Mangrove cyanobacterial diversity as a source of bioactive natural products
Arup Ratan Roy, Shayontani Basu, Sergio de los Santos Villalobos, Joydeep Mukherjee

TL;DR
This paper reviews cyanobacterial diversity in mangroves and their potential to produce useful natural products like drugs and biofuels.
Contribution
It is the first global review connecting cyanobacterial taxonomy with their bioactive metabolites in mangrove ecosystems.
Findings
Mangroves host high cyanobacterial diversity, including novel taxa.
Cyanobacteria produce diverse bioactive compounds like antimicrobials and anticancer agents.
Modern taxonomic methods are essential for identifying cyanobacterial strains.
Abstract
•First global review linking polyphasic taxonomy to cyanobacterial metabolites in mangroves.•Catalogs cyanobacterial strains recognized only by modern morphological and molecular data.•Reveals high cyanobacterial diversity across mangrove ecosystem, including novel taxa.•Highlights diverse bioactivities: antimicrobial, anticancer, toxins, bioplastic.•Bridges taxonomy with biotech applications in drugs, agrochemicals, dyes, biofuels. First global review linking polyphasic taxonomy to cyanobacterial metabolites in mangroves. Catalogs cyanobacterial strains recognized only by modern morphological and molecular data. Reveals high cyanobacterial diversity across mangrove ecosystem, including novel taxa. Highlights diverse bioactivities: antimicrobial, anticancer, toxins, bioplastic. Bridges taxonomy with biotech applications in drugs, agrochemicals, dyes, biofuels. Extensive…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
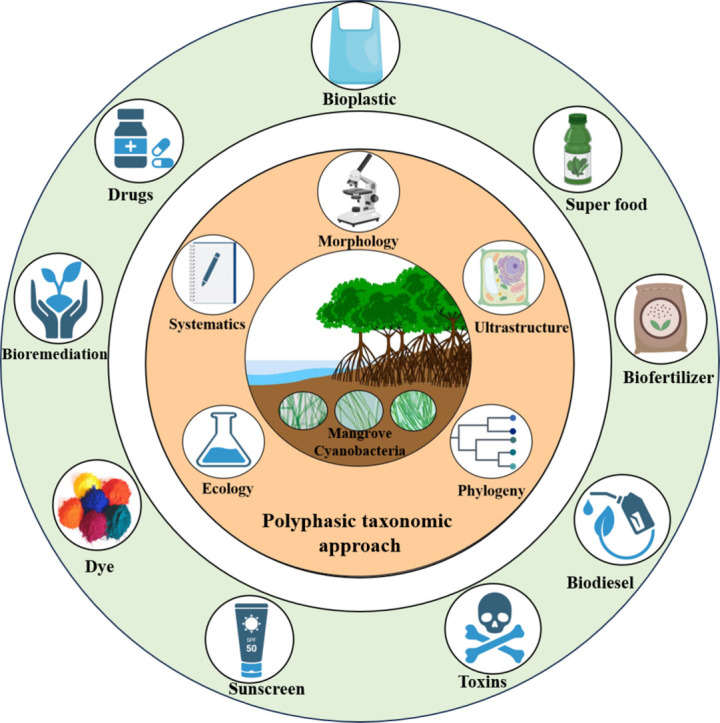 Figure 1
Figure 1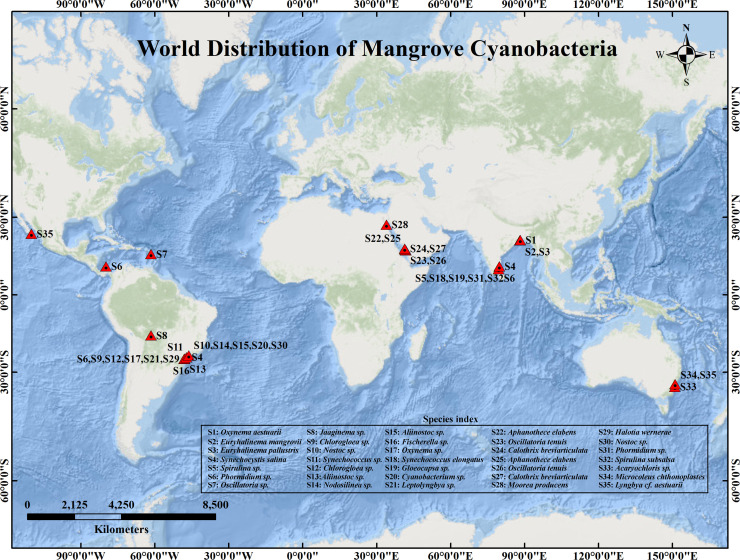 Figure 2
Figure 2 Figure 3
Figure 3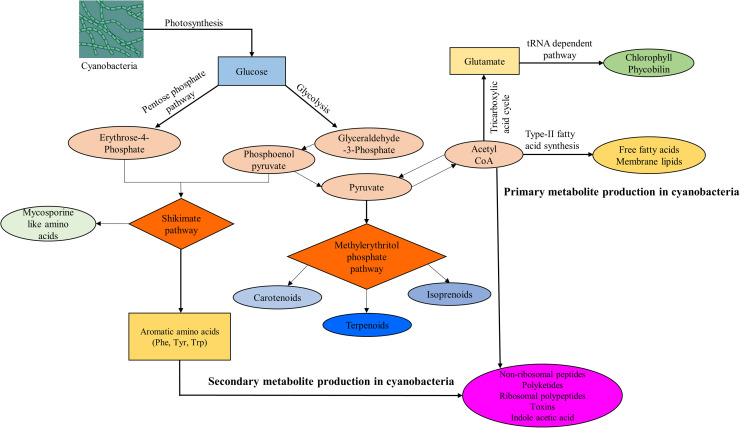 Figure 4
Figure 4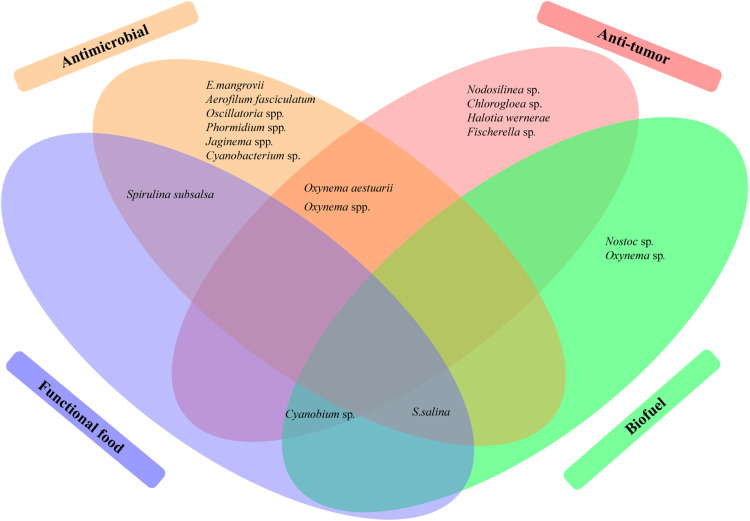 Figure 5
Figure 5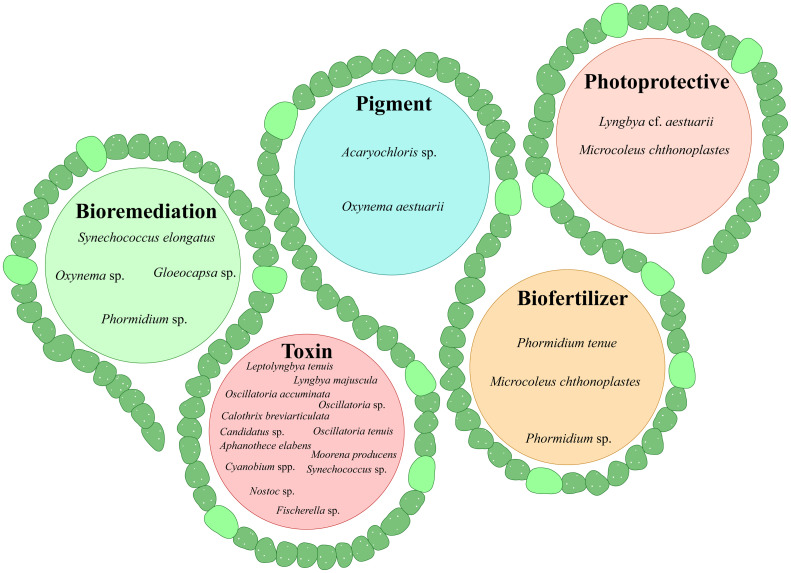 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Natural Products and Biosynthesis · Seaweed-derived Bioactive Compounds · Marine and coastal plant biology
Introduction
1
Mangroves are unique, highly susceptible, and unevenly distributed ecosystems located along the shorelines of tropical and subtropical regions, hosting a variety of genetically and morphologically diverse organisms. Various microbial communities that are adapted to survive the unstable ecological conditions prevalent in these ecotones reside here by virtue of the abundance of inorganic nutrients and carbon (Giri et al., 2011; Pramanik et al., 2011). These include mainly physical factors such as salinity, temperature, sunlight, availability of gases, tidal fluctuations, and moisture (Rigonato et al., 2013). Cyanobacteria are ancient, oxygenic photosynthetic prokaryotes that played a fundamental role in the evolution of Earth’s atmosphere and continue to be key contributors to global carbon and nitrogen cycles (Sánchez-Baracaldo and Cardona, 2020). Owing to their remarkable ecological diversity across freshwater, marine, terrestrial, mangrove, and intertidal ecosystems, as well as extreme environments such as deserts, hot springs, hypersaline lakes, and polar regions, and their ability to produce bioactive secondary metabolites, they have gained significant attention for biotechnological and environmental applications (Whitton and Potts, 2012). Cyanobacteria derived from mangrove are a notable