Comprehensive capsular, lipopolysaccharide, virulence, and antimicrobial resistance profiling of Pasteurella multocida isolated from buffaloes in Vietnam: First report of capsular type D and predominant L2 genotype
Thai Van Nguyen, T. T Hang Trinh, Trong Van Nguyen, Dinh Ng–Nguyen, Hieu Quoc Nguyen, Hung Vu–Khac

TL;DR
This study provides the first detailed molecular profile of Pasteurella multocida in Vietnamese buffaloes, identifying new capsular types and resistance genes.
Contribution
First report of capsular type D in Vietnamese buffalo P. multocida and comprehensive profiling of LPS, virulence, and resistance genes.
Findings
Capsular type D was identified for the first time in Vietnamese buffalo P. multocida isolates.
L2 was the most prevalent LPS genotype among the isolates.
Four antimicrobial resistance genes were detected, including floR and tetB.
Abstract
Pasteurella multocida is a primary cause of hemorrhagic septicemia (HS) in buffaloes across tropical regions, leading to rapid deaths and significant economic losses. In Vietnam, recurrent HS outbreaks have been reported, yet most studies have solely focused on bacterial isolation or species-level identification. Comprehensive molecular data, including capsular typing, lipopolysaccharide (LPS) genotyping, virulence–associated genes (VAGs), and antimicrobial resistance genes (ARGs), remain limited for buffalo-derived strains. This molecular gap hampers understanding of strain diversity, epidemiology, and vaccine development. Notably, no previous Vietnamese study has concurrently characterized capsule, LPS, VAG, and ARG profiles or reported atypical serogroups in buffaloes. Therefore, integrated molecular surveillance is crucial to detect emerging lineages and guide One Health–oriented…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1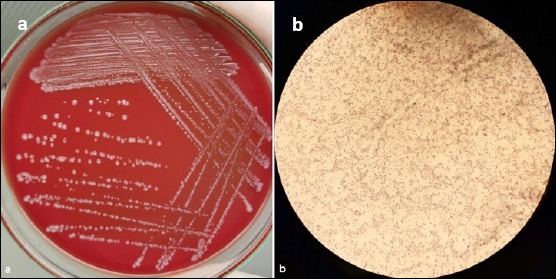 Figure 2
Figure 2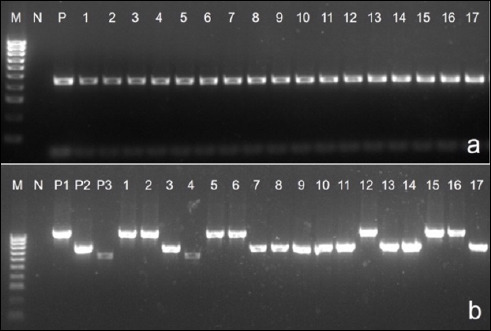 Figure 3
Figure 3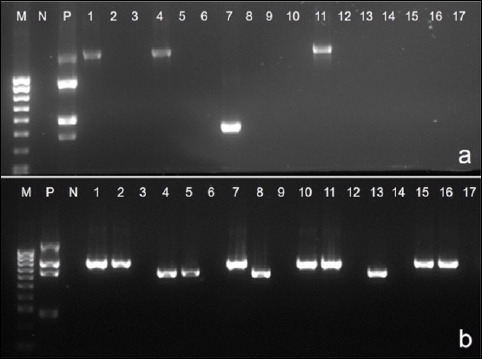 Figure 4
Figure 4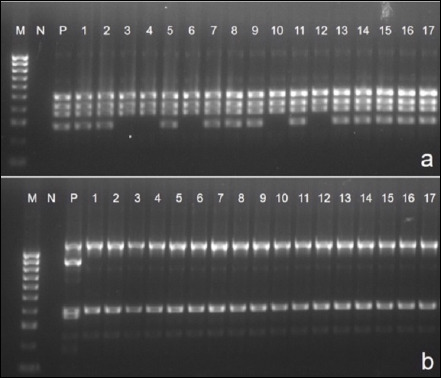 Figure 5
Figure 5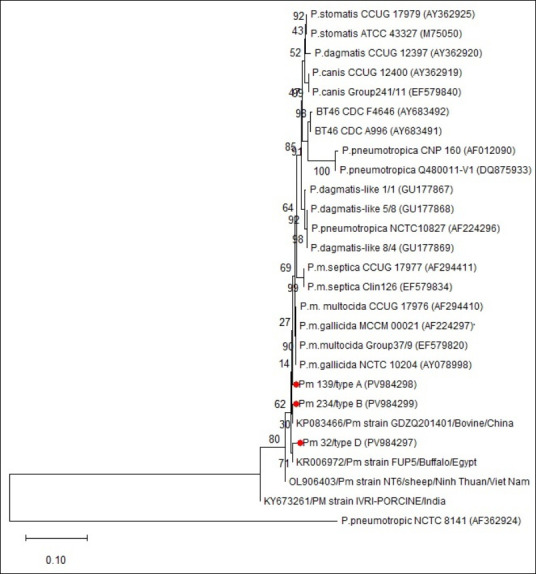 Figure 6
Figure 6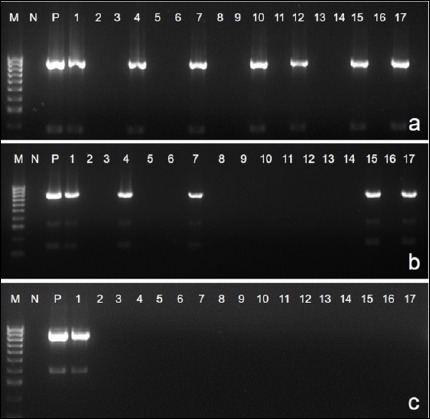 Figure 7
Figure 7| Gene | Function | Primer sequences (5’–3’) | Amplicon size (bp) | References |
|---|---|---|---|---|
|
| Species–specific marker for | F: ATCCGCTATTTACCCAGTGG | 460 | [ |
|
| Serogroup A cap | F: GATGCCAAAATCGCAGTCAG | 1044 | [ |
|
| Serogroup B cap | F: CATTTATCCAAGCTCCACC | 760 | [ |
|
| Serogroup D cap | F: TTACAAAAGAAAGACTAGGAGCCC | 657 | [ |
|
| LPS genotype L1 | F: ACATTCCAGATAATACACCCG | 1307 | [ |
|
| LPS genotype L2 | F: CTTAAAGTAACACTCGCTATTGC | 810 | [ |
|
| LPS genotype L3 | F: TGCAGGCGAGAGTTGATAAACCATC | 474 | [ |
|
| LPS genotype L4 | F: TTTCCATAGATTAGCAATGCCG | 550 | [ |
|
| LPS genotype L5 | F: AGATTGCATGGCGAAATGGC | 1175 | [ |
|
| LPS genotype L6 | F: TCTTTATAATTATACTCTCCCAAGG | 668 | [ |
|
| LPS genotype L7 | F: CCTATATTTATATCTCCTCCCC | 931 | [ |
|
| LPS genotype L8 | F: GAGAGTTACAAAAATGATCGGC | 255 | [ |
|
| Type IV fimbria (adhesion) | F: TGTGGAATTCAGCATTTTAGTGTGTC | 488 | [ |
