Diversity of macrofungi in Dali University, Yunnan Province, China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
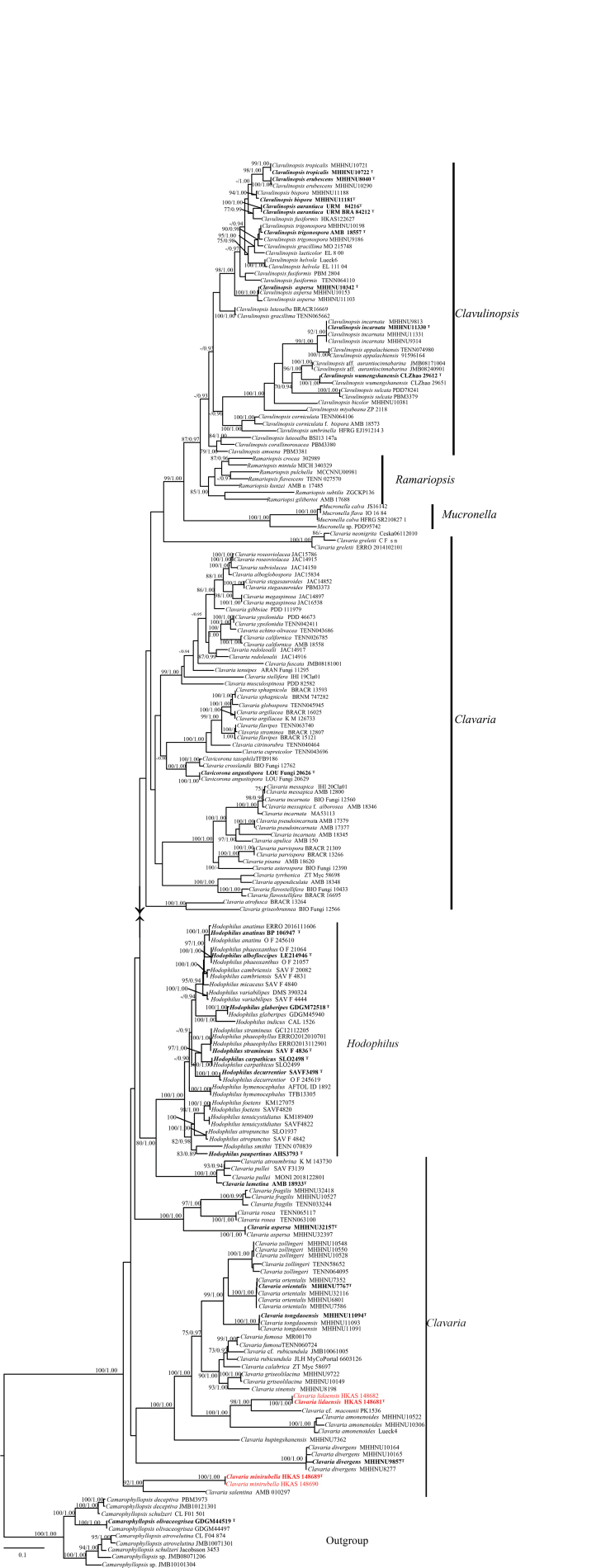 Figure 1
Figure 1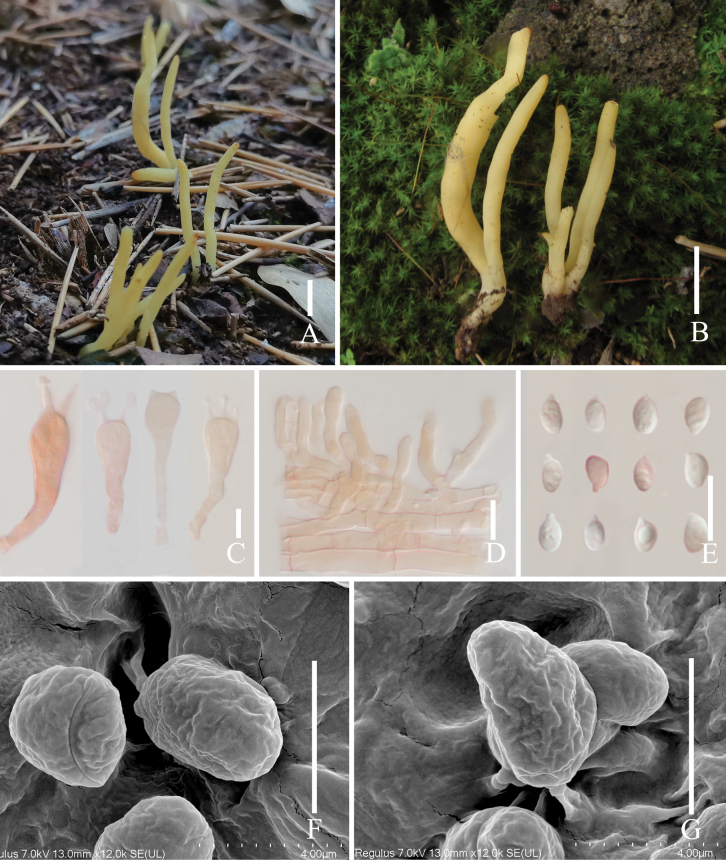 Figure 2
Figure 2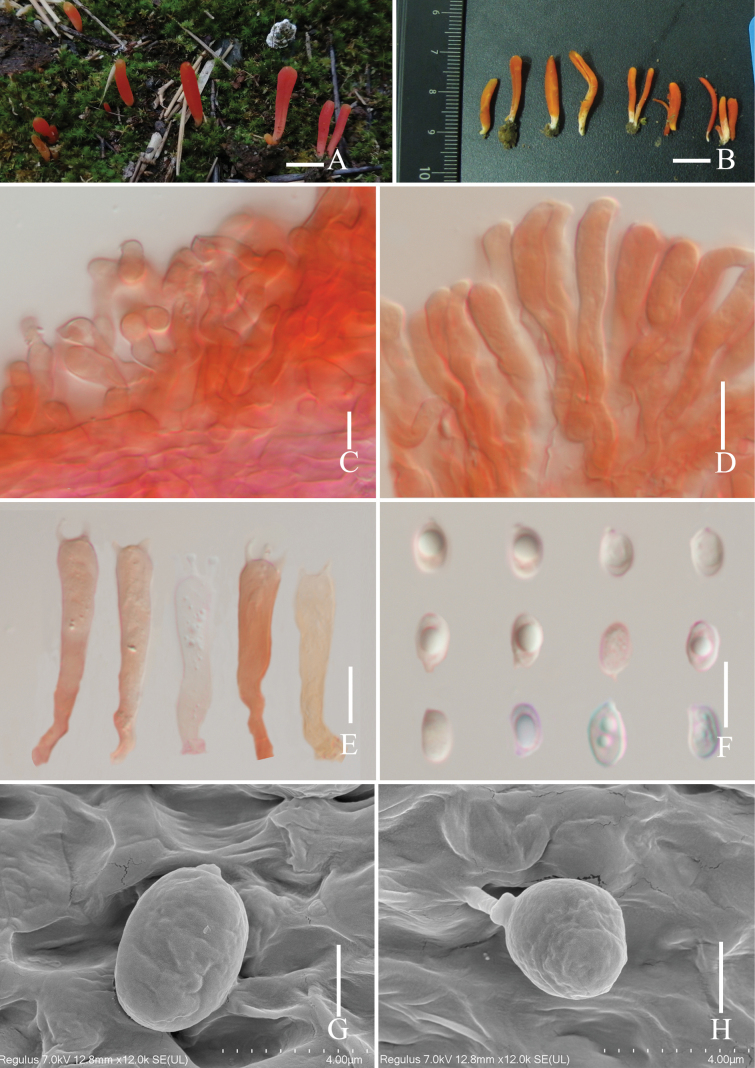 Figure 3
Figure 3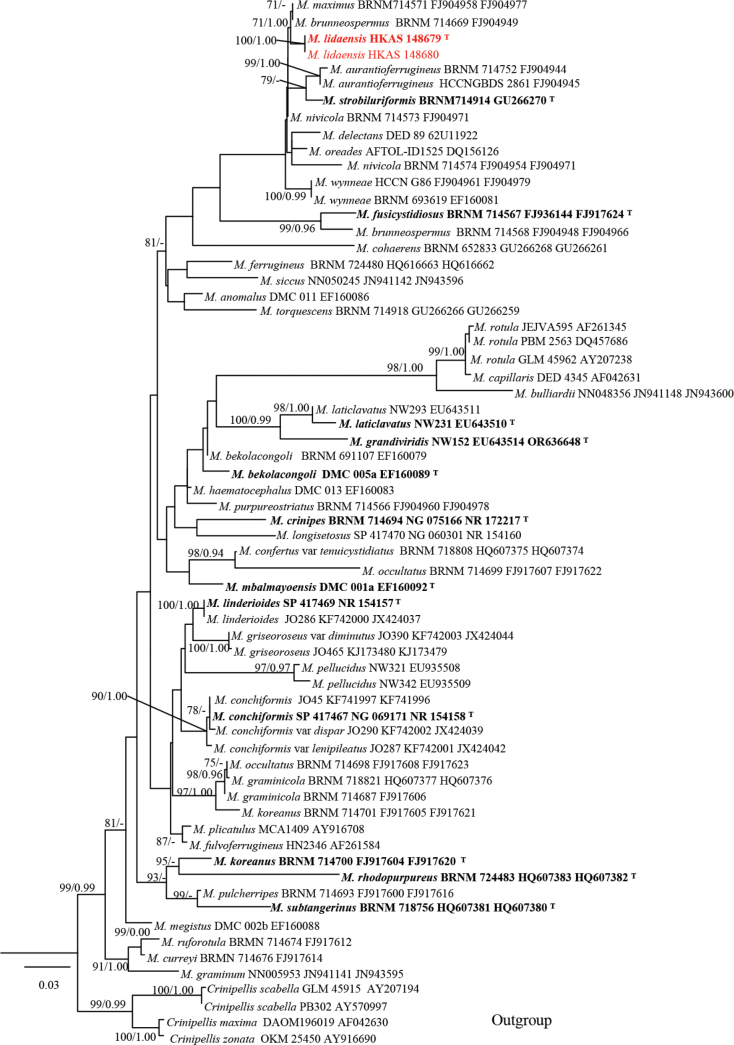 Figure 4
Figure 4 Figure 5
Figure 5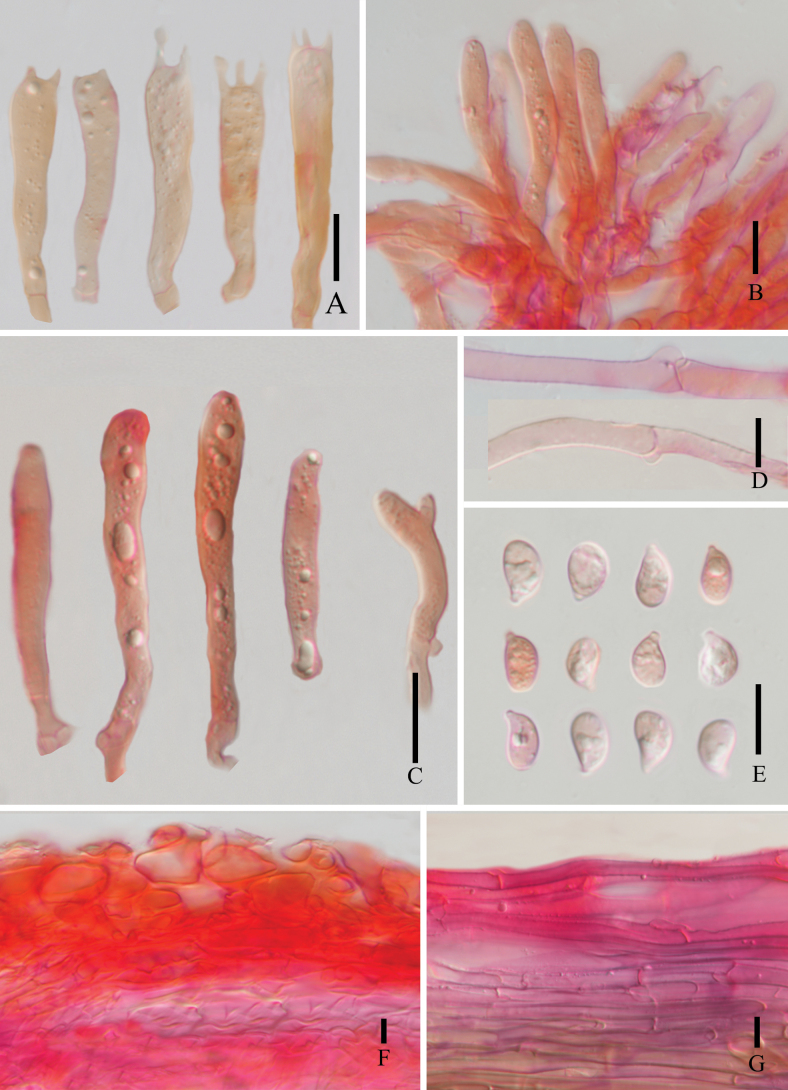 Figure 6
Figure 6 Figure 7
Figure 7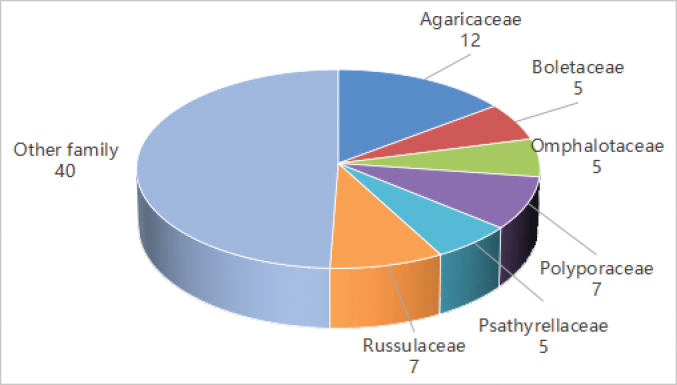 Figure 8
Figure 8| Taxon | Collection numbers | ITS | Family | Order |
|---|---|---|---|---|
|
| 20210721-02 |
|
|
|
|
| x20210718-03 |
|
|
|
|
| SJ1186 |
|
|
|
|
| x20210711-08 |
|
|
|
|
| x20210703-01 |
|
|
|
|
| x20210702-01 |
|
|
|
|
| x20210718-08 |
|
|
|
|
| Sj1743 |
|
|
|
|
| Sj1988 |
|
|
|
|
| Sj254 |
|
|
|
|
| 2020082517 |
| Incertae sedis |
|
|
| sj2386 |
|
|
|
|
| x20210727-06 |
|
|
|
|
| sj267 |
|
|
|
|
| sj268 |
|
|
|
|
| sj1759 |
|
|
|
|
| HKAS 148681 |
|
|
|
| HKAS 148682 |
| |||
|
| HKAS 148689 |
|
|
|
| HKAS 148690 |
| |||
|
| sj318 |
|
|
|
|
| x20210709-01 |
|
|
|
|
| x20210701-01 |
|
|
|
|
| x20210726-12 |
|
|
|
|
| sj600 |
|
|
|
|
| x20210818-04 |
|
|
|
|
| f20210902-01 |
|
|
|
|
| f20210902-10 |
|
|
|
|
| x20210710-03 |
|
|
|
|
| x20210727-07 |
|
|
|
|
| x20210708-03 |
|
|
|
|
| x20210711-01 |
|
|
|
|
| f20210901-23 |
|
|
|
|
| f20210901-28 |
|
|
|
|
| x20210727-12 |
|
|
|
|
| x20210707-06 |
|
|
|
|
| x20210708-06 |
|
|
|
|
| Sj1758 |
|
|
|
|
| Sj277 |
|
|
|
|
| sj356 |
|
|
|
|
| 20240723-6 |
|
|
|
|
| x20210717-01 |
|
|
|
|
| sj647 |
|
|
|
|
| x20210707-10 |
|
|
|
|
| x20210831-02 |
|
|
|
|
| f20210902 |
|
|
|
|
| x20210701-03 |
|
|
|
|
| Sj2384 |
|
|
|
|
| x20210726-08 |
|
|
|
|
| x20210903-05 |
|
|
|
|
| x20210714-02 |
|
|
|
|
| Sj630 |
|
|
|
|
| f20210902-02 |
|
|
|
|
| x20211024-05 |
|
|
|
|
| sj263 |
|
|
|
|
| SJ1170 |
|
|
|
|
| HKAS 148679 |
|
|
|
| HKAS 148680 |
| |||
|
| x20210720-03 |
|
|
|
|
| SJ1768 |
|
|
|
|
| Sj1737 |
|
|
|
|
| Sj628 |
|
|
|
|
| SJ1990 |
|
|
|
|
| x20210726-21 |
|
|
|
|
| x20210909-04 |
|
|
|
|
| ZS017 |
|
|
|
|
| f20210902-05 |
|
|
|
|
| x20210718-11 |
|
|
|
|
| x20210718-10 |
|
|
|
|
| SJ2387 |
|
|
|
|
| x20210726-17 |
|
|
|
|
| Sj268 |
|
|
|
|
| Sj1144 |
|
|
|
|
| Sj339 |
|
|
|
|
| f20210902-08 |
|
|
|
|
| f20210901-12 |
|
|
|
|
| x20210920-01 |
|
|
|
|
| Sj341 |
|
|
|
|
| Sj335 |
|
|
|
|
| x20210708-02 |
|
|
|
|
| f20210901-30 |
|
|
|
|
| Sj1201 |
|
|
|
|
| x20210717-06 |
|
|
|
|
| f20210902-11 |
|
|
|
|
| x20210901-05 |
|
|
|
|
| x20210714-05 |
|
|
|
| Scientific Name | Specimens | Sequence | Voucher | Max score | Total score | Query cover | Per. Ident. | Acc. length | Accession | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
|
| HKAS 148681 | ITS | PK1536 | 521 | 521 | 100% | 83.03% | 658 |
