Three new species and two new records of Hydnum (Hydnaceae, Cantharellales) from the Dabie Mountains, China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
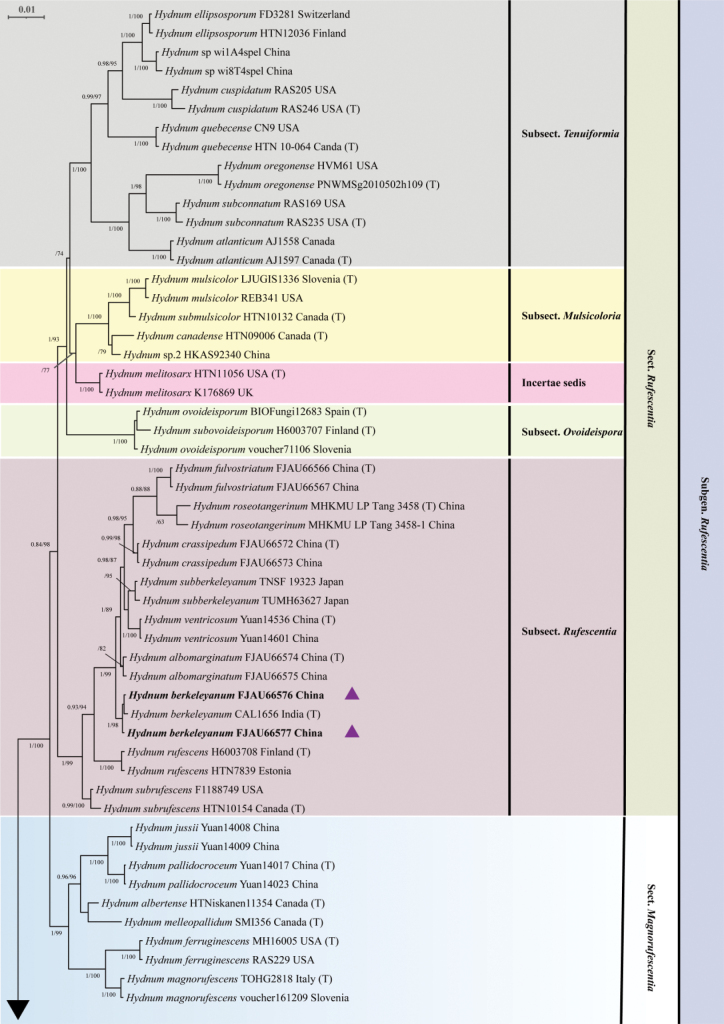 Figure 1
Figure 1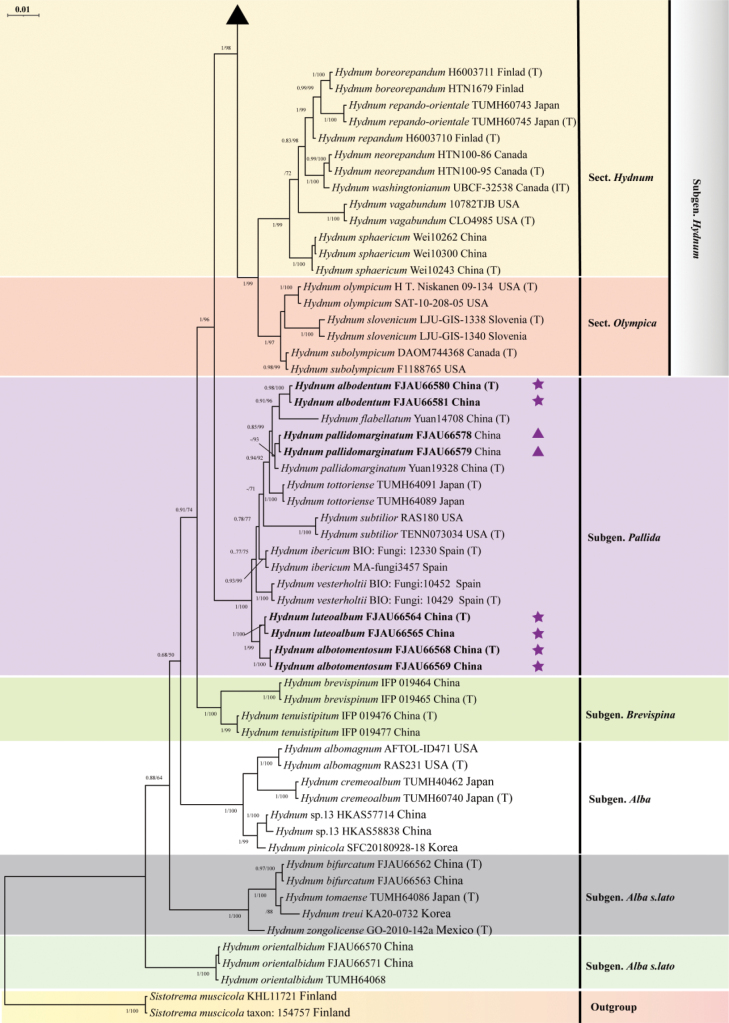 Figure 2
Figure 2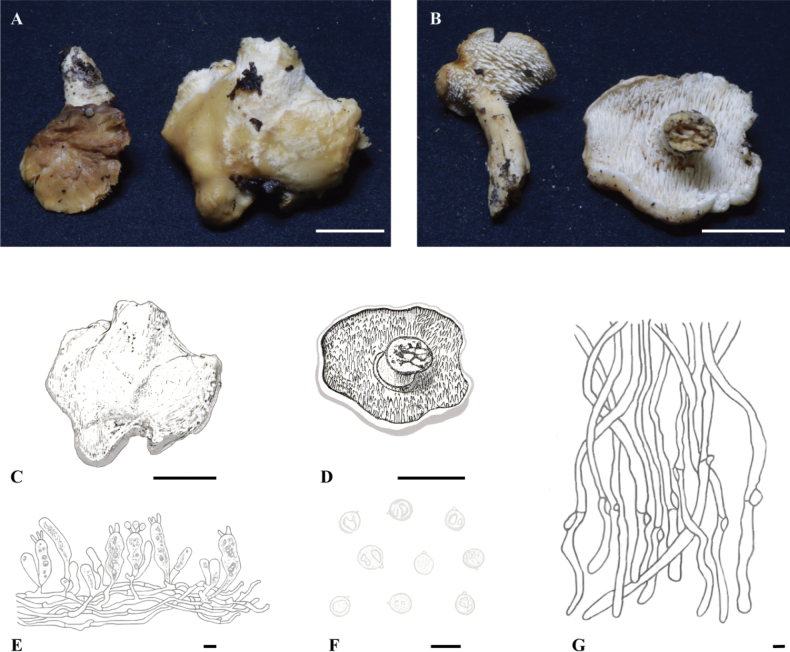 Figure 3
Figure 3 Figure 4
Figure 4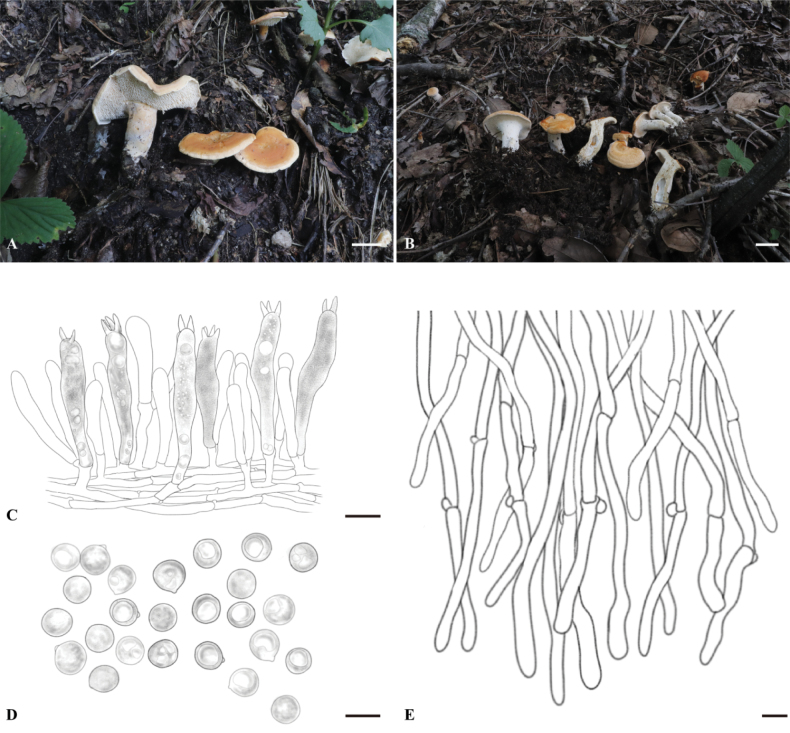 Figure 5
Figure 5 Figure 6
Figure 6 Figure 7
Figure 7| Species | Specimen voucher | GenBank No. | Country | References | ||
|---|---|---|---|---|---|---|
| nrLSU | ITS | |||||
|
| H T. Niskanen 11-354 ( | – |
| – | Canada | ( |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| AFTOL-ID 471 |
|
|
| USA | ( |
|
| RAS231 ( | – |
| – | USA | ( |
|
| FJAU66574 ( |
|
|
| China | ( |
|
| FJAU66575 |
|
|
| China | ( |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| AJ1597 ( | – |
|
| Canada | ( |
|
| AJ1558 | – |
|
| Canada | ( |
|
| CAL1656 ( |
|
| – | India | ( |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| FJAU66562 ( | – |
|
| China | ( |
|
| FJAU66563 | – |
|
| China | ( |
|
| HTN1679 | – |
| – | Finland | ( |
|
| H6003711 ( | – |
| – | Finland | ( |
|
| IFP 019464 ( |
|
| – | China | ( |
|
| IFP 019465 |
|
| – | China | ( |
|
| HTN09006 ( | – |
| – | Canada | ( |
|
| FJAU66572 ( |
|
|
| China | ( |
|
| FJAU66573 |
|
|
| China | ( |
|
| TUMH40462 | – |
| – | Japan | ( |
|
| TUMH60740 ( | – |
| – | Japan | ( |
|
| RAS246 ( | – |
| – | USA | ( |
|
| RAS205 | – |
| – | USA | ( |
|
| FD3281 |
|
| – | Switzerland | ( |
|
| HTN 12036 | – |
| – | Finland | ( |
|
| MH16005 ( | – |
| – | USA | ( |
|
| RAS229 | – |
| – | USA | ( |
|
| Yuan14708 ( |
|
| – | China | ( |
|
| FJAU66566 ( |
|
| – | China | ( |
|
| FJAU66567 |
|
| – | China | ( |
|
| BIO: | – |
| – | Spain | ( |
|
| MA-fungi 3457 | – |
| – | Spain | ( |
|
| Yuan14008 |
|
|
| China | ( |
|
| Yuan14009 |
|
|
| China | ( |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| voucher 161209 |
|
| – | Slovenia | ( |
|
| TOHG2818 ( | – |
| – | Italy | ( |
|
| HTN11056 ( | – |
| – | USA | ( |
|
| K 176869 | – |
| – | UK | ( |
|
| SMI356 ( | – |
| – | Canada | ( |
|
| LJUGIS1336 ( | – |
| – | Slovenia | ( |
|
| REB341 | – |
| – | USA | ( |
|
| HTN100-95 ( | – |
| – | Canada | ( |
|
| HTN100-86 | – |
| – | Canada | ( |
|
| H T. Niskanen 09-134 ( | – |
| – | USA | ( |
|
| SAT-10-208-05 | – |
| – | USA | ( |
|
| HVM61 | – |
| – | USA | ( |
|
| PNMSg2010502h109 ( | – |
| – | USA | ( |
|
| TUMH:64068 | – |
| – | Japan | ( |
|
| FJAU66570 |
|
|
| China | ( |
|
| FJAU66571 |
|
|
| China | ( |
|
| voucher71106 | – |
| – | Slovenia | ( |
|
| BIO Fungi12683 ( | – |
| – | Spain | ( |
|
| Yuan14017 ( |
|
|
| China | ( |
|
| Yuan14023 |
|
|
| China | ( |
|
| Yuan13928a ( |
|
|
| China | ( |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| SFC20180928-18 |
|
|
| Korea | ( |
|
| HTN 10-064 ( | – |
| – | Canada | ( |
|
| CN9 | – |
| – | USA | ( |
|
| TUMH60745 ( | – |
| – | Japan | ( |
|
| TUMH60743 | – |
| – | Japan | ( |
|
| H6003710 ( | – |
| – | Finland | ( |
|
| MHKMU LP Tang 3458 ( |
|
|
| China | ( |
|
| MHKMU LP Tang 3458-1 |
|
|
| China | ( |
|
| H6003708 ( | – |
| – | Finland | ( |
|
| HTN7839 | – |
| – | Estonia | ( |
|
| LJU-GIS-1338 ( | – |
| – | Slovenia | ( |
|
| LJU-GIS-1340 | – |
| – | Slovenia | ( |
| wi1A4spel | – |
| – | China | ( | |
| wi8T4spel | – |
| – | China | ( | |
| HKAS57714 |
|
| – | China | ( | |
| HKAS58838 |
|
| – | China | ( | |
| HKAS92340 |
|
| – | China | ( | |
|
| Wei10243 ( |
|
|
| China | ( |
|
| Wei10300 |
|
|
| China | ( |
|
| Wei10262 |
|
|
| China | ( |
|
| TNSF 19323 | – |
| – | Japan | ( |
|
| TUMH63627 | – |
|
| Japan | ( |
|
| RAS235 ( | – |
