Seasonal prevalence of extended-spectrum β-lactamase–producing bacteria in food-chain animals, humans, and the surrounding environment in Fayoum governorate: a one health approach
Ayatollah S. El-Zayat, Shrouk E. Khalil, Marwa N. Ahmed, Dina El-Sayed, Neveen Rabie, Enas A. H. Farag, Hanan A. Goda, Ahmad F. Al-Shahaby, Hala R. Ali

TL;DR
This study shows how the presence of drug-resistant bacteria varies with seasons in animals, humans, and environments in Egypt, highlighting the need for One Health surveillance.
Contribution
This is the first comprehensive investigation of seasonal ESBL prevalence in animals, humans, and environments across three seasons in Egypt.
Findings
ESBL-producing E. coli prevalence was highest in summer and fall, while K. pneumoniae peaked in winter.
Environmental samples showed fluctuating ESBL trends, indicating farm environments' role in persistence and spread.
Genotypic resistance genes like blaSHV and blaCTX-M1 correlated with phenotypic resistance and multidrug resistance patterns.
Abstract
Extended-spectrum β-lactamase (ESBL) producers, particularly Escherichia coli and Klebsiella pneumoniae, pose a growing One Health challenge influenced by seasonal variation. This study assessed seasonal impacts on ESBL prevalence among humans, animals, and farm environments. A total of 2,890 poultry samples, 864 samples from dairy cows (including 88 milk samples and 776 rectal swabs), 248 human fecal samples (118 farm workers and 130 hospitalized patients) and 583 environmental samples were collected from Fayoum governorate, across three seasons. The isolation data revealed significant seasonal impacts, particularly in dairy cows and environmental samples, with source-related differences evident within the same season. The phenotypic and genotypic ESBL- analysis of all isolates from different sources across seasons showed that ESBL-producing E. coli demonstrated comparable prevalence…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
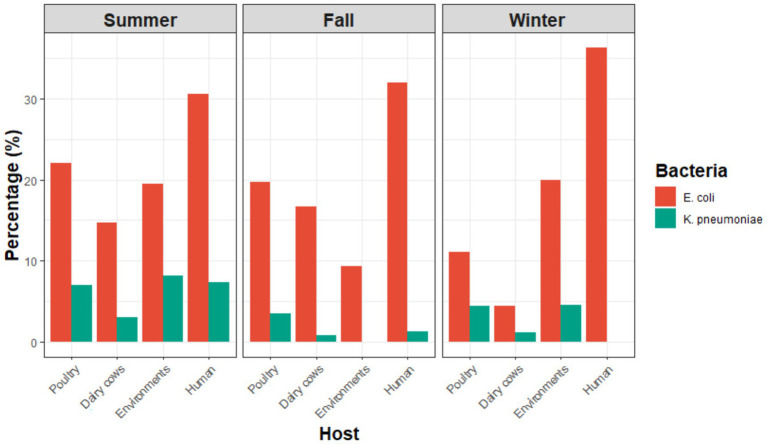 Figure 1
Figure 1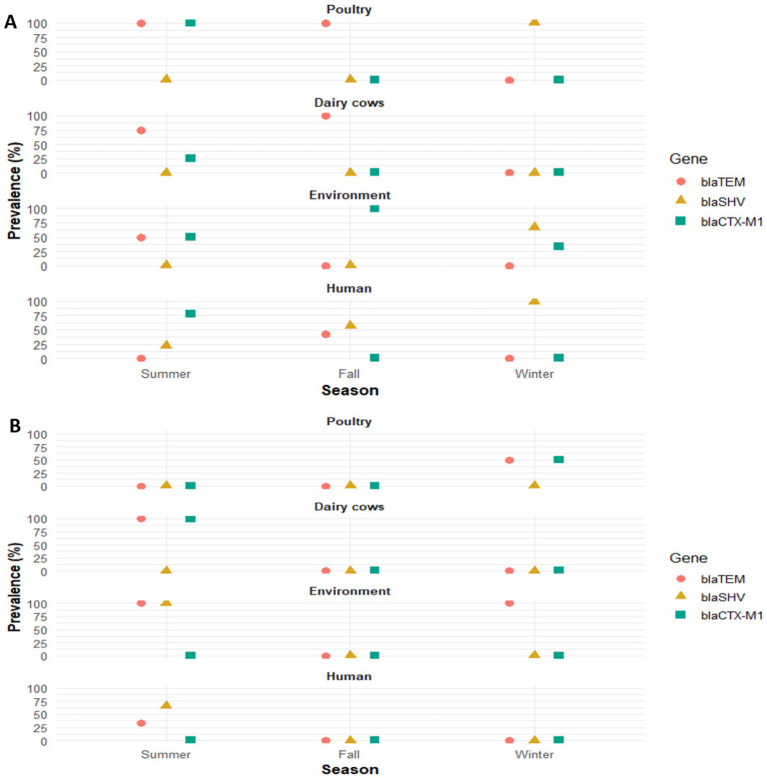 Figure 2
Figure 2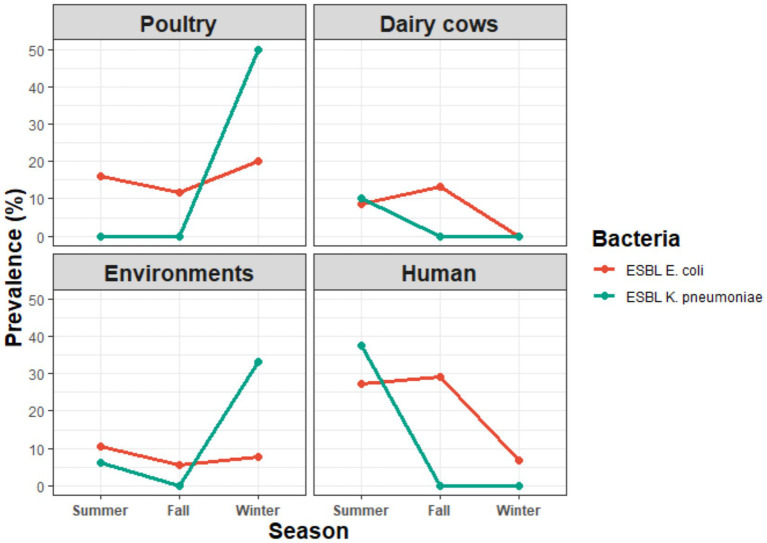 Figure 3
Figure 3 Figure 4
Figure 4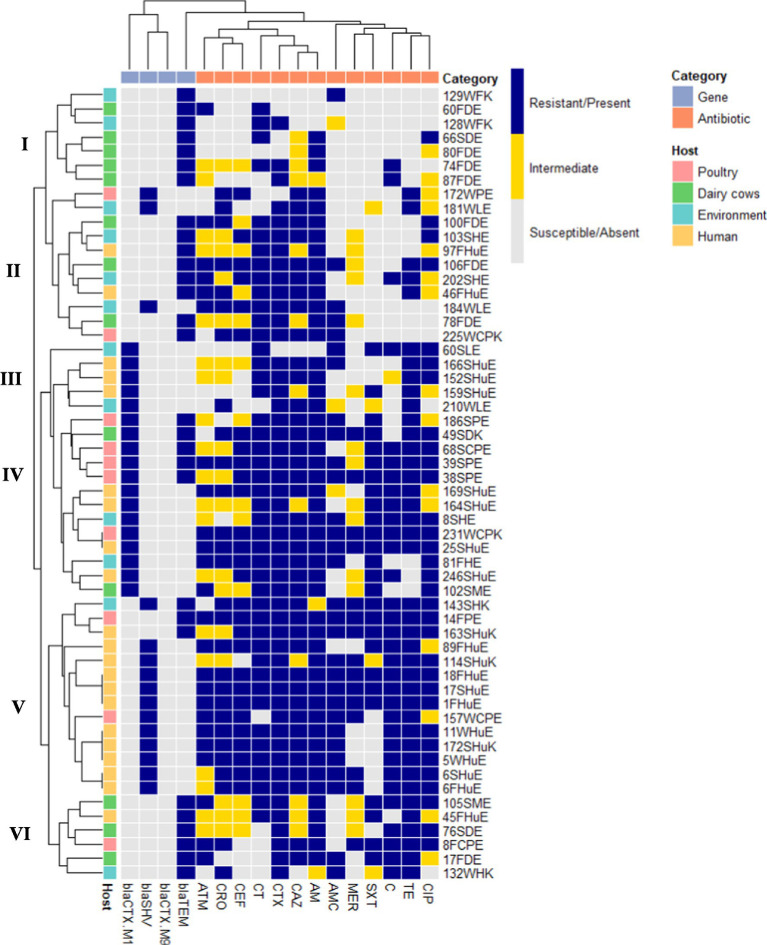 Figure 5
Figure 5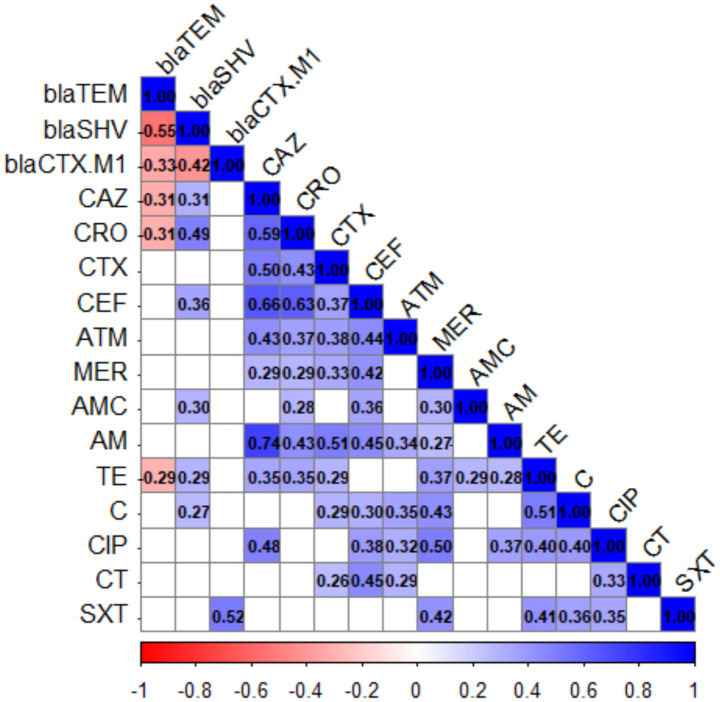 Figure 6
Figure 6| Target gene | Forward primer (5′–3′) | Reverse primer (5′–3′) | Amplicon size (bp) |
|---|---|---|---|
|
| CATTTCCGTGTCGCCCTTATTC | CGTTCATCCATAGTTGCCTGAC | 800 |
|
| AGCCGCTTGAGCAAATTAAAC | ATCCCGCAGATAAATCACCAC | 713 |
|
| TTAGGAAATGTGCCGCTGTA | CGATATCGTTGGTGGTACCAT | 878 |
|
| GCA GAT AAT ACG CAG GTG | CGG CGT GGT GGT GTC TCT | 164 |
| Sample source | Poultry farms | Dairy farms | Hospitals | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sample type | Cloacal swabs | Cecal core | Environment (n:169) | Farm Workers | Rectal swabs n:320 | Milk n:22 | Environment n:26 | Farm workers | Fecal swabs | ||||||||
| Live | Slaughtered | Water | Feed | Litter | Surface | Fecal swab | Healthy | Diseased | Mastitic | Normal | Water | Feed | Slurry | Fecal samples | |||
| Collected No. | 50 | 63 | 55 | 20 | 50 | 44 | 51 | 310 | 10 | 10 | 12 | 10 | 6 | 10 | 7 | 50 | |
| Bacterial species | 14 (28) | 11(17.46) | 10 (18.18) | 5 (25) | 12 (24) | 9 (20.45) | 26 (50.98) | 34 (10.96) | 4 (40) | 7 (70) | 2 (16.66) | 1 (10) | 0 | 1 (10) | 0 | 9 (14) | |
| 2 (4) | 6 (9.5) | 9 (16.36) | 1 (5) | 2 (4) | 2 (4.5) | 7 (13.72) | 8 (2.58) | 2 (20) | 2 (20) | 0 | 0 | 2 (33.3) | 0 | 0 | 1 (2) | ||
| 0.001* | 0.192 | 0.001* | 0.000* | 0.000* | 0.628 | 0.070 | 0.478 | 1 | a | 0.008* | |||||||
