Developmental and aging trajectories of 40-Hz auditory steady-state responses: A systematic review across the human lifespan
Aurimas Mockevičius, Danylo Machevskyi, Dariusz Majcherczyk, Inga Griškova-Bulanova

TL;DR
This review examines how 40-Hz auditory steady-state responses change across the human lifespan, highlighting key developmental patterns and gaps in current research.
Contribution
The paper provides a systematic synthesis of age-related changes in 40-Hz ASSRs, revealing nonlinear developmental and aging trajectories.
Findings
ASSR amplitude and phase-locking increase significantly from infancy to adolescence.
Adult and aging studies show inconsistent results, likely due to methodological differences.
Sparse data in early childhood and older adulthood limit understanding of full lifespan patterns.
Abstract
Auditory steady-state responses (ASSRs) are rhythmic neural oscillations that synchronize to periodic auditory stimulation and serve as a noninvasive index of cortical network dynamics. ASSRs, particularly those at 40 Hz, have received substantial attention as sensitive markers of temporal precision, excitation–inhibition balance, and functional connectivity in the auditory cortex, and have been widely applied in translational research on neurodevelopmental and neuropsychiatric disorders. Because gamma synchronization supports key cognitive functions, including auditory temporal processing, selective attention, speech perception, and early language development, mapping its lifespan trajectory provides insight into how maturing cortical dynamics underpin cognitive development. However, despite extensive clinical use, the normative developmental and aging trajectory of gamma-range ASSRs…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1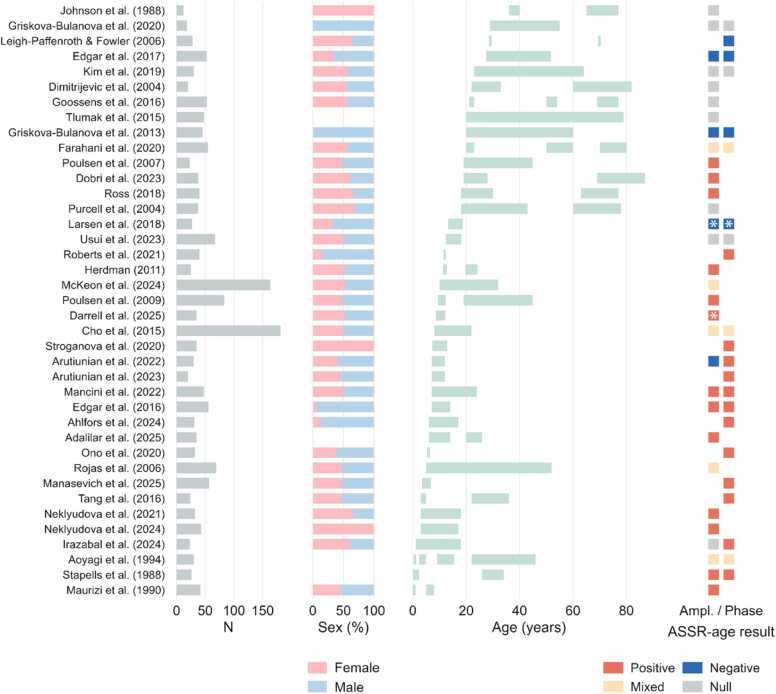 Figure 2
Figure 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeural dynamics and brain function · Neuroscience and Music Perception · Hearing, Cochlea, Tinnitus, Genetics
Introduction
1
Auditory steady-state responses (ASSRs) refer to rhythmic neural activities that align with the temporal patterns of periodic auditory stimuli (Picton et al., 2003). When sounds are modulated at specific frequencies, cortical populations synchronize their activity to the stimulus rhythm, producing a measurable outcome in EEG or MEG recordings (Legget et al., 2017).
While ASSRs can be elicited across a broad range of modulation frequencies, responses within the gamma range (30–100 Hz) have attracted particular attention because they reflect fast neural synchronization that stands as an essential mechanism of perceptual binding, attention, and working memory (Başar-Eroglu et al., 1996). Within this range, 40-Hz ASSR has emerged as a particularly robust outcome, linked to the functional integrity of GABAergic and glutamatergic networks within the auditory cortex (Toso et al., 2024). Consequently, the 40-Hz ASSR has become a tool reflecting excitation–inhibition (E/I) balance (Tada et al., 2020), temporal processing abilities (Kadowaki et al., 2022), and state of functional connectivity (Du et al., 2023; Ying et al., 2015) in the human brain. It stands as an attractive output for translational research offering a stable, noninvasive, and frequency-specific index of neural synchronization (Farahani et al., 2020a, Hirano et al., 2020).
Abnormalities in ASSR strength and synchronization level have been reported in neuropsychiatric and neurodevelopmental disorders, including schizophrenia (Thuné et al., 2016, Zouaoui et al., 2023), bipolar disorder (Jefsen et al., 2022), autism spectrum disorder (Arutiunian et al., 2023, Seymour et al., 2020), Alzheimer’s disease (van Deursen et al., 2011), and corresponding animal models (Jasinskyte et al., 2025) where alterations in network structure and E/I dynamics are well documented. Furthermore, evidence suggests that ASSR may reflect cognitive abilities (Parciauskaite et al., 2021). These observations make ASSRs a valuable candidate biomarker for probing cortical network function across both health and disease.
Despite extensive research in clinical populations, the normative trajectory of gamma-range ASSRs across the human lifespan remains poorly defined. While individual studies have investigated developmental or aging effects on ASSRs, findings have not yet been systematically integrated and full picture is not clear. Without a comprehensive understanding of how ASSRs mature during typical development and evolve with aging, the specificity and interpretability of abnormalities observed in clinical samples remain uncertain. Establishing normative patterns of ASSR properties from early childhood through late adulthood is therefore crucial for distinguishing pathological alterations from age-appropriate variability. A systematic synthesis of lifespan data is needed to determine whether changes observed in clinical populations reflect delayed maturation, accelerated decline, or distinct pathophysiological mechanisms.
At the neurobiological level, the ability to generate and maintain gamma-range ASSRs depends on the coordinated activity of excitatory pyramidal neurons and inhibitory parvalbumin-positive (PV⁺) interneurons (Tada et al., 2020), which regulate the precise timing of cortical firing (Buzsáki and Wang, 2012). Developmental changes in ASSRs may thus reflect the maturation of inhibitory circuitry, synaptic refinement, and progressive myelination (Toga et al., 2006) that enhance temporal precision within auditory pathways. Conversely, age-related changes in GABAergic function (Zuppichini et al., 2024), synaptic density (Toyonaga et al., 2024), and white-matter integrity (Salat et al., 2005) may underlie a diminished capacity for neural synchronization in later life. Together, these processes suggest a potentially non-linear lifespan pattern of gamma synchronization.
Although the cellular and neurochemical bases of ASSR generation are increasingly well understood, no previous synthesis has integrated these mechanistic insights with empirical findings across different life stages. The present review aims to systematically summarize evidence on age-related changes in low-gamma (30–60 Hz) ASSRs measured with EEG or MEG across the human lifespan, integrating developmental, adult, and aging data to delineate how neural synchronization to external stimulation evolves from early childhood to older adulthood.