| – | USA | ( |
|
| RAS169 | – |
| – | USA | ( |
|
| HTN10132 ( | – |
| – | Canada | ( |
|
| F1188765 |
|
| – | USA | ( |
|
| DAOM744368 ( | – |
| – | Canada | ( |
|
| H6003707 ( | – |
| – | Finland | ( |
|
| HTN10154 ( | – |
| – | Canada | ( |
|
| F1188749 |
|
| – | USA | ( |
|
| RAS180 | – |
| – | USA | ( |
|
| TENN073034 ( | – |
| – | USA | ( |
|
| IFP 019476 ( |
|
| – | China | ( |
|
| IFP 019477 |
|
| – | China | ( |
|
| TUMH64086 ( | – |
| – | Japan | ( |
|
| TUMH64091 ( | – |
|
| Japan | ( |
|
| TUMH64089 | – |
|
| Japan | ( |
|
| KA20-0732 |
|
|
| Korea | ( |
|
| 10782TJB | – |
| – | USA | ( |
|
| CLO4985 ( | – |
| – | USA | ( |
|
| Yuan14536 ( |
|
|
| China | ( |
|
| Yuan14601 |
|
|
| China | ( |
|
| BIO: | – |
| – | Spain | ( |
|
| BIO: | – |
| – | Spain | ( |
|
| UBCF-32538 (isotype) | – |
| – | Canada | ( |
|
| GO-2010-142a ( | – |
| – | Mexico | ( |
|
| KHL 11721 | – |
| – | Sweden | ( |
|
| taxon:154757 | – |
| – | Sweden | ( |
| 1 | Basidiomata more or less white to cream yellow |
|
| – | Basidiomata yellow to orange |
|
| 2 | Pileus white |
|
| – | Pileus cream yellow |
|
| 3 | Pileus ≤ 30 mm wide |
|
| – | Pileus ≥ 30 mm wide |
|
| 4 | Habitat in broad-leaved forests |
|
| – | Habitat in |
|
| 5 | Pileus ≥ 60 mm wide |
|
| – | Pileus < 60 mm wide |
|
| 6 | Basidiospores ≤ 5 μm long on average |
|
| – | Basidiospores > 5 μm long on average |
|
| 7 | Pileus > 60 mm wide |
|
| – | Pileus ≤ 60 mm wide |
|
| 8 | Habitat in broad-leaved forests |
|
| – | Habitat in |
|
| 9 | Spines ≤ 3 mm long |
|
| – | Spines > 3 mm long |
|
| 10 | Spines pale orange |
|
| – | Spines cream yellow |
|
| 11 | Basidiomata yellowish-white |
|
| – | Basidiomata orange |
|
| 12 | Basidiospores < 6 μm long on average |
|
| – | Basidiospores > 6 μm long on average |
|
| 13 | Habitat in angiosperm forests |
|
| – | Habitat in mixed forests |
|
| 14 | Basidiospores < 7.5 μm long on average |
|
| – | Basidiospores > 7.5 μm long on average |
|
| 15 | Pileus > 40 mm wide |
|
| – | Pileus < 40 mm wide |
|
| 16 | Spines > 3 mm long |
|
| – | Spines ≤ 3 mm long |
|
| 17 | Habitat in September |
|
| – | Habitat in August |
|
| 18 | Habitat in |
|
| – | Habitat in |
|
| 19 | Basidiospores 7–8 μm long on average |
|
| – | Basidiospores > 8 μm long on average |
|
| 20 | Pileus ≥ 60 mm wide |
|
| – | Pileus < 60 mm wide |
|
| 21 | Habitat in |
|
| – | Habitat in |
|
| 22 | Basidiospores 8–9 μm long on average |
|
| – | Basidiospores > 9 μm long on average |
|
| 23 | Spines white |
|
| – | Spines orange-white or cream-yellow |
|
| 24 | Spines ≤ 3 mm long |
|
| – | Spines ≥ 3 mm long |
|
| 25 | Spines cream-yellow |
|
| – | Spines orange-white to brownish-orange |
|
| 26 | Basidiospores 8.1–8.5 μm long on average |
|
| – | Basidiospores > 8.5 μm long on average |
|
| 27 | Spines < 2 mm long |
|
| – | Spines > 2 mm long |
|
| 28 | Habitat in angiosperm forests |
|
| – | Habitat in |
|
| 29 | Basidiospores Q < 1.1. |
|
| – | Basidiospores Q > 1.1 |
|
| 30 | Pileus < 50 mm wide |
|
| – | Pileus > 50 mm wide |
|
| 31 | Spines orange-white to pale-orange |
|
| – | Spines light-yellow to pale-orange |
|
| 32 | Pileus ≥ 30 mm wide |
|
| – | Pileus ≤ 30 mm wide |
|
| 33 | Basidiospores > 9.5 μm long on average |
|
| – | Basidiospores < 9.5 μm long on average |
|
| 34 | Basidiospores Q > 1.3 |
|
| – | Basidiospores Q < 1.3 |
|
| 35 | Pileus ≤ 40 mm wide |
|
| – | Pileus > 40 mm wide |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Plant Pathogens and Fungal Diseases · Lichen and fungal ecology
Introduction