|
| Filamentous hemagglutinin (adhesion) | F: AGCTGATCAAGTGGTGAAC | 275 | [ |
|
| Sialidase | F: GTCCTATAAAGTGACGCCGA | 554 | [ |
|
| Sialidase | F: GAATATTTGGGCGGCAACA | 360 | [ |
|
| Porin (protectin) | F: CGCGTATGAAGGTTTAGGT | 438 | [ |
|
| Porin (protectin) | F: ATGAAAAAACTTTTAATTGCGAGC | 948 | [ |
|
| Iron metabolism | F: GGTGGTGATATTGATGCGGC | 1144 | [ |
|
| Hemoglobin-binding protein (iron uptake) | F: TGGCGGATAGTCATCAAG | 419 | [ |
|
| Hemoglobin-binding protein (iron uptake) | F: ACCGCGTTGGAATTATGATTG | 788 | [ |
|
| Dermonecrotic toxin | F: CTTAGATGAGCGACAAGGTT | 865 | [ |
|
| Superoxide dismutate | F: TACCAGAATTAGGCTACGC | 361 | [ |
|
| Superoxide dismutate | F: AGTTAGTAGCGGGGTTGGCA | 235 | [ |
|
| Universal bacterial marker | F: AGAGTTTGATYMTGGC | 1502 | [ |
| Antibiotic class | Gene | Primer sequences (5’–3’) | Amplicon size (bp) | References |
|---|---|---|---|---|
| β-lactam |
| F: GAGTATTCAACATTTTCGT | 852 | [ |
|
| F: CATTAACGGCTTGTTCGC | 856 | [ | |
| Tetracycline |
| F: CCTTATCATGCCAGTCTTGC | 774 | [ |
|
| F: ATACTGCTGATCACCGT | 1076 | [ | |
|
| F: TAACTTAGGCATTCTGGCTC | 1801 | [ | |
| Chloramphenicol |
| F: CACGTTGAGCCTCTATATGG | 885 | [ |
| Aminoglycoside (Gentamycin) |
| F: CTCGAATGCCTGGCGTGTTT | 482 | [ |
| Gene | No. of positive isolates | Percentage | 95% CI |
|---|---|---|---|
|
| 21 | 31.3a | 20.5–43.8 |
|
| 42 | 62.7b | 50.1–74.2 |
|
| 4 | 5.9c | 1.7–14.5 |
| Gene | No. of positive isolates | Percentage (%) | 95% CI |
|---|---|---|---|
|
| 11 | 16.4a | 8.4-27.5 |
|
| 38 | 56.7b | 44.1-68.7 |
|
| 5 | 7.5a | 2.4-16.5 |
|
| 0 | – | – |
|
| 0 | – | – |
|
| 13 | 19.4a | 10.7-30.8 |
|
| 0 | – | – |
|
| 0 | – | – |
| Gene | No. of positive isolates | Percentage (%) | 95% CI |
|---|---|---|---|
|
| 67 | 100a | 94.6-100 |
|
| 39 | 58.2b | 45.5-70.1 |
|
| 67 | 100a | 94.6-100 |
|
| 0 | - | - |
|
| 67 | 100a | 94.6-100 |
|
| 67 | 100a | 94.6-100 |
|
| 67 | 100a | 94.6-100 |
|
| 67 | 100a | 94.6-100 |
|
| 23 | 34.3c | 23.1-46.9 |
|
| 0 | - | - |
|
| 67 | 100a | 94.6-100 |
|
| 67 | 100a | 94.6-100 |
| Gene | No. of positive isolates | Percentage (%) | 95% CI |
|---|---|---|---|
|
| 0 | - | - |
|
| 7 | 10.4ab | 4.3-20.3 |
|
| 8 | 11.9ab | 5.2-22.1 |
|
| 3 | 4.4a | 0.9-12.5 |
|
| 0 | - | - |
|
| 15 | 22.3b | 13.1-34.2 |
|
| 0 | - | - |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial infections and disease research · Rabies epidemiology and control · Animal health and immunology
INTRODUCTION
Pasteurella multocida is a Gram–negative bacterium responsible for a wide range of diseases in animals and humans, from mild infections to severe, often fatal conditions. These include hemorrhagic septicemia (HS) in cattle and buffaloes, pneumonic and septicemic pasteurellosis in sheep and goats, fowl cholera in poultry, and various infections in companion animals such as cats and dogs [1–4]. Among these diseases, HS is especially devastating in tropical regions, causing high morbidity and mortality. In Vietnam, recurrent HS outbreaks have been documented in both cattle and buffaloes, with buffaloes showing greater susceptibility and higher death rates. This highlights the need for focused molecular surveillance and targeted disease control strategies in this species.