group of simple filamentous or unicellular, Gram-negative, oxygen-evolving prokaryotes, present globally in virtually all well-illuminated terrestrial and aquatic habitats (Thatoi et al., 2013; Alvarenga et al., 2015) (Fig. 1). Though a primitive group of organisms, they have evolved adaptive strategies to survive environmental extremes such as high temperatures, prolonged UV exposure, hypersaline waters, arid conditions, and freezing temperatures (Alvarenga et al., 2015; Joset et al., 1996). Years of extensive research indicate that cyanobacteria from mangrove ecosystems are abundant sources of both established and novel bioactive compounds like dolastatins, scytonemins, microcystins, and xanthenes, with broad industrial relevance (Pramanik et al., 2011; Thatoi et al., 2013). In the mangrove intertidal zones, at the intersection of land, sea, and air, cyanobacteria endure regular shifts in salinity and tidal immersion, spending roughly half their life cycle exposed to dry, aerial conditions and the other half submerged in brackish water (Zhang et al., 2013) (Fig. 2A and B). All these distinctive features ensure the competitive success of mangrove cyanobacteria by inducing the production of unique bioactive molecules that are quite different from those produced by organisms inhabiting stable ecosystems (Pramanik et al., 2011; Jones et al., 2021). Despite their importance in ecosystem services as well as on a commercial scale for the manufacture of a host of products, relatively few reports on cyanobacterial NPs, especially from the mangrove and marine ecosystems, are currently available (Alvarenga et al., 2015). Detailed reports on bioactive compounds derived from mangrove cyanobacteria are limited to certain organisms only, which are described below. This is probably due to the use of traditional screening of naturally occurring cyanobacterial populations in the mangrove habitat (without the use of polyphasic taxonomy) for desirable compounds, which tend to be disorganized. As a result, expensive retrieval of previously known compounds rather than the discovery of novel molecules is fairly common (Bull et al., 1992). The polyphasic characterization of mangrove cyanobacteria increases our understanding of the mangrove microflora composition and, in turn, leads to a better realization of their potential applicability across various industries.Fig. 1. Global distribution of cyanobacterial genera in mangrove habitats. The genera are referred to as S1 to S35 in the map and the respective genera are explained in the key within the map.Fig 1: dummy alt textFig. 2(A and B) Cyanobacterial biofilm formed on the surface of soil sediments of the Sundarbans, India the world’s largest intertidal mangrove forest ecosystem. (C and D) Cyanobacterial biofilm formed on the surface of mangrove roots and soil sediments in the coastal areas of South Florida, USA. The arrow pointed at the cyanobacterial mats. (Image A, B reproduced from (Balu et al., 2020), copyright license No. 5237690888906 and image C, D reproduced from (Berthold et al., 2021), copyright license No. 5237000719750.Fig 2 dummy alt text
The present study is a systematic review undertaken to collate reported data on the mangrove cyanobacterial secondary metabolites of commercial importance and the justification of the taxonomic position of these cyanobacteria following the modern polyphasic approach.