|
|
| P3J09 | 516 | 516 | 99% | 82.95% | 950 |
| unpublished | |||
| 3163 | 381 | 381 | 65% | 84.95% | 397 |
| unpublished | |||
| 175487368 | 364 | 364 | 74% | 81.82% | 658 |
| unpublished | |||
| Lueck4 | 350 | 350 | 57% | 85.39% | 600 |
|
| |||
| LSU | AMB018217 | 1317 | 1317 | 99% | 94.24% | 865 |
| unpublished | ||
| PK1536 | 1291 | 1291 | 100% | 93.52% | 1375 |
|
| |||
| 1258 | 1258 | 100% | 92.79% | 1341 |
|
| ||||
| Lueck4 | 1258 | 1258 | 100% | 92.79% | 1026 |
|
| |||
| AFTOL984 | 1243 | 1243 | 100% | 92.45% | 1402 |
|
| |||
|
| HKAS 148689 | ITS | 6603126 | 278 | 278 | 41% | 86.09% | 734 |
|
|
| MTS4577 | 291 | 291 | 46% | 85.19% | 635 |
| unpublished | |||
| BMS24-11 | 291 | 291 | 46% | 85.19% | 622 |
| unpublished | |||
| CF 32637 | 291 | 291 | 46% | 85.19% | 656 |
|
| |||
| 291 | 291 | 46% | 85.19% | 614 |
|
| ||||
| LSU | F-684 | 1240 | 1240 | 100% | 92.20% | 1497 |
| unpublished | ||
| AB0532 | 1238 | 1238 | 100% | 92.31% | 1324 |
|
| |||
| GG AB05-32 | 1238 | 1238 | 100% | 92.31% | 1048 |
|
| |||
| BRACR16666 | 1232 | 1232 | 100% | 92.19% | 943 |
|
| |||
| CF32637 | 1229 | 1229 | 100% | 92.08% | 1326 |
|
| |||
|
| HKAS 148679 | ITS | B6 | 1140 | 1140 | 100% | 98.32% | 678 |
| unpublished |
| NS16081406 | 1120 | 1120 | 100% | 97.57% | 688 |
| unpublished | |||
| BRNM714571 | 1105 | 1105 | 100% | 97.26% | 877 |
|
| |||
| KG224 | 1105 | 1105 | 100% | 97.26% | 850 |
|
| |||
| BRNM714570 | 1098 | 1098 | 100% | 97.10% | 840 |
|
| |||
| LSU | BRNM714571 | 1633 | 1633 | 100% | 99.78% | 1381 |
|
| ||
| BRNM714569 | 1628 | 1628 | 100% | 99.66% | 1410 |
|
| |||
| BRNM714570 | 1628 | 1628 | 100% | 99.66% | 1420 |
|
| |||
| G1352 | 1609 | 1609 | 99% | 99.55% | 1282 |
| unpublished | |||
| GAL 1672 | 1598 | 1598 | 100% | 99.10% | 890 |
| unpublished |
| Taxa | Basidiomata branched or unbranched | Basidiospores size | Basidia size | Basidiomata color | Reference |
|---|---|---|---|---|---|
|
| Unbranched | 7.0–8.5 × 5.0–7.0 μm | 42–55 × 7–10 μm | Pale orange to yellow |
|
|
| Unbranched | 4.0–6.0 × 2.5–4.0 μm | 35–50 × 5–8 μm | Yellow to very pale orange-yellow |
|
|
| Mostly Unbranched, rarely branched | 9.0–11.0 × 4.0–5.0 μm | 51–75 × 6–8 μm | Bright yellow to pale yellow |
|
|
| Unbranched | 4.0–5.0 × 2.5–4.0 μm | 35–50 × 4–8 μm | White |
|
|
| Unbranched | 5.0–6.0 × 3.8–4.2 μm | 27–40 × 4–6 μm | White |
|
|
| Branched | 4.2–5.0 × 2.7–3.8 μm | 48–65 × 6–9 μm | White, becoming yellowish or tawny with age |
|
|
| Unbranched | 7.5–9.0 × 5.5–7.0 μm | 33–40 × 5–9 μm | White when young, becoming white yellow with age |
|
|
| Unbranched | 6.0–8.5 × 4.0–6.0 μm | 41–60 × 7–8 μm | Pale yellow to light orange |
|
|
| Unbranched | 4.0–6.0 × 3.0–5.0 μm | 26–40 × 5–8 μm | White when young, becoming white yellow or yellow with age |