| Sample source | Poultry farms | Dairy farms | Hospitals | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sample type | Cloacal swabs | Cecal core | Environment (n:170) | Farm workers | Rectal swabs n:328 | Milk n:38 | Environment n:22 | Farm workers | Fecal samples | ||||||||
| Live | Slaughtered | Water | Feed | Litter | Surface | Fecal samples | Healthy | Diseased | Mastitic | Normal | Water | Feed | Slurry | Fecal samples | |||
| Collected No. | 50 | 36 | 60 | 20 | 50 | 40 | 25 | 316 | 12 | 12 | 26 | 10 | 6 | 6 | 10 | 40 | |
| Bacterial species | 8 (16) | 9 (25) | 2 (3.33) | 0 | 9 (18) | 0 | 19 (76) | 57 (18.03) | 2 (16.66) | 2 (16.66) | 0 | 0 | 0 | 7 (116) | 0 | 5 (12.5) | |
| 3 (6) | 0 | 0 | 0 | 0 | 0 | 0 | 1 (0.31) | 0 | 0 | 2 (7.69) | 0 | 0 | 0 | 0 | 1 (2.5) | ||
| 0.110* | 0.002* | 0.001* | 0.000* | 0.000* | 0.478 | 0.478 | 0.490 | 0.009* | a | 0.201 | |||||||
| Source | Poultry farms | Dairy farms | Hospitals | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sample type | Cloacal swabs | Cecal core | Environment (n:169) | Farm Workers | Rectal swabs n:128 | Milk n:28 | Environment n:27 | Farm workers | Fecal swabs | ||||||||
| Live | Slaughtered | Water | Feed | Litter | Surface | Fecal swab | Healthy | Diseased | Mastitic | Normal | Water | Feed | Slurry | Fecal samples | |||
| Collected No. | 50 | 40 | 70 | 20 | 40 | 39 | 10 | 122 | 6 | 6 | 22 | 9 | 7 | 11 | 15 | 40 | |
| Bacterial species | 2 (4) | 8 (20) | 11 (15.71) | 1 (5) | 22 (55) | 2 (5.12) | 0 | 15 (12.29) | 0 | 0 | 0 | 1 (11.11) | 2 (28.57) | 0 | 13 (86.66) | 16 (40) | |
| 1 (2) | 3 (7.5) | 1 (1.42) | 4 (20) | 3 (7.5) | 0 | 0 | 4 (3.27) | 0 | 0 | 0 | 1 (11.11) | 0 | 0 | 0 | 0 | ||
| 1.000 | 0.105 | 0.000* | a | 0.009* | a | a | a | 0.610 | 0.000* | 0.000* | |||||||
| Source | Bacterial strains (n) | ESBL- phenotypes detection using disk diffusion method | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ESBL-screening antibiotics (Resistance %) | DDST Positive (n) | ||||||||||||||||
| 3rd generation cephalosporins | 4th generation cephalosporin | Monobactam | |||||||||||||||
| Ceftazidime (CAZ) | Ceftriaxone (CRO) | Cefotaxime (CTX) | Cefepime (CEF) | Aztreonam (ATM) | |||||||||||||
| R | I | S | R | I | S | R | I | S | R | I | S | R | I | S | |||
| Poultry | 13 (100) | 0 | 0 | 9 (69) | 3 (23) | 1 (7.6) | 13 (100) | 0 | 0 | 12 (92.3) | 1 (7.6) | 0 | 9 (69.2) | 4 (30.7) | 0 | 4 (30.7) | |
| 4 (80) | 1 (20) | 0 | 5 (100) | 0 | 0 | 5 (100) | 0 | 0 | 5 (100) | 0 | 0 | 5 (100) | 0 | 0 | 0 | ||
| Dairy cattle | 2 (40) | 3 (60) | 0 | 1 (20) | 3 (60) | 1 (20) | 4 (80) | 0 | 1 (20) | 1 (20) | 3 (60) | 1 (20) | 3 (60) | 1 (20) | 1 (20) | 4 (80) | |
| (100) | 0 | 0 | 1 (100) | 0 | 0 | 1 (100) | 0 | 0 | 1 (100) | 0 | 0 | 1 (100) | 0 | 0 | 1 (100) | ||
| Environment | 13 (72) | 4 (22.2) | 1 (5.5) | 12 (66.6) | 4 (22.2) | 2 (11.1) | 17 (94.4) | 0 | 1 (5.5) | 16 (88.8) | 1 (5.5) | 1 (5.5) | 8 (44.4) | 8 (44.4) | 2 (11.1) | 4 (22.2) | |
| 12 (100) | 0 | 0 | 11 (91.6) | 1 (8.3) | 0 | (12) | 0 | 0 | 11 (91.6) | 1 (8.3) | 0 | 10 (83) | 2 (16.6) | 0 | 1 (8.3) | ||
| Human | 13 (81.25) | 3 (18.75) | 0 | 9 (56) | 6 (37.5) | 1 (6.25) | 16 (100) | 0 | 0 | 12 (75) | 2 (12.5) | 2 (12.5) | 9 (56) | 6 (37.5) | 1 (6) | 9 (56.25) | |
| 4 (80) | 1 (20) | 0 | 3 (60) | 2 (40) | 0 | 5 (100) | 0 | 0 | 5 (100) | 0 | 0 | 4 (80) | 0 | 1 (20) | 3 (60) | ||
| Source | Bacterial strains (n) | ESBL- phenotypes detection using disk diffusion method | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ESBL-screening antibiotics (Resistance %) | DDST Positive (n) | ||||||||||||||||
| 3rd generation cephalosporins | 4th generation cephalosporin | Monobactam | |||||||||||||||
