retention rate in sand (Mretained) (Fig. 4b). Flagellar hydrophobicity showed a strong positive correlation with bacterial retention in the sand (R² = 0.89), indicating that increased hydrophobicity enhances adhesion to the sand matrix. Conversely, a moderate negative correlation was observed between hydrophobicity and effluent recovery (R² = 0.70), suggesting that more hydrophobic strains are less efficiently transported through the porous medium. Among the three flagellated strains, the Δfgt1 mutant, characterized by the highest flagellar hydrophobicity, exhibited the lowest recovery in the effluent and the highest retention in the sand compared to the WT and Δfgt2. These results suggest that flagellar hydrophobicity is a key factor driving bacterial deposition in porous media.
Bacterial preferential transport and their mechanisms of deposition
The BTCs for all mutants of P. amygdali pv. tabaci were well described using the MIM model based on the Hydrus-1D code (Fig. 2 and 3) with R^2^ >0.93. Mean values of transport and deposition parameters are reported in Fig. 5, and the fitted parameters of all replicates are shown in Material S3.
Fitted parameters of P. amygdali pv. tabaci WT, ΔfliC, Δfgt1, and Δfgt2 strains. Replicates are shown as mean values, and error bars represent standard deviations.
The dispersivity λ and the mobile water fraction θm/θ are key indicators of flow and transport behavior in porous media. Low λ and high θm/θ values suggest more uniform, less dispersive flow with reduced preferential transport pathways. All bacterial strains showed similar transport behavior, with mean θm/θ values ranging from 0.64 to 0.68 (Fig. 5), indicating that 64%–68% of the pore volume was involved in convective transport, while the remaining 32%–36% represented immobile zones that were either excluded from flow or available only for diffusive transport. No significant differences in θm/θ were observed among the strains. Dispersivity λ was highest for the WT strain (0.44 cm), followed by ΔfliC (0.33 cm), Δfgt2 (0.27 cm), and Δfgt1 (0.23 cm). Although the WT strain showed greater variability between replicates, all λ values remained within the same order of magnitude, suggesting broadly comparable transport dynamics across strains.
The first-order attachment (katt), detachment (k_d_), and straining (kstr) coefficients derived from simulations highlight the respective contributions of physicochemical attachment, reversible detachment, and physical straining to bacterial deposition. The WT, ΔfliC, and Δfgt1 strains exhibited similar attachment coefficients, with a mean katt value of 0.18 min^−^¹, indicating comparable attachment behavior. In contrast, the Δfgt2 mutant displayed a significantly reduced katt value (0.08 min^−1^) (Fig. 5; Material S3), approximately one order of magnitude lower, suggesting impaired attachment efficiency likely due to partial glycosylation of flagella. The WT strain exhibited a significantly lower detachment coefficient k_d_ of 0.39 min^−1^ compared to the ΔfliC mutant, which showed a value of 3.06 min⁻¹ (Material S3). This indicates that the ΔfliC strain is more prone to reversible detachment from sandy grains under hydrodynamic forces. A similar trend was observed for the Δfgt1 and Δfgt2 mutants with k_d_ of 2.05 and 1.29 min^−1^, respectively. These findings suggest that the WT strain is more likely to undergo irreversible attachment, likely driven by stronger physicochemical interactions with the sand surface. The kstr values obtained for all bacterial strains ranged from 0.86 to 2.04 min^−1^ (Fig. 5). Given the similar orders of magnitude and standard deviations of the kstr values across all strains, physical straining appears to be a consistent mechanism contributing equally to bacterial retention among the tested strains. As previously reported by Zheng et al. (17), some parameters derived from numerical simulations exhibited standard error coefficients (obtained from the Hydrus-1D model; see Material S3) that were comparable in magnitude to their corresponding absolute values. Nevertheless, comparisons between the different mutants can still reveal qualitative trends that may be linked to their distinct biological characteristics.