Taxonomy of mangrove cyanobacteria using a polyphasic approach
2
Cyanobacteria are among the oldest and most adaptable life forms, thriving in extreme places and in mangrove forests, encompassing various families (Abed et al., 2009). Extended evolution has produced great phylogenetic and metabolic variety in cyanobacteria, which produce unique biomolecules that are valuable for biotechnology (Dahms et al., 2006; Bhadury and Wright, 2004). Study of extremophilic cyanobacteria to understand their taxonomy, physiology and metabolic strategies for surviving harsh conditions is gradually gaining importance. Polyphasic taxonomy, enriched with ecological and biogeographic information, has yielded novel cyanobacterial genera and species, which is of paramount importance for discovering new enzymes, proteins, biopolymers, and potential drugs (Zammit et al., 2012; Zammit, 2018; Zammit et al., 2023). For measuring species-level alpha diversity in mangroves, researchers use a polyphasic approach that combines 16S rRNA and 16S–23S ITS sequence analyses with morphology, ultrastructure, and ecological information (Iteman et al., 2000; Nübel et al., 1997; Boyer et al., 2001; Casamatta et al., 2006; Berthold et al., 2021; Komárek, 2016;Komárek, 2018). Since cyanobacteria are morphologically plastic, this combined approach is essential to separate cryptic, morphological, or ecological taxa. Modern phylogeny has been enhanced by phylogenomics, which utilizes large-scale genetic data for exploring evolutionary relationships among cyanobacteria (Zammit et al. 2023). Addition of phylogenomics to polyphasic taxonomy has improved estimation of species limits, uncovered new orders and families, and resolved unclear genus relationships among cyanobacteria (Strunecký et al., 2023). Recent notable examples include the Indian Sundarbans mangrove cyanobacteria Oxynema aestuarii, Euryhalinema sp., Aerofilum fasciculatum, Leptoelongatus littoralis, and Almyronema epifania (Chakraborty et al., 2018, 2019, 2021; Roy et al., 2024) and Picosynechococcus mangrovensis, Sirenicapillaria, and Allocoleopsis from the Andaman coast mangrove environment of Thailand (Lim Chun Ginn et al., 2025). However, applying polyphasic identification without culturing individual cyanobacterial strains is challenging. Only a few mangrove-derived strains have been described completely by the polyphasic methods (Table 1), revealing significant lacunae in alpha-diversity studies.Table 1. Globally reported novel genus and species described from the mangrove ecosystem based on polyphasic taxonomic identification.Table 1: dummy alt textCyanobacteria strainSamples collection habitatCountryReferencesAerofilum fasciculatumSoil sedimentIndiaChakraborty et al., 2021Almyronema epifaneiaSoil sedimentIndiaRoy et al., 2024Candidatus Planktothricoides nigerBiofilm on the sediment from the periphytonGuadeloupeGuidi-Rontani et al., 2014Candidatus Planktothricoides roseaBiofilm on the sediment from the periphytonGuadeloupeGuidi-Rontani et al., 2014Dapisostemon apicaliramisBiofilmBrazilHentschke et al., 2016Euryhalinema mangroviiSoil sedimentIndiaChakraborty et al., 2019Euryhalinema pallustrisSoil sedimentIndiaChakraborty et al., 2021Foliisarcina bertiogensisLeaves of Avicennia schauerianaBrazilAlvarenga et al., 2016Halotia werneraeSoil sedimentBrazilGenuário et al., 2015Halotia longisporaBrackish waterBrazilGenuário et al., 2015Halotia branconiiSoil sedimentBrazilGenuário et al., 2015Kryptousia macronemaLeaves of Avicennia schaueriana and Merostachys neesiiBrazilAlvarenga et al., 2017Kryptousia microlepisLeaves of Avicennia schaueriana and Merostachys neesiiBrazilAlvarenga et al., 2017Leptolyngbya indicaSoil sedimentIndiaDebnath et al., 2017DLeptoelongatus litoralisSoil sedimentIndiaChakraborty et al., 2019Leptochromothrix engeneiBiofilm from sand sedimentUSABerthold et al., 2021Leptolyngbya sp.Soil sediment, Marine waterBrazilSilva et al., 2014Nodosilinea sp.Soil sediment, Marine waterBrazilSilva et al., 2014Oxynema sp.Soil sediment, Marine waterBrazilSilva et al., 2014Oxynema aestuariiSoilIndiaChakraborty et al., 2018Okeania hirsutaMarine waterPanamaEngene et al., 2013Ophiophycus aerugineusBiofilm from sand sedimentUSABerthold et al., 2021Phyllonema aviceniicolaLeaves of Avicennia schauerianaBrazilAlvarenga et al., 2016Streptostemon lutescensBiofilmBrazilHentschke et al., 2016Vermifilum ionodoliumBiofilm from sand sedimentUSABerthold et al., 2021
Overview of cyanobacterial biosynthetic pathways and strategies for upscaling metabolite production
3
Cyanobacterial processes such as photosynthesis and carotenogenesis produce both primary and secondary metabolites. Primary metabolites include antioxidants, structural proteins, and lipids, which support essential functions such as growth, reproduction, and cell division, and can be re‑engineered into products like dyes, biofertilizers, supplements, and bioplastics. Secondary metabolite production varies by species and environment, and commonly serves defensive roles. Examples of cyanobacterial secondary compounds include toxins, phenolics, alkaloids, and antibiotics (Żymańczyk-Duda et al., 2022). The main pathways for primary and secondary metabolites include the transfer ribonucleic acid (tRNA) dependent pathway, Type-II fatty acid synthesis pathway, shikimate pathway, and methylerythritol phosphate (MEP) pathway, with common intermediates and precursors produced by glycolysis and pentose phosphate pathway (Verma et al., 2022; Cao et al., 2023; Nagao et al., 2025). Fig. 3 gives a concise overview of the major biosynthetic pathways for the production of primary and secondary metabolites in cyanobacteria.Fig. 3. Core pathways for cyanobacterial metabolite biosynthesis, including tRNA-dependent, Type-II fatty acid, shikimate, and MEP pathways, from precursors generated by glycolysis and the pentose phosphate pathway.Fig 3: dummy alt text
Building on this metabolic framework, cyanobacterial natural-product discovery currently involves biosynthetic gene cluster (BGC) identification with compound characterization and scalable production. Approaches including OSMAC (one strain — many compounds) cultivation, native-host genetics, heterologous expression, and in vitro reconstitution are useful for the discovery of silent pathways and expansion of production options. Genomics, metabolomics, and machine-learning tools are used to accelerate dereplication, BGC-metabolite linking, and biotechnological exploitation (Pancrace et al., 2019; Baunach et al., 2024). High‑throughput OSMAC cultivation employing variable nutrients, trace elements, light, CO₂, temperature, desiccation, and chemical elicitors has resulted in conditional production of compounds such as anabaenopeptins, siderophores, and scytonemin (Soule et al., 2007; Demay et al., 2019). In model cyanobacteria such as Nostoc punctiforme PCC 73102, transcriptional reporters, forward/reverse mutagenesis, regulator overexpression, and shuttle plasmids may be used to enable BGC activation and mechanistic dissection (Dehm et al., 2019; Baunach et al., 2024; Krumbholz et al., 2022). Since many cyanobacteria yield low metabolite titers or are unfit for manipulation, heterologous routes in engineered E. coli, yeast, Streptomyces, or cyanobacterial hosts such as Anabaena sp. 7120 are used for complementary production and combinatorial chemistry options, though host choice remains empirical (Jones et al., 2012; Dhakal et al., 2021; Baunach et al., 2024). A notable example for enzyme engineering in the context of cyanobacteria associated with the mangrove environment is the development of transgenic Chlamydomonas reinhardtii by introducing the cyanobacterial cyanase (CYN) gene from Synechococcus elongates (a cyanobacteria with widespread occurrence in mangrove environments) resulting in strong gene overexpression. This engineered strain (TC. reinhardtii‑2) displayed enhanced growth, resilience, and efficient removal of cyanide, highlighting its potential as an eco-friendly bioremediation model for polluted freshwater environments (Sobieh et al., 2022). In vitro or chemo‑enzymatic “total biosynthesis” complements cell‑based methods for pathway elucidation and library generation when enzyme function and precursor chemistry are known, exemplified by aetokthonotoxin, cyanobacterin, and guanitoxin studies (Sattely et al., 2008; Adak et al., 2022; D’Agostino et al., 2022; Lima et al., 2022). Integrating genome mining, structural biology‑guided enzyme engineering, microfluidics‑compatible miniaturization, and life‑cycle considerations yields a robust framework that maximizes discovery, mechanistic insight, and routes toward sustainable biotechnological exploitation (Baunach et al., 2024).