The genus Hydnum (Hydnaceae, Cantharellales), established by Linnaeus in 1753 and typified by H. repandum L. (Niskanen et al. 2018), exhibits a transcontinental distribution, occurring in Asia (Cao et al. 2021; Kim et al. 2023; Qin et al. 2024; Su et al. 2024; Zhang et al. 2024), Australia (Massee 1910; McNabb 1971), Europe (Grebenc et al. 2009; Márquez-Sanz et al. 2023), and North America (Banker 1901; Niskanen et al. 2018; Swenie et al. 2018). Morphologically, it is characterized by a monomitic hyphal system, nodose-septate hyphae, 2–6-sterigmate basidia, and subglobose to obovoid-elliptical, smooth, hyaline to subhyaline (basidiospores filled with granular inclusions and oily droplets), thin-walled basidiospores (Hall and Stuntz 1971; Moncalvo et al. 2006; Niskanen et al. 2018; Swenie et al. 2018). Historically, species delimitation in Hydnum has largely depended on morphological traits, including basidiome dimensions, pileus coloration, and basidiospore size and morphology (Banker 1906; Huhtinen and Ruotsalainen 2006; Olariaga et al. 2012). Nevertheless, pronounced morphological characteristic overlaps – especially within the H. rufescens complex – have long posed challenges to accurate identification, resulting in an underestimation of the genus’s diversity (Sugawara et al. 2022).
The integration of molecular phylogenetics with traditional morphological analyses has substantially advanced our understanding of Hydnum diversity. For example, Feng et al. (2016), using multi-marker datasets (ITS, RPB1, and tef1-α), revealed at least 31 Hydnum species and preliminarily divided them into four groups (Albomagnum, Vesterholtii, Rufescens, and Repandum), although systematic taxonomic treatments of these lineages were not provided. Subsequently, Niskanen et al. (2018) combined morphological evidence with ITS data, increasing the number of recognized species worldwide to 49, including 22 newly described taxa. They also proposed a classification framework comprising four subgenera (Alba, Hydnum, Pallida, and Rufescentia), four sections (Hydnum, Olympica, Magnorufescentia, and Rufescentia), and three subsections (Mulsicoloria, Rufescentia, and Tenuiformia). This infrageneric system was further supported by Cao et al. (2021), who conducted a comprehensive multi-marker phylogenetic analysis (ITS, nrLSU, tef1-α, RPB2, and mtSSU), confirmed 17 genera within Hydnaceae, and introduced the new subgenus Brevispina. Collectively, these studies highlight the importance of integrative taxonomic approaches in resolving long-standing ambiguities and revealing previously unrecognized diversity within the genus.
Globally, more than 60 Hydnum species have been recognized, with 32 described from temperate regions such as Europe and North America (Grebenc et al. 2009; Olariaga et al. 2012; Vizzini et al. 2013; Feng et al. 2016; Márquez-Sanz et al. 2023; Qin et al. 2024). In contrast, records from subtropical regions were historically limited. Prior to 2016, only three species – H. repandum, H. repandum var. album, and H. rufescens – were reported from China (Ying and Zang 1994; Zang et al. 1996). In recent years, research on Hydnum in China has intensified, with 32 species now reported nationwide, more than half of which were originally described from tropical and temperate regions of the country (Feng et al. 2016; Cao et al. 2021; Qin et al. 2024; Su et al. 2024; Zhang et al. 2024). The Anhui Tianma National Nature Reserve, located in the core area of the Dabie Mountains, represents a critical transitional zone between subtropical and warm-temperate ecosystems in China and is recognized as a key area for biodiversity conservation (Xie et al. 2004; Xu and Pan 2019). However, the distribution of Hydnum in this region remains poorly documented, and no comprehensive taxonomic treatment has been published to date.