|
|
| Unbranched | 4.4–5.5 × 3.4–5.0 μm | 32–45 × 5–8 μm | Pale yellowish-brown, light gray, or smoky gray. |
|
|
| Branched | 4.0–7.0 × 3.0–5.0 μm | 42–57 × 8–10 μm | Pale purple to grayish purple |
|
|
| Unbranched | 7.0–10.0 × 6.0–8.0 µm | 35–50 × 6–9 µm | Whitish when young, becoming cream, yellowish brown |
|
|
| Branched | 4.0–6.0 × 3.5–5.0 μm | 36–50 × 5–8 μm | Rose-white to seashell-pink |
|
|
|
|
|
|
|
|
|
| Unbranched | 6.0–7.0 × 3.0–4.0 µm | 30–60 × 5–6 µm | Cream, pale orange-yellow, barium yellow |
|
|
|
|
|
|
|
|
|
| Branched | 5.0–6.0 × 4.0–5.0 μm | 34–48 × 5–8 μm | Deep amethyst, becoming yellowish or tawny with age |
|
|
| Unbranched | 6.0–7.0 × 3.5–4.2 μm | 32–45 × 6–9 μm | Wine-colored, light yellow, or dark pink |
|
|
| Unbranched | 6.0–8.0 × 3.0–4.0 µm | 40–45 × 8 µm | Rose pink, with a white or pallid stem |
|
|
| Branched | 5.0–6.0 × 3.5–4.5 μm | 30–40 × 6–8 μm | Pale purple to pinkish-purple |
|
|
| Branched | 3.5–5.0 × 3.0–4.2 μm | 26–43 × 6–8 μm | Pale purple to pale purplish pink, becoming yellowish or tawny with age. |
|
|
| Branched | 5.0–6.0 × 4.0–5.0 μm | 34–48 × 5–8 μm | Violet, lilac, or purplish-brown |
|
| Taxa | Pileus size | Pileus color | Basidiospores size | Basidia size | Reference |
|---|---|---|---|---|---|
|
| 30–70 mm wide | Orange | 11.5–15 × 4.5–6.0 μm | 26–28 × 8–11 μm |
|
|
| 20–50 mm wide | Yellowish white to light brown | 6.0–8.0 × 3.0–4.0 μm | 28–32 × 6–7 μm |
|
|
| 10–20 mm wide | Whitish darkened | 7–9 × 4 μm | – |
|
|
|
|
|
|
|
|
|
| 25–65 mm wide | Ochre-brown at center, otherwise pale brownish to pale ochraceous | 7.0–10.3 × 4.5–6.0 μm | 35–48 × 7–10 μm |
|
|
| 5–40 mm wide | White-off, whitish to yellowish white or pale yellow | 6.5–8.0 × 3.7–5.0 μm | 24–29 × 6–9 μm |
|
|
| 30–55 mm wide | Pale brownish to pale ochraceous | 7.5–9.5 × 3.6–5.0 µm | 35–48 × 7–10 μm |
|
|
| 10–50 mm broad | White or gray ochraceous when young, then milky white, gray, or gray violaceous | 6.5–8.0 × 3.7–4.5 μm | 24 × 7 μm |
|
- —Yunnan Provincial Department of Education 501100007846 https://ror.org/02yrxdp92 http://doi.org/10.13039/501100007846
- —Dali University 501100010617 http://doi.org/10.13039/501100010617
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Biocrusts and Microbial Ecology · Slime Mold and Myxomycetes Research
Introduction
Macrofungi constitute a highly significant category of fungi due to their diverse applications in food, medicine, and ecology (Wu et al. 2019; Corrales et al. 2022; Dong et al. 2024, 2025; Niu et al. 2025; Song et al. 2025; Tang et al. 2025a; Wang et al. 2025; Xu et al. 2025; Yang et al. 2025; Zhang et al. 2025). These fungi are highly valued for their edible properties, providing a rich source of nutrients and flavor. In medicine, many macrofungi contain bioactive compounds with potential therapeutic applications, including antioxidants, antimicrobial agents, and immune modulators. Ecologically, they play vital roles in nutrient cycling and soil health. However, it is also important to note that certain macrofungi can act as forest pathogens or produce toxic compounds, posing potential risks to both humans and ecosystems (Sun et al. 2024).