Over the past decade, genomics, metabolomics, and bioinformatics have converged to upend natural‐product discovery in cyanobacteria (Dittmann et al., 2015; D’Agostino, 2023; Baunach et al., 2024). Automated platforms such as AntiSMASH predict and annotate BGCs, while spectral networks from Global Natural Products Social Molecular Networking (GNPS) and curated libraries like CyanoMetDB, NPAtlas, and LOTUS accelerate dereplication (Van Der Hooft et al., 2020; Jones et al., 2021; Van Santen et al., 2021). Emerging in silico tools, viz., peptidogenomics suites (NRPquest, MetaMiner, DeepRipp), correlation‐driven metabologenomics, and hybrid frameworks like NPLinker facilitate the application of machine learning to score and visualize BGC–metabolite relationships, rapidly expanding the known chemical space of cyanobacteria (Mohimani et al., 2018; Cao et al., 2019; Merwin et al., 2020). In-silico tools have also been proven useful to detect features relevant for genome editing, notably CRISPR-Cas in the mangrove cyanobacterium Oxynema aestuarii AP17. Multiple Type I and Type III CRISPR arrays were characterized in the genome of O. aestuarii AP17, along with a previously unreported Type I‑G cas operon, illustrating that mangrove cyanobacteria harbor novel CRISPR systems that could be harnessed for editing or anti‑phage strategies (Basu et al., 2023).
Genome‐scale metabolic models (GSMs) now support both systems biology and biotechnological engineering in cyanobacteria (Goodall et al., 2018; Gale et al., 2019). Updated models for Synechocystis PCC 6803, PCC 7942 (iJB785 and iJB792), and UTEX 2973 integrated photosynthetic electron transport, light capture, and O₂ evolution, demonstrating 78–98% agreement with experimental fluxes (Joshi et al., 2017; Broddrick et al., 2019; Gale et al., 2019). Comparative models uncovered strain‐specific carbon uptake and Calvin‐cycle flux differences, guiding in silico optimization of 2,3‐butanediol, n‐butanol (OptFlux), and limonene (OptForce) yields (Ranganathan et al., 2010; Rocha et al., 2010). Yet GSM refinement is limited by sparse proteomic, transcriptomic, and regulatory data (Gale et al., 2019). To close these gaps, the CyanoSource platform has been launched, with a standardized knockout library of over 3,400 genes in PCC 6803 using modular cloning and conditional alleles. Such resources will enhance functional annotation, reproducibility, and the scalable exploitation of mangrove cyanobacterial biosynthetic capacity.
Types of bioactive compounds produced by mangrove cyanobacteria
4
The taxonomic advances and the biosynthetic strategies outlined in the previous sections establish mangrove cyanobacteria as a valuable resource for natural product discovery. Their diverse lineages, backed by polyphasic taxonomy and phylogenomics, provide a strong basis for linking genetic identity with metabolite production. At the same time, modern approaches for biosynthetic pathway analysis and scaling methods demonstrate how mangrove cyanobacteria can be applied for utilization across various industries. Within this framework, the following section provides a comprehensive overview of the major classes of bioactive compounds reported from mangrove cyanobacteria, emphasizing their ecological functions and commercial relevance in antimicrobial, anti-inflammatory, toxin, photoprotective, pigment, and functional food applications. Table 2 and Fig. 4 display the major categories of NPs from various genera of mangrove cyanobacteria.Table 2. Overview of the bioactive compounds and beneficial primary metabolites produced by mangrove cyanobacteria.Table 2: dummy alt textStrainsCompound/ Extract andTaxonomic classificationReferencePlanktothrix mougeotii PMC 1114.19Godavarin K (Anti-helminthic)16S rRNA and morphologicalWang et al., 2023Oxynema aestuarii, Euryhalinema mangrovii, Euryhalinema pallustris, Aerofilum fasciculatum, Leptoelongatus littoralisMethanol extracts, Antimicrobial (Inhibition of S. aureus, E. coli, B. subtilis, P. aeruginosa)Polyphasic taxonomyPramanik et al., 2011; Chakraborty et al., 2018, 2019, 2021Synechocystis salina, Oscillatoria salina, Spirulina subsalsa, Phormidium fragile, Oscillatoria cortiana, Oscillatoria willei, Phormidium tenueMethanol / methanol:chloroform:water extracts, Antifungal (Inhibition of fungi F. solani, A. flavus)Morphological (Desikachary)Sakthivel and Kathiresan, 2013Synechococcus sp., Gloeocapsa sp.Aqueous extracts Antimicrobial (inhibit P. vulgaris and P. aeruginosa)16S rDNAAnburaj et al., 2020Oscillatoria sp., Jaaginema sp.Methanol:acetonitrile:water extracts, Antimicrobial Inhibit E. coli and P. atlantica16S rRNADuperron et al., 2019Cyanobium, Chlorogloea, Cyanobacterium, Oxynema, Synechococcus, Leptolyngbya, Nodosilinea, NostocEthyl acetate, chloroform, methanol extracts, Antimicrobial (inhibit B. cereus, S. typhimurium and Candida sp*.)16S rRNASilva et al., 2014; Silva-Stenico et al., 2014Fischerella* sp., Aliinostoc sp.Methanol extracts (Inhibition of Staphylococcus sp. and Candida albicans)16S rRNAShishido et al., 2019Oxynema sp. AP17Ethyl acetate extract (anti-angiogenic)Polyphasic taxonomyBasu et al., 2021Cyanobium sp., Oxynema sp.Ethanol extracts (CT-26 colon carcinoma inhibition)16S rRNASilva-Stenico et al., 2014; Genuário et al., 2019Nodosilinea sp., Chlorogloea sp., Cyanobium sp., Halotia wernerae, Fischerella sp.Methanol extracts (cytotoxic to MOLM-13 leukemia cells)16S rRNASilva-Stenico et al., 2014; Shishido et al., 2019Aphanothece elabens, Lyngbya majuscula, Leptolyngbya tenuis, Oscillatoria accuminata, Oscillatoria tenuis, Calothrix breviarticulata, Aliinostoc bakau sp. nov.Microcystins, saxitoxinMorphology, Polyphasic taxonomy for A. bakauMohamed & Al-Shehri, 2015; Merican et al., 2023Moorena producensApratoxins, aeruginosin16S rRNAThornburg et al., 2013Oscillatoria sp. Gwada, Candidatus Planktothricoides niger, Candidatus Planktothricoides rosea, Cyanobium sp. CENA166, Synechococcus CENA170, Nostoc (Aliinostoc) sp. CENA175, Cyanobium sp. CENA153, CENA154, CENA185, Fischerella sp. CENA161Cyanotoxins16S rRNAGuidi-Rontani et al., 2014; Genuário et al., 2019; Silva-Stenico et al., 2012; Shishido et al., 2019Lyngbya cf. aestuarii, Microcoleus chthonoplastesScytonemin, MAAsMorphologyKarsten et al., 1998; Singh et al., 2020Oxynema aestuariiPhycocyaninPolyphasic taxonomyHazra & Saha Kesh, 2017;Acaryochloris sp.Chlorophyll-d16S rRNALarkum et al., 2012Spirulina subsalsaCarbohydrates, proteins, lipids, vitamins, mineralsMorphologicalSakthivel & Kathiresan, 2013Synechocystis sp.Fatty acids (linoleic acid, α-linolenic acid)Matsudo et al., 2020Phormidium sp., Microcoleus chthonoplastes, Phormidium, Microcoleus, OscillatoriaIndole acetic acid, EPS, secondary metabolite mixture16S rRNA; morphologicalBoopathi et al., 2013; Sakthivel & Kathiresan, 2015; Bashan et al., 2000Synechocystis sp.Polyhydroxyalkanoates16S rRNAGracioso et al., 2021Fig. 4. Oval flower diagrams of mangrove cyanobacterial genera with antimicrobial, anti-tumor, biofuel, and functional food activities. The overlaps denote cyanobacteria that have multiple bioactivities.Fig 4: dummy alt text