Capsular typing, particularly the identification of serogroup B, remains essential for determining P. multocida strains involved in HS outbreaks across Asia [5–7]. Lipopolysaccharide (LPS) genotyping adds an extra layer of strain differentiation, as LPS types (L1–L8) display host- and disease-specific patterns; however, data on buffalo-derived strains, especially in Vietnam, are limited [8]. Virulence–associated genes (VAGs), which encode factors related to adhesion, iron acquisition, outer membrane stability, and enzymatic activity, further influence the pathogenic profile of P. multocida [1, 9]. Although the distribution of these genes varies among host species and regions, few studies have investigated VAGs patterns in Vietnamese buffalo isolates.
Antimicrobial resistance genes (ARGs) have been detected in P. multocida strains from pigs in Vietnam [10], but similar studies on buffalo-derived isolates are lacking. Although P. multocida has been isolated from various domestic animals, including cattle, buffaloes, pigs, poultry, goats, and sheep [11–13], most previous Vietnamese research focused only on species-level identification. Few studies have investigated capsular types, and almost none have assessed LPS genotypes or virulence profiles in buffalo strains. Consequently, essential molecular epidemiological data are still missing. Recent research across Asia emphasizes the importance of combining capsular and LPS genotyping to monitor emerging P. multocida lineages [14, 15]; however, similar molecular insights for Vietnamese buffalo populations are still unavailable.
Despite the frequent occurrence of HS in Vietnam and the recognized role of P. multocida as a major pathogen in buffaloes, there is still a significant lack of molecular epidemiological data on the circulating strains in this host. Previous Vietnamese studies have mainly focused on bacterial isolation and species confirmation, with limited work on capsular typing and almost no research combining LPS genotyping, virulence–associated gene (VAG) profiles, and ARG patterns in buffalo-derived isolates. Consequently, critical information about strain diversity, pathogenic potential, and the molecular factors that influence disease severity in buffalo populations is missing. Importantly, buffaloes in the Central Highlands are raised within multispecies livestock systems, often near cattle, pigs, poultry, and dogs, which heightens the risk of cross-species transmission, the emergence of unusual capsular or LPS types, and the spread of ARG. While recent Asian studies highlight the importance of combined capsular–LPS genotyping for tracking new P. multocida lineages, comparable integrated data for Vietnamese buffaloes are still lacking. This significant gap hampers the development of targeted vaccines, accurate diagnostic tools, and effective regional control strategies.