Methods
2
Review design and objectives
2.1
This study was conducted in line with the Preferred Reporting Items for Systematic reviews and Meta-Analyses extension for Scoping Reviews (PRISMA-ScR) (Tricco et al., 2018). The protocol was not prospectively registered. The aim was to summarize age-related differences in ASSRs recorded with EEG or MEG across the human lifespan. The research question was defined using the PICOS framework: Population (P) - humans across the lifespan, including typically developing and clinical groups; Intervention/Exposure (I) - EEG or MEG measurement of ASSRs elicited by periodic auditory stimulation; Comparison (C) - between-age group comparisons or age as a continuous predictor; Outcomes (O) - quantitative ASSR measures (amplitude, inter-trial phase coherence, phase-locking factor, power, signal-to-noise ratio, latency, or topographic features, etc.); Study design (S) - peer-reviewed human studies with cross-sectional or longitudinal designs.
Search strategy
2.2
The literature search aimed to identify all peer-reviewed human studies examining age-related effects on ASSRs measured with EEG or MEG. Two databases, PubMed and Scopus, were searched to ensure comprehensive coverage across biomedical and interdisciplinary sources. No date restrictions were applied, and only English-language full-text publications were considered. The search targeted three conceptual domains: (i) developmental stages (childhood and adolescence), (ii) aging and older adulthood, and (iii) general age or maturation effects across the lifespan. Each domain was represented by a dedicated Boolean query, detailed below.
Search procedure
2.3
The search was conducted systematically in accordance with the predefined strategy. Both PubMed and Scopus databases were queried, with no temporal limitations. Three complementary searches were performed in each database, targeting studies on childhood/adolescence, aging, and general maturation.
The search strategy combined terminology related to ASSRs (auditory steady-state response, steady-state auditory evoked potential, ASSR, SSAEP) with population-specific terms for developmental stages (child, infant, adolescent, teen, pediatric) and aging (aging, older adults, elderly, senescence). It also included broader age-related concepts (age-related, maturation, lifespan, development, age effects). Finally, all searches constrained results to studies using EEG or MEG methods (EEG, electroencephalography, MEG, magnetoencephalography). Complete search formulations are provided in Supplementary material.
Eligibility criteria
2.4
Studies were included if they involved human participants of any age, used EEG or MEG to record ASSRs elicited by periodic amplitude- or frequency-modulated sounds within the gamma range (typically 30–80 Hz), and reported age-related analyses through group comparisons or correlations. Eligible studies provided quantitative electrophysiological measures and were not limited to hearing-threshold precision estimation. Only original, peer-reviewed articles published in English were included.
Exclusion criteria encompassed non-human or invasive studies, the absence of age-related data, and investigations limited to behavioral or hemodynamic measures. Reviews, case reports, editorials, and conference abstracts, as well as duplicates and non-English full texts, were excluded.
Study selection process
2.5
All search results were imported into Rayyan (rayyan.ai, Ouzzani et al., 2016) for reference management and screening, resulting in a total of 550 records. Automatic and manual de-duplication were performed prior to screening. Three reviewers independently assessed titles and abstracts against the eligibility criteria, resolving disagreements through discussion or, when necessary, consultation with a fourth reviewer. After removing 246 duplicates, 304 unique records were screened, of which 26 studies met all inclusion criteria and were retained for qualitative synthesis. Subsequently, the reference lists of the retained articles were screened to identify relevant studies. Additionally, a nonsystematic search was conducted in Google Scholar to identify any potentially missed articles. The additional searches resulted in the inclusion of 14 more papers in this review. The full selection process is summarized in the PRISMA flow diagram (Fig. 1).Fig. 1PRISMA diagram representing the article inclusion process.Fig. 1
Data extraction
2.6
For each included study, relevant data were extracted into a structured spreadsheet. Extracted information included study identifiers (authors, year, DOI), participant characteristics (sample size, age range, sex distribution, and clinical status), and details of auditory stimulation and recording. Stimulation parameters comprised modulation type and frequency, carrier signal, intensity, duration, and presentation mode. Recording information specified whether EEG or MEG was used, the number of channels, and whether analyses were performed at the sensor or source level. Reported ASSR outcomes (such as amplitude, power, inter-trial phase coherence (ITPC), phase-locking factor (PLF), signal-to-noise ratio (SNR), and latency) were recorded along with the direction and significance of any age-related effects and hemispheric asymmetries. Results were extracted with a focus on responses in the 30–60 Hz frequency range (for single frequency stimulation) or at peak gamma frequency (for sweep-like stimulation). Data extraction was conducted independently by two reviewers, and discrepancies were resolved through verification and consensus.
Results
3
A formal quality appraisal of the included studies was not conducted, as the review aimed to map the existing evidence rather than evaluate study bias. The findings were integrated through qualitative narrative synthesis. Data were thematically organized according to developmental stage (childhood/adolescence, adulthood, aging) and clinical status, as well as by methodological factors such as recording modality, stimulus type, and modulation frequency. The extracted information is provided in Table 1.Table 1. Available information extracted from the included studies regarding samples (sample and group types and sizes, sex composition, mena age, standard deviation and ranges), auditory stimulation (stimulation frequency and carrier, stimulus and inter-stimulus-interval duration, number of repetitions, monaural/binaural stimulation, intensity and instruction during the stimulation), data acquisition (EEG/MEG and the number of electrodes/channels), analysis (measure and ROI) and results (ASSR relationship with age or between-group comparisons).Table 1. StudySampleStimulusStimulationRecordingAnalysisAge-related findings1Adalilar et al. (2025)Healthy subjectsGroup 1 (N = 18)Age: 20–26 yearsGroup 2 (N = 10)Age: 6–10 yearsGroup 3 (N = 7)Age: 11–14 years37, 40, 43, 77, 80, 83 Hz AMwhite noise carrierStimulus condition (6 frequencies x left/right/binaural) lasted 307.2 sMonaurally and binaurally70 dB SPLSilent movieEEG, 64 electrodesAmplitude, SNR, phase lag, lateralizationSource level:TP7, CP5, P9, P5, P7, PO7, PO3, O1, TP8, CP6, P10, P6, P8, PO8, PO4, and O2Age was positively related to 40-Hz ASSR amplitude (only right ear stimulation), SNR and latency2Ahlfors et al. (2024)Subjects with ASD (N = 22, 4 F)Age: 13.6 ± 0.6 (7−17) yearsHealthy subjects (N = 31,4 F)Age: 13.1 ± 0.6 (6−17) years25 and 43 Hz AMbroadband noise carrier1200 ms duration (300–400 ms ISI)100 repetitionsBinaurally65 dB SPLMEG, 306 channelsITPCSource level:bilateral auditory cortexAge positively predicted 43-Hz ASSR ITPC3Aoyagi et al. (1994)Healthy subjectsGroup 1 (N = 5)Age: 8.6 (4−15) monthsGroup 2 (N = 5)Age: 3 years 4 months (2 years 3 months – 4 years 11 months)Group 3 (N = 10)Age: 11 years 4 months (9 years 1 month – 15 years 6 months)Group 4 (N = 10)Age: 31.4 (22−46) years20–200 Hz (20 Hz steps) AM 1000-Hz carrierMonaurally50 dB nHLDuring sleepEEG,1 electrodeAmplitude, SNR and ITPCSensor level: vertex40-Hz ASSR ITPC and SNR was higher in adults and older children compared to younger children4Arutiunian et al. (2022)Healthy subjects (N = 30, 12 F)Age: 9.1 ± 1.5 (7.06–12.03) years40 HzAM 1000 Hz carrier1000 ms duration (2000 ms ISI)90 repetitionsBinaurally83.7 dB SPLFixation crossMEG,306 channelsITPC and amplitudeSource level: bilateral temporal cortices40-Hz ASSR ITPC correlated positively, while amplitude correlated negatively with age, only in the right hemisphere5Arutiunian et al. (2023)Subjects with ASD (N = 20, 5 F)Age: 10.03 ± 1.7 (8.02–14.01) yearsHealthy subjects (N = 20, 9 F)Age: 9.11 ± 1.3 (7.02–12.03) years40 HzAM 1000 Hz carrier1000 ms duration (2000 ms ISI)90 repetitionsBinaurally83.7 dB SPLFixation crossMEG,306 channelsITPCSource level: bilateral auditory cortex40-Hz ASSR ITPC correlated positively with age in both hemispheres in healthy group only6Cho et al. (2015)Healthy subjects (N = 181, 89 F)Age: 14.5 ± 4.4 (8−22) yearsGroup 1 (N = 44, 21 F)Age: 8–10 yearsGroup 2 (N = 32, 16 F)Age: 11–13 yearsGroup 3 (N = 37, 19 F)Age: 14–16 yearsGroup 4 (N = 36, 18 F)Age: 17–19 yearsGroup 5 (N = 32, 15 F)Age: 20–22 years20, 30, 40 Hz click trains500 ms durationBinaurally65 ± 5 dBOddball paradigmEEG, 128 electrodesAmplitude and ITPCSensor level: 14 electrodes centered at FCz40-Hz ASSR amplitude and ITPC linearly increased from age group 1–3, but decreased from age group 3–5. No significant differences in 30-Hz ASSR measures7Darrell et al. (2025)Subjects with ASD (N = 53, 10 F)Age: 10.84 ± 1.5 yearsHealthy subjects (N = 35, 18 F)Age: 10.4 ± 1.8 years27, 40 Hz click trains500 ms duration (488–788 ms ISI)200 repetitionsBinaurally60 dB SPLOddball paradigmEEG,64 electrodesPowerSensor level: FC3, FCz, FC440-Hz ASSR power correlated positively with age in a combined and ASD group8Dimitrijevic et al. (2004)Hearing-impaired subjects (N = 10, 6 F)Age: 75 (57−86) yearsHealthy subjectsGroup 1 (N = 10, 5 F)Age: 28 (22−33) yearsGroup 2 (N = 10, 6 F)Age: 68 (60−82), years39–55 and 78–100 Hz AM and FM 500, 1000, 1500 and 2000 Hz carrierBinaurally70 dB SPLDuring sleepEEG,1 electrodeAmplitudeSensor level: vertexNo significant differences in ASSR amplitudes between normal-hearing age groups9Dobri et al. (2023)Healthy subjectsGroup 1 (N = 19, 11 F)Age: 23.8 ± 6.2 (19−28) yearsGroup 2 (N = 19, 12 F)Age: 76.1 ± 6.2 (69−87) years40 Hz AM 400 Hz carrier2050 ms total duration, consisting of 400-ms bursts separated by 12.5 ms pauses (1950-ms ISI)Presented alone combined with babble noise300 repetitions per conditionBinaurally60 dB above individual thresholdPassive listeningMEG, 151 channelsAmplitudeSource level: bilateral auditory corticesSignificantly higher 40-Hz ASSR amplitude in older vs young group only in noise condition.Significant positive correlation between 40-Hz ASSR amplitude with age in older group10Edgar et al. (2016)Subjects with ASD (N = 55, 4 F)Age: 10.6 ± 1.5 (8−14) yearsHealthy subjects (N = 56, 3 F)Age: 10.1 ± 1.5 (7−14) years40 HzAM 500 Hz carrier1 s duration (4 s ISI)Binaurally45 dB above hearing thresholdSilent movieMEG,306 channelsPower and ITPCSource level: bilateral auditory cortexAge was a positive predictor of 40-Hz ASSR power (right hemisphere) and ITPC (both hemispheres)11Edgar et al. (2017)Healthy subjects (N = 53, 18 F)Age: 39.6 ± 12.1 years40 HzAM 500 Hz carrier1 s duration (4 s ISI)88 ± 18 average repetitionsBinaurally35 dB above individual hearing thresholdFixation crossMEG,306 channelsEEG,60 electrodesPower and ITPCSource level: bilateral auditory cortexSensor level: Cz, Fz and 18 electrodes per hemisphereSignificant negative correlation of age with 40-Hz ASSR ITPC and power in the left hemisphere at source level12Edgar et al. (2018)Subjects with SZ (N = 41, 7 F)Age: 40.3 ± 11.6 (20–60) yearsHealthy subjects (N = 55, 19 F)Age: 39.6 ± 11.9 (21–58) years40 Hz AM 500 Hz carrier1 s duration (4 s ISI)87 ± 18 (SZ) or 92 ± 22 (healthy) average repetitionsBinaurally35 dB above individual hearing thresholdMEG,306 channelsEEG,60 electrodesPower and ITPCSource level:Bilateral temporal corticesSignificant negative correlation of age with 40-Hz ASSR ITPC and power in healthy controls but not SZ patients13Farahani et al., 2020b, Farahani et al., 2020aHealthy participantsGroup 1 (N = 19, 10 F)Age: 20–30 yearsGroup 2 (N = 20, 10 F)Age: 50–60 yearsGroup 3 (N = 16, 11 F)Age: 70–80 years4, 2, 40 and 80 HzAM white noise carrier300 s durationMonaurally70 dB SPLSilent videoEEG,64 electrodesAmplitude and ITPCSource level: multiple cortical and subcortical ROIs40-Hz ASSR amplitude was higher in multiple ROIs in older compared to younger and middle-aged groups; higher in younger compared to middle-aged group;40-Hz ASSR ITPC was higher in frontal cortical ROIs in older compared to younger and middle-aged group14Goossens et al. (2016)Healthy subjectsGroup 1 (N = 19, 10 F)Age: 22 ± 1 yearsGroup 2 (N = 20, 10 F)Age: 52 ± 2 yearsGroup 3 (N = 14, 10 F)Age: 74 ± 3 years4, 20, 40, 80 HzAM white noise carrier300 s durationMonaurally and binaurally70 dB SPLSilent movieEEG, 64 electrodesSNR, sensor level: TP7/8, CP5/6, P9/10, P7/8, P5/6, PO7/8, PO3/4, O1/2No differences in 40-Hz ASSR SNR between age groups15Griskova-Bulanova et al. (2013)Healthy subjects(N = 46, all M)Group 1 (N = 13)Age: 20–30 yearsGroup 2 (N = 13)Age: 30–40 yearsGroup 3 (N = 9)Age: 40–50 yearsGroup 4 (N = 11)Age: 50–60 years40 Hz click trains500 ms duration (1–1.5 s ISI)60 repetitionsBinaurally60 dB SPLFixation cross with passive listeningEEG,9 electrodesAmplitude and ITPCSensor level: F3, Fz, F4, C3, Cz, C4, P3, Pz, P4Age