Based on detailed morphological observations and multi-marker phylogenetic evidence, the present study describes three new species of Hydnum and reports two newly recorded Hydnum species from the Dabie Mountains. These findings contribute to a more complete understanding of the diversity and distribution of Hydnum in subtropical China, particularly within the Dabie Mountains region.
Materials and methods
Specimen collecting
Ten specimens were collected from July to October 2023 in the Tianma National Nature Reserve (31°10'–31°20'N, 115°20'–115°50'E), Anhui Province, China. After we thoroughly documented the fresh macroscopic features, the specimens were oven-dried at 40–45 °C and deposited in the Mycology Herbarium of Jilin Agricultural University (HJAU).
Morphological studies
Macroscopic characters were documented using field notes and basidiomata photographs. Colorimetric data followed the Methuen Handbook (Kornerup and Wanscher 1978). For micromorphological analysis, dried specimens were examined under a Carl Zeiss Axio Lab A1 compound microscope (Carl Zeiss AG, Jena, Germany). The micrographs of basidia and basidiospores were acquired using a Zeiss Scope A1 microscope (Carl Zeiss AG, Jena, Germany) (Appendix 2). Specimens (collected from each basidiocarp: five spines, approximately 0.2 cm^2^ of pileipellis, and 0.2 cm^2^ of stipitipellis) were mounted in a staining solution prepared with 3% (w/v) KOH (Sangan Biotech, Shanghai, China, A610441-0500), supplemented with 1% (w/v) Congo Red (Sangan Biotech, A427695-0500) when necessary. Amyloid reactions were tested using Melzer’s reagent [3 g KI (Sangan Biotech, A610443-0050), 1 g I_2_ (Sangan Biotech, A500538-0100), 40 g CCl_3_CH(OH)2 (Sangan Biotech, A500288-0250), dissolved in 40 mL distilled water]. Spore dimensions (n = 100 per species) were recorded as (a) b–c (d) (with 99% of values falling within the b–c range), where mean length (avg. L) and width (avg. W) were calculated by excluding extreme values; Q denotes the L/W ratio (L: spore length, W: spore width).
DNA extraction, PCR, and sequencing
We extracted genomic DNA from samples using the NuClear Plant Genomic DNA Kit (CoWin Biotech Co., Ltd., Jiangsu, China), following the manufacturer’s protocols. We selected the primer pairs ITS1F/ITS4 (White et al. 1990; Gardes and Bruns 1993) and tef1F/tef1R (Feng et al. 2016) to amplify and sequence the ITS and tef1-α markers, respectively. For PCR amplification, we prepared a 25 μL reaction mixture containing 2 μL genomic DNA template, 12.5 μL SanTaq PCR Master Mix (Sangon Biotech, Shanghai, China), 8.5 μL ddH_2_O, and 1 μL of each forward and reverse primer (10 μM). The PCR cycling parameters were set as follows: initial denaturation at 95 °C for 4 min, followed by 35 cycles of 45 s at 95 °C, 45 s at 56 °C, and 1.5 min at 72 °C, with a final extension at 72 °C for 10 min for ITS; and 95 °C for 3 min, followed by 35 cycles of 40 s at 95 °C, 45 s at 56 °C, and 1.5 min at 72 °C, with a final extension at 72 °C for 7 min for tef1-α. After amplification, PCR products were purified with the Genview High-Efficiency Agarose Gels DNA Purification Kit (Gen-View Scientific Inc., Galveston, TX, USA). Purified amplicons were then subjected to Sanger sequencing by Comate Bioscience Co., Ltd. (Jilin, China). Sequence assembly was performed using SeqMan software (DNASTAR, v7.1), and the consensus sequences were deposited in the NCBI GenBank database.
Phylogenetic analyses
The newly generated sequences have been deposited in the National Center for Biotechnology Information (NCBI) database (https://www.ncbi.nlm.nih.gov/). The sequences for the multi-marker (ITS–nrLSU–tef1-α) phylogenetic tree were derived from previous studies (Feng et al. 2016; Cao et al. 2021; Qin et al. 2024; Su et al. 2024; Zhang et al. 2024) and highly relevant taxonomic unit sequences identified via BLAST searches (coverage: 97–100% and identity: 97–100%) (Table 1). Sistotrema muscicola (Nilsson et al. 2006) was used as the outgroup. The workflow included (1) independent alignments of the ITS, nrLSU, and tef1-α sequences, which were aligned in batches with MAFFT v7.505 (Katoh and Standley 2013) using the auto strategy and normal alignment mode; and (2) the sequence alignments were manually adjusted in BioEdit v7.1.3 (Hall 1999). Subsequently, the ITS, nrLSU, and tef1-α regions were concatenated using the sequence concatenation tool in the Alignment toolbar of PhyloSuite v1.2.3 (Xiang et al. 2023); (3) ModelFinder v2.2.0 (Kalyaanamoorthy et al. 2017) was used to select the best-fit model using the BIC criterion; (4) Bayesian inference was performed using MrBayes v3.2.7a (Ronquist et al. 2012) with 5 million generations (sampling every 2000 generations, 25% burn-in, and split frequency = 0.004); (5) maximum likelihood analysis was performed using IQ-TREE v2.2.0 (Nguyen et al. 2015) with 4950 ultrafast bootstraps, as well as the Shimodaira–Hasegawa-like approximate likelihood-ratio test (Guindon et al. 2010); and (6) the tree was visualized for topological structure using the online platform ITOL v7.2.1 (https://itol.embl.de). The phylogenetic tree, macroscopic photographs, and microscopic photographs were edited and typeset using Adobe Illustrator 2020 (Adobe, San Jose, CA, USA). In accordance with phylogenetic reporting standards (Leebens-Mack et al. 2006), we deposited the results of phylogenetic analyses (BI and ML trees) and alignment data of concatenated sequences into the online database figshare (https://doi.org/10.6084/m9.figshare.29858450, V2).
Results
Phylogenetic analyses
In the concatenated (ITS–nrLSU–tef1-α) dataset, 99 samples and 156 sequences underwent phylogenetic analysis. The multiple sequence alignment featured 111 sequences, with 3831 column sites and 1056 distinct patterns, including 3128 (81.65%) constant sites, 125 singleton sites, and 578 (15.09%) parsimony-informative sites. In both the maximum likelihood (ML) phylogenetic analysis and Bayesian inference (BI) analysis, the best-fit partitioned models were selected using the Bayesian Information Criterion (BIC). Specifically, for the maximum likelihood (ML) analysis, the best-fit partitioned models were HKY+F+G4 for ITS, HKY+F+R2 for nrLSU, and TNe+G4 for tef1-α. Similarly, for the Bayesian inference (BI) analysis, the best-fit partitioned models were HKY+F+G4 for ITS, HKY+F+I+G4 for nrLSU, and SYM+G4 for tef1-α. There was topological consistency between the maximum likelihood (ML) and Bayesian inference (BI) methods, and the BI tree is provided in this study (Figs 1, 2). A total of 20 newly generated sequences from this study, including ten ITS sequences and ten tef1-α sequences, were deposited in GenBank.
Phylogenetic tree inferred from Bayesian inference (BI) based on the combined dataset (ITS–nrLSU–tef1-α). The values on the nodes represent Bayesian posterior probability (left, BPP ≥ 0.95) and maximum likelihood bootstrap support (right, MLBS ≥ 75%), respectively. Novel sequences generated in this study appear in boldface type; holotype specimens are marked with (T); new taxa are marked with filled pentagrams (Purple star); and newly recorded taxa are marked with solid triangles (Purple up-pointing triangle).