To address these gaps, this study aimed to perform the first comprehensive molecular characterization of P. multocida isolated from buffaloes in Vietnam’s Central Highlands. Using a PCR-based multilocus approach, we examined capsular types, LPS genotypes, and VAG profiles, along with screening for key antimicrobial resistance determinants. Additionally, representative isolates underwent 16S rRNA sequencing to confirm species identity and determine phylogenetic relationships with global reference strains. By integrating capsule, LPS, virulence, and resistance markers, this research offers a comprehensive molecular epidemiological assessment of P. multocida in Vietnamese buffalo populations. The data generated aims to improve diagnostic accuracy, guide vaccine antigen selection based on regional strain patterns, strengthen antimicrobial stewardship, and support future One Health surveillance efforts in mixed-species farming settings.
MATERIALS AND METHODS
Ethical approval
This study did not involve any experimental procedures on live animals. All samples used in this research were collected from buffaloes that were either clinically affected by HS or had died naturally and were submitted to provincial veterinary authorities for routine diagnostic investigation. No animals were handled, restrained, or sampled for research purposes. Accordingly, the work did not require a separate Institutional Animal Care and Use Committee approval under the ethical policies of Tay Nguyen University and the Institute of Veterinary Research and Development of Central Vietnam.
Sampling and laboratory procedures complied with the national regulations of Vietnam on animal welfare, biosafety, and disease surveillance, as well as guidelines outlined by the World Organization for Animal Health (OIE/WOAH) for the humane use of animals in research and diagnosis. All submitted samples were processed anonymously, and no identifying information related to farms or owners was disclosed.
Owner consent was obtained at the time of sample submission for diagnostic purposes, including permission to use leftover clinical materials for research aimed at improving disease control in the region. All laboratory analyses, bacterial isolation, polymerase chain reaction (PCR) assays, virulence and resistance gene screening, and sequencing, were performed on stored isolates without requiring any interaction with live animals. Thus, the study fully adhered to the principles of Replacement, Reduction, and Refinement (3Rs) in animal research.
Study period and location
The study was conducted from April 2022 to May 2025. All samples were collected from buffaloes raised in the Dak Lak, Gia Lai, Kon Tum, and Lam Dong provinces of the Central Highlands in Vietnam. Sampling took place over a three-year span (Figure 1).
Map of Vietnam showing the study location.
Isolation and identification of P. multocida strains
A total of 67 P. multocida isolates were obtained from the lungs, bone marrow, and nasal swabs of buffaloes showing clinical signs of pasteurellosis, such as fever, edema of the mandible, neck, and brisket, nasal discharge, respiratory distress, and loss of appetite. All isolates were cultured on 5% sheep blood agar (Oxoid, UK) and incubated at 37°C for 24 h. Initial identification was based on characteristic morphology (smooth, grayish-white, non-hemolytic colonies), Gram staining, absence of growth on MacConkey agar (HiMedia, India), and standard biochemical profiles (positive catalase and oxidase tests; fermentation of glucose and mannitol). Species confirmation was carried out using a P. multocida–specific PCR assay with ATCC 12945 (Microbiologics, USA) as the positive-control, following the previously described method [16].
DNA extraction
Confirmed isolates were cultured in brain–heart infusion broth (Oxoid, UK) at 37°C for 24 h. A 1 mL aliquot of the overnight culture was processed for DNA extraction using the QIAamp DNA Mini Kit (Qiagen, Germany) following the manufacturer’s instructions. Extracted DNA was stored at –20°C until analysis.
Capsular typing
Capsular types were identified using multiplex PCR targeting the capA, capB, and capD loci [17, 18]. Each 20 μL reaction contained 2 μL of DNA template, 10 μL of 2× MyTaq Mix (Bioline, USA), 0.5 μL of each primer (25 μM; Phusa Genomics, Vietnam), and nuclease-free water. The thermal cycle included 94°C for 4 min; 35 cycles of 94°C for 45 s, 55°C for 45 s, and 72°C for 45 s; followed by a final extension at 72°C for 10 min. Positive-control DNA provided by the Institute of Veterinary Research and Development of Central Vietnam (IVRD) was included in each run.
LPS genotyping
LPS genotypes were identified using two multiplex PCR sets targeting eight LPS-related genes, following a previously described method with minor modifications [8]. In the original protocol, eight LPS-specific genes were amplified in a single multiplex reaction; in the present study, these eight targets were divided into two separate four-plex multiplex PCR assays:
- Multiplex set 1: pcgD–pcgB, gatF, latB, ppgB
- Multiplex set 2: nctA, rmlA–rmlC, nctB, natG
Each 50 μL reaction included 25 μL of MyTaq Mix, 0.5 μL of each primer (25 μM), 5 μL of DNA template, and 18 μL of nuclease-free water. Cycling conditions consisted of 96°C for 5 min; 30 cycles of 96°C for 30 s, 56°C for 30 s, and 72°C for 2.5 min; and a final extension at 72°C for 8 min. IVRD positive-control DNA was used to verify assay performance.