| Ceftazidime (CAZ) | Ceftriaxone (CRO) | Cefotaxime (CTX) | Cefepime (CEF) | Aztreonam (ATM) | |||||||||||||
| R | I | S | R | I | S | R | I | S | R | I | S | R | I | S | |||
| Poultry | 7 (77) | 2 (22) | 0 | 7 (77) | 1 (11) | 1 (11) | 9 (100) | 0 | 0 | 7 (77) | 1 (11) | 1 (11) | 7 (77) | 2 (22) | 0 | 2 (22) | |
| 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| Dairy cattle | 8 (47) | 8 (47) | 1 (5.88) | 8 (47) | 5 | 4 (23) | 15 (88.2) | 0 | 2 (11) | 8 (47) | 5 (29) | 4 (23) | 12 (70.6) | 4 (23) | 1 (5) | 8 (47) | |
| 0 | 1 (100) | 0 | 1 (100) | 0 | 0 | 1 (100) | 0 | 0 | 0 | 1 (100) | 0 | 0 | 1 (100) | 0 | 0 | ||
| Environmental | 3 (100) | 0 | 0 | 3 (100) | 0 | 0 | 3 (100) | 0 | 0 | 2 (66) | 1 (33) | 0 | 3 (100) | 0 | 0 | 1 (33) | |
| 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| Human | 13 (86) | 2 (13) | 0 | 13 (86) | 2 (13) | 0 | 14 (93.3) | 0 | 1 (6.6) | 11 (73) | 4 (26.6) | 0 | 13 (86) | 2 (13) | 0 | 7 (46.6) | |
| 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| Source | Bacterial strains (n) | ESBL- phenotypes detection using disk diffusion method | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ESBL-screening antibiotics (Resistance %) | DDST positive (n) | ||||||||||||||||
| 3rd generation cephalosporins | 4th generation cephalosporin | Monobactam | |||||||||||||||
| Ceftazidime (CAZ) | Ceftriaxone (CRO) | Cefotaxime (CTX) | Cefepime (CEF) | Aztreonam (ATM) | |||||||||||||
| R | I | S | R | I | S | R | I | S | R | I | S | R | I | S | |||
| Poultry | 4 (80) | 1 (20) | 0 | 5 (100) | 0 | 0 | 4 (80) | 0 | 1 (20) | 5 (100) | 0 | 0 | 4 (80) | 0 | 1 (20) | 2 (40) | |
| 2 (66.6) | 1 (33.3) | 0 | 2 (66.6) | 0 | 1 (33.3) | 3,100 | 0 | 0 | 3,100 | 0 | 0 | 2 (66.6) | 0 | 1 33.3 | 2 (66.6) | ||
| Dairy cattle | 0 | 2 (100) | 0 | 1 (50) | 0 | 1 (50) | 2 (100) | 0 | 0 | 2 (100) | 0 | 0 | 2 (100) | 0 | 0 | 0 | |
| 1 (100) | 0 | 0 | 1 (100) | 0 | 0 | 1 (100) | 0 | 0 | 1 (100) | 0 | 0 | 1 (100) | 0 | 0 | 0 | ||
| Environment | 8 (88) | 1 (12.5) | 0 | 8 (88) | 0 | 1 (12.5) | 8 (88) | 0 | 1 (12.5) | 6 (66.6) | 0 | 3 (33) | 6 (66.6) | 0 | 3 (33) | 3 (33) | |
| 1 (25) | 0 | 3 (75) | 2 (50) | 0 | 2 (50) | 3 (75) | 0 | 1 (25) | 0 | 1 (25) | 3 (75) | 0 | 1 (25) | 3 (75) | 3 (75) | ||
| Human | 3 (75) | 1 (25) | 0 | 3 (75) | 1 (25) | 0 | 4 (100) | 0 | 0 | 3 (75) | 1 (25) | 0 | 3 (75) | 1 (25) | 0 | 2 (50) | |
| 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Infections and bacterial resistance · Pharmaceutical and Antibiotic Environmental Impacts
Introduction
1
The rapid emergence and global spread of antimicrobial resistance in animals represent a critical challenge that pose a threat to the public health, livestock production, and environmental safety (Hamed et al., 2025; Fathy et al., 2025; Peirano and Pitout, 2019; Van Duin and Paterson, 2016). Among the most alarming resistance mechanisms is the production of extended-spectrum β-lactamases (ESBLs), enzymes that inactivate a broad spectrum of β-lactam antibiotics, including cephalosporin, penicillin, and monobactam (El-Sayed et al., 2022; Superti et al., 2009). Clinically, the most significant ESBL-producing variants belong to the CTX-M, TEM, and SHV families. The frequent localization of bla-ESBL genes on plasmids, together with their close association with mobile genetic elements such as transposons and insertion sequences, has facilitated their rapid and widespread dissemination. Horizontal gene transfer further accelerates this process, enabling efficient transmission of ESBL genes to a wide variety of hosts (Ewers et al., 2021). ESBL-producing Enterobacteriaceae, particularly