Antimicrobial compounds from cyanobacteria
4.1
Cyanobacteria have been shown to produce potent antimicrobial compounds, which are gradually gaining importance as many traditional antibiotics fail due to rising resistance (O’Neill, 2016). The majority of the cyanobacterial NPs showing bioactivity are products of polyketide synthase (PKS), nonribosomal peptide synthetase (NRPS), hybrid (NRPS‐PKS), or post-translationally modified ribosomes (RiPPs) (Alsenani et al., 2020). Unsaturated fatty acids and terpenoids of microalgae and cyanobacteria have chiefly been found to possess potent antimicrobial properties (Carpine and Sieber, 2021; Welker et al., 2012, (Kini et al., 2020). A recent study found that Planktothrix mougeotii PMC 1114.19, a filamentous cyanobacterial strain isolated from mangrove biofilms in Mayotte island, produces the anti-helmintic and anti-biofilm triterpenoid godavarin K (Wang et al., 2023). Organic solvent extracts of various mangrove-dwelling cyanobacteria isolated from Indian and Brazilian mangrove environments (further elaborated below) have been demonstrated to show antimicrobial activity and can therefore be considered as potential candidates for drug development (Fig. 5). Research indicates that methanol extracts from newly described cyanobacteria, including Oxynema aestuarii, Euryhalinema mangrovii, Euryhalinema pallustris, Aerofilum fasciculatum, and Leptoelongatus littoralis, effectively inhibit the growth of Staphylococcus aureus, Escherichia coli, Bacillus subtilis, Pseudomonas aeruginosa, and multiple drug-resistant bacterial isolates (Pramanik et al., 2011; Chakraborty et al., 2018, 2019, 2021). Based on the modern taxonomic identification using a polyphasic approach, which includes 16S rRNA phylogeny and 16S-23S ITS secondary structure analysis, O. aestuarii belongs to Microcoleaceae, E. mangrovii and L. litoralis belong to Leptolyngbyaceae, and A. fasciculatum belongs to Oculatellaceae and E. pallustris to Prochlorotrichaceae families, respectively (Chakraborty et al., 2018, 2019, 2021). Research has demonstrated that methanol and methanol:chloroform: water extracts from cyanobacteria, including Synechocystis salina, Oscillatoria salina, Spirulina subsalsa, Phormidium fragile, Oscillatoria cortiana, O. willei, and Phormidium tenue, inhibit various human pathogens and fungi such as Fusarium solani and Aspergillus flavus (Sakthivel and Kathiresan, 2015). All the above-mentioned cyanobacteria were classified using only on morphological features following Desikachary (1959) (Desikachary, 1959). Aqueous extracts of Synechococcus sp. and Gloeocapsa sp. were shown to hinder the development of Proteus vulgaris and Pseudomonas aeruginosa (Anburaj et al., 2020b). These cyanobacterial strains were identified based on 16s rDNA identification, but phylogeny and NCBI accession numbers were undisclosed. Methanol:acetonitrile: water extracts of members of the Oscillatoria and Jaaginema genera have been shown to inhibit pathogenic E. coli and environmental P. atlantica (Duperron et al., 2019). In this literature, the authors studied ten strains of the Oscillatoriales family and eight strains of the Synechococcales family, including Jaaginema sp. Although these strains were identified based on only 16s rRNA sequence analysis, it is essential to review and update the taxonomic positions of these cyanobacteria of potential value using the polyphasic approach, to determine the cultivable strains and choices for strain improvement (Bagchi and Singh, 2019; Baunach et al., 2024).Fig. 5. Diagram of mangrove cyanobacterial genera producing pigments, biofertilizers, photoprotective compounds, and toxins, including bioremediating species.Fig 5: dummy alt text
Extracellular ethyl acetate and chloroform extracts and intracellular methanolic extracts of several members of the genera Cyanobium, Chlorogloea, Cyanobacterium, Oxynema, Synechococcus, Leptolyngbya, Nodosilinea, and Nostoc were proven to be effective inhibitors of several pathogenic bacteria and yeast such as Bacillus cereus, Salmonella typhimurium, and Candida sp*.* (Silva et al., 2014; Silva-Stenico et al., 2013).
Methanolic intracellular extracts of various isolates of Fischerella sp. and Aliinostoc sp. were also reported to inhibit Staphylococcus sp. and Candida albicans (Shishido et al., 2019). Fischerella is reported under the Haplosiphonaceae family according to modern cyanobacteria taxonomic classification (Komárek, 2014). The taxonomic assignment of all the strains was based solely upon a 16S rRNA phylogenetic tree study, which is appropriate for genus-level identification. Precise species-level identification necessitates examining the secondary structures of the 16S-23S ITS regions, which are lacking for these strains.
Antitumor and anticancer activity
4.2
Owing to their unique biological profiles, several marine cyanobacterial NPs, such as curacin A, dolastatins 10 and 15, and their chemically synthesized analogs, are currently undergoing studies for further drug development (Tan, 2007). These compounds and their analogs which display anticancer activity can cause the potential destruction of various cancer cells through inducing apoptosis, or alteration of cell signaling activity (primarily involving the proteins belonging to the kinase- C family), and various other modes of cancer cell impairment viz. arrest of the cancer cell cycle, induction of dysfunctions of cancer cell mitochondria and oxidative damage to cancer cells (Qamar et al., 2021). Investigations of metabolites extracted from mangrove cyanobacteria isolated from the Indian Sundarbans and various mangrove forests in Brazil (described below) revealed potent anti-inflammatory and anti-cancer activity. Recent findings, indicate that ethyl acetate extracts of the mangrove-inhabiting cyanobacterium Oxynema sp. AP17 exhibits noteworthy anti-angiogenic activity (Basu et al., 2021). Research shows that ethanol extracts from Cyanobium sp. and Oxynema sp. effectively suppress the growth of CT-26 colon carcinoma cells (Silva-Stenico et al., 2014; Genuário et al., 2019). Methanol extracts of Nodosilinea sp., Chlorogloea sp., Cyanobium sp., Halotia wernerae, and Fischerella sp. were reported to be cytotoxic to the human leukaemia MOLM-13 cell line (Silva-Stenico et al., 2014; Shishido et al., 2019). However, though Oxynema sp. AP17 was identified by the polyphasic approach (Chakraborty et al., 2018); the rest of the strains mentioned in this section were identified on the sole basis of 16S rRNA sequence.