Phylogenetic tree inferred from Bayesian inference (BI) based on the combined dataset (ITS–nrLSU–tef1-α). The values on the nodes represent Bayesian posterior probability (left, BPP ≥ 0.95) and maximum likelihood bootstrap support (right, MLBS ≥ 75%), respectively. Novel sequences generated in this study appear in boldface type; holotype specimens are marked with (T); new taxa are marked with filled pentagrams (Purple star); and newly recorded taxa are marked with solid triangles (Purple up-pointing triangle).
The phylogenetic tree revealed six major clades, corresponding to six subgenera: Rufescentia, Hydnum, Pallida, Brevispina, Alba s.l., and Alba. The newly generated sequences were divided into five species-level clades with high support values (BPP = 0.98–1; MLBS = 93–100%). Among them, FJAU66576/FJAU66577 (BPP/MLBS = 1/98) were nested within the H. berkeleyanum clade and formed a distinct lineage within the subsect. Rufescentia; H. pallidomarginatum (MLBS = 93), H. albodentum (BPP/MLBS = 0.98/100), H. luteoalbum (BPP/MLBS = 1/100), and H. albotomentosum (BPP = 1, MLBS = 100) each formed a distinct lineage within the subgenus Pallida; FJAU66578/FJAU66579 were nested within the H. pallidomarginatum clade; H. albodentum is closely related to H. flabellatum; and H. luteoalbum and H. albotomentosum form a sister clade (Figs 1, 2).
Taxonomy
Hydnum
albodentum
Taxon classificationFungiCantharellalesHydnaceae
Yonglan Tuo, Bo Zhang & Yu Li sp. nov.
BF0FE5E1-968E-524B-90C2-E7AA66EAA8FC
Fungal Names: FN 572951
Etymology.
The specific epithet “albodentum” refers to the pure white coloration of the fungal spines.
Holotype.
China • Anhui Province, Lu’an City, Jinzhai County, Yanzihe Town, 31°18'20"N, 115°55'32"E, Elevation: ca. 752 m. on soil in mixed forests (Quercus serrata Thunb. and Cunninghamia lanceolata (Lamb.) Hook.), 10 September 2023, Yonglan Tuo FJAU66580. GenBank accession numbers: ITS = PV997562, tef1-α = PX026328.
Macroscopic and microscopic features of H. albodentum (FJAU66580, holotype). A–D. Basidiomata; E. Hymenium and subhymenium; F. Basidiospores; G. Pileipellis. Scale bars: 2 cm (A–D); 10 μm (E–G).
Diagnosis.
H. albodentum differs from other Hydnum species primarily in its medium-sized (7.93–6.78 μm), elliptical spores (Q = 1.13–1.25), and the presence of a white tomentum covering the pileus margin.
Description.
Basidiocarps small, solitary, 25.5–40.5 mm in height, fleshy when fresh. Pileus 27.5–55.5 mm wide, brownish orange to light brown (6C6–6D7), convex to plano-convex when mature, surface smooth, margin incurved and covered with white (4A1) tomentum. Context 2.5–4.5 mm thick, white (4A1). Hymenophore hydnoid, spines fleshy, non-decurrent, conical, sparse, 1–2 spines/mm^2^; surface white (4A1), 3.0–7.5 mm long, 0.5–1.5 mm diameter. Stipe cylindrical, 21.5–35.0 × 8.5–10.5 mm, solid, central, equal to slightly enlarged at the base, white to light brown (4A1–6C6), covered with white basal mycelium, brownish orange when handled. Odor mild or fruity.
Basidiospores (7.0) 8.0–8.5 (9.0) × (5.5) 6.0–7.0 (8.0) μm, av. L = 7.93 μm, av. W = 6.78 μm, Q = (1.0) 1.13–1.25 (1.33), av. Q = 1.17, ellipsoid, thin-walled, smooth, hyaline in 3% KOH, some with granular contents; hilar appendix 0.5–1.0 μm long. Basidia (30.0) 32.5–45.0 (55.0) × (8.0) 9.0–10.0 (12.0) μm, subcylindric, some with granular contents and hyaline oily droplets; sterigmata 2–5, 4.0–5.0 × 0.5–1.0 µm, conical, thin-walled, smooth. Basidioles 25.0–32.0 × 7.0–9.0 μm, numerous, smaller than basidia, some with granular contents. Subhymenium trama filamentous, hyphae 2.5–3.0 μm wide, thin-walled, hyaline in 3% KOH. Hyphae of spines 5.0–7.0 μm, thin-walled, apex cylindrical. Pileipellis composed of cylindrical hyphae, slightly interwoven; terminal elements cylindrical at apex, cells 65.5–105.5 × 8.5–10.0 μm. Stipitipellis composed of cylindrical hyphae, slightly interwoven, 6.0–7.5 μm wide, terminal elements rounded at apex. Clamp connections present.
Habitat and distribution.
The species occurs solitarily in a mixed forest (Q. serrata and C. lanceolata) within the buffer zone of Tianma National Nature Reserve, China (ca. 700–800 m, subtropical monsoon climate). The forest is subject to human disturbance (grazing, thinning, and proximity to villages at approximately 1.5–2.5 km). The specimen was collected at a distance of 45.6 cm from a C. lanceolata tree (DBH: 23.6 cm) and at a distance of 85.6 cm from a Q. serrata tree (DBH: 15.6 cm). Soil temperature was 12.1 °C, soil volumetric water content was 25.3%, soil pH was 5.9, air temperature was 24.2 °C, and relative humidity was 88.5%. Soil temperature, moisture content, and pH were measured at the time of sample collection; air temperature and humidity were monitored over the long term within the sample plot (measurements were taken at 10-minute intervals; these data are mean values calculated over the 3 days preceding and following sample collection).
Additional specimens examined.
China • Anhui Province, Lu’an City, Jinzhai County, 31°18'41"N, 115°55'01"E, Elevation: ca. 742 m. on soil in Q. serrata and C. lanceolata mix forests, 10 September 2023, Yonglan Tuo FJAU66581. GenBank accession numbers: ITS = PV997563, tef1-α = PX026329.
Notes.
Morphologically, H. albodentum is similar to H. pallidomarginatum in having small basidiocarps (27.5–55.5 mm vs. 20.0–35.0 mm), but it is distinct from H. pallidomarginatum in having smaller basidiospores (7.93 × 6.78 μm vs. 8.75 × 6.99 μm) and a brownish orange to light brown pileus.
In the phylogenetic tree, the sequences of H. albodentum cluster together, forming a distinct lineage and a sister clade with H. flabellatum within subgenus Pallida.
Hydnum
albotomentosum
Taxon classificationFungiCantharellalesHydnaceae
Yonglan Tuo, Bo Zhang & Yu Li sp. nov.
A37B082C-C343-56B9-9393-98EFB170648B
Fungal Names: FN 572699
Etymology.
The specific epithet “albotomentosum” refers to the white tomentum covering the pileus and stipe of immature basidiocarps.
Holotype.
China • Anhui Province, Lu’an City, Tianma National Nature Reserve, 31°13'10"N, 115°46'53"E, Elevation: ca. 817 m. on soil in mixed forests (Quercus variabilis Blume and Pinus hwangshanensis W.Y.Hsia), 8 August 2023, Yonglan Tuo FJAU66568. GenBank accession numbers: ITS = PV329851, tef1-α = PP357256.