E. coli and K. pneumoniae, are currently recognized as critical priority pathogens due to their widespread occurrence and association with serious infections in humans and animals (Ribeiro et al., 2024; Ibrahim et al., 2023). The epidemiology of ESBL-producing pathogens has shifted from being exclusively confined to healthcare facilities to becoming a significant problem in the community, food chain, and environment. Food-chain animals serve as an important reservoir of ESBL-producing pathogens, with numerous studies documenting their colonization of poultry (Badr et al., 2022; Dierikx et al., 2013) and dairy cattle worldwide (Kürekci et al., 2024). These super-variants can spread widely via contamination at multiple stages of the food production chain, including on farms, during slaughter or processing, and in retail stores (Perestrelo et al., 2022). Such dynamics not only pose a threat to animal health and livestock productivity, but also represent potential sources of zoonotic disease transmission to humans, either through direct contact, environmental exposure, or consumption of contaminated animal byproducts.
Workers in close contact with animals, such as farmers, slaughterhouse staff, and food handlers, are particularly at risk of colonization with ESBL-producing pathogens (Subramanya et al., 2021); (Ribeiro et al., 2024). The occupational interface enables resistant bacteria to move between animals and humans, providing opportunities for horizontal gene transfer and persistence in human populations. Beyond the direct animal–human interactions, the surrounding environment of soil, water, and animal manure represents an additional reservoir and pathway for the spread of resistant bacteria and related genes (Wielders et al., 2020; Blaak et al., 2015). These interconnected domains illuminate the significance of adopting a One Health approach, recognizing that human, animal, and environmental health are intrinsically linked.
A significant and underexplored dimension of ESBL epidemiology is the role of seasonal variation. Environmental and management conditions change with seasons, influencing bacterial survival, shedding, and transmission (Rose et al., 2001). For example high temperatures and humidity during summer may affect bacterial survival and persistence in the environment, while seasonal changes in animal production cycles, antimicrobial use, and human–animal interactions may also affect prevalence rates. Some studies suggest that resistance levels in foodborne pathogens may peak during certain times of the year (Rose et al., 2001; Hellberg and Chu, 2016) but comprehensive data remain limited. Understanding these temporal patterns is critically important for identifying periods of high risk for disease outbreaks and designing timely intervention approaches.
Studying the seasonal influence on the prevalence of ESBL-producing bacteria among humans, animals, and the environment in Egypt is significantly important for designing targeted and cost-effective interventions. Seasonal characteristics in Egypt, such as hot summers, mild winters, and irrigated agriculture, create fluctuating temperature and humidity conditions that can affect the survival, shedding, and transmission of bacteria along the food chain from farms and live bird markets to slaughterhouses and retail outlets. Seasonal shifts in animal production cycles, disease burden, and antimicrobial use may amplify selective pressure on ESBL producers in poultry and dairy cattle, while increased contact with surface water and irrigation canals may facilitate environmental spread and human exposure.