Toxins
4.3
Mangrove-dwelling cyanobacterial species such as Aphanothece elabens, Lyngbya majuscula, Leptolyngbya tenuis, Oscillatoria accuminata, Oscillatoria tenuis, Calothrix breviarticulata (Mohamed and Al-Shehri, 2015), Moorena producens (Thornburg et al., 2013), Oscillatoria sp. Gwada, Candidatus Planktothricoides niger, Candidatus Planktothricoides rosea (Guidi-Rontani et al., 2014), Cyanobium sp. CENA166, Synechococcus CENA170, and Nostoc (Aliinostoc) sp. CENA175 (Genuário et al., 2019; Silva-Stenico et al., 2012), Cyanobium sp. CENA153, Cyanobium sp. CENA154, Fischerella sp. CENA161, Cyanobium sp. CENA185 (Shishido et al., 2019) have been known to produce toxins that are lethal to most commercially important aquatic finfish and shellfish (and subsequently to the population consuming them). Toxins are usually produced by mat-like massive cyanobacterial growths known as harmful blooms (or cyano-habs), such as those formed by Oscillatoria sp. Gwada, Candidatus Planktothricoides niger, Candidatus Planktothricoides rosea (Guidi-Rontani et al., 2014). 16S rRNA analysis shows that these organisms belong to the Oscillatoriales family of cyanobacteria. Anthropogenic activity-mediated degradation of the mangroves is believed to be the chief cause of the uncontrolled growth of cyano-hab-forming cyanobacteria (Singh and Babele, 2020). The discharge of industrial effluents containing massive volumes of phosphorus and nitrogen into riverine and estuarine waters and gradually escalating temperature and CO_2_ levels are some of the factors contributing to the worldwide increase of cyano-hb occurrences (Singh and Babele, 2020; Paerl and Otten, 2013; Sandrini et al., 2016). The NPs produced by cyano-habs (cyanotoxins) are known to possess distinct structures and are highly toxic to animals and plants even at trace levels (Pearson et al., 2010).. Phylogenetic analysis using 16S rRNA shows the strain belongs to Moorena producens, which was formerly misspelled as Moorea sp. (Thornburg et al., 2013). Moorena producens is reported under the Oscillatoriacae family of cyanobacteria. Microcystin is a hepatotoxin that is produced by Leptolyngbya tenuis, Oscillatoria tenuis, Aphanothece elabens, Cyanobium, Fischerella, and Calothrix breviarticulata. Alkaloids (such as saxitoxin produced by Lyngbya majuscula, Oscillatoria accuminata, and Leptolyngbya tenuis) act as neurotoxins (Shishido et al., 2019; Mohamed and Al-Shehri, 2015). Merican and colleagues (2023) reported a novel species, Aliinostoc bakau sp. nov., as a producer of microcystins. This cyanobacterium was isolated from a mangrove-associated biofilm in Malaysia and subsequently classified using polyphasic taxonomic methods. Some of these above-described toxins, notably apratoxins, and aeruginosin have potential anti-cancer, and trypsin and thrombin inhibiting activity respectively (Thornburg et al., 2013; Silva-Stenico et al., 2012). The toxin-secreting capability of cyano-habs is a cause of concern to the overall health of the mangrove ecosystems due to the risk of biomagnification of the toxins in higher trophic levels of the food web initiated by primary consumers that feed on toxic cyanobacteria. Thus arises the need to monitor mangrove regions for detecting the presence of cyano- habs and their secreted cyanotoxins in a bid to prevent their undesirable effects (Singh and Babele, 2020). Thorough polyphasic and systematic identification of toxin-producing cyanobacteria, incorporating both molecular markers and morphological traits, is vital for fully assessing the risks they pose to animal and human health when present in drinking water (Wilmotte et al., 2017). This is because different members of the same or different species, as well as various cyanobacterial populations, can be either toxic or nontoxic, with toxic members having variable levels of toxins per cell.
Photoprotective compounds
4.4
Cyanobacteria, especially those dwelling in the mangrove habitats (notably Lyngbya cf. aestuarii and Microcoleus chthonoplastes), are exposed daily to intense solar radiation by their latitudinal location in the tropical and subtropical coastal intertidal regions (Karsten et al., 1998; Singh et al., 2020). Lyngbya is a member of the Oscillatoriaceae family capable of producing photoprotective compounds. Like other photosynthetic organisms, mangrove cyanobacteria synthesize UV protective molecules such as scytonemin to overcome UV exposure-induced cellular damage (caused chiefly by shortwave UV- B radiations of wavelength ranging from 280 to 350 nm) which may lead to altered cell physiology and biochemistry, and extensive cellular DNA/ RNA and protein damage (Richa et al., 2016). The water-soluble transparent mycosporine-like amino acids (MAAs) are the most widespread intracellular UV-absorptive NPs synthesized by cyanobacterial species, exhibiting molecular weight ranging from 188 to 1050 Da (Singh et al., 2020; Pathak et al., 2019), while the hydrophobic scytonemins are less commonly found among cyanobacteria (Pathak et al., 2019). These UV-absorbing compounds harmlessly convert harmful solar energy into heat, avoiding the production of reactive oxygen species and thereby preventing oxidative processes linked to tumor formation (Bhatia et al., 2010). They also reduce the formation of UV-B-induced thymine dimer formation in hairless albino mice (Singh et al., 2020). Antioxidant properties (Rastogi et al., 2016) and antiproliferative effects on neoplastic cells (Singh et al., 2020) are some other characteristics exhibited by MAAs apart from UV screening activity. Scytonemin is a (naturally) dark brown UV-absorbing compound that appears in the outer mucilaginous sheath of cyanobacterial cells, particularly those bare to prolonged sunlight irradiation (Garcia-Pichel and Castenholz, 1991). Scytonemin not only provides UV protection but also delivers anti-inflammatory and anti-proliferative effects by inhibiting a serine/threonine kinase, specifically polo-like kinase-1, which is essential for mitotic spindle formation in fibroblasts (Stevenson et al., 2002). Detailed reports on photoprotective compounds derived from mangrove cyanobacteria are limited to the two aforementioned strains. As random screening is usually employed (methods in the quest for organisms producing bioactive compounds) is often incapable of removing recurring species in the initial stages (Bull et al., 1992), the polyphasic method of identification would probably help find new organisms producing such compounds.
Dyes and coloring agents
4.5
Biopigments and coloring agents derived from cyanobacteria, such as chlorophylls, carotenoids, and phycobiliproteins, have appeared as an alternative source of biodegradable colorants that are favorable for human health and environmentally sustainable, as opposed to chemically manufactured dyes, which are toxic to humans as well as the ecosystem (Saini et al., 2018; Mandal et al., 2020). Cyanobacterial chlorophylls (a, b, d, and f), on account of selectively absorbing light of the red and blue colors in the visible light spectrum, produce green and yellow pigments that are used as food color (Saini et al., 2018). They have also proved capable of functioning as a biomordant to increase the dyes of textiles, in addition to an antimicrobial colorant for fabrics (Miazek et al., 2015). Phycobiliproteins extracted from cyanobacteria are classified into different groups, such as allophycocyanin, allophycocyanin B, phycoerythrocyanin, blue C-phycocyanin, phycoerythrin, and phycoerythrin. All are water-soluble proteins that absorb red, orange, yellow, and green lights of the visible spectrum, respectively (Saini et al., 2018; Mandal et al., 2020). A phycocyanin extracted from Oxynema aestuarii obtained from the Indian Sundarbans was recently reported to show antioxidant activity (Hazra and Saha Kesh, 2017). This organism was identified using the polyphasic taxonomy approach. A mangrove-dwelling member of the genus Acaryochloris was also reported to contain chlorophyll-d as its major photosynthetic pigment (Larkum et al., 2012). Identification of this organism, however, is based solely on its 16S rRNA sequence.