negatively correlated to 40-Hz ASSR ITPC and amplitude16Griskova-Bulanova et al. (2020)Subjects with SZ (N = 18, all M)Age: 38 ± 14 yearsHealthy subjects (N = 18, all M)Age: 42 ± 13 years1–120 Hz sweeps AM 440 Hz carrier500 ms duration(700–1000 ms ISI)450 repetitionsBinaurally60 dBASilent movieEEG,9 electrodesAmplitude and ITPCSensor level: Fz and CzNo correlation of ASSR with age17Herdman (2011)Healthy subjectsGroup 1 (N = 12, 6 F)Age: 12 ± 0.7 yearsGroup 2 (N = 13, 7 F)Age: 22 ± 2.3 years40 HzAM 800 or 1200 Hz carrier500 or 175 ms duration (1000–1500 ms ISI)96 or 384 repetitionsBinaurally70 dB SPLOddball paradigmMEG,151 channelsAmplitudeSource level: bilateral auditory cortexSignificantly higher 40-Hz ASSR amplitude in adults compared to children18Irazabal et al. (2024)Healthy subjects(N = 23, 14 F)Age: 115.26 ± 57.65 months (1–18 years)1–120 Hz sweeps AM 1200 Hz carrier1600 ms duration (400 ms ISI)500 repetitionsBinaurally85 dB SPLSilent videoEEG,19 electrodesEvoked power and ITPCSensor-level: F3, Fz, F4Significant positive correlation between ITPC at peak low gamma frequency and age; no correlation between evoked power and age19Johnson et al. (1988)Healthy subjectsGroup 1 (N = 5, all F)Age: 38 (36−40) yearsGroup 2 (N = 7, all F)Age: 69.6 (65−77) years40 Hz trains of 1000 Hz bursts500 repetitionsBinaurallyEyes-closed passive listeningEEG,21 electrodesAmplitudeSensor level: whole-headNo difference between age groups in 40-Hz ASSR20Kim et al. (2019)Subjects with SZ (N = 33, 17 F)Age: 42.21 ± 10.99 (21−60) yearsHealthy subjects (N = 30, 17 F)Age: 43.33 ± 12.95 (23–64) years40 Hz click trains500 ms duration (3050–3500 ISI)150 repetitionsBinaurally80 dB SPLFixation cross with active listeningEEG,64 electrodesPower and ITPCSensor level: CzNo significant correlation between age and 40-Hz ASSR21Larsen et al. (2018)Subjects with 22q11.2 deletion syndrome (N = 18, 5 F)Age: 15.39 ± 2.45 yearsHealthy subjects (N = 27, 9 F)Age: 15.96 ± 2.71 years40 Hz click trains1 s duration (2 s ISI)85 dB SPL120 repetitionsBinaurallyFixation cross with passive listeningEEG,128 electrodesPower and ITPCSensor level: CzAge negatively predicted 40-Hz ASSR power and ITPC in combined 22q11.2 and control group22Leigh-Paffenroth and Fowler (2006)Healthy subjectsGroup 1 (N = 16, 12 F)Age: 29 yearsGroup 2 (N = 12, 6 F)Age: 69.9 years20, 40 or 90-Hz AM 500 Hz or 1000 Hz carrierMonaurallyEEG,1 electrodeITPCSensor level: Fz40-Hz ASSR ITPC was significantly higher in younger subjects compared to older only at 500 Hz carrier frequency23Manasevich et al. (2025)Healthy subjects (N = 57, 27 F)Age: 64.35 ± 11.96 (42−82) months40 Hz click trains500 ms duration (500–800 ms ISI)Binaurally65 dB SPLSilent videoEEG,32 electrodesITPCSource level: FzSignificant positive correlation of 40-Hz ASSR with age24Mancini et al. (2022)Subjects with 22q11.2 deletion syndrome (N = 58, 26 F)Age: 17.6 ± 6.9 yearsHealthy subjects (N = 48, 24 F)Age: 17.7 ± 6.2 yearsBoth groups divided into age bins:1) 7–13 years2) 14–18 years3) ≥ 19 years40 HzAM 1000-Hz carrier2 s duration (1.5–2.5 s ISI)100 repetitionsBinaurallyOddball paradigmEEG,256 electrodesPower and ITPCSensor level: cluster of electrode around FCzSignificant linear increase in 40-Hz ASSR power and ITPC from childhood to adulthood in control group only25Maurizi et al. (1990)Healthy subjectsGroup 1 (N = 32, 16 F)Age: 1–3 daysGroup 2 (N = 10, 3 F)Age: 5–8 years40 Hz trains of 500 Hz bursts1000 burstsMonaurallyDecreasing intensity from 70 dB nHL to thresholdDuring sleep (neonates) or passive listening (children)EEG,1 electrodeAmplitude, test-retest reliabilitySensor level: upper foreheadIncreased 40-Hz ASSR amplitude and test-retest reliability in children as compared to neonates26McKeon et al. (2024)Healthy subjects(N = 164, F = 87)Age: 10–32 years20, 30, 40 Hz click trains500 ms duration (605 ms ISI)150 repetitionsBinaurallyPassive listeningEEG,64 electrodesEvoked power, spontaneous power and SNRSensor level: F3, F5, F7, F1, F2, F4, F6, F8, AFz, AF1, AF2, Fp1, Fp2, Fz, AF5, AF6ASSR SNR increased through adolescence only; no change in evoked power with age27Neklyudova et al. (2021)Healthy subjectsGroup 1 (N = 13, 7 F)Age: 16.04 ± 1.9 (12−18) yearsGroup 2 (N = 19, 14 F)Age: 7.8 ± 2.6 (3−12) years40 Hz click tains500 ms duration (500–800 ms ISI)150 repetitionsBinaurally80 dB SPLSilent videoEEG32 electrodesAmplitudeSensor level: Fz40-Hz ASSR amplitude was significantly higher in “old” group compared to “young” group28Neklyudova et al. (2024)Subjects with RTT (N = 43, all F)Age: 8.19 ± 3.84 (2.92–17.1) yearsHealthy subjects (N = 43, 26 F)Age: 8.27 ± 3.87 (2.58–17.45) years40 Hz click trains500 ms duration (500–800 ms ISI)150 repetitionsBinaurally65 dB SPLSilent videoEEG, 28 electrodesAmplitudeSensor level: FCzSignificant positive correlation between age and 40-Hz ASSR amplitude only in healthy group29Ono et al. (2020)Subjects with ASD (N = 23, 5 F)Age: 74.8 ± 11.2 monthsHealthy subjects (N = 32, 12 F)Age: 69.7 ± 6.2 months20, 40 HzAM 1000 Hz carrier1 s duration (900–1100 ms ISI)Binaurally70 dB SPLSilent videoMEG,151 channelsPower and ITPCSource level: bilateral auditory cortexPositive correlation of 40-Hz ASSR ITPC and age in the right hemisphere only in healthy group30Poulsen et al. (2007)Healthy subjects (N = 23, 11 F)Age: 29 (19−45) years40 HzFM1000 Hz carrier10–100 Hz sweepsAM white noise carrierFM stimulus: 1000 ms duration (1000 ± 100 ms ISI)400 repetitionsAM stimuli: 15.36 s up part, 15.36 s down part10 repetitions65 dB SPL (FM stimuli) and 55 dB SPL (AM stimuli)BinaurallySilent videoEEG,128 electrodesAmplitude Sensor level: CzSource level: bilateral temporal cortex and brainstemPositive relationship between 40-Hz ASSR amplitude (FM stimuli) and age, both in sensor and source level;No relationship between ASSR amplitude and age at peak frequency (AM stimuli)31Poulsen et al. (2009)Healthy subjectsGroup 1 (N = 60, 28 F)Age (T1): 10 ± 0.39 (9.4 – 10.8) yearsAge (T2): 11.5 ± 0.39 (10.9 – 12.3) yearsGroup 2 (N = 23, 11 F)Age: 29 (19−45) years40 HzFM1000 Hz carrier10–100 Hz sweepsAM white noise carrierFM stimulus: 1000 ms duration (1000 ± 100 ms ISI)400 repetitionsAM stimuli: 15.36 s up part, 15.36 s down part10 repetitions65 dB SPL (FM stimuli) and 55 dB SPL (AM stimuli)BinaurallySilent videoEEG,128 electrodesAmplitudeSensor level: CzSource level: bilateral temporal cortex and brainstemSignificantly higher 40-Hz ASSR in adults vs children; significant increase in amplitude at T2 compared to T1 in children group; no significant difference in EFR amplitude at the peak frequency between adult and children groups and within children group32Purcell et al. (2004)Healthy subjectsGroup 1 (N = 25, 20 F)Age: 18–43 yearsGroup 2 (N = 13, 6 F)Age: 60–7820–600 Hz sweepsAM white noise carrier30 s