By 2007, global estimates suggested that 53,000–110,000 species of macrofungi existed worldwide (Mueller et al. 2007; Arenas et al. 2018). In China, a comprehensive survey reported 9,302 macrofungi species, including 1,789 edible fungi, 798 medicinal fungi, and 561 species serving both purposes (Dai et al. 2021). More recently, a study documented 558 macrofungal species in southeast Xizang province (Cui et al. 2025); this indicates that southwestern China is rich in fungal resources. Additionally, research has been actively conducted on the species diversity of macrofungi across different regions of China. These studies highlight the ecological, economic, and cultural significance of macrofungi, which play crucial roles in nutrient cycling, soil formation, and plant health through mycorrhizal associations. They also underscore the need for conservation efforts to protect these valuable resources from threats such as habitat loss and overharvesting.
According to the Catalogue of Life China 2025 Annual Checklist, a total of 27,996 fungal species have been discovered in China. In 2025 alone, 1,247 new species and infraspecific taxa were published, including 856 species in the Ascomycota and 20 species in the Basidiomycota.
In China, numerous universities have conducted large-scale surveys on fungal species diversity, with Jiangxi Agricultural University reporting 169 species (Zhang et al. 2018), Hanjiang Normal University 96 species (Wang et al. 2017), Hainan University 88 species (Deng and Wu 2014), Jiangxi Meilin University 77 species (Chai et al. 2014), and Northwest A&F University 73 species (Dou et al. 2015). However, reports on macrofungi diversity from universities in Yunnan Province remain relatively scarce (Lu et al. 2023, 2024).
Dali University, located in Yunnan Province, southwestern China, is a beautiful institution with rich vegetation types and abundant macrofungal resources. Although ecologically important, the diversity of macrofungi in this university campus has been poorly characterized in previous studies. From 2020 to 2024, a comprehensive investigation of macrofungi on campus was conducted to fill this knowledge gap.
Through morphological examination and molecular analyses, we identified 83 macrofungal species, including three new species, they are Clavaria lidaensis, C. minirubella and Marasmius lidaensis.
