First report of Ophiotaenia sp. in frogs (Amietophrynus kassasii) from Egypt and in vitro anticestodal activity of Sinularia sp. extract
Barakat Shehata Abd elmaleck, Mahmoud Abdelhamid, Abdallah Alian, Hind Alzaylaee, George D. Zouganelis, Gaber El-Saber Batiha, Marwa Adel Thabet, Fatma A. S. Anwar

TL;DR
This study reports the first case of a tapeworm in Egyptian frogs and shows that a coral extract can effectively kill the parasite in the lab.
Contribution
The first documentation of Ophiotaenia sp. in Egyptian frogs and evidence of Sinularia sp. extract's anticestodal activity.
Findings
Ophiotaenia sp. was found in 5.9% of frogs with high parasite load and identified using microscopy.
Sinularia sp. extract killed tapeworms in a dose-dependent manner, with faster mortality at higher concentrations.
SEM analysis revealed significant structural damage to tapeworms treated with the extract.
Abstract
Ophiotaenia species are globally distributed proteocephalidean cestodes that commonly parasitize amphibians and reptiles. Despite the ecological importance of frogs in controlling insect populations and maintaining food-web stability, data on cestode infections in Egyptian amphibians remain scarce. This study provides the first documentation of Ophiotaenia sp. infecting Amietophrynus kassasii in Egypt and evaluating the in vitro anticestodal activity of Sinularia sp. extract against adult tapeworms. A total of 85 frogs were collected from freshwater ponds in New Valley Governorate, Egypt, between February and September 2024. Intestines were examined for cestodes, which were identified morphologically using light microscopy and scanning electron microscopy (SEM). Soft coral Sinularia sp. extract was prepared by methanolic extraction, and three concentrations (25, 50, 100 µg/mL) were…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1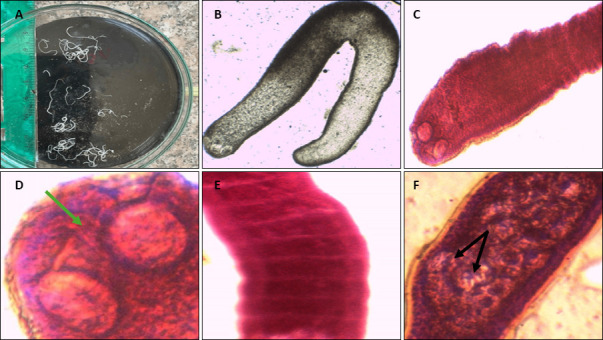 Figure 2
Figure 2 Figure 3
Figure 3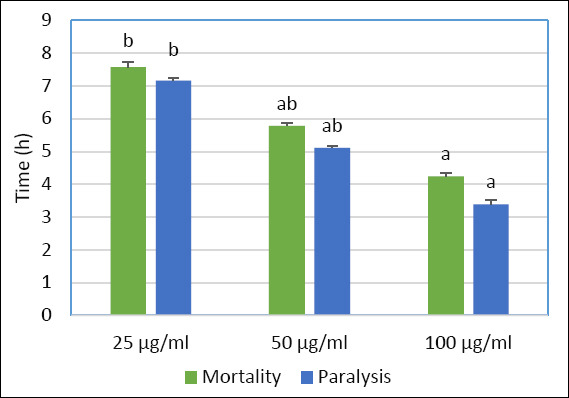 Figure 4
Figure 4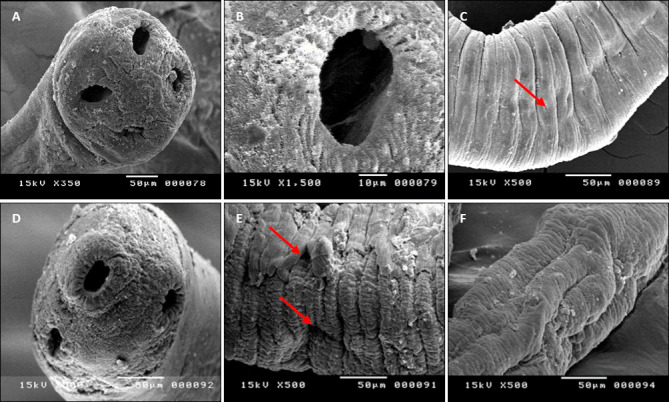 Figure 5
Figure 5| Parasite | Number of infected frogs | Infection rate (95% CI) | Intensity mean (95% CI) | Mean abundance (95% CI) |
|---|---|---|---|---|
| 5 | 5.9 (1.93–13.20) | 70 (35.00–90.00) | 4.12 (1.18–8.82) |
| Species |
|
| |||
|---|---|---|---|---|---|
| Host |
|
|
|
| |
| Region | Michigan, USA | Wisconsin, USA | Wisconsin, Mississipi, USA | Arkansas, USA | New Valley Governorate, Egypt |
| Reference | Osler [ | Scholz | Scholz | Scholz | Present study |
| Total length (mm) | 40–280 | 108 | 39–42 | 72–119 | 12–30 |