duration, 15-s up and 15-s down partsMonaurally50 or 60 dB SPLSilent movieEEG, 1 electrodeAmplitude, sensor level: CzNo differences in ASSR amplitude at peak frequency between age groups33Roberts et al. (2021)Subjects with ASD (N = 80, 12 F)Age: 11.72 ± 0.26 yearsHealthy subjects (N = 40, 6 F)Age: 11.94 ± 0.44 years10–100 Hz sweepAM 500 Hz carrier30 s sweeps of 15-s up and 15-s down parts (9 s ISI)20 repetitionsBinaurally45 dB SLMEG,275 channelsITPCSource level:bilateral auditory cortexAge positively predicted ASSR ITPC at the peak frequency34Rojas et al. (2006)Healthy subjects (N = 69, 32 F)Age: 25.62 ± 13.05 (5−52) years40 Hz click trains500 ms duration (1.5 s ISI)150 repetitionsBinaurally65 dB SPLSilent videoMEG,37 channelsPowerSensor level: channel with maximal powerExponential relationship between 40-Hz ASSR and age, with sharp increase in early age and plateau in adulthood35Ross (2018)Healthy participantsGroup 1 (N = 19, 12 F)Age: 22.4 (18−30) yearsGroup 2 (N = 21, 14 F)Age: 70.9 (63−77) years40 HzAM 400, 800, 1200, 1600 and 2400 Hz carrier13.6 s duration (16 s ISI)22 repetitionsBinaurally60 dB SPLFixation crossMEG,151 channelsAmplitudeSource level: bilateral auditory cortexAge positively predicted 40-Hz ASSR amplitude36Stapells et al. (1988)Healthy subjectsGroup 1 (N = 18)Age: 42.1 ± 32.4 weeks (3 weeks – 28 months)Group 2 (N = 8)Age: 26–34 years9–59 Hz (5 Hz steps) trains of 1000 Hz bursts or43.4 Hz click trains4096 repetitions of 1000-Hz bursts or 8192 repetitions of clicksMonaurally70 dB nHLDuring sleepEEG,5 electrodesAmplitude, phaseSensor level: CzASSR around 40 Hz was significantly higher in adults compared to children37Stroganova et al. (2020)Subjects with ASD (N = 35, all M)Age: 9.69 ± 1.5 (7.2–12.3) yearsHealthy subjects (N = 35, all M)Age: 10.08 ± 1.5 (7.3–12.9) years40 Hz click trains500 ms duration (1000 ms ISI)100 repetitionsMonaurally60 dB SPLSilent videoMEG,306 channelsITPCSource level: bilateral auditory cortex40-Hz ASSR ITPC correlated positively with age in both ASD and controls38Tang et al. (2016)Healthy subjectsGroup 1 (N = 12, 4 F)Age: 49.3 months (3–5 years)Group 2 (N = 12, 7 F)Age: 28.8 (22−36) years1–80 Hz sweepAM white noise carrier9 s duration (900–1000 ms ISI)100 (children) or 200 (adults) repetitionsBinaurally75 dB SPLSilent movieMEG,64 (children) and 160 (adults) channelsITPCSource level: bilateral auditory cortexASSR ITPC was higher in adults compared to children at 15–80 Hz39Tlumak et al. (2015)Healthy participantsGroup 1 (N = 16)Age: 29.69 ± 4.76 (20−39) yearsGroup 2 (N = 16)Age: 51.94 ± 5.27 (40−59) yearsGroup 3 (N = 16)Age: 63.37 ± 3.9 (60−79)0.75, 1.25, 2.5, 5, 10, 20, 40 HzAM 1000 Hz carrierRight ear70 dB SPLEEG,2 electrodesAmplitudeSensor level: CzNo differences in 40-Hz ASSR amplitude among age groups40Usui et al. (2023)Healthy subjects (N = 67, 33 F)Age (T1): 13.4 ± 0.5 (12.3–14.3) yearsAge (T2): 16.1 ± 0.8 (14.4–18.2) years20, 40 Hz click trains500 ms duration (500 ms ISI)200 repetitionsBinaurally80 dBPassive listening with eyes openEEG,2 electrodesPower and ITPCSensor level: Fz and CzNo significant differences in 40-Hz ASSR power and ITPC between T1 and T2 measurements
Of the 40 studies included in this review, 3 investigated infant samples below one year of age (Aoyagi et al., 1994, Maurizi et al., 1990, Stapells et al., 1988), demonstrating that measurable 40-Hz ASSRs can already be detected in early infancy. Eighteen studies examined ASSR in early life, covering ages up to mid 20 s (Adalilar et al., 2025, Ahlfors et al., 2024, Arutiunian et al., 2022, Arutiunian et al., 2023, Cho et al., 2015, Darrell et al., 2025; Edgar et al., 2016; Herdman et al., 2011; Irazabal et al., 2024; Larsen et al., 2018; Manasevich et al., 2025; Mancini et al., 2022; Maurizi et al., 1990; Neklyudova et al., 2021, Neklyudova et al., 2024; Ono et al., 2020; Roberts et al., 2021; Stroganova et al., 2020), predominantly highlighting developmental increases in low-gamma ASSR. Ten studies primarily addressed adulthood and aging, including those that compared young, middle-aged, and older adult groups (Dimitrijevic et al., 2004, Dobri et al., 2023, Farahani et al., 2020b, Goossens et al., 2016; Griskova-Bulanova et al., 2013; Johnson et al., 1988; Leigh-Paffenroth and Fowler, 2006; Purcell et al., 2004; Ross, 2018; Tlumak et al., 2015), and reflecting non-uniform ageing effects, with 40-Hz synchronization ranging from diminished to preserved or even increased. Three studies employed a longitudinal design (McKeon et al., 2024, Poulsen et al., 2009, Usui et al., 2023), showing mixed findings in adolescent and adult participants. The remaining works involved adult cross-sectional or broad lifespan samples, ranging from early childhood or adolescence into late adulthood (Aoyagi et al., 1994, Edgar et al., 2017, Edgar et al., 2018, Griskova-Bulanova et al., 2020, Kim et al., 2019, Poulsen et al., 2007, Rojas et al., 2006, Stapells et al., 1988, Tang et al., 2016), further contributing to the overall picture of nonlinear developmental and aging trajectories. Sample characteristics and reported ASSR-age relationships are summarized in Fig. 2. Only data from healthy participants were included in this visual synthesis.Fig. 2. Summary of study characteristics: from the left, total sample size, sex distribution, age span and reported ASSR-age relationship. Only healthy cohorts are shown. Age spans were taken directly from reported minimum–maximum values when available or estimated as mean ± SD. ASSR-age relationships are shown for amplitude (left) and phase-based (right) measures separately. Red (positive) and blue (negative) represent ASSR increase and decrease with age (based either on correlations or between-group comparisons), respectively. Beige (mixed category) refers to studies showing non-linear relationships or variable results when comparing multiple age groups. Studies reporting null relationships between ASSR and age are labeled in grey (null). White asterisk marks the studies that assessed ASSR-age relationship in a combined sample of patients and controls. Edgar et al. (2018) was not included in the plot due to largely overlapping healthy participant sample with a previous study (Edgar et al., 2017). For more details, refer to Results section and Table 1.Fig. 2
Additionally, 11 studies included clinical or neurodivergent populations alongside healthy controls. These comprised individuals with autism spectrum disorder (Ahlfors et al., 2024, Arutiunian et al., 2023, Darrell et al., 2025, Edgar et al., 2016, Stroganova et al., 2020), Rett syndrome (Neklyudova et al., 2024), 22q11.2 deletion syndrome (Larsen et al., 2018, Mancini et al., 2022), and schizophrenia-spectrum disorders (Edgar et al., 2018, Griskova-Bulanova et al., 2020, Kim et al., 2019).