Macroscopic and microscopic features of H. albotomentosum (FJAU66568, holotype). A–C. Basidiomata; D. Hymenium and subhymenium; E. Basidiospores; F. Pileipellis. Scale bars: 2 cm (A–C); 10 μm (D–F).
Diagnosis.
H. albotomentosum differs from other Hydnum species in its relatively small basidiocarps, short spines (0.5–2.0 mm), and the presence of a white tomentum covering the pileus surface when immature, which disappears upon maturity.
Description.
Basidiocarps small, solitary, 22.0–30.0 mm in height, fleshy when fresh. Pileus 16.5–29.5 mm wide, pale orange to deep orange (5A3–4A8), convex to plano-convex when mature, margin incurved, surface covered with white (4A1) tomentum when immature, tomentum disappearing upon maturity. Context 2.5–5.0 mm thick, white (4A1). Hymenophore hydnoid, spines fleshy, non-decurrent, conical, sparse, 1–2 spines/mm^2^; surface white to yellowish white (4A1–4A2), 0.5–2.0 mm long, 0.25–0.5 mm diameter. Stipe cylindrical, 20.0–30.0 × 7.0–10.0 mm, solid, central, equal to slightly enlarged at the base, white to deep orange (4A1–4A8), covered with a large amount of white basal mycelium; Stipe at first whitish, staining pale brownish orange when handled. Odor mild or fruity.
Basidiospores (6.0) 7.0–8.0 (8.5) × (5.5) 6.5–7.0 (8.0) μm, av. L = 7.11 μm, av. W = 6.72 μm, Q = 1.0–1.14, av. Q = 1.06, globose to subglobose, thin-walled, smooth, hyaline in 3% KOH, some with granular contents; hilar appendix 0.5–1.0 μm long. Basidia (33.0) 36.0–42.0 (46.0) × (7.5) 9.0–10.0 (12.0) μm, clavate to suburniform, some with granular contents and hyaline oily droplets; sterigmata 2–4, 4.0–6.0 × 1.0–2.0 µm, conical, thin-walled, smooth. Basidioles (28.0) 30.0–38.0 (42.0) × (6.0) 8.0–10.0 (12.0) μm, numerous, smaller than basidia, some with granular contents. Subhymenium trama filamentous, hyphae 3.0–4.0 μm wide, thin-walled, hyaline in 3% KOH. Hyphae of spines 6.5–7.0 μm, thin-walled, apex cylindrical. Pileipellis composed of cylindrical hyphae, subparallel to slightly interwoven; terminal elements cylindrical at apex, cells 56.0–105.0 × 5.5–7.0 μm. Stipitipellis composed of cylindrical hyphae, slightly interwoven, 4.0–6.0 μm wide, terminal elements rounded at apex. Clamp connections present.
Habitat and distribution.
The species occurs solitarily in a mixed forest (Q. variabilis and P. hwangshanensis) within the buffer zone of Tianma National Nature Reserve, China (ca. 800–900 m, subtropical monsoon climate). The forest is subject to human disturbance (grazing, thinning, and proximity to villages and towns at approximately 0.5–3.5 km). Specimens were collected at a distance of 139.5 cm from a P. hwangshanensis tree (DBH: 50.6 cm) and at a distance of 85.6 cm from a Q. variabilis tree (DBH: 25.6 cm). Soil temperature was 24.1 °C, soil volumetric water content was 24.2%, soil pH was 5.2, air temperature was 25.6 °C, and relative humidity was 85.6%. Soil temperature, moisture content, and pH were measured at the time of sample collection; air temperature and humidity were monitored over the long term within the sample plot (measurements were taken at 10-minute intervals; these data are mean values calculated over the 3 days preceding and following sample collection).
Additional specimens examined.
China • Anhui Province, Lu’an City, Tianma National Nature Reserve, 31°13'14"N, 115°46'55"E, Elevation: ca. 856 m. on soil in Q. variabilis and P. hwangshanensis mixed forests, 17 July 2023, Yonglan Tuo FJAU66569. GenBank accession numbers: ITS = PV329852, tef1-α = PP357257.
Notes.
Morphologically, H. albotomentosum is similar to H. ibericum in having a white to cream pileus and small basidiocarps (16.5–29.5 mm vs. 22.0–30.0 mm), but it is distinct from H. ibericum in having smaller basidiospores (7.11 × 6.72 μm vs. 8.2 × 6.4 μm).
In the phylogenetic tree, the sequences of H. albotomentosum form a distinct lineage and group within a large subclade with strong support (100% ML and 1.00 BPP) together with H. vesterholtii, H. ibericum, H. subtilior, H. tottoriense, H. flabellatum, H. pallidomarginatum, and H. luteoalbum. Based on the morphological characteristics described above and the phylogenetic results, H. albotomentosum should be classified as a member of subgenus Pallida.
Hydnum
berkeleyanum
Taxon classificationFungiCantharellalesHydnaceae
K. Das, Hembrom, Baghela & Vizzini, 2018
0B977CEE-7F80-5100-9C17-A6B00B29B0CF
Habitat and distribution.
The species grows gregariously in a Q. serrata forest within the core zone of Tianma National Nature Reserve, China (ca. 1000–1100 m, subtropical monsoon climate). It is currently known from India, Pakistan, Anhui Province, and Yunnan Province in China. The specimen was collected at a distance of 105.6 cm from a Q. serrata tree (DBH: 56.8 cm). Soil temperature was 20.6 °C, soil volumetric water content was 17.5%, soil pH was 5.9, air temperature was 23.2 °C, and relative humidity was 91.2%. Soil temperature, moisture content, and pH were measured at the time of sample collection; air temperature and humidity were monitored over the long term within the sample plot (measurements were taken at 10-minute intervals; these data are mean values calculated over the 3 days preceding and following sample collection).
Macroscopic and microscopic features of H. berkeleyanum. A, B. Basidiomata; C. Hymenium and subhymenium; D. Basidiospores; E. Pileipellis. Scale bars: 2 cm (A, B); 10 μm (C–E).
Specimens examined.
China • Anhui Province, Lu’an City, Tianma National Nature Reserve, 31°15'20"N, 115°42'09"E, Elevation: ca. 1049 m, 23 July 2023, Yonglan Tuo (FJAU66576), GenBank accession numbers: ITS = PV875855, tef1-α = PV889364); • Anhui Province, Lu’an City, Tianma National Nature Reserve, 31°15'21"N, 115°42'07"E, Elevation: ca. 1052 m, 16 July 2023, Yonglan Tuo (FJAU66577), GenBank accession numbers: ITS = PV875856, tef1-α = PV889365.
Notes.
No major differences were found between our specimens and the type; however, the habitat of our specimens (Q. serrata forest) differs from that in the original description (mixed forest). Phylogenetic analyses based on multi-marker datasets indicate that the two specimens from Anhui are well nested within the H. berkeleyanum clade.
Hydnum
luteoalbum
Taxon classificationFungiCantharellalesHydnaceae
YonglanTuo, Bo Zhang & Yu Li sp. nov.
EFBEA113-92FF-5D41-AD99-6CB3F1AACF96
Fungal Names: FN 572683
Etymology.
The specific epithet “luteoalbum” refers to the yellowish-white color of the entire basidiocarp.
Holotype.
China • Anhui Province, Lu’an City, Tianma National Nature Reserve, 31°15'43"N, 115°41'41"E, Elevation: ca. 1207 m. on soil in Quercus serrata Thunb. forest, 4 August 2023, Yonglan Tuo FJAU66564. GenBank accession numbers: ITS: PV329847, tef1-α: PP357254.