The Clavaria Vaill. ex L. is a member of Clavariaceae Chevallier. Species of this genus are characterized by their simple clavarioid or branched basidiomata (Corner 1950, 1970; Petersen 1988; Yan et al. 2022). At present, approximately 40 species are recognized worldwide, and 13 species have been recorded in China (Tai 1979; Olariaga et al. 2015; Chen and Zhang 2019; Yan et al. 2020, 2022, 2023, 2025).
Marasmius Fr. was established by Fries (1836); it is characterized by the small to medium-sized basidiomata, white spore prints, inamyloid basidiospores, and hymeniform pileipellis (Retnowati and Desjardin 2022). Recent molecular phylogenetic analyses have demonstrated the polyphyletic nature of Marasmius (Wilson and Desjardin 2005), with 17 species of this genus having been originally described from China (Zhang et al. 2023; Chen et al. 2025).
This study not only enriches the understanding of macrofungal diversity in the region but also highlights the importance of further exploration and conservation efforts. A checklist of the identified macrofungi species is provided, along with GenBank numbers for the sequences obtained from the studied samples. Detailed illustrations, descriptions, and phylogenetic analysis results of the three new species are also included. The findings emphasize the need for continued research on macrofungal diversity in similar environments across China and beyond. Such studies are crucial for understanding the ecological roles of macrofungi, their potential economic value, and the threats they face from habitat loss and climate change. Furthermore, documenting macrofungal diversity contributes to the broader understanding of ecosystem complexity and resilience, while providing opportunities for public education and science outreach. By engaging local communities and students in fungal surveys and conservation efforts, researchers can raise awareness of the importance of fungi in sustaining healthy ecosystems and the need for biodiversity conservation.
Materials and methods
Morphological studies
Specimens were photographed in the field; all the important collections data were recorded (Rathnayaka et al. 2025), and specimens were separately wrapped in aluminium foil or kept in a plastic collection box and taken to the laboratory of the Fungal Diversity Conservation and Utilization Team (FDCU) in Northwest Yunnan (Dali University). The fresh basidiomes were macro-morphologically described on the same day of collection. Color identification was performed using the Color Hexa website (www. colorhexa. com) to assign codes (Tang et al. 2025b). After thoroughly drying at 50 °C in a food drier, the specimens were stored in sealed plastic bags and deposited in the Herbarium of Cryptogams, Kunming Institute of Botany, Academia Sinica (KUN-HKAS), or deposited in the Herbarium of Dali University. Dried materials were sectioned under a stereo microscope, transferred onto slides, and mounted in a 5% KOH solution. The morphological structures were observed as described by reference. For microscopic characters, anatomical and cytological characteristics such as basidia, basidiospores, and cystidia were observed and photographed using a Nikon ECLIPSE Ni-U microscope at magnifications up to × 1000. For SEM studies, fragments of the lamellae of the dried material were taken, sputter-coated with gold, and analyzed with a TM4000Plusll in Dali University (Hitachi Japan). The notation [x/y/z] specifies that measurements were made on x basidiospores measured from y basidiomata of z collections. At least 50 basidiospores and 20 basidia were measured from one basidiomata. Basidiospores dimensions are given as (a–) b–c (–d). Where “a” and “d” refer to the minimum and maximum values of all measurements, respectively, b–c presents the range of 95% of the measured values; Q is the length/width ratio of basidiospores, Qm is the average Q of all basidiospores, and is given as Qm ± standard deviation; av. represents the average value of all basidiospores.
DNA extraction, PCR amplification and sequencing
DNA extraction was conducted according to the protocols provided by Genomic DNA, which was extracted from dried specimens using the Ezup Column Fungi Genomic DNA extraction kit (Sangon China) following the manufacturer’s protocol. Primer pairs for PCR were ITS1/ITS4 for the ITS and LR5/LR0R for the LSU locus. ITS and LSU were amplified as described by Tang et al. (2025a). The PCR amplicons were sent to Sangon Biotech (China) for Sanger sequencing. Sequence reads were assembled in SeqMan II (DNA STAR Inc.).
Sequence alignment and phylogenetic analyses
The newly generated sequences were checked using BioEdit Sequence Alignment Editor version 7.0.4 (Hall 1999) and assembled using SeqMan (DNAstar, Madison, WI, USA). The sequences were then blasted against the GenBank database using the Basic Local Alignment Search Tool (BLAST) to identify the most closely related sequences. Reference sequences were retrieved, minimally adjusted by hand in BioEdit v.7.0.4 first and then aligned using TrimAl (Salvador et al. 2009). Maximum likelihood (ML) analysis was performed separately for each locus, and the concatenated dataset using RAxML-HPC2 v. 8.2.12 (Stamatakis 2014) as implemented on the CIPRES portal, with the GTR+G model for both genes and 1,000 rapid bootstrap (BS) replicates. The GTR+G model was obtained by MrModeltest 2.2. For Bayesian Inference (BI), the best substitution model for each character set was determined with MrModeltest 2.2 on CIPRES, using the Akaike information criterion. Bayesian analysis was performed using MrBayes ver. 3.2.7a as implemented on CIPRES.
Identification of species
All collected specimens were subjected to molecular data acquisition. The resulting sequences were compared with those available in the NCBI database using BLAST searches. Sequence similarity was first evaluated, and specimens showing a sequence similarity of ≥ 99% were preliminarily regarded as conspecific. Subsequently, morphological comparisons were conducted based on diagnostic characters of closely related taxa. When no stable or significant morphological differences were observed, the specimens were further considered to represent the same species.
Results
Phylogenetic analyses
The phylogeny of Clavariaceae, was constructed using LSU and ITS sequences (Fig. 1); 194 reference specimens and 4 new collected specimens were used for phylogenetic analysis, which comprised 1,707 characters with gaps (LSU = 893, ITS = 814). Single-gene analyses were also performed, and topology and clade stability were compared with those from the combined gene analyses. The final ML optimization likelihood is -38361.761232. The matrix included 1,208 distinct alignment patterns, with 29.77% of characters undetermined or missing. Estimated base frequencies were obtained as follows: A = 0.276304, C = 0.188061, G = 0.250759, T = 0.284876; substitution rates AC = 1.486723, AG = 3.234500, AT = 1.882358, CG = 0.530891, CT = 5.979178, GT = 1.0. ML and BI analyses generated similar topologies, so only the ML tree is presented, along with Maximum likelihood bootstrap (BS ≥ 75%) support values and Bayesian inference (BI) posterior probabilities (PP ≥ 0.90) (Fig. 1). The phylogeny revealed that Clavariaceae was divided into six genera: Camarophyllopsis Herink. and Clavaria Vaill. ex L., Clavulinopsis Overeem, Hodophilus R. Heim, Mucronella Fr., and Ramariopsis (Donk) Corner. Our specimens belong to the Clavaria clade and are clearly distinct from other Clavaria species.
Maximum likelihood phylogeny of ITS and LSU sequence data of Clavaria, Clavulinopsis, Hodophilus, Mucronella and Ramariopsis, Camarophyllopsis was chosen as the outgroup. The new species are highlighted in red; the holotype of each species is in bold.