Detection of VAGs
Twelve VAGs related to adhesion, iron acquisition, outer membrane integrity, and enzymatic activity were screened using multiplex PCR, following protocols adapted from previous studies [19–21].
- Multiplex set 1: sodA, hgbA, ptfA, pfhA
- Multiplex set 2: ompH, tonB, nanH, toxA
- Multiplex set 3: oma87, sodC, hgbB, nanB
Each 25 μL reaction contained 2 μL DNA template, 12.5 μL 2× MyTaq Mix, 0.5 μL of each primer (10 μM), and 6.5 μL nuclease-free water. Amplification conditions were: 96°C for 5 min; 30 cycles of 96°C for 60 s, 56°C for 60 s, and 72°C for 60 s; and a final extension at 72°C for 10 min. IVRD positive-control DNA was used for quality assurance. Primer sequences for all assays are listed in Table 1 [8, 16, 17, 20, 21, 22].
Detection of ARGs
ARGs were screened using simplex PCR with primers listed in Table 2 [23–25]. Each 25 μL reaction included 2 μL of DNA template, 12.5 μL of 2× MyTaq Mix, 0.5 μL of each primer (10 μM), and 9.5 μL of water. Cycling conditions were the same as those used for 16S rRNA PCR, except for tetO, where the extension step was increased to 1 min [10]. Positive-control DNA from IVRD was included in all runs.
16S rRNA gene sequencing and phylogenetic analysis
Three isolates representing capsular types A, B, and D and harboring multiple VAGs were selected for 16S rRNA sequencing using universal primers [22]. PCR reactions (25 μL) contained 2 μL of DNA template, 0.5 μL of each primer (10 μM), 12.5 μL of 2× MyTaq Mix, and 9.5 μL of water. Conditions were: 94°C for 5 min; 30 cycles at 94°C for 45 s, 48°C for 45 s, and 72°C for 90 s; and a final extension at 72°C for 8 min.
Amplicons were sequenced through Sanger sequencing (1st BASE, Singapore). Sequences were aligned in BioEdit (version 5.0.9) using the ClustalW algorithm. Phylogenetic trees were generated in MEGA version 6.06 (Molecular Evolutionary Genetics Analysis; www.megasoftware.net) with the neighbor-joining method, performing 1,000 bootstrap replicates; bootstrap values ≥70% indicated strong support. Rooting was carried out using P. pneumotropica NCTC 8141 (AF362924).
All PCR assays were conducted in duplicate on a C1000 Touch thermal cycler (Bio-Rad, USA). PCR products were separated on 1% TBE agarose gels at 130 V for 60 min, stained with ethidium bromide, and visualized under UV light using a 100-bp DNA ladder (Bioline, USA).
Statistical analysis
Prevalence was calculated as the proportion of positive isolates among total samples and expressed as percentages with 95% confidence intervals (R version 4.4.3, R Core Team, Vienna, Austria). Differences between proportions were assessed using chi-square tests, with p < 0.05 considered statistically significant. QGIS version 3.10.0 was used to generate the study site map [26].
RESULTS
Identification and confirmation of P. multocida
Biochemical characterization confirmed that all 67 isolates showed typical features of P. multocida, including Gram–negative coccobacilli morphology (Figures 2a and 2b), lack of growth on MacConkey agar, non-hemolytic colonies on blood agar, and positive catalase and oxidase reactions. Glucose and sucrose fermentation were observed, while lactose was not fermented. Species–specific PCR further verified that all isolates belonged to P. multocida (Figure 3a).
Morphological characteristics of Pasteurella multocida isolates. (a) Colonies of P. multocida after 24 h incubation at 37oC on 5% sheep blood agar showing smooth, glistening, non–hemolytic morphology. (b) Gram–strained smear demonstrating small Gram–negative coccibacilli observed under oil immersion (100×).
Species–specific Polymerase chain reaction (PCR) and multiplex PCR for capsular typing of Pasteurella multocida. (a) P. multocida confirmation by species–specific PCR. 1-17: representative samples; N: negative control; P: positive-control (460 bp); M: 100 bp DNA ladder. (b) Multiplex reaction of capA, capB, capD. 1-17: representative samples; P1: capA positive-control (1044 bp); P2: capB positive-control (760 bp); P3: capD positive-control (657 bp); M: 100 bp DNA ladder.