Functional food
4.6
Driven by the urgent need for sustainable and scalable food sources in the wake of global population growth, there is increasing interest in exploring non-traditional alternatives beyond conventional crops. Growing concerns about feeding an expanding global population have intensified interest in functional foods as a promising way to meet nutritional needs sustainably, sparking increased research and consumer demand. This has promoted interest in the use of functional foods to meet the nutritional demands of the growing human population (Dixit and Suseela, 2013; Panjiar et al., 2017). Cyanobacteria (such as Spirulina subsalsa) and microalgae have long been consumed as functional foods due to their dense composition of essential nutrients, including polysaccharides, complete proteins, essential fatty acids, vitamins, and minerals supporting their role in human nutrition (Milledge, 2011). High levels of a multitude of nutrients, ease of cultivation without stringent requirements for freshwater or fertile farmland, and the ability of cyanobacteria and their products to withstand temperature and pH fluctuations make cyanobacteria an attractive candidate for exploration as a possible non-conventional source of nutrients (Panjiar et al., 2017). Spirulina subsalsa, a dominant cyanobacterial species found in three mangrove forests across the Southeastern coast of India, is identified as a source of carbohydrates, proteins, and lipids along with several important minerals and vitamins (Sakthivel and Kathiresan, 2013). However, these cyanobacteria were solely identified based on the taxonomic keys provided by Desikachary (1959). Polyphasic taxonomic studies of functional food-producing candidate cyanobacteria would be beneficial for accurate strain identification and assessing the non-toxic nature of cyanobacteria used in nutritional supplements by combining morphology, molecular phylogeny, chemistry, and ecology for preventing misidentification and detection of toxic or adulterant strains (Komárek, 2016). Brazilian mangrove forests harbor a member of the genus Synechocystis, a known producer of significant amounts of fatty acids such as linoleic acid and alpha-linolenic acid that have gained importance as nutritional supplements (Matsudo et al., 2020). A member of the genus Cyanobium (occurring in Brazilian mangroves) is a source of adipic acid, which is utilized in the bakery industry as a leavening compound and an acidulant (Armstrong et al., 2019).
Biofuels
4.7
Fossil fuel usage has been proven to cause several large-scale global disturbances, namely a rise in the average global temperature, alterations in climate patterns, and adverse impacts on the environment caused by mining operations to recover coal and petroleum for use (Behera et al., 2015; Martens, 2013; Sitther et al., 2020). Thus, the quest for alternative, clean, and renewable sources of energy has prompted the exploration of other directly biologically derived options, such as fats derived from food crops, viz., soybean oil, sugarcane lipids, and corn oils, which are subsequently used as feedstocks for producing diesel alternatives and ethanol (Sitther et al., 2020; Goldemberg, 2007). The idea of cyanobacterial cells used as a potential biofuel source is increasingly gaining popularity, as they might prove to have shorter generation time than common food crops, they are capable of growing in minimal light and nutrient availability, and do not compete for agricultural fields with other valuable cash crops (Sitther et al., 2020; Rodolfi et al., 2009). The biodiesel extracted from cyanobacteria has also proved to be eco-friendly as it has significantly lower levels of sulfur and aromatic hydrocarbon emissions, and efficient combustion (Sitther et al., 2020). Various methods of biochemical processing can be used on the cyanobacterial biomass in order to manufacture a broad range of energy sources such as biogas, biohydrogen, cellulosic ethanol, and biodiesel (Machado and Atsumi, 2012). Cyanobacterial isolates from the sugars can readily ferment into bioethanol, while cellular lipids can be chemically transformed into fatty acid methyl esters (FAMEs), which are the primary constituents of biodiesel. The above-mentioned advantages coupled with the efficacy of capture and transformation of solar energy to fuels make cyanobacteria the candidates of choice for exploitation, offering significant financial and logistical benefits compared to the manufacture of biofuel from cash crops (Sitther et al., 2020; de Farias Silva and Bertucco, 2016). Mangrove-dwelling members of the genera Oxynema, Nostoc, Synechocystis, and Cyanobium have recently been reported to produce intermediates important in biofuel production (Matsudo et al., 2020; Armstrong et al., 2019). Cyanobacterial polyphasic taxonomy in combination with phylogenomics helped in the precise identification of conserved functional genes responsible for the production of polyunsaturated fatty acids (PUFAs) in the diatom Phaeodactylum tricornutum, and linked these genes to well‑characterized taxa (Rehmanji et al., 2022). This integrated approach is anticipated to facilitate the understanding of mangrove cyanobacterial metabolism and guide targeted enhancements to boost yields in cyanobacterial biorefineries (Zammit et al., 2023).
Bio-fertilizers
4.8
Sustainable agriculture is currently an urgent need as the earth strains to recover from the damage caused by claiming more land for agricultural purposes and enhancing the productivity of land already under cultivation using synthetic fertilizers and pesticides. Food security is essential for the purpose of accommodating and feeding the staggering global population of ∼7.9 billion people (Joshi et al., 2020). Bio-fertilizers are gaining popularity as sustainable, eco-friendly fertilizers (as opposed to chemical fertilizers) as they can produce a host of beneficial organic compounds in soil that stimulate the growth of seeds and plants and also maintain a proper balance of macro-nutrients nitrogen, phosphate, potassium (NPK), and other micronutrients (Joshi et al., 2020). Mangrove cyanobacteria have added advantages, such as secretion of plant hormone like Indole-3-acetic acid (IAA) from Phormidium sp. (identified by 16S rRNA analysis) (Boopathi et al., 2013), and other secondary metabolites (such as seed and foliage lipid content enhancer secreted by Microcoleus chthonoplastes isolated from a Mexican mangrove forest) (Bashan et al., 2000) that supplement healthy crop growth; fixing of atmospheric nitrogen in the soil (notably by Microcoleus chthonoplastes) to replenish the nitrogen content of agricultural fields (Bashan et al., 1998; Kathiresan, 2000); and acting as a biocontrol agent against phytopathogens by secreting antifungal and antibacterial NPs (Sakthivel and Kathiresan, 2015; Joshi et al., 2020). Cyanobacteria may also act as soil stabilizers by forming three-dimensional extracellular polysaccharide (EPS) filamentous sheaths (Lynch and Bragg, 1985). Cyanobacteria used in agriculture as cost-effective alternatives to harmful chemically synthesized fertilizers benefit the environment, as well as human and livestock health by recycling nutrients and restoring soil fertility through the benign method of nitrogen fixation, heading towards the improved production of nutritious and safe-to-consume food (Joshi et al., 2020; Bashan et al., 1998). Additional systematic and phylogenomic investigation of mangrove-dwelling members of Phormidium, Microcoleus, and Oscillatoria genera may prove profitable for agricultural applications, as well as for determining the roles of these cyanobacteria in the mangrove ecosystem (Joshi et al., 2020). It is also expected to be helpful to identify lineages with metabolic pathways relevant to IAA and EPS production and mapping these genes across clades of neighbouring cyanobacterial strains, thus broadening the candidate pool for biofertilizer production (Zammit et al., 2023).
Polyhydroxyalkanoate bioplastics
4.9
Plastics are used in most aspects of daily life, owing to their versatile mechanical properties, durability, thermal and electrical insulating properties, lightweight, and cheap production costs (Samantaray et al., 2014). However, wastes generated from plastic articles do not easily undergo natural microbe-mediated decomposition (i.e., biodegradation) (Barnes et al., 2009). Attempts to dispose of petrochemical-derived plastics by incineration lead to the production of toxic gases such as dioxins, polycyclic aromatic hydrocarbons (PAHs), furans, and hydrogen cyanide ((Samantaray et al., 2014; Barnes et al., 2009). The increasing awareness about the detrimental effects of the petroleum-based plastic wastes on the environment has led to the search for novel raw materials that can be utilized for the manufacture of plastics developed from biodegradable polymers (Lee, 1996, Samantaray et al., 2014). Polyhydroxyalkanoates (PHAs) are polyesters of hydroxyl acids produced and accumulated intracellularly by several prokaryotes (including cyanobacteria) and are completely degraded by environmental microbiota (Lee, 1996). The products made out of PHAs are suitable for numerous applications, including the production of medical implants, drug delivery carriers, printing, and photographic material, as well as more conventional practices in the production of packing and single-use materials (Samantaray et al., 2014; Wang et al., 2008; Yao et al., 2008). Members of Synechocystis isolated from a Brazilian mangrove environment have been shown to produce and accumulate high volumes of PHAs when subjected to high-intensity illumination (Gracioso et al., 2021). Further improvement of these strains for scaling up bioplastic production, field application, and realistic performance predictions requires meticulous exploration of their physiological and ecological parameters and tolerance levels, which is an integral part of polyphasic taxonomy (Komárek, 2016).