Identifying peak transmission times and locations enables public health and veterinary services to time screening, biosafety measures, hygiene enhancements, waste and water management controls, and antimicrobial management strategies to achieve maximum impact. Adopting a seasonally- based One Health strategy in Egypt, can therefore highlight transmission hotspots at the human-animal-environment interface, improve risk assessment related to foodborne and occupational exposure, and ultimately reduce the risk of resistant infections while preserving the effectiveness of critically important antibiotics.
Therefore, the current study aims to investigate the impact of seasonal variation on the prevalence of ESBL-producing E. coli and K. pneumoniae across food-chain animals, co-workers, and the surrounding environment in Fayoum Governorate, Upper Egypt, an area around 1800 km2 characterized by high animal population density and intensive farming activities with wide varieties of livestock, mostly poultry and cattle.
Materials and methods
2
Study area
2.1
We planned to screen different samples from multiple food chain sources, including live food-producing animals (cattle and poultry), surrounding environment (feed, water, farm slurry), farm workers and hospitalized patients, from of seven districts (Markas El-Fayoum, Pander El-Fayoum, Yousef-Elsadik, Senores, Tamiya, Ebshway and Itsa) of Fayoum governorate (29.3565° N, 30.6200° E).
Samples collection
2.2
Poultry samples
2.2.1
A total of 2,890 samples were collected from poultry farms and slaughterhouses across three seasons: summer (n = 1,130; 500 flocks, 630 slaughtered), fall (n = 860; 500 flocks, 360 slaughtered), and winter (n = 900; 500 flocks, 400 slaughtered). For each flock, ten cloacal swabs were pooled into a single composite sample. From each slaughterhouse, ten cecal content samples were collected and pooled together.
Dairy cattle samples
2.2.2
From 16 dairy farms, milk samples were collected in summer (n = 22; 12 bulk tank, 10 mastitis cases), fall (n = 38; 26 bulk tanks, 12 mastitis cases), and winter (n = 28; 22 bulk tanks, 6 mastitis cases). Rectal swabs were also collected: summer (n = 320; 310 from apparently healthy cows, 10 from diseased cows), fall (n = 328; 316 from apparently healthy cows, 12 from diseased cows), and winter (n = 128; 122 from apparently healthy cows, 6 from diseased cows).
Environmental samples
2.2.3
From poultry farms, 508 environmental samples were collected: summer (n = 169), fall (n = 170), and winter (n = 169). Summer samples comprised water (n = 55), feed (n = 20), litter (n = 50), and surfaces of knives/cutting boards (n = 44). Fall samples included water (n = 60), feed (n = 20), litter (n = 50), and knife/cutting board surfaces (n = 40). Winter samples included water (n = 70), feed (n = 20), litter (n = 40), and knife/cutting board surfaces (n = 39).
From dairy cattle farms, 75 environmental samples were collected: summer (n = 26), fall (n = 22), and winter (n = 27). Summer samples comprised water (n = 10), feed (n = 6), and slurry (n = 10). Fall samples included water (n = 10), feed (n = 6), and slurry (n = 6). Winter samples included water (n = 9), feed (n = 7), and slurry (n = 11).
Human samples
2.2.4
Anonymized fecal specimens were obtained from bacteriological laboratories conducting routine occupational health monitoring of farm workers and also receive samples from nearby hospitals, where samples were accessed after completion of diagnostic testing and prior to disposal. No direct contact occurred between the research team and donors; all specimens were received in fully de-identified form (Supplementary Figure S1). Seasonal collections included: summer (n = 58 farm workers, n = 50 hospitalized patients), fall (n = 35 farm workers, n = 40 hospitalized patients), and winter (n = 25 farm workers, n = 40 hospitalized patients).
All samples were collected under aseptic conditions, immediately placed on ice, and transferred to a bacteriological laboratory for further analysis.
Isolation and biochemical identification of Escherichia coli and Klebsiella pneumoniae from the collected samples
2.3
Cloacal swabs, rectal swabs from dairy cattle, and fecal samples from hospitalized patient or from farm workers were streaked onto Eosin Methylene Blue Agar (EMBA; Himedia, Mumbai, India) plates (Badr et al., 2022; Mustika et al., 2024). Environmental samples were also obtained, including swabs from knives and cutting boards in slaughterhouses (20–40 cm^2^ surface area). These were collected using sterile cotton swabs moistened with buffered peptone water (Sebsibe and Asfaw, 2020), aseptically transferred into 10 mL sterile peptone water, and vortexed for 30 s to release microorganisms. Additionally, 10 g of cecal content, 25 g each of feed, slurry, and litter, and 25 mL each of water, tank milk, and mastitis milk samples were collected. All samples were subjected to serial decimal dilution in sterile saline solution (0.85% NaCl). Aliquots (1 mL) from appropriate dilutions were dispensed into sterile Petri dishes, overlaid with molten EMBA, gently mixed, and allowed to solidify (Sebsibe and Asfaw, 2020; International Organization for Standardization, 2013). Plates were incubated at 37 °C for 24–48 h to isolate and identify E. coli and K. pneumoniae. On EMBA media, E. coli colonies typically exhibit a metallic green sheen, whereas K. pneumoniae colonies typically appear mucoid pink. Colonies were subsequently enumerated and purified for further identification.
Presumptive E. coli and K. pneumoniae colonies were subjected to primary biochemical examination using IMViC tests (indole, methyl red, Voges-Proskauer, and citrate utilization), along with oxidase and catalase tests. In addition, identification of suspected E. coli and K. pneumoniae isolates was performed using the VITEK® 2 Compact system (bioMérieux, Marcy-l’Étoile, France) (Funke et al., 1998). Isolates were prepared as standardized suspensions in 0.45% NaCl, inoculated into GN (Gram-negative) identification cards, and incubated in the automated system. Biochemical profiles were interpreted by the VITEK® 2 software, and species-level identification was determined according to the manufacturer’s guidelines.