Distribution of capsular biosynthesis genes
Multiplex PCR analysis revealed the following capsular gene distribution (Table 3; Figure 3b):
- capA: 21 isolates (31.3%)
- capB: 42 isolates (62.7%)
- capD: 4 isolates (5.9%)
Thus, capsular type B predominated among buffalo-derived isolates, followed by types A and D.
LPS genotype distribution
Four LPS genotypes were detected among the 67 isolates (Table 4; Figures 4a and 4b):
Polymerase chain reaction (PCR) for lipopolysaccharide genotypes of isolated Pasteurella multocida strains. 1-17: samples, N: negative control, P: positive-control, M: 100 bp DNA ladder. (a) PCR product sizes: L1 = 1307 bp; L3 = 474 bp. (b) PCR product sizes: L2 = 810 bp; L6 = 668 bp.
- L2: 38 isolates (56.7%) - most prevalent)
- L6: 13 isolates (19.4%)
- L1: 11 isolates (16.4%)
- L3: 5 isolates (7.5%)
Genotypes L4, L5, L7, and L8 were not detected in any isolate.
VAGs
All isolates carried eight conserved VAGs: ptfA, nanB, ompH, oma87, exbB–tonB, hgbA, sodA, and sodC. Variable gene frequencies included:
- pfhA: 58.2%
- hgbB: 34.3%
The nanH and toxA genes were completely absent across all isolates. Representative PCR amplifications are shown in Figures 5a and 5b, with detailed prevalence in Table 5.
Multiplex polymerase chain reaction (PCR) detection of virulence–associated genes in Pasteurella multocida isolates. M: 100 bp DNA ladder, N: negative control, P: positive-control, 1-17: samples. (a) PCR product sizes: ptfA = 488 bp; hgbA = 419 bp; sodA = 361 bp; pfhA = 275 bp. (b) PCR product sizes: tonB = 1144 bp; ompH = 438 bp.
Phylogenetic analysis of 16S rRNA sequences
Phylogenetic analysis showed that isolates Pm 139 (type A), Pm 234 (type B), and Pm 32 (type D) clearly grouped within the P. multocida species, confirming their identity (Figure 6).
Neighbor-joining dendrogram illustrating the phylogenetic relationships of 16S rRNA gene sequences of Pasteurella multocida isolates.
- Pm 139/type A and Pm 234/type B grouped closely with P. multocida reference strains (e.g., NCTC 10204), indicating high genetic similarity.
- Pm 32/type D formed a distinct but related sub-branch, suggesting minor genetic divergence while remaining within the species cluster.
All three isolates were distinctly separated from other Pasteurella species (Pasteurella stomatis, Pasteurella dagmatis, Pasteurella canis, and Pasteurella pneumotropica). International reference strains from China, Egypt, India, and Vietnam also grouped within the P. multocida cluster, showing conservation of 16S rRNA sequences across different geographic regions.
The neighbor-joining tree was built using 1,000 bootstrap replications, with bootstrap values of 70% or higher considered strong support P. pneumotropica NCTC 8141 (AF362924) served as the outgroup.
Occurrence of ARGs
ARG detection revealed variable resistance gene prevalence (Table 6 and Figures 7a–c):
Polymerase chain reaction assays for detecting the prevalence of (a) floR, (b) tetB, and (c) blaROB1.
- floR: 22.3% (15/67) - highest prevalence
- tetB: 11.9% (8/67)
- blaROB1: 10.4% (7/67)
- tetH: 4.4% (3/67)
The blaTEM, tetO, and aacA4 genes were not detected in any isolate.
DISCUSSION
Overall significance of the study
This investigation offers the first in-depth molecular characterization of P. multocida isolates from buffaloes in the Central Highlands of Vietnam. Previous studies in Vietnam mainly focused on species-level identification, leaving significant gaps in knowledge about capsular types, LPS genotypes, virulence–associated genes (VAGs), and ARGs. By incorporating these molecular markers, the current study provides new epidemiological insights and creates a baseline dataset for regional surveillance and disease management.
Capsular type distribution and epidemiological implications
Capsular type B was the most common serogroup identified, aligning with reports from other Asian countries where serogroup B is strongly linked to HS in cattle and buffaloes [7, 14, 27]. Capsular type A was also found, consistent with findings from different geographic regions [27–30].
Importantly, the detection of capsular type D (5.9%; 4/67) in Vietnamese buffaloes is a new and epidemiologically important discovery, as this serogroup has not been previously reported in buffalo-derived isolates in Vietnam. Although uncommon, type D has been found in buffaloes in Iran (1.8%) [27] and India (4.3%) [29], indicating its sporadic yet widespread presence across Asia.