DISCUSSION
Flagella not only drive flagellar motility but also function as a factor in bacterial adhesion to host cells (13, 15, 35, 36) and other surfaces, including polystyrene (37). In this study, the ΔfliC strain lacking flagella was the most efficiently recovered in the effluent and exhibited the lowest retention within the sand (Table 2). Lower retardation factors and earlier breakthroughs observed for the ΔfliC strain relative to the WT (Fig. 2a and b) suggest faster transport, potentially due to the absence of flagella and the resulting loss of motility. The ΔfliC mutant exhibits a significantly reduced capacity to elicit disease symptoms compared to the WT strain (38). In agreement with the present work, the S. Typhimurium ΔflgKL mutant, lacking flagella, demonstrated faster transport through porous media than both the WT and the non-motile, flagellated ΔmotAB strain (17). This mutant also showed lower retention in sand compared to flagellated strains, further supporting the role of flagella in promoting surface interactions. Similarly, previous studies have reported that non-flagellated bacterial strains exhibit greater mobility than flagellated ones in sandy media (39). Overall, the ΔfliC mutant in our study exhibited greater mobility, likely due to reduced surface adhesion, resulting in shorter residence times and more efficient transport through porous media; however, as in many studies involving biological cells, a small fraction of bacteria (~20%) could not be recovered because some cells remain irreversibly attached to sand grains despite the standardized extraction procedure.
The WT strain showed more dispersive flow behavior, as indicated by a higher mean dispersivity relative to the ΔfliC strain, which may be attributed to the influence of flagella on bacterial movement and interaction with the porous medium (17). Lower mean dispersivity and more uniform flow patterns, reflected by higher θm/θ values, were observed for the S. Typhimurium ΔflgKL mutant lacking flagella, compared to both the WT and the non-motile, flagellated ΔmotAB strain in porous media. These differences were attributed to the presence or absence of flagella, suggesting that flagellated bacterial strains tend to exhibit less uniform flow and more preferential transport pathways in porous environments. While a similar trend in dispersivity was observed in our study, the close θm/θ values between the WT (64%) and ΔfliC (66%) suggest only minor differences in flow uniformity, making it difficult to establish a clear link between flagellum presence and preferential transport behavior. Post-translational modification of flagella significantly influenced bacterial transport, as reflected by effluent recovery rates. The Δfgt1 strain, lacking flagellar glycosylation, exhibited the lowest recovery in the effluent compared to the glycosylated WT and the partially glycosylated Δfgt2 strain. Glycosylation may enhance the structural stability of the flagellar filament by stabilizing polymerized flagellin proteins, as reported for P. amygdali pv. tabaci (24). This structural reinforcement could explain the reduced recovery and increased retention of the non-glycosylated strain in the sand matrix. As observed for flagellum presence, its post-translational modification via glycosylation did not appear to influence flow uniformity under the experimental conditions of this study, as indicated by the similar θm/θ values across strains. The P. amygdali pv. tabaci ΔfliC mutant exhibited the lowest deposition in porous media among all tested strains, likely due to the absence of flagella, which may reduce its adhesion capacity in the sand matrix. This observation is consistent with previous findings showing that Escherichia coli O157:H7 strains lacking FliC exhibited significantly lower adhesion to bovine intestinal and plant tissues compared to the WT (12, 40). Moreover, non-flagellated S. Typhimurium and E. coli mutants exhibited reduced deposition onto sand grains compared to the WT strain (17, 41). Our findings underscore the significance of flagella in facilitating bacterial deposition by augmenting surface adhesion, both in vitro and in vivo. The mechanisms driving adhesion of the WT strain to surfaces remain to be fully elucidated. In many motile bacteria, adhesion often involves the formation of non-motile, multicellular communities called biofilms, which enable stable surface attachment and enhanced environmental persistence (42). In these biofilms, adhesins such as pili and flagella, as well as extracellular matrix components like DNA and exopolysaccharides, along with bacterial motility, play crucial roles in early attachment and biofilm development. This presents a greater challenge for bacteria to detach from surfaces (43–45), thereby enabling them to thrive and persist in diverse environments (46).