Macroscopic and microscopic features of H. luteoalbum (FJAU66564, holotype). A–C. Basidiomata; D. Hymenium and subhymenium; E. Basidiospores; F. Pileipellis. Scale bars: 2 cm (A–C); 10 μm (D–F).
Diagnosis.
H. luteoalbum differs from other species of Hydnum by its ensiform-dagger spines, white pileus, and smaller basidiospores (7.0–7.5 × 6.0–6.5 μm).
Description.
Basidiocarps small to medium, solitary, 25.0–30.0 mm in height. Pileus 25.0–60.5 mm wide, white to yellowish white (4A1–4A2), round to irregular, shallowly depressed in the center, discoloration not observed; margin incurved, azonate, white (4A1) tomentum; pileus context 2.0–4.5 mm thick. Hymenophore hydnoid, spines non-decurrent, ensiform, slightly sparse, 1–2 spines/mm^2^, white to yellowish white (4A1–4A2), 5.0–10.0 mm long, 0.5–1.5 mm diameter. Stipe 20.0–35.0 × 7.0–15.0 mm, central to slightly eccentric, solid, surface covered with white (4A1) tomentum. Odor mild or fruity.
Basidiospores (6.0) 7.0–7.5 (8.0) × (5.5) 6.0–6.5 (7.0) μm, av. L = 7.08 μm, av. W = 6.10 μm, Q = (1.00) 1.08–1.17 (1.33), av. Q = 1.16, broadly ellipsoid, thin-walled, smooth, hyaline in 3% KOH, some with granular contents, inamyloid, hilar appendix 0.5–1.0 μm long. Basidia (35.0) 36.0–50.0 (55.0) × (7.0) 8.0–10.0 (11.0) μm, clavate to suburniform, some with granular contents, sterigmata 2–4, 4.0–6.5 × 1.0–2.0 µm, conical, thin-walled, smooth. Basidioles (25.0) 30.0–40.0 (50) × (6.0) 7.0–10.0 (11.0) μm, smaller than basidia, some with granular contents. Subhymenium trama filamentous, hyphae 4.0–5.0 μm wide, thin-walled, hyaline in 3% KOH. Hyphae of spines 7.0–8.0 μm, thin-walled, apex cylindrical. Pileipellis composed of cylindrical hyphae, slightly interwoven, terminal elements cylindrical at apex, cells 60.0–125.0 × 7.0–11.0 μm. Stipitipellis composed of cylindrical hyphae, slightly interwoven, 6.0–14.0 μm wide, terminal elements dilated at apex. Clamp connections present.
Habitat and distribution.
The species occurs solitarily in a Q. serrata forest within the core zone of Tianma National Nature Reserve, China (ca. 1100–1300 m, subtropical monsoon climate), with minimal human disturbance. Specimens were collected at a distance of 42.5 cm from a Q. serrata tree, with a diameter at breast height (DBH: 18.14 cm). Soil temperature was 22.5 °C, soil volumetric water content was 20.5%, soil pH was 5.8, air temperature was 24.2 °C, and relative humidity was 90.1%. Soil temperature, moisture content, and pH were measured at the time of sample collection; air temperature and humidity were monitored over the long term within the sample plot (measurements were taken at 10-minute intervals; these data are mean values calculated over the 3 days preceding and following sample collection).
Additional specimens examined.
China • Anhui Province, Lu’an City, Tianma National Nature Reserve, 31°15'40"N, 115°41'46"E, Elevation: ca. 1193 m. on soil in Q. serrata Thunb. forest, 4 August 2023, Yonglan Tuo FJAU66565. GenBank accession numbers: ITS = PV329848, tef1-α = PP357255.
Notes.
Morphologically, H. luteoalbum has medium-sized basidiocarps and is similar to H. flavidocanum in pileus and stipe color, but it has smaller basidiospores (7.08 × 6.10 μm vs. 9.81 × 9.03 μm) and smaller basidia (36.0–44.0 × 8.0–10.0 μm vs. 45.0–82.0 × 10.0–14.0 μm).
Phylogenetic analysis indicates that H. luteoalbum forms a distinct lineage and forms a sister branch to H. albotomentosum. Based on the above morphological characters and phylogenetic results, H. luteoalbum is easily distinguishable from other species of Hydnum and should be classified in subgenus Pallida.
Hydnum
pallidomarginatum
Taxon classificationFungiCantharellalesHydnaceae
T. Cao & H.S. Yuan, 2021
B988FF2A-54C5-5777-827F-9D98C524667C
Habitat and distribution.
The species grows gregariously in a Q. serrata forest within the core zone of Tianma National Nature Reserve, China (ca. 900–1000 m, subtropical monsoon climate). It is currently documented in Anhui Province, Yunnan Province, Heilongjiang Province, and Liaoning Province in China. The specimen was collected at a distance of 10.6 cm from a Q. serrata tree (DBH: 36.2 cm). Soil temperature was 19.8 °C, soil volumetric water content was 23.2%, soil pH was 6.2, air temperature was 22.0 °C, and relative humidity was 92.5%. Soil temperature, moisture content, and pH were measured at the time of sample collection; air temperature and humidity were monitored over the long term within the sample plot (measurements were taken at 10-minute intervals; these data are mean values calculated over the 3 days preceding and following sample collection).
Macroscopic and microscopic features of H. pallidomarginatum. A, B. Basidiomata; C. Hymenium and subhymenium; D. Basidiospores; E. Pileipellis. Scale bars: 2 cm (A, B); 10 μm (C–E).
Specimens examined.
China • Anhui Province, Lu’an City, Tianma National Nature Reserve, 31°15'14"N, 115°42'10"E, Elevation: ca. 960 m, 11 October 2023, Yonglan Tuo (FJAU66578), GenBank accession numbers: ITS = PV875857, tef1-α = PV889366. • Anhui Province, Lu’an City, Tianma National Nature Reserve, 31°15'09"N, 115°42'13"E, Elevation: ca. 954 m, 12 October 2023, Yonglan Tuo (FJAU66579), GenBank accession numbers: ITS = PV875858, tef1-α = PV889367.
Notes.
H. pallidomarginatum is characterized by its white to yellowish-white pileus, 2–4 sterigmata, and globose to subglobose basidiospores. It was described from Yunnan Province, China. We examined collections from Yunnan and several DNA sequences from China annotated as H. pallidomarginatum in the database. No major differences were found between our specimens and the holotype, except for the color of the pileus. The multi-marker phylogenetic analyses revealed that the two Chinese H. pallidomarginatum specimens form a monophyletic cluster within H. pallidomarginatum.
Key to Species of Hydnum in China
**: **
Discussion
Through a comprehensive analysis integrating morphological and molecular methods, we identified and described three novel species (H. luteoalbum, H. albodentum, and H. albotomentosum) and two newly recorded species from the Dabie Mountains (H. berkeleyanum and H. pallidomarginatum). Morphological observations revealed significant differences between H. luteoalbum, H. albodentum, H. albotomentosum, and other Hydnum species in key taxonomically informative characteristics, such as pileus color, size, and basidiospore dimensions. Phylogenetic analyses further confirmed that these three species form distinct monophyletic clades, indicating pronounced genetic divergence. In contrast, H. berkeleyanum and H. pallidomarginatum align with previous taxonomic findings, including both morphological descriptions and phylogenetic placements (Cao et al. 2021; Su et al. 2024).