ESBL-phenotypic characterization of Escherichia coli and Klebsiella pneumoniae isolates
2.4
The biochemically/VITEK positive isolates for E. coli and K. pneumoniae were then cultured on HiCrome ESBL agar media (Himedia ®, Mumbai, India). This is a chromogenic screening medium designed for the selective isolation of ESBL-producing organisms. The bacteria that produce ESBL are distinguished by color using a chromogenic combination. ESBL producing E. coli grow as either pink or purple colonies whereas ESBL producing K. pneumoniae grow as bluish green colonies. Moreover, the phenotypic identification of ESBL-producing bacteria also obtained through double synergy test (Badr et al., 2022; Bubpamala et al., 2018) and antibiotic resistance pattern (Ryoo et al., 2005; Balta et al., 2024).
Double disc synergy (DDST)
2.4.1
An amoxicillin-clavulanic disk (AMC, 30 μg) was placed 20 mm (center to center) from each antibiotic disc third-generation cephalosporins: Ceftazidime, Cefotaxime, and Ceftriaxone (30 μg each); and fourth-generation cephalosporin: Cefepime (30 μg) on Mueller-Hinton Agar plate inoculated with the test isolates (Badr et al., 2022). Following a 24-h incubation at 37 °C, an abrupt rise or augmentation of the inhibition zone of any cephalosporin discs toward to the AMC disc was deemed indicative of ESBL production.
Antibiotic resistance pattern
2.4.2
Antibiotics-ESBL screening was carried out by Kirby-Bauer technique employed the disk diffusion test to produce qualitative classifications of susceptibility, including sensitive, intermediate, and resistant evaluations. This method based on various cephalosporins according to the Clinical and Laboratory Standards Institute (CLSI) standard. The antibiotics concentrations were selected according to CLSI guidelines. Isolates with an inhibition zone size of ≤22 mm with ceftazidime (30 μg), ≤25 mm with ceftriaxone (30 μg), ≤27 mm with cefotaxime (30 μg), ≤18 mm Cefepime (30 μg), and ≤15 mm with aztreonam (10 μg) were identified as potential ESBL producers CLSI (Clinical and Laboratory Standards Institute) (2021) Performance standards for antimicrobial susceptibility testing. 25th informational supplement, Clinical and Laboratory Standards Institute, Wayne, M100-S25. CLSI (2023). Performance Standards for Antimicrobial Susceptibility Testing. 33rd ed. CLSI supplement M100. Furthermore, antibiotic susceptibility profiling was performed using the Kirby–Bauer disk diffusion method (Kirby and Bauer, 1966) against eight antibiotics (Bioanalyse, Ankara, Turkey): oxytetracycline (TE, 30 µg), colistin (CT, 10 µg), meropenem (MEM, 10 µg), ampicillin (AM, 10 µg), ciprofloxacin (CIP, 5 µg), chloramphenicol (C, 30 µg), amoxicillin/clavulanate (AMC, 20/10 µg), and sulfamethoxazole/trimethoprim (SXT, 1.25/23.75 µg). The results of antibiotic susceptibility testing were interpreted according to the guidelines of the Clinical and Laboratory Standards Institute (CLSI) (Balta et al., 2024).
ESBL-genotypic characterization of Escherichia coli and Klebsiella pneumoniae isolates
2.5
The phenotypically ESBL-positive E. coli and K. pneumoniae isolates were examined by multiplex PCR to detect the presence of ESBL genes, including blaTEM, blaSHV, and blaCTX-M (groups 1 and 9) as previously described (Bubpamala et al., 2018; Ryoo et al., 2005). Briefly, DNA was extracted from the phenotypically ESBL-positive isolates using the GeneDireX DNA Extraction Kit (Taiwan) following the manufacturer’s instructions. The extracted DNA was then amplified using gene-specific multiplex PCR reactions. The cycling conditions were as follows: initial denaturation at 94 °C for 10 min, followed by 30 cycles of denaturation at 94 °C for 30 s, annealing at 60.1 °C for 35 s, and extension at 72 °C for 1 min. A final extension step was performed at 72 °C for 9 min. The primers sequences used in this study are listed in Table 1.
Additionally, in order to study the distribution of ESBL genes among bacterial isolates was analyzed in relation to their antibiotic resistance profiles using hierarchical heat-map visualization.
Correlation analysis of ESBL genes and antibiotic resistance profiles
2.6
Pairwise correlations between antibiotic resistance genes and antimicrobial susceptibility profiles were calculated using the Pearson correlation coefficient (r). Only numeric variables were included, with categorical data converted to ordinal values (0 = sensitive/absent, 1 = intermediate, 2 = resistant/present). Zero-variance variables were excluded to avoid bias. Correlation matrices were computed using the cor() function with pairwise complete observations. Corresponding p-values were obtained using the cor() function and subsequently adjusted for multiple comparisons using the Benjamini–Hochberg false discovery rate (FDR) method. Only correlations that remained significant after FDR correction were interpreted.
Statistical analysis
2.7
Statistical analysis of the data was conducted using SPSS version 22 via Chi-Square tests and Fisher’s Exact Test. The Chi-Square test is used to determine the association between samples sources, seasons and isolation rate for E. coli and K. pneumoniae and ESBL-production. The result of E. coli and K. pneumoniae in different demographic variables by Chi-Square test and for cells with expected count below 5 were tested using Fisher’s exact test and all statistical analysis were performed at p < 0.05. Statistical significance of correlations was assessed using cor.test, and corresponding p-values were adjusted at confidence level 95%. Pearson correlation coefficient (r) and all data was showed and plotted using R version 4.3.2 Software.
Results
3
Prevalence rate of Escherichia coli and Klebsiella pneumoniae isolated from the collected samples across three seasons
3.1
During summer season, a total of 500 cloacal swabs and 630 cecal cores were collected from poultry and pooled into 50 and 63 samples, respectively (Table 2). From these, 14 E. coli and 2 K. pneumoniae isolates were recovered from cloacal swabs, while 11 E. coli and 6 K. pneumoniae were obtained from cecal cores. Poultry farm environmental samples (n = 169; water, feed, litter, and surface swabs), yielded 36 E. coli and 14 K. pneumoniae isolates. Additionally, samples from poultry farm workers (n = 51) recovered 26 E. coli and 7 K. pneumoniae. In dairy farms, rectal swabs from healthy cows (n = 320) yielded 34 E. coli and 8 K. pneumoniae, while diseased cows (n = 10) produced 4 E. coli and 2 K. pneumoniae. Mastitic milk samples (n = 10) yielded 7 E. coli and 2 K. pneumoniae, whereas bulk tank milk (n = 12) produced 2 E. coli only. Environmental samples (n = 26) recovered 2 E. coli and 2 K. pneumoniae, but no isolates were obtained from dairy workers (n = 7). From hospitalized patients, fecal samples (n = 50) yielded 9 E. coli and 1 K. pneumoniae.