The WT strain exhibited a lower retention rate compared to the non-glycosylated Δfgt1 mutant, highlighting the critical role of flagellar glycosylation in enhancing bacterial adhesion and biofilm formation (27, 47). Although glycosylation-defective mutants show reduced swimming activity in highly viscous media (48), their swimming motility under standard conditions, most relevant to transport through porous media, does not differ from that of the WT. However, P. amygdali pv. tabaci strains lacking flagellar glycosylation display markedly diminished swarming motility and impaired biofilm formation on plant surfaces, along with reduced adhesion to polystyrene (23). Swarming, a collective surface-associated behavior, plays a key role in early colonization and biofilm establishment (49–51). Thus, the reduced adhesion and impaired swarming motility of the non-glycosylated strain likely weaken its ability to initiate and sustain biofilms within porous media during column experiments. This diminished biofilm-forming capacity provides a mechanistic basis for the altered transport dynamics and lower retention observed for this strain. Together, these results highlight how flagellar glycosylation influences not only structural properties but also the interplay between motility, adhesion, and surface colonization, shaping bacterial persistence and spatial organization in heterogeneous environments such as porous substrates. The non-glycosylated Δfgt1 strain exhibited a higher propensity for detachment from sandy grains, which may explain its elevated detachment coefficient (k_d_) compared to the glycosylated WT and Δfgt2 strains. The Δfgt1 strain demonstrated greater retention than the WT strain due to physicochemical attachment forces, a process that is reversible. Additionally, the Δfgt1 strain could still be retained through physical straining, as evidenced by the higher kstr values observed in our study compared to the other strains (Fig. 5). Our findings reveal that bacterial strains deficient in flagellar glycosylation display the highest surface hydrophobicity (Fig. 4a). This observation aligns with previous studies demonstrating that the presence of pilin glycans in Pseudomonas aeruginosa reduces the hydrophobicity of both purified pili and whole cells (52). As anticipated, the observed positive correlation between flagellum hydrophobicity and retention rates (Fig. 4b) aligns with previous research findings on the relationship between bacterial surface hydrophobicity and retention under unsaturated flow conditions (53). Hydrophobicity was identified as the primary factor influencing bacterial deposition. The Δfgt1 strain demonstrated higher hydrophobicity compared to the WT, resulting in increased deposition on sandy surfaces. In contrast, the Δfgt2 strain exhibited flagellar hydrophobicity like that of the WT, which led to enhanced deposition relative to the WT strain. Similarly, S. Typhimurium lacking flagellar methylation showed significantly reduced deposition compared to the WT (17), with its flagellar hydrophobicity being lower than that of the WT (30). Research has extensively explored flagellar post-translational modifications, particularly in bacteria from natural aquatic environments such as Shewanella oneidensis (54) and various other bacterial species, including Enterobacteriaceae (20, 55). This study underscores the critical role of flagellar glycosylation in promoting the dissemination of plant pathogens via water runoff and their potential transport into groundwater systems. These post-translational modifications may enhance bacterial motility and surface interactions, offering an advantage in colonizing diverse environments by finely tuning the balance between dispersal and biofilm formation. However, our understanding of how glycosylation contributes to the environmental dissemination of pathogens remains limited. A key challenge lies in dissecting the specific contributions of glycosylation, as current approaches such as creating mutants of glycosyltransferases or altering glycosylation target sites through amino acid substitution each come with limitations. Glycosyltransferase mutations may have pleiotropic effects on other proteins or pathways, even when genes are located within operons containing other flagellum-related elements. Conversely, amino acid substitutions on flagellins may not fully recapitulate the structural or functional consequences of glycan removal and may affect flagellar integrity or function in unintended ways (23). Moreover, because we could not include the single Ser/Ala-substituted mutants (S143A, S164A, S176A, S183A, S193A, and S201A) or perform genetic complementation of the ΔfliC, Δfgt1, and Δfgt2 strains, we cannot fully exclude potential off-target effects associated with the knockout approach. As such, part of the observed phenotypes may reflect unintended genetic alterations rather than solely the loss of flagellin glycosylation. Future work using site-specific substitutions and complemented strains will be necessary to confirm the direct contribution of these modifications. Despite these constraints, these complementary strategies offer valuable insights and highlight the need for more refined tools to study glycosylation in situ. Future work should therefore focus on developing targeted approaches such as site-specific glycan editing or real-time imaging of motility and biofilm formation under flow conditions to better understand how glycosylation of surface structures like flagella and pili influences environmental survival and pathogen spread (56).