| Maximum width (mm) | 1.59 | 0.90 | 0.70–0.86 | 0.89 | 0.7–0.9 |
| Mature proglottid (length/width ratio) | 1.07 | N/A | N/A | 0.84–1.22 | 0.555–0.525 |
| Gravid proglottid (length/width ratio) | 1.19 | N/A | N/A | 1.61–2.46 | 1.8–2.85 |
| Scolex width (μm) | 270–320 | 390–470 | 295–370 | 340–425 | 170–250 |
| Scolex length (μm) | 200–250 | 290–410 | 185–275 | 245–310 | 150–170 |
| Sucker width (μm) | 120–150 | 175–200 | 115–180 | 155–175 | 135–165 |
| Apical organ | Present (degenerate) | Present | Present | Absent | Present |
| Apical organ width (μm) | 23 | 95–130 | 75–130 | Not applicable | 75–105 |
| Relative length of the cirrus sac | About 17% | 22% | 19%–27% | 19%–23% | 10%–15% |
| Genital pore (position) | Approximately 33% (23%–26%) | 28% | 17%–21% | 16%–22% | 20% |
| No. | Chemical constituents | Results |
|---|---|---|
| 1 | Carbohydrates/glycosides | + |
| 2 | Flavonoids | − |
| 3 | Sterols | + |
| 4 | Diterpenes | + |
| 5 | Anthraquinones | − |
| 6 | Saponins | − |
| 7 | Tannins | − |
| 8 | Alkaloids | − |
| 9 | Cardiac glycosides | − |
| 10 | Coumarins | − |
| Parameter | Control group (tapeworms treated with PBS) | 25 µg/mL | 50 µg/mL | 100 µg/mL |
|---|---|---|---|---|
| Mortality (%) | 70.39 ± 1.23ᵃ | 7.58 ± 0.15ᵇ | 5.79 ± 0.08ᵃᵇ | 4.247 ± 0.09ᶜ |
| Paralysis (%) | 67.77 ± 1.52ᵃ | 7.16 ± 0.09ᵇ | 5.12 ± 0.05ᵃᵇ | 3.40 ± 0.11ᶜ |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParasite Biology and Host Interactions · Aquaculture disease management and microbiota · Parasites and Host Interactions
INTRODUCTION
The frogs Amietophrynus kassasii are commonly found across stagnant freshwater habitats within Egypt’s Nile Valley and Delta regions [1]. Members of the genus Ophiotaenia, originally described under the order Proteocephalidea Mola, 1928 and now placed within the order Onychophora (Caira, Jensen, Waeschenbach, Olson, and Littlewood, 2004), represent a diverse group of cestodes widely distributed among reptiles, small fish, and amphibians, particularly frogs, with more than 130 recognized species [2, 3]. In Africa, most records of proteocephalidean tapeworms belonging to Ophiotaenia date back to the early 20^th^ century. The genus Ophiotaenia La Rue, 1911 is the second most species-rich group within the Proteocephalidae, comprising over 60 species that parasitize reptiles worldwide [4]. Despite this diversity, the true number of proteocephalids infecting amphibians and reptiles is likely underestimated, largely due to the high biodiversity of hosts and the limited research attention historically devoted to their parasitic fauna [4–7].