In the phylogeny of Marasmius, which was constructed using ITS and LSU sequences, 65 reference specimens were analyzed, including 38 ITS and 60 LSU. The concatenated ITS and LSU sequence dataset comprised 2,104 characters. The final ML optimization likelihood is -11743.475326. The matrix included 774 distinct alignment patterns, with 33.06% of characters undetermined or missing. Estimated base frequencies were obtained as follows: A = 0.258687, C = 0.198439, G = 0.278994, T = 0.263879; substitution rates AC = 1.195069, AG = 2.359518, AT = 1.442852, CG = 0.692961, CT = 6.013312, GT = 1.0. ML and BI analyses generated similar topologies (Fig. 4), so only the ML tree is presented, along with Maximum likelihood bootstrap (BS ≥ 75%) support values and Bayesian inference (BI) posterior probabilities (PP ≥ 0.90) (Fig. 1). The phylogeny inferred from the ITS and LSU sequences showed that Marasmius lidaensis sp. nov. clustered within the Marasmius clade. The new species M. lidaensis formed a sister lineage to M. maximus Hongo (BRNM714571) and M. brunneospermus (BRNM 714669).
Taxonomy
Between 2020 and 2024, a comprehensive investigation was conducted to document the macrofungal species present on campus. The study identified a total of 83 macrofungal species across 11 orders and 35 families (Table 1), based on morphology and molecular sequence data of 485 collections.
The results of the ITS and LSU molecular data comparisons of the three type specimens are shown in Table 2.
Clavaria
lidaensis
Taxon classificationFungiAgaricalesClavariaceae
S.M. Tang & Z.L. Luo sp. nov.
43245800-F955-5D2A-A659-826413B25EDF
Fungal Names: FN 572831
Fig. 2 Chinese Name: 大理大学珊瑚菌(Da Li Da Xue Shan Hu Jun)
Etymology.
The epithet “lidaensis” refers to the type locality, Dali University (abbreviated as “lida”), where the holotype of this species was collected.
Holotype.
China • Yunnan Province, Dali Autonomous Prefecture, Dali University, elev. 2,105 m, September 13, 2020, Jun He, Sj615 (HKAS 148681).
Description.
Basidiomata incrustations fragile, simple, 10–30 mm tall, 1–4 mm wide, mostly solitary, rarely scattered to gregarious, mostly caespitose-connate at the base; mostly tubular with acute apex, smooth or irregularly ridged. Fertile part subcylindric to fusiform, soft yellow (#deca86) to dark moderate orange (#af9755), apex rounded, concolorous with fertile part, becoming dark orange (#824725) with age. Sterile part narrow, concolorous, without tomentum or mycelial patch at the base. Context fragile, hymenium concolorous. Taste and odor were not recorded.
Clavaria lidaensis. A, B. Basidiomata; C. Basidia; D. Basidioles in the basidiomata top; E. Basidiospores on the optical microscope; F, G. Basidiospores on the SEM. Scale bars: 1 cm (A, B); 10 μm (C–E); 5 μm (F, G).
Basidiospores (3.5–) 4.3–6.8 (–7.1) × (3.1–) 3.3–4.5 (–4.9) µm, av. 6.0 ± 0.6 × 3.8 ± 0.4 µm, thin-walled, hyaline, smooth, broadly ellipsoid to ellipsoid. Basidia 30–51 × 7–8 µm, av. 38 ± 7.5 × 7.4 ± 0.5 µm, thin-walled, hyaline, clavate to subcylindrical, rarely spheropedunculate, with a clamp connection, mostly 4–spores, rarely 2–spores, sterigmata 5–7 µm long. Basidioles in the basidiomata middle, 21–36 × 2–7 µm, av. 31.6 ± 6.4 × 4.5 ± 1.9 µm, clavate to subcylindrical. Basidioles in the basidiomata top, 10–15 × 1–5 µm, av. 12.6 ± 1.4 × 2.5 ± 0.9 µm, flexuose, clavate or subcylindrical. Subhymenium clearly delimited from the context, composed of densely interwoven hyphae. Hyphae near subhymenium 3.4–7.6 µm wide, cylindrical to inflated, thin-walled, hyaline, parallel, without clamp connections. Hyphae distant from subhymenium 12.1–21.2 (–30.3) µm wide, cylindrical to inflated, thin-walled, hyaline, parallel, without clamp connections.
Habitat and distribution.
Solitary in mixed forest dominated by Pinus sp. and various broadleaved trees; only known from Yunnan Province of Dali Autonomous Prefecture.
GenBank accession numbers.
ITS: HKAS 148681, PX353115; HKAS 148682, PX353116; LSU: HKAS 148681, PV802692; HKAS 148682, PV802693.
Additional materials examined.
China • Yunnan Province, Dali Autonomous Prefecture, Dali University, elev. 2,108 m, September 13, 2020, Jun He, Sj617 (HKAS 148682).
Notes.
Clavaria lidaensis and C. amoenoides Corner, K.S. Thind & Anand are similar by yellow basidiomata. However, C. amoenoides has a larger basidiomata (10–30 vs 50–100 mm tall), and longer basidia ( 30–51 vs 45–60 µm long), and basidiospores 4.3–6.8 × 3.3–4.5 vs 6–9.5 × 3–4.5 µm (Roberts 2008); the ITS base differences between C. lidaensis (sj615, holotype) and C. amoenoides (MHHNU10551) is 15.3% (85/555 bp).
Phylogenetically, C. lidaensis is related to C. macounii Peck; however, C. macounii, originally distributed from Canada, has a greenish-yellow or pale-cinereous basidiomata, and smaller basidiomata (1.8–3.5 µm; Peck 1894). The ITS base difference between C. lidaensis (sj615, holotype) and C. macounii (PK1536) is 10.5% (85/810).
Clavaria
minirubella
Taxon classificationFungiAgaricalesClavariaceae
S.M. Tang & Z.L. Luo sp. nov.
4C40C5BB-C938-515C-BF2E-6D8EA23C4332
Fungal Names: FN 572832
Fig. 3 Chinese Name: 小红珊瑚菌(Xiao Hong Shan Hu Jun)
Etymology.
The species epithet “minirubella” refers to the small and red basidiomata.
Holotype.
China • Yunnan Province, Dali Autonomous Prefecture, Dali University, elev. 2,005 m, September 13, 2020, Jun He, Sj619 (HKAS 148689).
Description.
Basidiomata fragile, simple, 10–20 mm tall, 1–3 mm wide, mostly solitary, rarely scattered to gregarious; mostly tubular with obtuse apex, smooth. Fertile part subcylindric to fusiform, dark red (#99302d), apex rounded, concolorous with fertile part. Sterile part narrow, slightly dark moderate red (#a95a51), without tomentum or mycelial patch at the base. Context fragile, hymenium concolorous. Taste and odor were not recorded.
Basidiospores (4.1–) 5.1–6.8 (–7.1) × (2.7–) 2.9–3.6 (–3.9) µm, av. 5.4 ± 0.4 × 3.3 ± 0.2 µm, thin-walled, hyaline, smooth, broadly ellipsoid to ellipsoid. Basidia 31–41 × 6–7 µm, av. 35 ± 4.8 × 6.5 ± 0.5 µm, thin-walled, hyaline, clavate to subcylindrical, with a clamp connection, 4–spores, sterigmata 1–3 µm long. Basidioles in the basidiomata middle, 22–33 × 3–6 µm, av. 28.3 ± 3.9 × 4.2 ± 0.7 µm, clavate to subcylindrical. Basidioles in the basidiomata top, 10–20 × 1–4 µm, av. 15.3 ± 1.2 × 2.8 ± 0.5 µm, mostly flexuose, rarely clavate. Subhymenium clearly delimited from the context, composed of densely interwoven hyphae. Hyphae near subhymenium 2.8–7.6 µm wide, cylindrical to inflated, thin-walled, hyaline, parallel, without clamp connections. Hyphae distant from subhymenium 7.2–12.1 µm wide, cylindrical to inflated, thin-walled, hyaline, parallel, without clamp connections.