Conclusion
Through laboratory column transport experiments, we investigated and quantified the transport and deposition behaviors of bacterial strains in sandy porous media, focusing on the role and glycosylation of flagella. By integrating observations with modeling, we elucidated the impact of flagellar hydrophobicity on bacterial recovery and retention. Our findings clearly demonstrate that flagella impede bacterial transport and enhance deposition, with the WT showing higher retention compared to the ΔfliC strain, which exhibited greater potential for detachment. The Δfgt1 strain displayed faster transport and lower recovery rates, indicating that flagellar glycosylation facilitates effluent recovery and reduces deposition. Furthermore, increased flagellar hydrophobicity was found to hinder transport while promoting deposition. This study highlights critical insights into the effects of glycosylation, which are essential for evaluating the risk posed by phytopathogenic bacteria in farmland and for developing effective soil bioremediation strategies.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kim S-B. 2006. Numerical analysis of bacterial transport in saturated porous media. Hydrol Process 20:1177–1186. doi:10.1002/hyp.5930 · doi ↗
- 2Bai H, Cochet N, Pauss A, Lamy E. 2016. Bacteria cell properties and grain size impact on bacteria transport and deposition in porous media. Colloids Surf B Biointerfaces 139:148–155. doi:10.1016/j.colsurfb.2015.12.01626705829 · doi ↗ · pubmed ↗
- 3Safadoust A, Mahboubi AA, Mosaddeghi MR, Gharabaghi B, Unc A, Voroney P, Heydari A. 2012. Effect of regenerated soil structure on unsaturated transport of Escherichia coli and bromide. J Hydrol (Amst) 430–431:80–90. doi:10.1016/j.jhydrol.2012.02.003 · doi ↗
- 4Yang L, Kang J, Chen X, Ripp SA, Johnson WP, Zhuang J. 2021. Real-time bioluminescent imaging of spatiotemporal variation of microbial retention during transport through porous media under variably saturated flow conditions. J Hydrol (Amst) 601:126603. doi:10.1016/j.jhydrol.2021.126603 · doi ↗
- 5Redman JA, Walker SL, Elimelech M. 2004. Bacterial adhesion and transport in porous media: role of the secondary energy minimum. Environ Sci Technol 38:1777–1785. doi:10.1021/es 034887 l 15074689 · doi ↗ · pubmed ↗
- 6Ning Z, Li R, Lian H, Zhou S, Liao P, Liu C. 2021. Effects of flow-interruption on the bacteria transport behavior in porous media. J Hydrol (Amst) 595:125677. doi:10.1016/j.jhydrol.2020.125677 · doi ↗
- 7Bai H, Cochet N, Drelich A, Pauss A, Lamy E. 2016. Comparison of transport between two bacteria in saturated porous media with distinct pore size distribution. RSC Adv 6:14602–14614. doi:10.1039/C 5RA 21695 H · doi ↗
- 8Kim HN, Walker SL. 2009. Escherichia coli transport in porous media: influence of cell strain, solution chemistry, and temperature. Colloids Surf B Biointerfaces 71:160–167. doi:10.1016/j.colsurfb.2009.02.00219278837 · doi ↗ · pubmed ↗