In the fall season, cloacal swabs (n = 50 pools) yielded 8 E. coli and 3 K. pneumoniae, while cecal cores (n = 63 pools) produced 9 E. coli with no K. pneumoniae. Poultry environmental samples (n = 170) recovered 11 E. coli only, and farm workers (n = 25) yielded 19 E. coli with no K. pneumoniae. In dairy farms, rectal swabs from healthy cows (n = 316) yielded 57 E. coli and 1 K. pneumoniae, while diseased cows (n = 12) produced 2 E. coli only. Dairy environmental samples (n = 22) yielded 7 E. coli. From hospitalized patients, fecal samples (n = 40) yielded 5 E. coli and 1 K. pneumoniae (Table 3).
In the winter season, cloacal swabs (n = 50) yielded 2 E. coli and 1 K. pneumoniae, while pooled cecal samples (n = 40) yielded 8 E. coli and 3 K. pneumoniae. Poultry farm environmental samples (n = 169) recovered 36 E. coli and 12 K. pneumoniae, whereas no isolates were obtained from poultry worker fecal samples (n = 10). In dairy farms, rectal swabs from healthy cows (n = 122) yielded 15 E. coli and 4 K. pneumoniae, while samples from diseased cows (n = 6) were negative for both organisms. Neither mastitic milk (n = 6) nor normal tank milk (n = 22) yielded any isolates. Environmental samples (n = 27) recovered 3 E. coli and 1 K. pneumoniae, whereas dairy farm workers (n = 15) yielded 13 E. coli but no K. pneumoniae. Similarly, fecal samples of hospitalized patients (n = 40) recovered only 16 E. coli (Table 4).
As shown in Figure 1 and Supplementary Table S1, E. coli prevalence in poultry samples ranged from 22.1% in summer to 19.8% in fall and 11.1% in winter, while K. pneumoniae varied between 7.07% and 3.48–4.44%, with no significant seasonal effect observed. In dairy cows, E. coli prevalence decreased sharply from 14.68–16.7% in summer and fall to 4.4% in winter, and K. pneumoniae dropped from 3.1% to below 1.2%, showing significant seasonal variation with winter as the lowest-prevalence season (p = 0.043). Environmental samples showed fluctuating trends, with E. coli peaking in summer (19.5%) and winter (19.9%) but dropping in fall (9.4%), while K. pneumoniae was present in summer (8.2%) and winter (4.6%) but absent in fall, both demonstrating significant seasonal effects (E. coli p = 0.007 and K. pneumoniae p = 0.000). Among farm workers, E. coli prevalence remained consistently high across seasons (44.8–54.3%) with no significant variation, while K. pneumoniae was detected only in summer (12.1%), reflecting significant seasonality (p = 0.021). In hospitalized patients, E. coli prevalence ranged from 12.5 to 29.1%, and K. pneumoniae remained very low (2–2.5%), with no significant seasonal differences. When comparing across sources within the same season, E. coli showed highly significant differences in all seasons (p = 0.000), while K. pneumoniae differed significantly in summer (p = 0.016) and winter (p = 0.045) but not fall.
The percentage of Escherichia coli and Klebsiella pneumoniae isolates revered from different hosts (Poultry, dairy cows, environments, co-workers and hospitalized patients) across three seasons: Summer, fall, and winter. Data show variations across sources and seasons, with significant seasonal effects.
ESBL- identification
3.2
Phenotypic detection of ESBL-producing isolates across seasons
3.2.1
The E. coli and K. pneumoniae isolates recovered from different hosts across the three seasons were subjected to phenotypic characterization using HiCrome ESBL agar. HiCrome-positive isolates were subsequently confirmed by the ESBL screening antibiotic test and the double-disk synergy test (DDST). The presence of either resistance to at least one third- or fourth-generation cephalosporin, or a synergistic effect of amoxicillin-clavulanic with any cephalosporin antibiotic, was considered phenotypically positive, as detailed in Tables 5–?7.
Table 5 summarizes the phenotypically confirmed ESBL producers among the summer isolates. Of the 13 E. coli isolates recovered from poultry, 4 were confirmed as ESBL producers, whereas all K. pneumoniae isolates from poultry tested negative. In dairy cattle, 4 of 5 E. coli isolates and the single K. pneumoniae isolate were ESBL positive. Among environmental isolates, 4 of 18 E. coli and 1 of 12 K. pneumoniae demonstrated ESBL activity. From human samples, 9 of 16 E. coli isolates and 3 of 5 K. pneumoniae isolates were confirmed as ESBL producers.
In total, 21 of 52 E. coli isolates and 5 of 23 K. pneumoniae isolates recovered during summer were phenotypically confirmed as ESBL producers.
In the fall season, 2 of 9 E. coli isolates from poultry, 8 of 17 E. coli isolates from dairy cattle, 1 of 3 E. coli isolates from environmental samples, and 7 of 15 E. coli isolates from humans were confirmed as ESBL producers. Overall, 18 of 44 E. coli isolates were ESBL producers. In contrast, none of the K. pneumoniae isolates recovered from any host exhibited ESBL activity during fall (Table 6).
In the winter season, 2 of 5 E. coli and 2 of 3 K. pneumoniae from poultry were confirmed as ESBL producers. Neither the 2 E. coli nor the single K. pneumoniae recovered from dairy cattle tested positive. Among environmental isolates, 3 of 9 E. coli and 3 of 4 K. pneumoniae were ESBL positive, while from human samples 2 of 4 E. coli were positive and no K. pneumoniae were recovered. Overall, 7 of 20 E. coli and 5 of 8 K. pneumoniae obtained in winter were confirmed as phenotypic ESBL producers as shown in Table 7.