Ophiotaenia is considered a non-monophyletic assemblage of tapeworms capable of parasitizing a wide range of amphibian and reptilian hosts across the globe. Although species are generally thought to be host-specific within tetrapod groups, they are particularly significant because they serve as valuable model taxa for studies of host–parasite coevolution, diversification, and the biogeographical processes shaping parasite distribution. A striking example is the detection of the North American species Ophiotaenia perspicua (La Rue, 1911), originally from Nerodia rhombifer, in Europe, an instance of “parasite spillover” that highlights how global trade and the movement of animals can introduce novel parasite lineages into new ecosystems [8].
While the life cycles of a few Ophiotaenia species have been partially elucidated, the developmental biology of many, particularly Neotropical taxa, remains unknown. Copepods serve as the first intermediate hosts, whereas insects, tadpoles, and small fish may act as paratenic or secondary intermediate hosts [9].
Marine natural products represent an important reservoir of bioactive compounds derived from marine microorganisms, plants, and animals. These compounds possess diverse pharmacological activities, including antibacterial, antifungal, anti-inflammatory, and antiviral properties [10–12]. Within this context, soft corals of the genus Sinularia are notable for producing a wide variety of biologically active metabolites, such as diterpenes, steroids, and fatty acids, conferring antibacterial, anti-inflammatory, and antioxidant properties [13]. These compounds play essential ecological roles as chemical defenses, mediate predator–prey interactions, and hold considerable promise as sources of novel therapeutic agents [14].
Although helminth infections in amphibians have been documented in several regions of the world, information on proteocephalidean cestodes, particularly those belonging to the genus Ophiotaenia, remains extremely limited across North Africa and the Middle East. Existing surveys in Egypt focus largely on amphibian biodiversity, while parasitological investigations are outdated, geographically restricted, or incomplete. No previous study has reported Ophiotaenia species parasitizing A. kassasii, leaving a significant gap in understanding the distribution, host range, and morphological variability of these cestodes in Egyptian amphibians. Furthermore, most available data on Ophiotaenia life cycles and intermediate hosts remain fragmented, hindering ecological interpretation of transmission dynamics in local freshwater ecosystems. In parallel, despite global interest in marine-derived natural products for antiparasitic drug discovery, the anticestodal potential of Sinularia, a chemically rich soft coral genus, has not been evaluated against frog-infecting cestodes. The absence of baseline epidemiological data, combined with the lack of experimental evidence on natural marine extracts against Ophiotaenia species, highlights the need for integrative parasitological and pharmacological research.
The present study was designed to address these critical knowledge gaps by conducting the first investigation of Ophiotaenia species infection in A. kassasii frogs from the New Valley Governorate, Egypt, supported by detailed morphological and SEM examination for accurate species-level characterization. Additionally, this work aimed to evaluate the in vitro anticestodal efficacy of methanolic Sinulariasp. extract against adult Ophiotaenia specimens using motility, paralysis, and mortality indicators, coupled with ultrastructural assessment of tegumental alterations induced by treatment. By integrating ecological, taxonomic, and pharmacological approaches, this study seeks to generate foundational epidemiological data, elucidate key morphological traits of the isolated cestodes, and explore the therapeutic potential of Sinularia-derived bioactive metabolites as alternative anthelmintic candidates. Ultimately, the findings aim to enhance understanding of cestode–amphibian interactions in Egypt and contribute to the growing field of marine natural-product-based antiparasitic research.