Across the reviewed studies, 18 reported positive associations between age and ASSR measures, i.e., increased amplitude and/or phase-locking with age (Adalilar et al., 2025, Ahlfors et al., 2024, Arutiunian et al., 2023, Darrell et al., 2025, Dobri et al., 2023, Edgar et al., 2016, Herdman, 2011, Manasevich et al., 2025, Mancini et al., 2022, Maurizi et al., 1990, Neklyudova et al., 2021, Neklyudova et al., 2024, Ono et al., 2020, Roberts et al., 2021, Ross, 2018, Stapells et al., 1988, Stroganova et al., 2020, Tang et al., 2016), 5 reported negative associations (Edgar et al., 2017, Edgar et al., 2018; Griskova-Bulanova et al., 2013; Larsen et al., 2018; Leigh-Paffenroth and Fowler, 2006), and 8 found no significant age effects (Dimitrijevic et al., 2004, Goossens et al., 2016, Griskova-Bulanova et al., 2020, Johnson et al., 1988, Kim et al., 2019, Purcell et al., 2004, Tlumak et al., 2015, Usui et al., 2023). Other studies described mixed findings, including different results for amplitude and frequency-based measures (Arutiunian et al., 2022, Irazabal et al., 2024), non-linear ASSR trajectories with age (McKeon et al., 2024, Rojas et al., 2006) or divergent ASSR changes depending on age groups (Aoyagi et al., 1994, Cho et al., 2015, Farahani et al., 2020b).
ASSRs were most frequently elicited by 40-Hz amplitude-modulated (AM) tones (Adalilar et al., 2025, Ahlfors et al., 2024, Aoyagi et al., 1994, Arutiunian et al., 2022, Arutiunian et al., 2023, Dobri et al., 2023, Edgar et al., 2016, Edgar et al., 2017, Edgar et al., 2018, Farahani et al., 2020b, Goossens et al., 2016, Herdman, 2011, Leigh-Paffenroth and Fowler, 2006, Mancini et al., 2022, Ono et al., 2020, Ross, 2018, Tlumak et al., 2015) or click trains (Cho et al., 2015, Darrell et al., 2025; Griskova-Bulanova et al., 2013; Johnson et al., 1988; Kim et al., 2019; Larsen et al., 2018; Manasevich et al., 2025; Maurizi et al., 1990; McKeon et al., 2024; Neklyudova et al., 2021, Neklyudova et al., 2024; Rojas et al., 2006; Stapells et al., 1988; Stroganova et al., 2020; Usui et al., 2023). Several studies employed multi-frequency sweeps or chirp stimuli (Irazabal et al., 2024, Griskova-Bulanova et al., 2020, Purcell et al., 2004, Roberts et al., 2021, Tang et al., 2016), or compared AM and frequency-modulated (FM) tones to examine resonance profiles across the gamma range (Dimitrijevic et al., 2004, Poulsen et al., 2007, Poulsen et al., 2009).
EEG was used in 28 studies, either employing lower density setups of 1–28 electrodes (Aoyagi et al., 1994, Dimitrijevic et al., 2004, Irazabal et al., 2024, Griskova-Bulanova et al., 2020, Johnson et al., 1988, Leigh-Paffenroth and Fowler, 2006, Maurizi et al., 1990, Neklyudova et al., 2024, Purcell et al., 2004, Stapells et al., 1988, Tlumak et al., 2015) or higher density recordings with 32–256 channels (Adalilar et al., 2025, Cho et al., 2015, Darrell et al., 2025, Edgar et al., 2017, Edgar et al., 2018, Farahani et al., 2020b, Goossens et al., 2016, Kim et al., 2019, Larsen et al., 2018, Manasevich et al., 2025, Mancini et al., 2022, McKeon et al., 2024, Neklyudova et al., 2021, Poulsen et al., 2007, Poulsen et al., 2009, Usui et al., 2023), while MEG was applied in 14 studies (Ahlfors et al., 2024, Arutiunian et al., 2022, Arutiunian et al., 2023, Dobri et al., 2023, Edgar et al., 2016, Edgar et al., 2017, Edgar et al., 2018, Herdman, 2011, Ono et al., 2020, Roberts et al., 2021, Rojas et al., 2006, Ross, 2018, Stroganova et al., 2020, Tang et al., 2016), employing 151–306-channel whole-head systems.
At the analysis level, 26 studies reported sensor-space measures, typically at frontocentral electrodes such as Fz, FCz or Cz (Aoyagi et al., 1994, Cho et al., 2015, Darrell et al., 2025, Dimitrijevic et al., 2004, Edgar et al., 2017, Irazabal et al., 2024, Goossens et al., 2016, Griskova-Bulanova et al., 2020, Johnson et al., 1988, Kim et al., 2019, Larsen et al., 2018, Leigh-Paffenroth and Fowler, 2006, Manasevich et al., 2025, Mancini et al., 2022, Maurizi et al., 1990, McKeon et al., 2024, Neklyudova et al., 2021, Neklyudova et al., 2024, Poulsen et al., 2007, Poulsen et al., 2009, Purcell et al., 2004, Rojas et al., 2006, Stapells et al., 1988, Tlumak et al., 2015, Usui et al., 2023), but also temporal-parietal electrodes (Adalilar et al., 2025), while 16 conducted source-space analyses (Ahlfors et al., 2024, Arutiunian et al., 2022, Arutiunian et al., 2023, Dobri et al., 2023, Edgar et al., 2016, Edgar et al., 2017, Edgar et al., 2018, Farahani et al., 2020b, Herdman, 2011, Ono et al., 2020, Poulsen et al., 2007, Poulsen et al., 2009, Roberts et al., 2021, Ross, 2018, Stroganova et al., 2020, Tang et al., 2016). Nearly all MEG investigations localized activity to superior temporal regions and several demonstrated right-hemisphere dominance or asymmetric developmental trajectories (Ahlfors et al., 2024, Arutiunian et al., 2022, Edgar et al., 2016, Ono et al., 2020, Roberts et al., 2021).
Discussion
4
This review indicates that gamma-range ASSRs follow a broadly non-linear lifespan pattern, with robust developmental strengthening, relative stabilization in early adulthood, and heterogeneous changes in aging. However, coverage across the full lifespan remains uneven, with sparse data in early childhood, late adulthood, and longitudinal designs, limiting precise delineation of maturation and decline trajectories. Consequently, our understanding of how gamma-range ASSRs evolve across the lifespan remains incomplete and biased toward developmental stages, underscoring the need for harmonized longitudinal and lifespan-spanning investigations. To contextualize this uneven coverage, findings are summarized below across three main domains: developmental trajectory, adulthood and aging, and clinical or neurodivergent populations.
Developmental trajectory (childhood to early adulthood)
4.1
Developmental studies indicate that the strength of ASSRs increases markedly from infancy through adolescence. Evidence from fetal MEG further suggests that the neural capacity for rhythmic auditory synchronization begins to emerge before birth (Niepel et al., 2020): steady-state responses were detected in fetuses between 30 and 38 weeks of gestation at 27 Hz, whereas responses around 42 Hz were absent, indicating that higher-frequency gamma synchronization develops only later in postnatal life.