Clavaria minirubella. A, B. Basidiomata; C. Basidioles in the basidiomata top; D. Basidioles in the basidiomata middle; E. Basidia; F. Basidiospores on the optical microscope; G, H. Basidiospores on the SEM. Scale bars: 1 cm (A, B); 10 μm (C–E); 2 μm (F, G).
Habitat and distribution.
Solitary in mixed forest dominated by Pinus sp. and various broadleaved trees; only known from Yunnan Province of Dali Autonomous Prefecture.
GenBank accession numbers.
ITS: HKAS 148689, PX353114; HKAS 148690, PX353113; LSU: HKAS 148689, PV802694; HKAS 148690, PV802695.
Additional materials examined.
China • Yunnan Province, Dali Autonomous Prefecture, Dali University, elev. 2,005 m, September 13, 2020, Jun He, Sj620 (HKAS 148690).
Notes.
Our multi-locus phylogenetic analysis indicates that C. minirubella is related to C. salentina Agnello & Baglivo. However, C. salentina is originally distributed from Italy, and the ITS base difference between the specimen C. salentina (AMB_010297) and C. minirubella (sj619, holotype) is 12.0% (98/814).
Maximum likelihood phylogeny of ITS1-5.8S-ITS2 and LSU sequence data of Marasmius, Cirnioellis scabella, C. maxima and C. zonata were chosen as the outgroup. ML bootstrap (≥ 70%) and posterior probabilities (≥ 0.90) are indicated above branches or in front of the branch leading to each node. The new species are highlighted in red; the holotype of each species is in bold.
Morphologically, Clavaria minirubella can be easily confused with Clavulinopsis sulcata Overeem because of its strong orange basidiomata (van Overeem 1923). However, the ITS base difference between the specimen C. minirubella (sj619, holotype) and Clavulinopsis sulcata (PDD_111965) is 31.8% (205/644), thus they are considered to be separate genera.
Marasmius
lidaensis
Taxon classificationFungiAgaricalesMarasmiaceae
S.M. Tang & Z.L. Luo sp. nov
418F2522-99DE-5ABF-9973-4BF31EBB1587
Fungal Names: FN 572833
Figs 5, 6 Chinese Name: 大理大学小皮伞(Da Li Da Xue Xiao Pi San)
Etymology.
The epithet “lidaensis” refers to the type locality, Dali University (abbreviated as “lida”), where the holotype of this species was collected.
Holotype.
China • Yunnan Province, Dali Autonomous Prefecture, Dali University, elev. 2,005 m, September 1, 2020, Jun He, sj523 (HKAS 148679).
Description.
Basidiomata medium-sized. Pileus 20–40 mm in diam., hemispherical when young, becoming convexo-applanate to plano-concave or concave with age, dark orange (#b07530) at center, becoming slightly desaturated yellow (#cabc88) with margin, umbonate of center; aspects exceeding lamellae of margin; distinct, straight, and smooth on the surface; context 1–2 mm thin, tough. Lamellae 2–4 mm in diam., arcuate when young, ventricose with age, white (#ffffff); attachment emarginate; lamella edge even or entire. Stipe 30–60 × 1–3 mm, cylindrical, white (#ffffff) on the upwards, becoming dark orange (#724620) with downward, hollow; context soft and tough surface, white (#ffffff) to pale cream; abundant floccose on the surface and mycelium grown at the base. Taste not distinctive.
Basidiomata of Marasmius lidaensis. A. Basidiomata grow in the forest; B. Basidiomata grow on the grass. Scale bars: 1 cm.
Basidiospores (5.2–) 5.6–10.2 (–11.5) × (4.2–) 4.5–6.3 (–5.8) μm, av. 8.6 ± 1.03 × 5.5 ± 0.48, Q = 1.1–1.7 μm, ellipsoid, smooth, hyaline, thin-walled. Basidia 35–42 × 5–7 μm, av. 38.7 ± 4.5 × 5.8 ± 1.32 μm, clavate, with a clamp connection, 4–spored, sterigmata 1–3 μm. Basidioles 25–52 × 4–6 μm, av. 31 ± 5.2 × 5.2 ± 1.42 μm, scattered, cylindrical, clavate. Cheilocystidia 24–47 × 4–7 μm, av. 30.5 ± 4.6 × 6.1 ± 0.8 μm, mostly clavate, flexuose, rarely branched, thin-walled. Pleurocystidia 21–41 × 4–7 μm, av. 31.3 ± 4.1 × 5.7 ± 0.7 μm, abundant, clavate, flexuose, thin-walled. Lamellae trama hyphae cylindrical or swelling, 4–8 μm in diam., av. 5.9 ± 1.6 μm, thin-walled, with a clamp connection, hyaline in 5% KOH solution. Stipitipellis a cutis of cylindrical, parallel, thin walled, hyaline in 5% KOH solution, 19–90 × 3–5 μm, av. 50.9 ± 23.7 × 4.0 ± 0.6 μm, present connection camp. Pileipellis a hymeniderm composed of cells, swelling, 7–17 × 4–11 μm, av. 14.8 ± 3.6 × 8.8 ± 2.0 μm, parallel, hyaline in 5% KOH solution, with a clamp connection.
Marasmius lidaensis (HKAS 148679, holotype). A. Basidia; B. Pleurocystidia; C. Cheilocystidia; D. Clamp connection; E. Basidiospores; F. Pileipellis; G. Stipitipellis. Scale bars: 10 μm.
Habitat and distribution.
Solitary in mixed forest dominated by Pinus sp. and various broadleaved trees, sometimes basidiomata occur on grass. Only known from the Yunnan Province of Dali Autonomous Prefecture.
GenBank accession numbers.
ITS: HKAS 148679, PX353111; HKAS 148680, PX353112; LSU: HKAS 148679, PV802690; HKAS 148680, PV802691.
Additional materials examined.
China • Yunnan Province, Dali Autonomous Prefecture, Dali University, elev. 2,007 m, July 8, 2021, Jun He, sj525 (HKAS 148680); June 6, 2024, Song-Ming Tang, 2024060601.
Notes.
Marasmius lidaensis is distinguished from other Marasmius species by its clearly striated pileus surface, dark orange at the center, becoming slightly desaturated yellow with a margin; the stipe is white on the upper side, becoming dark orange on the lower side, with a soft and rigid context surface, and is abundant with floccose hairs on the surface.
According to our multi-locus phylogenetic analyses, M. lidaensis was clustered together with M. maximus Hongo (BRNM714571) and M. brunneospermus Har. Takah. (BRNM 714669). However, M. maximus has an ochre-brown to pale brownish to pale ochraceous pileus, and slightly broadened at stipe base, pleurocystidia absent, and irregular lobate cheilocystidia (Hongo 1962); and the ITS base difference between the specimen M. maximus (BRNM 714571) and M. lidaensis (HKAS 148679, holotype) is 1.3% (10/778). Marasmius brunneospermus has a brown to brownish orange pileus, and smaller basidia (35–42 × 5–7 μm vs. 23–27 × 3.3–4.6 μm) (Takahashi 1999); and the ITS base difference between the specimen M. brunneospermus (BRNM 714571) and M. lidaensis (HKAS 148679, holotype) is 8.7% (68/778).