MATERIALS AND METHODS
Ethical approval
All procedures involving amphibian collection, handling, transportation, and laboratory experimentation were conducted in strict accordance with the ethical guidelines for the care and use of animals in scientific research. The study protocol was reviewed and approved by the Animal Care and Use Committee of the Faculty of Sciences, Assiut University, Egypt (Approval No. 01.2025.0004). Permission for the collection of A. kassasii specimens from natural freshwater habitats in the New Valley Governorate was granted by the local environmental authorities.
Frogs were collected and handled using non-invasive methods to minimize stress and avoid unnecessary injury. Only freshly dead frogs were used for parasitological examination, and no animals were euthanized solely for research purposes. All dissections were carried out by trained personnel under controlled laboratory conditions using sterile instruments to ensure animal welfare and biosafety.
Experimental procedures involving cestodes isolated from the frog intestines complied with national and international ethical standards, including the Guide for the Care and Use of Laboratory Animals (National Research Council, USA), the World Organization for Animal Health standards for animal welfare, and the animal research: Reporting of In Vivo experiments guidelines for transparent reporting of animal-based research. All samples were disposed of in accordance with institutional biosafety and environmental regulations.
Study period and location
The study was conducted from February to September 2024. A total of 85 A. kassasii frogs (Figure 1) were collected from freshwater ponds located in the New Valley Governorate, Egypt (25.5°N, 29.5°E). These semi-permanent ponds, primarily used for irrigation and livestock watering, support dense aquatic vegetation dominated by Typha and Phragmites species, providing suitable habitats for amphibians and associated invertebrates.
The frogs (Amietophrynus kassasii) appear yellowish with brown spots on their back, besides dark to black dots on the abdomen of their forelimb toes. The average length and width were 11–13.5 and 1.8–2.7 cm, respectively.
Environmental parameters, including temperature (24°C–30°C), pH (7.0–7.8), and dissolved oxygen (6–8 mg/L), were recorded and found to be within normal freshwater ranges. Sampling was conducted during spring and early summer to coincide with periods of high amphibian activity and increased parasite transmission potential. Freshly dead frogs were transported to the Zoology Department laboratory, New Valley University, for parasitological examination. Intestines were dissected, placed in saline solution, and screened under a stereomicroscope. Recovered tapeworms were thoroughly washed in 0.85% saline solution for subsequent morphological identification and in vitro assays.
Morphological examination of tapeworms
Whole-mount preparations of isolated cestodes were stained with acetic acid-alum carmine, followed by dehydration through ascending ethanol concentrations. Specimens were differentiated in acid alcohol (1 mL concentrated HCl per liter of 70% ethanol), dehydrated fully through 70%–100% ethanol, and cleared in xylene. Mounting was performed using Canada balsam under a coverslip, and slides were dried at 37°C before examination under light microscopy [15].
Preparation and chemical profiling of Sinularia sp. extract
Soft coral samples of Sinularia sp. were collected from the Egyptian Red Sea coast and transported immediately to the laboratory. Approximately 250 g of tissue was cut into small pieces and extracted in methanol at room temperature. The mixture was filtered (Whatman, Merck, Germany), and the solvent was evaporated using a rotary evaporator at 50°C, yielding 4 g of crude methanolic extract.
The resulting extract underwent preliminary chemical screening following established protocols to determine its major chemical constituents [16].
In vitro anticestodal assay
Adult Ophiotaenia sp. tapeworms (n = 6) were incubated in Petri dishes containing Sinularia sp. extract at concentrations of 25, 50, and 100 µg/mL (triplicate dishes per concentration). Controls were maintained in phosphate-buffered saline (PBS) without extract. All experiments were performed at 37°C ± 1°C. Tapeworm motility, paralysis, and mortality were recorded at 30-min intervals to assess cestocidal efficacy [17].
Scanning electron microscopy (SEM)
Tapeworms from the control group and those exposed to the highest effective concentration of Sinularia sp. extract were prepared for SEM analysis. Samples were fixed in 5% glutaraldehyde for 24 h, rinsed 3 times with PBS, post-fixed in osmium tetroxide for 1.5 h, and dehydrated through 90%, 95%, and 100% ethanol. Specimens were mounted on metallic stubs, sputter-coated with gold–palladium and examined using a JOEL JSM-T200 SEM (JOEL, Japan) operating at 20 kV [18].