The appearance of type D in this study may be due to pathogen spillover within the multispecies farming systems typical of the Central Highlands, where buffaloes, cattle, pigs, poultry, and dogs are raised closely together [31]. Such interfaces promote interspecies transmission, genetic exchange, and the introduction of unusual serogroups. This finding underscores One Health concerns regarding the circulation of various P. multocida lineages and emphasizes the need for expanded molecular surveillance.
The dominance of capsular type B and its alignment with region-specific patterns highlight its significance as a target for better vaccine development in Vietnam.
LPS genotype variation and ecological drivers
LPS genotype L2 was the most common, followed by L6, L1, and L3. This distribution is different from patterns typically seen in cattle, pigs, and poultry, where L3 and L6 are mostly found [32–34]. For example:
- L6 predominates in pig-derived isolates in China [33].
- L3 is common in bovine respiratory pasteurellosis [32].
The prominence of L2 in buffaloes may reflect host-specific ecology, antibiotic use, and local husbandry practices. In the Central Highlands, buffaloes often graze communally under extensive or semi-intensive systems, which promotes the transmission of hemorrhagic septicemia–associated strains. Seasonal humidity and flooding may also help P. multocida persist in the environment. These ecological factors probably contribute to the co-occurrence of capsular type B and L2, the dominant profile observed in this study.
Virulence gene profiles and their biological relevance
All isolates carried conserved VAGs responsible for adhesion (ptfA), iron acquisition (exbB–tonB and hgbA), and outer membrane integrity (omph and oma87). The universal presence of ptfA and nanB is consistent with their known importance in colonization and mucosal invasion, mirroring reports from India [29] and Iran [35].
The moderate prevalence of pfhA (58.2%) and hgbB (34.3%) aligns with previous findings that the distribution of virulence genes varies by capsular type. Serogroup B isolates, which were predominant here, are historically linked to extensive adhesion and iron uptake capabilities [30]. The absence of toxA and nanH (0%) aligns with the host specificity of these toxin-related genes, which are more commonly associated with disease syndromes in pigs and poultry than in ruminants [21, 29].
These patterns support the idea that capsule genotype influences virulence gene composition and reflect a virulence profile adapted to buffalo-related HS.
ARG patterns and One Health context
Among the seven ARGs screened, four, blaROB1, tetB, tetH, and floR, were detected at different frequencies, while blaTEM, tetO, and aacA4 were not present. This is the first report of ARG profiles in buffalo-derived P. multocida isolates from Vietnam.
The overall ARG prevalence was significantly lower than that observed in Vietnamese pig isolates, in which all seven genes have been documented [10]. This difference probably stems from species–specific antimicrobial exposure and treatment strategies, highlighting the need for host-targeted antimicrobial stewardship.
International comparisons reveal substantial variation. For instance:
- Multidrug-resistant isolates in Egypt commonly carry tetH and β-lactamase genes [36].
- In Germany, floR was found in only 12.5% [37], which is less than the 22.3% observed in this study.
In Vietnam’s mixed-farming systems, close animal proximity heightens the risk of horizontal ARG gene transfer between species. Detection of tetB, tetH, and blaROB1 matches the common field use of tetracyclines and β-lactams, while floR aligns with routine florfenicol administration [10, 38]. These findings highlight the importance of integrated molecular surveillance and careful antimicrobial use across livestock sectors within a One Health approach.
Phylogenetic insights and evolutionary considerations
Phylogenetic analysis confirmed that all isolates clustered closely with global P. multocida reference strains from cattle, buffaloes, pigs, and sheep, indicating strong species-level genetic conservation. However, the capsular type D isolate (Pm 32) formed a distinct minor sub-branch, suggesting possible local microevolution within the multispecies farming ecosystems of the Central Highlands. These ecological interfaces may facilitate interspecies transmission, genetic recombination, and the emergence of new sub-lineages. Therefore, ongoing genomic surveillance, including MLST and whole–genome sequencing, is crucial to monitor evolutionary trends and transmission pathways.
CONCLUSION
This study offers the first comprehensive molecular characterization of P. multocida isolated from buffaloes in the Central Highlands of Vietnam, providing important insights into the pathogen’s capsular composition, LPS diversity, virulence factors, antimicrobial resistance patterns, and phylogenetic placement. Among the 67 isolates examined, capsular type B was most common (62.7%), followed by type A (31.3%). Notably, capsular type D (5.9%) was identified for the first time in Vietnamese buffaloes, an epidemiologically important discovery indicating possible spillover within multispecies farming systems. LPS genotyping showed L2 as the dominant genotype (56.7%), with all isolates carrying eight conserved virulence genes (ptfA, nanB, ompH, oma87, exbB–tonB, hgbA, sodA, and sodC), and variable presence of pfhA (58.2%) and hgbB (34.3%). Four antimicrobial resistance genes, floR, tetB, blaROB1, and tetH, were detected, marking the first report of ARGs in buffalo-derived P. multocida in Vietnam.