Discussion
In this study, we conducted a comprehensive investigation of the diversity of macrofungi on the Dali University campus and in surrounding habitats in Yunnan Province, China. Through a combination of detailed morphological examination and molecular phylogenetic analysis, we identified 83 macrofungi species. These species were distributed across 11 orders and 35 families, reflecting the rich fungal diversity in this region. The majority of the recorded species were widely distributed taxa, further confirming the ecological stability and suitability of the study area for fungal growth.
Remarkably, during our investigation, we discovered and described three species, Clavaria lidaensis, C. minirubella, and Marasmius lidaensis. So far, 22 species of the genus Clavaria have been reported in China (Table 3). These species are distributed across ten provinces, namely Anhui, Guizhou, Hubei, Hunan, Jiangxi, Jilin, Shanxi, Xizang, Yunnan and Zhejiang (Yan et al. 2020, 2022, 2025), indicating a wide distribution of the genus in China.
China exhibits high species richness of the genus Marasmius, with more than 140 species recorded to date (Song et al. 2009), which reflects the country’s abundant Marasmius germplasm resources. In this study, we systematically collated and comparatively analyzed species morphologically similar to Marasmius lidaensis (Table 4), and ultimately confirmed that this fungus represents a distinct species.
Table 4.: Selected key morphological characteristics of related Marasmius lidaensis species.
The recognition of these novel taxa not only enriches the known diversity of Clavaria and Marasmius but also highlights the importance of continued field surveys and integrative taxonomic approaches in understudied areas such as Dali. Our findings not only expand the known fungal diversity of this region but also contribute valuable taxonomic and ecological data for the global fungal inventory. Overall, this study demonstrates that Dali University harbors a surprisingly high diversity of macrofungi and emphasizes the potential for discovering new fungal resources in this region.
Throughout this survey, 500 macrofungi specimens were collected from the campus and surrounding areas of Dali University. After careful morphological observation combined with molecular analysis, a total of 83 distinct species were recognized (habitat photos of some species see Fig. 7). Among these, the family Agaricaceae proved to be the most species-rich group (see Fig. 8), reflecting its ecological adaptability and dominance in the study area. Following this, representatives of the Polyporaceae and Russulaceae were also frequently encountered, indicating their significant presence and diversity. These findings highlight the broad taxonomic distribution of macrofungi at Dali University and provide important data for regional fungal diversity studies.
Habitat photos of species collected from Dali University campus. A. Abortiporus biennis; B. Agaricus campestris; C. Agaricus cupreobrunneus; D. Agaricus guizhouensis; E. Aleuria aurantia; F. Amanita parvipantherina; G. Auricularia villosula; H. Calvatia craniiformis; I. Candolleomyces candolleanus; J. Cerrena zonata; K. Entoloma clypeatum; L. Flammulina filiformis; M. Gymnopus dysodes; N. Omphalotus flagelliformis; O. Helvella cremeoinvoluta; P. Hygrocybe coccinea; Q. Lactarius kesiyae; R. Cyptotrama asprata.
Dominant families of macrofungi in Dali University.
Taken together, our findings reveal that the Dali University area harbors a surprisingly high diversity of macrofungi despite its limited spatial extent. This diversity is likely supported by the region’s unique climatic conditions, influenced by the Hengduan Mountain system and the Erhai Lake basin, which generate microclimatic gradients favorable for a wide range of fungal guilds. Additionally, the coexistence of natural, semi-natural, and human-managed habitats around the campus may promote species turnover and maintain fungal diversity through enhanced habitat heterogeneity.
Beyond its scientific implications, this study also holds substantial educational and outreach value. The identification of abundant and visually distinctive macrofungi on campus provides an excellent opportunity for field-based teaching and public science education. Fungal diversity surveys can serve as practical training modules for undergraduate and graduate students, helping them develop skills in taxonomy, ecology, and molecular systematics. Moreover, the documentation and illustration of local fungal species can be incorporated into fungal biodiversity exhibitions, campus nature trails, and digital databases, promoting ecological awareness among students and the broader community. Such initiatives not only enhance biodiversity literacy but also foster a sense of environmental stewardship. The discovery of new species within a university setting vividly demonstrates that scientific discovery can occur in everyday environments, inspiring public curiosity and participation in biodiversity conservation.
In conclusion, this study provides a baseline assessment of macrofungal diversity in the Dali University area and underscores the ecological, taxonomic, and educational significance of campus-based biodiversity research. Continued and expanded surveys across seasons and habitats will be crucial for capturing the full spectrum of fungal diversity and for integrating these findings into long-term monitoring, conservation planning, and science communication efforts in Yunnan and beyond.
Supplementary Material
XML Treatment for Clavaria lidaensis
XML Treatment for Clavaria minirubella
XML Treatment for Marasmius lidaensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Antonín V, Ryoo R, Shin HD (2010) Marasmioid and gymnopoid fungi of the Republic of Korea. 2. Marasmius sect. Globulares. Persoonia 24(1): 49–59. 10.3767/003158510 X 496107 PMC 289015820664760 · doi ↗ · pubmed ↗
- 2Arenas MC, Tadiosa ER, Reyes RG (2018) Taxonomic inventory based on physical distribution of macrofungi in Mt. Maculot, Cuenca, Batangas, Philippines. International Journal of Biology, Pharmacy and Allied Sciences 7(5): 672–687. 10.31032/IJBPAS/2018/7.5.4420 · doi ↗
- 3Azteni M, Gennari ANT, Nicoletti L (2022) Prima segnalazione italiana di Clavariaflavostellifera. Rivista micologica romana. Bollettino dell’Associazione Micologica Ecologica Romana 116: 69–80. 10.57624/AMER.2022.02 · doi ↗
- 4Chai ZY, Luo GX, Huo GH, Chen MH, Ye YJ (2014) Diversity research on macrofungi on campus of Meiling Mountain in Jiangxi. Biological Disaster Science 37(1): 41–49. 10.1155/2014/761257 [In Chinese] · doi ↗
- 5Chen H, Xu YQ, Zeng H, Hu YP, Wang SN, Yan JQ (2025) Four new species of Marasmius subgenus Globulares (Marasmiaceae, Agaricales) from subtropical regions of China. Myco Keys 120: 317–338. 10.3897/mycokeys.120.157997 PMC 1237417140859949 · doi ↗ · pubmed ↗
- 6Corner EJH (1950) A monograph of Clavaria and allied genera. Oxford University press, London.
- 7Corner EJH (1970) Supplement to “A monograph of Clavaria and allied genera”. Beihefte zur Nova Hedwigia 33: 1–299.
- 8Corrales A, Koch RA, Vasco-Palacios AM, Smith ME, Ge ZW, Henkel TW (2022) Diversity and distribution of tropical ectomycorrhizal fungi. Mycologia 114(6): 919–933. 10.1080/00275514.2022.211528436194092 · doi ↗ · pubmed ↗