Statistical analysis
Data were expressed as mean ± standard error. Differences among treatments were evaluated using one-way analysis of variance, followed by Duncan’s post hoc test using the Statistical Package for the Social Sciences version 20 (IBM Corp., NY, USA). Statistical significance was set at p < 0.05. Epidemiological descriptors, including infection rate, mean intensity, and abundance, were calculated using Quantitative Parasitology on the Web (QP 3.0; http://www.zoologia.hu/qp/).
RESULTS
Epidemiological analysis of Ophiotaenia sp. infection
Parasitological examination revealed that Ophiotaenia sp. infection occurred in 5 out of 85 examined A. kassasii frogs, yielding an infection rate of 5.9% (95% confidence interval [CI]: 1.93%–13.20%). Although the prevalence was relatively low, the mean intensity among infected frogs was remarkably high, with an average of 70 parasites per host (95% CI: 35.00–90.00). This indicates a substantial parasite burden in the few frogs that harbored infections. The mean abundance, representing the average number of parasites across all sampled frogs (infected and uninfected), was 4.12 (95% CI: 1.18–8.82) (Table 1).
Morphological characteristics of recovered tapeworms
The recovered adult tapeworms measured 2–30 mm in length and 0.7–0.9 mm in width. The scolex displayed two prominent spherical suckers accompanied by a distinct apical organ. Mature proglottids exhibited a length-to-width ratio of 0.555–0.525, whereas gravid proglottids were filled with approximately 25–30 eggs per segment (Figure 2). A comparative morphometric analysis showed notable similarities and differences relative to previously described Ophiotaenia species (Table 2) [7, 19].
Morphological features of Ophiotaenia sp. outlining: (A) Gross morphology. (B) Unstained whole mount showing the scolex. (C) Magnified photomicrographs of Ophiotaenia sp. stained with acetic acid-alum carmine showing the scolex and two suckers (black arrows) (x40). (D) Two suckers and the apical organ (green arrow) (x100). (E) Mature segments are broader than long (x100). (F) Gravid segments are filled with eggs (x100).
Examination of the intestinal contents of infected frogs revealed multiple insect fragments, suggesting ingestion of potential intermediate or paratenic hosts. These included coleopteran parts (elytra of ground beetles and bodies of darkling beetles), orthopteran structures (legs of mole crickets), and hymenopteran remains (ants, bees, and wasps) (Figure 3).
Multiple insect fragments isolated from the infected frogs showing: (A) The elytra wing of a ground beetle. (B) Body of the darkling beetles. (C) Digging leg of a mole cricket. (D) Body of an ant. (E) A bee. (F) A wasp. (x40).
Chemical profiling of Sinularia sp. extract and the in vitro anticestodal activity
Preliminary chemical screening of the methanolic extract of Sinularia sp. indicates the presence of glycosides, sterols, and diterpenes (Table-3). The methanolic extract of Sinularia sp. demonstrated dose-dependent anticestodal efficacy. Mortality of adult tapeworms occurred at 7.58 ± 0.15 h and 5.79 ± 0.08 h when exposed to 25 µg/mL and 50 µg/mL concentrations, respectively. The highest concentration, 100 µg/mL, induced significantly faster mortality at 4.247 ± 0.09 h, whereas tapeworms in the control group remained active for 70.39 ± 1.23 h (Table 4 and Figure 4).
In vitro anticestodal effect of different concentrations of Sinularia sp. extract on adult stages of Ophiotaenia sp. in terms of mortality and paralysis.
Ultrastructural alterations observed by SEM
SEM analysis of control tapeworms revealed the typical morphology of Ophiotaenia sp., including a scolex with four spherical suckers and a cirrus sac containing a large central cavity (~25 × 45 μm). Mature proglottids displayed clear genital pores.