H. luteoalbum, H. albodentum, H. albotomentosum, and H. pallidomarginatum are characterized by small to medium-sized basidiomata, with pilei ranging in color from white to orange and stipes that are initially white (turning orange upon treatment), which is consistent with the taxonomic criteria of Hydnum subgenus Pallida (Niskanen et al. 2018). However, basidiospore morphology (spherical to sub-elliptical) and Q values (1.0–1.23) of H. luteoalbum, H. albotomentosum, and H. pallidomarginatum differ significantly from the traditional diagnostic features of subgenus Pallida (ovoid to broadly elliptical basidiospores with Q > 1.25). Based on the latest phylogenetic data (Niskanen et al. 2018; Cao et al. 2021; Qin et al. 2024; Su et al. 2024; Zhang et al. 2024) incorporating newly generated sequencing data from this study, these three species are recovered in three strongly supported monophyletic clades within subgenus Pallida. This discrepancy between morphological traits and phylogenetic relationships indicates that the taxonomic diagnostic criteria for the subgenus may require re-evaluation. By integrating phylogenetic evidence with traditional morphological traits, such as basidioma morphology and spore parameters, we propose expanding the defined range of basidiospore Q values within subgenus Pallida.
Global Hydnum species exhibit significant regional distribution patterns: approximately 60% of known species, or 32 species, are concentrated in temperate regions such as Europe and North America, with only a few distributed in subtropical areas (Feng et al. 2016; Niskanen et al. 2018). Compared with the three early recorded species in China, recent studies have remarkably expanded the species diversity of this genus – currently, 32 Hydnum species have been reported in China, over half of which were newly discovered in subtropical regions such as Sichuan Province, Hunan Province, Hubei Province, and Anhui Province (Cao et al. 2021; Qin et al. 2024; Su et al. 2024; Zhang et al. 2024). This indicates that subtropical regions in China are also critical habitats for Hydnum species.
Additionally, as an ectomycorrhizal fungus, Hydnum forms stable symbiotic relationships with plants in the Pinaceae (Agerer et al. 1996) and Fagaceae (McNabb 1971; Feng et al. 2016; Niskanen et al. 2018) families, and its distribution is closely linked to that of its host plants (Niskanen et al. 2018; Swenie et al. 2018; Feng et al. 2016). Detailed recording of host species and other relevant information is of great significance for understanding the ecological distribution characteristics of species and habitat conservation (Durkin et al. 2020). The five species (H. crassipedum, H. albomarginatum, H. fulvostriatum, H. bifurcatum, and H. orientalbidum) we previously described (Tuo et al. 2025), together with the five species identified here, are all distributed in Quercus forests or Quercus-dominated forests. A comprehensive analysis of Hydnum species in China further indicates that nearly all species grow in Pinus–Quercus mixed forests or Quercus-dominated broad-leaved forests (Appendix 1). This suggests a potential direct ecological association between Hydnum distribution and Pinaceae/Fagaceae – previous ectomycorrhizal tip sequencing studies have directly confirmed the ectomycorrhizal association between Hydnum and these plant families (Agerer et al. 1996; Tanaka and Nara 2009; Gao et al. 2013). From a regional distribution perspective, Hydnum species in China exhibit a pattern of “subtropical dominance with temperate supplementation”: 20 species have been recorded in subtropical regions, while only four species are found in temperate regions. This distribution pattern is highly consistent with the geographical distributions of Quercus (Wang et al. 2022) and Pinus (Yue et al. 2025) in China. This indicates that subtropical and temperate regions – particularly the distribution ranges of Quercus and Pinus – may harbor rich Hydnum species resources, and strengthening the protection of habitats dominated by Quercus and Pinus should be an important practical direction for maintaining Hydnum species diversity.
Phenology is another key factor influencing fungal occurrence (Andrew et al. 2018; Krah et al. 2023). Although we conducted long-term macrofungal specimen sampling in the Dabie Mountains region from 2019 to 2024 (two to three times per week during April–October each year), Hydnum specimens were almost exclusively observed in August. Further analysis of the phenology of Hydnum species in China reveals that their peak occurrence mainly occurs from mid-August to mid-September (Appendix 1). This phenological characteristic is likely closely related to the precipitation and temperature conditions in this period of China – characterized by abundant rainfall and high temperatures (Zhan et al. 2019; Deng et al. 2022) – as suitable temperature and humidity conditions are conducive to spore germination (Setlow 2003) and mycelial growth (Querejeta et al. 2009). Ectomycorrhizal fungi act as symbiotic partners with plant roots, extracting water and inorganic nutrients from the soil through mycorrhizal hyphae and transferring them to the roots of the trees they colonize (Lehto and Zwiazek 2011). In return, the fungi receive carbohydrates from the host plants (Courty et al. 2010). The timing of carbohydrate uptake could potentially influence the phenology of the fruiting bodies (Sato et al. 2012). Consequently, ectomycorrhizal (EM) fungi are favored for fruiting body production during the host’s active photosynthetic period, typically in the summer (Sato et al. 2012).
Supplementary Material
XML Treatment for Hydnum albodentum
XML Treatment for Hydnum albotomentosum
XML Treatment for Hydnum berkeleyanum
XML Treatment for Hydnum luteoalbum
XML Treatment for Hydnum pallidomarginatum
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Agerer R, Kraigher H, Javornik B (1996) Identification of ectomycorrhizae of Hydnum rufescens on Norway spruce and the variability of the ITS region of H. rufescens and H. repandum (Basidiomycetes). Nova Hedwigia 63: 183–194. 10.1127/nova.hedwigia/63/1996/183 · doi ↗
- 2Andrew C, Heegaard E, Høiland K, Senn-Irlet B, Kuyper TW, Krisai-Greilhuber I, Kirk PM, Heilmann-Clausen J, Gange AC, Egli S, Bässler C, Büntgen U, Boddy L, Kauserud H (2018) Explaining European fungal fruiting phenology with climate variability. Ecology 99: 1306–1315. 10.1002/ecy.223729655179 · doi ↗ · pubmed ↗
- 3Baird R, Wallace LE, Baker G, Scruggs M (2013) Stipitate hydnoid fungi of the temperate southeastern United States. Fungal Diversity 62: 41–114. 10.1007/s 13225-013-0261-6 · doi ↗
- 4Banker HJ (1901) A preliminary contribution to a knowledge of the Hydnaceae. Bulletin of the Torrey Botanical Club 28: 199–222. 10.2307/2478788 · doi ↗
- 5Banker HJ (1906) A contribution to a revision of the North American Hydnaceae. Memoirs of the Torrey Botanical Club 12: 99–194. 10.5962/bhl.title.97394 · doi ↗
- 6Cao T, Hu YP, Yu JY, Wei TZ, Yuan HS (2021) A phylogenetic overview of the Hydnaceae (Cantharellales, Basidiomycota) with new taxa from China. Studies in Mycology 99: 100121. 10.1016/j.simyco.2021.100121 PMC 871757535035603 · doi ↗ · pubmed ↗
- 7Courty PE, Buée M, Diedhiou AG, Frey-Klett P, Le Tacon F, Rineau F, Turpault MP, Uroz S, Garbaye J (2010) The role of ectomycorrhizal communities in forest ecosystem processes: New perspectives and emerging concepts. Soil Biology & Biochemistry 42: 679–698. 10.1016/j.soilbio.2009.12.006 · doi ↗
- 8Deng HJ, Pepin N, Chen YN, Guo B, Zhang SH, Zhang YQ, Chen XW, Gao L, Liu MB, Chen Y (2022) Dynamics of diurnal precipitation differences and their spatial variations in China. Journal of Applied Meteorology and Climatology 61: 1015–1027. 10.1175/JAMC-D-21-0232.1 · doi ↗