The earliest post-natal investigations, including infant cohorts (Aoyagi et al., 1994, Maurizi et al., 1990, Stapells et al., 1988) demonstrated weak or unreliable 40-Hz responses during the first months of life, followed by progressive strengthening across early childhood. Subsequent studies of typically developing children and adolescents (e.g., Ahlfors et al., 2024; Edgar et al., 2016) reported positive correlations between age and response magnitude or phase-locking, frequently with right-hemisphere predominance (e.g., Arutiunian et al., 2022; Ono et al., 2020). Group-based comparisons further demonstrated higher amplitudes in adolescents or adults relative to younger children (e.g., Herdman, 2011; Neklyudova et al., 2021), reinforcing a picture of gradual refinement of neural synchrony during early development. Longitudinal data also support this trajectory: in a repeated-measures design, Usui et al. (2023) found stable 40-Hz responses between mid-adolescent assessments, suggesting that the major phase of ASSR maturation likely concludes by early adolescence.
Similar developmental increases in gamma activity have been observed in resting-state and task-based EEG/MEG studies, consistent with progressive maturation of inhibitory circuitry and excitation–inhibition balance (Candelaria-Cook et al., 2022, McKeon et al., 2023) and reflecting progressive stabilization, inhibitory refinement, and efficiency of cortical networks (Rhodes et al., 2025). These developmental changes have been attributed to the maturation of inhibitory (GABAergic) circuitry and refinement of excitation–inhibition balance (Gogolla et al., 2009, Kilb, 2012, Uhlhaas and Singer, 2010), as well as structural processes such as myelination and synaptic pruning that enhance long-range synchrony and network efficiency (Fair et al., 2009, Giedd et al., 2008, Whitford et al., 2007).
Collectively, these findings delineate a developmental trajectory that begins with weak or inconsistent gamma synchronization in infancy, progresses through childhood and adolescence, and stabilizes in early adulthood as auditory cortical circuits reach functional maturity, providing a continuous framework for interpreting the lifespan changes described below.
Aging trajectory (adulthood and aging)
4.2
In adulthood and aging, the trajectory of gamma-range ASSRs becomes increasingly heterogeneous. Several studies reported attenuated 40-Hz responses with advancing age (e.g., Griskova-Bulanova et al., 2013; Kim et al., 2019). Conversely, other investigations demonstrated enhanced amplitude or phase-locking in older adults (e.g., Dobri et al., 2023; Ross, 2018). The majority of studies found no significant age-related differences in 40-Hz responses (e.g., Goossens et al., 2016; Purcell et al., 2004).
Comparable age-related patterns have been observed across broader indices of gamma-band activity, with resting-state or task-based EEG/MEG studies showing that gamma power and peak frequency tend to decline with advancing age (Güntekin et al., 2023, Murty et al., 2020, Park et al., 2022). These findings are consistent with age-related alterations in inhibitory tone and cortical synchronization (Heise et al., 2022, Porges et al., 2021).
Beyond neurochemical and network-level factors, peripheral and subcortical auditory changes also contribute to age-related variability in ASSRs. Cochlear synaptopathy and brainstem dysfunction reduce hearing sensitivity and temporal precision, weakening early auditory coding (Ouda et al., 2015, Parthasarathy and Kujawa, 2018, Sergeyenko et al., 2013). Notably, the integrity of both peripheral hearing and cortical structure has been identified as essential for robust ASSRs, with smaller auditory cortical volumes and compromised white-matter connectivity predicting weaker gamma synchronization (Kim et al., 2019, Koshiyama et al., 2024, Schuler et al., 2022). Neuroimaging evidence further demonstrates diminished auditory cortical activation and reorganization of tonotopic maps in aging (Profant et al., 2015), alongside gray- and white-matter alterations in auditory and association regions reflecting large-scale cortical remodeling across the lifespan (de Mooij et al., 2018). These sensory and structural factors, together with neurochemical decline, likely interact with cortical E/I dynamics, amplifying the interindividual variability observed in aging cohorts.
Collectively, these findings suggest that, rather than a uniform decline, the adult–elderly transition in gamma synchronization is characterized by greater interindividual variability and regionally differentiated modulation of cortical oscillations. This variability likely reflects the combined influence of neurochemical decline, compensatory network recruitment, and methodological diversity across studies, underscoring that age-related changes in ASSRs are not purely degenerative but represent a complex reorganization of auditory cortical dynamics with aging.
Clinical populations and atypical development
4.3
Studies involving clinical or neurodivergent populations provide additional insight into how the normative age-related trajectory of 40-Hz ASSRs may be altered by neurodevelopmental or neuropsychiatric conditions affecting cortical synchronization.
In autism spectrum disorder (ASD), several studies have reported positive correlations between age and 40-Hz ASSR power or phase-locking (e.g., Arutiunian et al., 2023; Darrell et al., 2025). However, these associations are typically weaker, delayed, or lateralized compared with typically developing controls (Ono et al., 2020; Ahlfors et al., 2024). In studies that included 22q11.2 deletion syndrome (Mancini et al., 2022) and Rett syndrome (Neklyudova et al., 2024) patients, positive age–ASSR relationships were observed only in healthy participants. Although the available evidence remains limited, these findings suggest that the typical age-related strengthening of gamma synchronization may be delayed or dysregulated in neurodevelopmental conditions, reflecting atypical maturation and E/I balance within auditory cortical networks. This interpretation is further supported by recent animal-model findings: in Fragile X and PTEN-deletion mouse models, the maturation of 40-Hz temporal processing is delayed or dysregulated, showing region- and sex-specific differences (Croom et al., 2023, Croom et al., 2024, Croom et al., 2024).
Similarly, in schizophrenia-spectrum disorders, Kim et al. (2019), Griskova-Bulanova et al. (2020), and Edgar et al. (2018) reported no significant associations between age and ASSR measures. Meta-analytic evidence supports this interpretation, showing reductions in 40-Hz ASSR amplitude and phase-locking with only weak age dependence, manifesting as slightly larger deficits in younger patients and a developmentally anchored disruption of gamma synchronization (Thuné et al., 2016, Zouaoui et al., 2023).
Taken together, findings across clinical groups demonstrate that disturbances in NMDA-receptor-mediated E/I balance can delay, attenuate, or uncouple the typical age-related evolution of the 40-Hz ASSR. These deviations from the normative trajectory provide a sensitive window into atypical neurodevelopment and the pathophysiology of altered cortical synchronization across neuropsychiatric conditions.
Methodological and interpretative considerations
4.4
Several methodological and interpretative considerations should be acknowledged when evaluating the present synthesis.
First, sex distribution was not uniformly balanced across studies, and detailed sex-specific analyses were infrequently reported. Given accumulating evidence that gamma-band synchronization and excitation–inhibition balance may be modulated by biological sex and sex-steroid fluctuations (Anazawa et al., 2023, Jasinskyte et al., 2023, Hyer et al., 2018, Griskova-Bulanova et al., 2014, Melynyte et al., 2018), uneven sex ratios and the absence of control for hormonal status (e.g., menstrual cycle phase, contraceptive use, or menopausal status) may contribute to variability in reported ASSR–age relationships. Because such factors were rarely documented in the primary literature, their potential influence could not be systematically evaluated within the present review.
Second, although the synthesis focused primarily on age-related effects in healthy populations, a proportion of the included healthy samples were derived from control groups in clinical case–control studies. In such designs, matching procedures typically prioritize age and sex, whereas other demographic variables (e.g., educational level, socioeconomic status, or subclinical traits) are not consistently reported. As a result, the extent to which these factors may have influenced reported ASSR–age relationships remains unclear.