In contrast, tapeworms treated with 100 µg/mL Sinularia sp. extract exhibited profound structural damage. Observed alterations included:
- Constriction of the cirrus sac cavity
- Shrinkage of two suckers and invagination of the remaining two
- Presence of calcareous corpuscles and vitelline cells in gravid segments
- Tegumental peeling on one side of the proglottids
- Pronounced surface erosion, wart-like projections, and segment contraction
- Genital pores positioned closer together due to proglottid distortion.
These findings confirm that Sinularia sp. extract causes severe tegumental disruption, compromising essential structural and functional features of the cestode (Figure 5).
Scanning electron microscopy photomicrographs of Ophiotaenia sp. showing: (A–C) The control group of Ophiotaenia sp. (A) The scolex with four spherical suckers. (B) The cirrus-shaped sucker has a large central cavity measuring approximately 25 × 45 μm. (C) Mature segments with the genital pore (red arrow). (D-F) The treated group Ophiotaenia sp. (100 µg/mL). (D) The cirrus with a narrow cavity in the scolex, shrunken in two suckers, and invaginated in another two suckers. (E) Mature segments showing eroded tegmental surface, conspicuous warts, and eruptions are visible on every segment, and contraction caused the genital pores of two segments to be located at the same level due to alteration in their site (red arrows). (F) Gravid proglottids appear to be severely shrunken with acicular and capilliform filitriches.
DISCUSSION
Epidemiological significance of Ophiotaenia sp. infection
This study provides the first confirmed isolation and identification of Ophiotaenia species infecting A. kassasii in the New Valley Governorate of Egypt. The infection rate observed (5.9%) was relatively low; however, the mean intensity of 70 parasites per infected frog indicates a substantial parasitic burden, suggesting that although few frogs were affected, those individuals likely experienced considerable physiological stress. This pattern reflects the characteristic epidemiology of helminths, where a minority of hosts harbor disproportionately high parasite loads, a form of aggregated distribution frequently reported in natural amphibian populations.
Reported prevalence of Ophiotaenia species varies widely across regions and host species, with some studies documenting extremely low infection rates (0.41%–3%) [20], while others report much higher levels, such as the 75% infection rate in Telmatobius dankoi from the Andes [21]. These differences are influenced by environmental conditions, host behavior, and availability of intermediate hosts. As amphibian populations decline worldwide due to habitat alteration, climate change, and anthropogenic pressures [22], understanding parasite–host dynamics becomes increasingly critical. Human encroachment into wildlife habitats and increased contact with amphibians and reptiles also raise concern about potential zoonotic risks, although such risks remain largely hypothetical for Ophiotaenia species [23].
Morphological characteristics and host–parasite associations
Members of the family Proteocephalidae, including the genus Ophiotaenia La Rue, 1911, typically parasitize reptiles and amphibians. Although generally host-specific, Ophiotaenia species have been documented in various congeneric hosts across multiple continents [4, 5, 8, 24–26]. In the present study, adult tapeworms measured 12–30 mm × 0.7–0.9 mm, possessing a scolex with two spherical suckers and a distinct apical organ. Mature proglottids displayed a length-to-width ratio of 0.555–0.525, and gravid segments contained numerous eggs. Morphometric comparisons revealed several differences from previously described species by Scholz et al. [7] and Osler [19].
Analysis of intestinal contents from infected frogs revealed fragments of coleopteran, orthopteran, and hymenopteran insects, suggesting their potential role as intermediate or paratenic hosts. Despite the importance of these hosts, the life cycles of proteocephalidean tapeworms remain poorly understood. Most Ophiotaenia species are believed to require two intermediate hosts, starting with small aquatic or terrestrial insects that harbor larval stages. Some species, such as those infecting Ameiurus nebulosus, may utilize paratenic hosts [8]. Although no human infections have been reported, maintaining proper hygiene when handling wild animals or insects is recommended [25].
Transmission pathways for species infecting terrestrial vertebrates, such as Thaumasioscolex didelphidis, remain largely unknown, though frogs, fish, and insects are believed to mediate transmission [27]. In temperate aquatic systems, many proteocephalids rely solely on copepods as intermediate hosts, wherein the plerocercoid stage develops [8, 28].