The predominance of capsular type B and L2 LPS genotype has direct implications for HS vaccine development, as including regional dominant antigens may improve immunoprotection. The presence of specific virulence and resistance genes emphasizes the need for cautious antimicrobial use and the development of diagnostic panels tailored to buffalo-associated strains. The detection of capsular type D highlights the importance of monitoring cross-species pathogen transmission in mixed-farming systems.
This work employs multiple molecular methods, capsular typing, LPS genotyping, VAG profiling, ARG screening, and phylogenetic analysis, offering a multilayered epidemiological overview not previously available in Vietnam. The use of control strains, standardized PCR assays, and sequencing enhances the accuracy of the data. However, the study did not include phenotypic antimicrobial susceptibility testing, limiting direct correlation between ARGs and clinical resistance. Additionally, sampling was restricted to four provinces in the Central Highlands, and whole–genome sequencing was not conducted, which limits deeper insights into evolutionary pathways and transmission dynamics. Future research involving MLST and whole–genome sequencing across buffalo, cattle, pigs, and poultry will help clarify strain relationships, transmission routes, and the emergence of new lineages. Incorporating phenotypic resistance testing and environmental sampling could strengthen One Health surveillance strategies.
Overall, this study provides essential baseline molecular data for P. multocida in Vietnamese buffaloes and underscores the need for ongoing surveillance, targeted vaccination, and responsible antimicrobial use. The findings significantly contribute to regional disease management efforts and lay the groundwork for advanced genomic studies in Vietnam’s livestock sector.
DATA AVAILABILITY
The 16S rRNA gene sequences generated in this study have been deposited in GenBank under the accession numbers PV984297, PV984298, and PV984299. Supplementary Table 1. Molecular characteristics (capsular type, LPS genotype, VAGs, and ARGs) of all P. multocida isolates included in this study.
AUTHORS’ CONTRIBUTIONS
TVN: Performed the experiments, analyzed and interpreted the data, and, with support from TVN, TTHT, and HVK: Analyzed and interpreted the data, drafted and revised the manuscript. HVK: Conceived the study, provided project supervision, and critically revised the manuscript. TVN, DNN, and HQN: Collected the samples and contributed to data acquisition and interpretation. All authors have read and approved the final version of the manuscript.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Shivachandra S.B Viswas K.N Kumar A.AA review of hemorrhagic septicemia in cattle and buffalo Anim. Health Res. Rev 201112(1)67822167634110.1017/S 146625231100003 X · doi ↗ · pubmed ↗
- 2Piorunek M Brajer-Luftmann B Walkowiak J Pasteurella multocida infection in humans Pathogens 202312(10)1810.3390/pathogens 12101210 PMC 1061006137887726 · doi ↗ · pubmed ↗
- 3Wei B Liu C Zhu J Zou X.Y Zhang Z Pasteurella multocida infection:a retrospective study of 482 cases of P. multocida infection in patients of different ages BMC Infect. Dis 202525(1)1134003859610.1186/s 12879-025-10711-1PMC 11881477 · doi ↗ · pubmed ↗
- 4Andrés-Lasheras S Jelinski M Zaheer R Mc Allister T.A Bovine respiratory disease:culture-independent approaches to studying antimicrobial resistance Antibiotics 202211(4)22610.3390/antibiotics 11040487 PMC 902527935453238 · doi ↗ · pubmed ↗
- 5Almoheer R Wahid Abd Zakaria M.E Jonet M.A.B Al-shainabi M.M Al-Gheethi A Addis S.N.K Global patterns of hemorrhagic septicemia (2005–2019)Vaccines 202210(2)11710.3390/vaccines 10020315 PMC 888027735214771 · doi ↗ · pubmed ↗
- 6Weerathunga M.W.D.C Ubeyratne J.K.H Nadheer M.A An outbreak of haemorrhagic septicaemia in Navithanveli veterinary range in Ampara District, Sri Lanka Sri Lanka Vet. J 201764(2)712
- 7Biswas A Shivachandra S.B Saxena M.K Kumar A.A Singh V.P Srivastava S.K Molecular variability among strains of Pasteurella multocida isolated from an outbreak of haemorrhagic septicaemia in India Vet. Res. Commun 200428(4)28729810.1023/b:verc.0000026656.77847.5b 15222733 · doi ↗ · pubmed ↗
- 8Harper M John M Turni C Edmunds M St. Michael F Adler B Blackall P.J Cox A.D Boyce J.D Development of a rapid multiplex PCR assay to genotype Pasteurella multocida strains J. Clin. Microbiol 201553(2)4774852542814910.1128/JCM.02824-14PMC 4298526 · doi ↗ · pubmed ↗