Third, variability existed across studies in participant state and task demands during ASSR recording. Recordings were conducted under passive listening (with or without distraction), active attention, or sleep conditions. Given that gamma-band synchronization is sensitive to arousal (Griskova et al., 2007, Wang et al., 2019) and attentional engagement (Matulyte et al., 2024), differences in recording context may have contributed to heterogeneous findings. However, systematic monitoring and reporting of vigilance state were infrequent, limiting cross-study comparability.
Fourth, methodological heterogeneity across stimulation parameters, recording configurations, and analytic approaches constrains direct comparison of findings. Variability in modulation frequency, carrier type, number of trials, sensor versus source analysis, and the selection of amplitude- versus phase-based metrics complicates efforts to delineate precise normative trajectories, and based on observed mixed results when different paradigms were used in the same study (Poulsen et al., 2007, Poulsen et al., 2009), could be a substantial source of variability. Although such diversity reflects the evolving nature of the field, it restricts the possibility of quantitative integration and standardization.
Fifth, most available evidence is cross-sectional. Inferred nonlinear lifespan patterns should therefore be interpreted cautiously, as cohort effects cannot be excluded. While cross-sectional comparisons provide important initial mapping, harmonized longitudinal paradigms spanning contiguous developmental and aging stages are needed to delineate trajectories of maturation, stabilization, and decline in gamma synchronization.
Finally, integration of ASSR measures with multimodal approaches, including structural imaging, magnetic resonance spectroscopy, and behavioral assessments, remains limited but may be helpful in clarifying the neurobiological mechanisms underlying lifespan changes in cortical synchronization.
Together, these considerations underscore the importance of improved methodological harmonization and comprehensive reporting. Such advances may support the development of standardized ASSR protocols suitable for both cross-sectional and longitudinal investigations, thereby strengthening the role of gamma-range ASSRs as a mechanistic biomarker across the human lifespan.
Generalization
4.5
Across the lifespan, the auditory gamma-range steady-state responses follow a non-linear developmental trajectory (Arutiunian et al., 2022, Cho et al., 2015, Edgar et al., 2016). When integrated with recent fetal evidence (Niepel et al., 2020), the available data now trace this trajectory from the late prenatal period through senescence, highlighting a lifespan continuum of oscillatory maturation and decline. During childhood and adolescence, 40-Hz ASSRs show progressive strengthening of amplitude and phase coherence, reflecting the maturation of auditory cortical circuits and refinement of excitation–inhibition balance (Candelaria-Cook et al., 2022, McKeon et al., 2023, Uhlhaas and Singer, 2010). This maturation appears to plateau by early adulthood, after which findings become more heterogeneous (Usui et al., 2023). In adulthood and aging, results diverge: some studies report attenuation of ASSR amplitude or phase consistency (Edgar et al., 2017; Griskova-Bulanova et al., 2013), others find enhanced responses (Dobri et al., 2023, Ross, 2018), and several observe no clear changes (Goossens et al., 2016, Tlumak et al., 2015). Such variability may stem from differences in individual variation in cortical plasticity, neurochemical balance, and compensatory recruitment (ElShafei et al., 2020, Murty et al., 2020) combined with methodological aspects.
Understanding lifespan changes in ASSRs has important translational implications. Establishing normative developmental and aging trajectories enables the differentiation of pathological alterations from age-appropriate variability, improving the interpretability of ASSR abnormalities in clinical contexts such as schizophrenia (Thuné et al., 2016, Zouaoui et al., 2023), autism spectrum disorder (Darrell et al., 2025, Seymour et al., 2020), and dementia (Mao et al., 2025, van Deursen et al., 2011). Moreover, by delineating when cortical synchronization is most plastic or vulnerable, lifespan data can inform the timing and personalization of interventions targeting gamma activity, whether through auditory or multi-modal stimulation (Chen et al., 2025, Olson, 2021), pharmacological modulation (Homma et al., 2025) or noninvasive brain stimulation (Mockevičius et al., 2025). Ultimately, integrating lifespan and clinical evidence will advance the use of ASSRs as a mechanistic biomarker for monitoring neural network integrity and treatment response across neurodevelopmental, psychiatric, and neurodegenerative conditions.
CRediT authorship contribution statement
Aurimas Mockevičius: Writing – review & editing, Visualization, Validation, Methodology. Danylo Machevskyi: Writing – review & editing, Investigation. Dariusz Majcherczyk: Writing – review & editing, Investigation. Inga Griškova-Bulanova: Writing – review & editing, Writing – original draft, Visualization, Methodology, Conceptualization.
Declaration of Generative AI and AI-assisted technologies in the writing process
During the preparation of this work, the authors used ChatGPT and Grammarly to improve language and readability. After using this tool/service, the authors reviewed and edited the content as needed and take full responsibility for the content of the publication.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Adalilar I.Gransier R.Wouters J.van Wieringen A.Response strength and latencies of auditory steady-state responses from age six Int. J. Audiol.647202573574410.1080/14992027.2024.241999839773278 · doi ↗ · pubmed ↗
- 2Ahlfors S.P.Graham S.Bharadwaj H.Mamashli F.Khan S.Joseph R.M.Losh A.Pawlyszyn S.Mc Guiggan N.M.Vangel M.Hämäläinen M.S.Kenet T.No differences in auditory steady-state responses in children with autism spectrum disorder and typically developing children J. Autism Dev. Disord.54520241947196010.1007/s 10803-023-05907-w 36932270 PMC 11463296 · doi ↗ · pubmed ↗
- 3Anazawa S.Yamashiro K.Makibuchi T.Ikarashi K.Fujimoto T.Ochi G.Sato D.Sex differences in excitatory and inhibitory function in the primary somatosensory cortex during the early follicular phase: a preliminary study Brain Sci.135202376110.3390/brainsci 1305076137239233 PMC 10216438 · doi ↗ · pubmed ↗
- 4Aoyagi M.Kiren T.Furuse H.Fuse T.Suzuki Y.Yokota M.Koike Y.Effects of aging on amplitude-modulation following response Acta Oto-Laryngol. Suppl. 5111994152210.3109/000164894091282958203219 · doi ↗ · pubmed ↗
- 5Arutiunian V.Arcara G.Buyanova I.Gomozova M.Dragoy O.The age-related changes in 40 Hz auditory steady-state response and sustained event-related fields to the same amplitude-modulated tones in typically developing children: a magnetoencephalography study Hum. Brain Mapp.431720225370538310.1002/hbm.2601335833318 PMC 9812253 · doi ↗ · pubmed ↗
- 6Arutiunian V.Arcara G.Buyanova I.Davydova E.Pereverzeva D.Sorokin A.Tyushkevich S.Mamokhina U.Danilina K.Dragoy O.Neuromagnetic 40 Hz Auditory Steady-State Response in the left auditory cortex is related to language comprehension in children with Autism Spectrum Disorder Prog. Neuro-Psychopharmacol. Biol. Psychiatry 122202311069010.1016/j.pnpbp.2022.11069036470421 · doi ↗ · pubmed ↗
- 7Başar-Eroglu C.Strüber D.Schürmann M.Stadler M.Başar E.Gamma-band responses in the brain: A short review of psychophysiological correlates and functional significance Int. J. Psychophysiol.241199610111210.1016/S 0167-8760(96)00051-78978437 · doi ↗ · pubmed ↗
- 8Buzsáki G.Wang X.-J.Mechanisms of gamma oscillations Annu. Rev. Neurosci.35201220322510.1146/annurev-neuro-062111-15044422443509 PMC 4049541 · doi ↗ · pubmed ↗