Diversity and phylogenetic considerations
Proteocephalidean cestodes in Africa most commonly infect snakes and catfish. In Egypt, Ophiotaenia tessellata has been recorded in Natrix tessellata [29]. Phylogenetic analyses indicate that Ophiotaenia is polyphyletic, comprising morphologically similar yet distantly related taxa, likely due to convergent evolution [29]. Overall, the diversity, host range, and life cycles of proteocephalids in anurans remain underexplored. Research limitations stem from declining amphibian populations, protection status, and the lack of comprehensive morpho-molecular characterizations. Moreover, the absence of experimental infections and histological analyses restricts the understanding of parasite impacts on native and non-native hosts [26].
Anticestodal activity of Sinularia sp. extract
The methanolic extract of Sinularia sp. exhibited clear dose-dependent anticestodal activity against adult Ophiotaenia species. Incubation with increasing concentrations of the extract significantly reduced parasite motility, with the fastest mortality observed at 100 µg/mL (4.247 ± 0.09 h). SEM examination demonstrated severe ultrastructural damage in treated specimens, including narrowing of the scolex cirrus, pronounced tegumental erosion, and architectural disruption of proglottids. In contrast, control tapeworms retained intact scoleces, suckers, and tegumental surfaces.
The tegument plays a vital role in nutrient absorption, immune evasion, osmoregulation, and protection from host digestive enzymes. Thus, structural disruption directly interferes with parasite survival. Effective anthelmintics typically target the tegument [30, 31], supporting the strong cestocidal potential observed in Sinularia sp. extract.
Bioactive potential of marine-derived compounds
In the current study, preliminary chemical screening of the methanolic extract of Sinularia sp. indicates the presence of glycosides, sterols, and diterpenes*. Sinularia*, a widely distributed genus of soft corals, consists of interconnected polyps forming a colonial organism. They produce diverse bioactive metabolites, including sesquiterpenes, diterpenes, norditerpenes, polyhydroxylated steroids, and polyamines, with demonstrated antimicrobial, antifungal, and anti-inflammatory effects [32, 33]. Essential oils from Sinularia species have also been isolated from samples collected in the Iranian coastline [34]. Numerous studies [35–38] support the anthelmintic efficacy of marine-derived phytochemicals, often in a dose-dependent manner. For instance, phenolic compounds from Acanthophora spp. inhibit Haemonchus contortus larvae [39], while Bifurcaria bifurcata extracts show activity across developmental stages of Heligmosomoides polygyrus bakeri [40]. A halogenated β-bisabolene sesquiterpenoid from Laurencia scoparia demonstrated potent antinematodal activity against L4-stage Nippostrongylus brasiliensis [41], emphasizing the therapeutic promise of marine natural products.
CONCLUSION
This study provides the first confirmed report of Ophiotaenia sp. infection in A. kassasii frogs in the New Valley Governorate of Egypt, establishing essential baseline data for amphibian parasitology in the region. Although the overall prevalence was low (5.9%), infected frogs carried a remarkably high mean intensity of 70 parasites per host, indicating a substantial trematode burden capable of impairing host physiology and potentially affecting population health. Detailed morphological and SEM examinations confirmed key diagnostic features of the species, while gut-content analysis, revealing coleopteran, orthopteran, and hymenopteran fragments, supports the role of insect taxa in parasite transmission.
The methanolic extract of Sinularia sp. demonstrated strong dose-dependent anticestodal activity, with the highest concentration (100 µg/mL) producing the fastest mortality of adult tapeworms (4.247 ± 0.09 h). SEM findings revealed extensive tegumental damage in treated worms, including sucker shrinkage, cirrus cavity constriction, and severe erosion of proglottids, highlighting the extract’s potent cestocidal properties. These observations underscore the promising therapeutic potential of marine-derived natural compounds as alternative anthelmintic agents.