Environmental detoxification
4.10
Cyanobacteria are gaining popularity as an effective tool for the elimination of cytotoxic heavy metals coming from the groundwater, which are discharged mainly as a result of improper industrial effluent disposal containing huge volumes of toxic heavy metals eg, mangrove-derived Synechococcus elongatus and Gloeocapsa sp. effectively remove lead and cadmium, respectively (Genuário et al., 2019; Anburaj et al., 2020, 2017). All the strains were identified based on their external morphology. Heavy metals cause various disorders in both humans and wildlife, and several species of heavy metals are capable of causing a thinning of eggshells (Faber and Hickey, 1973). Cyanobacteria-mediated bioremediation involves highly efficient uptake of complex pollutants using sensitive polysaccharide receptors on their surface (Malik, 2004). The trapped pollutants are then bound passively to the cellular structure of the cyanobacterial cell aggregates through biosorption (Malik, 2004; Kulal et al., 2020). Mangrove-dwelling cyanobacteria are also garnering attention as potential degraders of some dyes, eg, a Brazilian species of the Phormidium genus has been reported to effectively decolorize indigo blue (Silva-Stenico et al., 2012). The current identification of these strains, however, was based on utilizing the 16S rRNA sequence only, highlighting a limited understanding of the taxonomical position of these cyanobacteria. Polyphasic taxonomy is anticipated to be helpful in correctly linking pollutant-degrading phenotypes to relevant cyanobacterial strains for reproducible bioremediation studies and further upscaling (Komárek, 2016; Zammit et al., 2023). A species of Phormidium derived from an Indian mangrove forest has also been reported to effectively degrade the textile dye malachite green (Kulal et al., 2020).
Conclusion: challenges and future scope
5
Cyanobacteria represent an ancient lineage of a morphologically varied group of photosynthetic, Gram-negative bacteria that have functioned as key players in the evolution of life forms on Earth through oxygenic photosynthesis, and they still play a role of paramount importance in the universal carbon and nitrogen cycles (Ducat et al., 2011). Approximately 50% of the ocean’s primary productivity is supplied by cyanobacteria, and they are potential alternative sources of various commercially and pharmaceutically important NPs (Pramanik et al., 2011; Jones et al., 2021). Despite having a ubiquitous presence, Cyanobacterial biotechnology and natural product (NP) discovery face technical and practical barriers that limit both NP discovery and commercial translation. Only a small subset of cyanobacteria is suitable for genetic engineering, limiting activation of BGCs and the scope of strain improvement, while many desirable metabolites appear at low, inconsistent titers that complicate purification and scale‑up (Covington et al., 2021; Romano et al., 2018). Heterologous expression is often solely experimental and resource‑intensive, requiring host redesign for compatible promoters, activating enzymes, and precursor supply, and bypassing cloning and expression challenges for large NRPS/PKS clusters (Liu and Pakrasi, 2018; Wells et al., 2018; Yang et al., 2017). Large-scale cultivation poses further obstacles: high water, nutrient, and energy inputs, contamination and phage risk, and ecological stability concerns in open systems, which undermine reliability (Iwaki et al., 2006; Qiang et al., 1998; Zahra et al., 2020). Miniaturized OSMAC screens, microfluidics, in vitro reconstitution and chemo‑enzymatic synthesis can circumvent some culture limitations and accelerate mechanistic insight, but these approaches may be costly, hard to scale and carry environmental trade‑offs from chemical steps (Baunach et al., 2024). Integrated strategies involving multiomics, enzyme engineering, and scalable, low‑impact workflows are therefore essential to bridge discovery with sustainable industrial use (Zahra et al., 2020).
Funding
This work was supported by the Council of Scientific and Industrial Research (CSIR), India, in the form of NET-Senior Research fellowship to ARR [09/096(0997)/2019-EMR-I] and the University Grants Commission (UGC), India, in the form of NET-Senior Research fellowship to SB [award number 200510616803].
CRediT authorship contribution statement
Arup Ratan Roy: Data curation, Methodology, Writing – original draft. Shayontani Basu: Data curation, Methodology, Writing – original draft. Sergio de los Santos Villalobos: Writing – review & editing. Joydeep Mukherjee: Supervision, Writing – review & editing.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abed R.M.M.Dobretsov S.Sudesh K.Applications of cyanobacteria in biotechnology J. Appl. Microbiol.106200911210.1111/j.1365-2672.2008.03918.x 19191979 · doi ↗ · pubmed ↗
- 2Adak S.Lukowski A.L.Schäfer R.J.B.Moore B.S.From tryptophan to toxin: nature’s convergent biosynthetic strategy to aetokthonotoxin J. Am. Chem. Soc.14420222861286610.1021/jacs.1c 1277835142504 PMC 9004672 · doi ↗ · pubmed ↗
- 3Alsenani F.Tupally K.R.Chua E.T.Eltanahy E.Alsufyani H.Parekh H.S.Schenk P.M.Evaluation of microalgae and cyanobacteria as potential sources of antimicrobial compounds Saudi Pharm. J.2820201834184110.1016/j.jsps.2020.11.01033424272 PMC 7783216 · doi ↗ · pubmed ↗
- 4Alvarenga D.O.Andreote A.P.D.Branco L.H.Z.Fiore M.F.Kryptousia macronema gen. nov., sp. nov. and Kryptousia microlepis sp. nov., nostocalean cyanobacteria isolated from phyllospheres Int. J. Syst. Evol. Microbiol.6720173301330910.1099/ijsem.0.00210928875896 · doi ↗ · pubmed ↗
- 5Alvarenga D.O.Rigonato J.Henrique Zanini Branco L.Soares Melo I.Fiore M.F.Phyllonema aviceniicola gen. nov., sp. nov. and Foliisarcina bertiogensis gen. nov., sp. nov., epiphyllic cyanobacteria associated with Avicennia schaueriana leaves Int. J. Syst. Evol. Microbiol.66201668970010.1099/ijsem.0.00077426582479 · doi ↗ · pubmed ↗
- 6Alvarenga D.O.Rigonato J.Branco L.H.Z.Fiore M.F.Cyanobacteria in mangrove ecosystems Biodivers. Conserv.24201579981710.1007/s 10531-015-0871-2 · doi ↗
- 7Anburaj R.Jebapriya G.R.Asmathunisha N.Kathiresan K.Antibacterial activity of mangrove-derived marine cyanobacteria Int. J. Sci. Res. Eng. Dev.32020542550
- 8Anburaj R.Jebapriya R.Kathiresan K.Biosorption of cadmium by mangrove-derived cyanobacteria (Gloeocapsa sp ARKK 3)Statistical Approaches on Multidisciplinary Research 20178294