The study’s strengths include being the first regional documentation of Ophiotaenia in A. kassasii, the combined use of light microscopy and SEM for accurate morphological validation, the incorporation of ecological evidence through gut-content analysis, and the clear experimental demonstration of the cestocidal effects of Sinularia sp. extract. However, certain limitations remain, including the geographically restricted sample, the absence of molecular identification for species-level confirmation, the lack of seasonal prevalence data, and the reliance on in vitro assays, which may not capture the full complexity of host–parasite interactions.
Future research should incorporate molecular sequencing to resolve phylogenetic placement, broaden sampling across multiple habitats and seasons, and investigate the full life cycle to identify intermediate and paratenic hosts. In vivo trials are also needed to validate the safety and efficacy of Sinularia extracts, along with chemical characterization studies to isolate and identify the specific bioactive compounds responsible for cestocidal activity.
Overall, this study advances the understanding of cestode infections in Egyptian amphibians and highlights the potential of Sinularia species as a promising source of natural anthelmintic agents. These findings contribute valuable insights into regional biodiversity, parasite ecology, and the future development of marine-derived antiparasitic therapies.
DATA AVAILABILITY
All the generated data are included in the manuscript.
AUTHORS’ CONTRIBUTIONS
BSA, MA, AA, MAT, and FASA: Conceptualized the study, methodology, and manuscript preparation. HA, GDZ, and GEB: Formal analysis and drafted the manuscript. All authors have reviewed and approved the final version of the manuscript.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ibrahim A.A Amphibians of Egypt:A troubled resource Basic Appl. Herpetol 201327107117
- 2De Chambrier A Beveridge I Scholz T Tapeworms (Cestoda:Proteocephalidae) of Australian reptiles:Hidden diversity of strictly host-specific parasites Zootaxa 2018446144774983031406310.11646/zootaxa.4461.4.2 · doi ↗ · pubmed ↗
- 3Scholz T Kuchta R Fish tapeworms (Cestoda) in the molecular era:Achievements, gaps and prospects Parasitol 2022149(14)1876189310.1017/S 0031182022001202 PMC 1101052236004800 · doi ↗ · pubmed ↗
- 4De Chambrier A Binh T.T Scholz T Ophiotaenia bungari n. sp. (Cestoda), a parasite of Bungarus fasciatus (Schneider) (Ophidia:Elapidae) from Vietnam, with comments on relative ovarian size as a new and potentially useful diagnostic character for proteocephalidean tapeworms Syst. Parasitol 201281(1)39502213900810.1007/s 11230-011-9320-0 · doi ↗ · pubmed ↗
- 5Diard L De Chambrier A Waeschenbach A Scholz TA new tapeworm from Compsophis infralineatus (Pseudoxyrhophiidae), an endemic snake of Madagascar:Scratching the surface of undiscovered reptilian parasite diversity Parasitol. Int 2022881025383500776310.1016/j.parint.2022.102538 · doi ↗ · pubmed ↗
- 6Torres-Carvajal O Pazmiño-Otamendi G Salazar-Valenzuela D Reptiles of Ecuador:A resource-rich online portal, with dynamic checklists and photographic guides Amphib. Reptile Conserv 2019131209229
- 7Scholz T De Chambrier A Kudlai O Tkach V.V Mc Allister C.T.AA global survey of tapeworms (Cestoda:Proteocephalidae) of “true“frogs (Amphibia:Ranidae), including a tabulated list of all proteocephalids parasitising amphibians Folia Parasitol. (Praha)202370(9)11910.14411/fp.2023.00937114799 · doi ↗ · pubmed ↗
- 8De Chambrier A Kudlai O Mc Allister C.T Scholz T Discovering high species diversity of Ophiotaenia tapeworms (Cestoda:Proteocephalidae) of watersnakes (Colubridae) in North America Int. Parasitol. Parasites Wildl 20232225527510.1016/j.ijppaw.2023.10.006PMC 1072469838107547 · doi ↗ · pubmed ↗