Genotypic identification and seasonal distribution of ESBL-producers
3.2.2
The PCR analysis revealed variable distribution of ESBL genes among isolates of different sources across seasons (Figure 2). In summer isolates, the blaTEM gene was widely detected in E. coli isolates from poultry (100%), dairy (75%), and environment (50%), but was absent in human isolates. In K. pneumoniae, blaTEM was found in isolates from dairy (100%), environment (100%), and some human samples (33.3%). The blaSHV gene was absent in E. coli from animal and environmental sources but was present in human isolates (22.2%). In K. pneumoniae, blaSHV was detected in environmental isolates (100%) and in humans (66.6%). The blaCTX-M group 1 gene showed high prevalence in E. coli from poultry (100%) and humans (77.7%) and was also present in dairy (25%) and environment (50%). In K. pneumoniae, it was detected only in dairy isolates (100%). None of the examined E. coli or K. pneumoniae isolates harbored blaCTX-M group 9. Among the fall isolates, the most prevalent ESBL gene was blaTEM, detected in 100% of E. coli isolates from both poultry and dairy sources. In human isolates, blaSHV was the most frequent (57.1%), whereas blaTEM was present in 42.85%. In contrast, environmental E. coli isolates predominantly harbored blaCTX-M. While the winter isolates, the blaTEM gene was absent in E. coli from all examined sources but was detected in K. pneumoniae isolates from poultry (50%) and from all environmental samples (100%). The blaSHV gene was consistently present in E. coli isolates from poultry (100%), environment (66.6%), and humans (100%), whereas it was not detected in K. pneumoniae. The blaCTX-M group 1 gene was detected in E. coli isolates from the environment (33.3%) and in K. pneumoniae isolates from poultry (50%). However, blaCTX-M group 1 gene was absent in dairy and human isolates. None of the E. coli or K. pneumoniae isolates harbored blaCTX-M group 9.
Genotypic identification and seasonal distribution of ESBL genes (blaTEM. blaSHV, blaCTX-M1) in Escherichia coli (A) and Klebsiella pneumoniae (B) isolated from poultry, dairy cows, environment, and human.
Prevalence of ESBL-producing Escherichia coli and Klebsiella pneumoniae across seasons
3.3
As shown in Figure 3 and Supplementary Table S2, ESBL-producing E. coli from poultry was detected at comparable rates in summer (16%) and fall (11.76%), with a slight increase in winter (20%). In contrast, ESBL-producing K. pneumoniae from poultry showed significant seasonal variation (p = 0.042), being detected only in winter (50%), but absent in summer and fall. Among dairy cow isolates, E. coli displayed moderate ESBL prevalence in summer (8.5%) and fall (13.11%), but none were detected in winter, while ESBL-K. pneumoniae (10%) was detected only in summer. Environmental E. coli isolates showed fluctuating ESBL-detection rates in summer (10.5%), Fall (5.5%) and winter (7.7%), while K. pneumoniae showed high ESBL prevalence in winter (33.3%) compared to summer (6.25%) and but absent in fall. Farm worker isolates demonstrated seasonal differences, with ESBL-producing E. coli identified in summer (23.08%) and fall (21.0%) but absent in winter. Human fecal samples from hospitalized patients also showed variation, with E. coli ESBL prevalence of 42.8% in summer, rising to 60% in fall, but decreasing to 12.5% in winter. Overall, ESBLs production in E. coli was comparable in summer (14.68%) and fall (15%) before declining in winter (7.5%), while in K. pneumoniae, the highest prevalence was in winter (29.4%) compared to lower detection in summer (11.9%) and complete absence in fall. Statistical analysis indicated significant seasonal differences in E. coli prevalence between sources in the fall (p = 0.039), whereas K. pneumoniae variation was significant in poultry across seasons (p = 0.042).
Seasonal prevalence of phenotypically and genotypically ESBL-producing Escherichia coli and Klebsiella pneumoniae in poultry, dairy cows, environment, and human. Statistical significance was determined at p < 0.05.
Antibiotic resistance-pattern of ESBL-producing Escherichia coli and Klebsiella pneumoniae
3.4
All the ESBL-isolates from poultry, dairy cows, environment and human were tested against 8 antibiotics including TE, CT, MEM, AM, CIP, C, AMC, and SXT. The result of E. coli isolates across seasons highlighted the highest and most consistent resistance with AM (100%) in poultry and human isolates during summer and fall, persisting in winter with poultry. CT resistance was also very high in summer, reaching 100% in humans, environmental, and poultry isolates. In fall, resistance remained at 100% in both human and environmental isolates, but declined in winter to 33% in environmental isolates. MEM resistance increased in fall (57.1% in humans and 100% in poultry) but declined in winter (50% in poultry and absent in the environment). CIP resistance peaked in fall (100%) among human, poultry and environment but ranged from 66.6% in human to 100% in dairy during summer but dropped to 0% in winter human isolates. STX resistance was moderate to high in summer (50–100%) and ranged from low to high (12.5–100%) in fall but declined in winter, with environmental isolates fully susceptible. Overall, resistance levels were greatest in fall, especially in human and poultry isolates, whereas winter isolates, particularly from the environment, displayed comparatively lower resistance (Figure 4A and Supplementary Tables S3, S5).
Antibiotic resistance patterns of (A) ESBL-Escherichia coli and (B) ESBL-Klebsiella pneumoniae across hosts and seasons against eight different antibiotics: oxytetracycline (TE), Colistin (CT), meropenem (MEM), ampicillin (AM), ciprofloxacin (CIP), chloramphenicol (C), amoxicillin/clavulanate (AMC), and sulfamethoxazole/trimethoprim (SXT).
Antibiotic resistance profile of ESBL- producing K. pneumoniae from dairy and environment displayed complete resistance to most antibiotics in summer, with the dairy isolate resistant to all agents except C, while the environmental isolates were susceptible only to MEM. Human isolates of summer exhibited 100% resistance to AM, C, TE, CT and CIP, with lower resistance to MEM (33.3%), AMC (66.6%), and SXT (33.3%). In winter, Poultry isolates demonstrated high resistance (50–100%) to most agents. Environmental isolates showed resistance ranged from 33.3 to 66.6%, where resistance was absent (0%) in MEM, AMC, CIP, and SXT Overall, although both summer and winter isolates displayed extensive resistance, winter isolates particularly from environment showed relatively greater susceptibility compared to the uniformly resistant summer isolates (Figure 4B and Supplementary Tables S6, S7).
Distribution of ESBL-genes among isolates in relation to antibiotic profiling
3.5
A total of 56 ESBL-producing isolates of E. coli and K. pneumoniae were screened for the presence of four β-lactamase genes (blaTEM, blaSHV, blaCTX-M1, and blaCTX-M9) and thirteen antibiotic resistance phenotypes (CAZ, CRO, CTX, CEF, ATM, MER, AMC, AM, TE, C, CIP, CT, and SXT). The hierarchical clustering heatmap (Figure 5) revealed distinct distribution patterns of ESBL genes in relation to resistance against a panel of antibiotics.
Distribution of ESBL genes (blaTEM, blaSHV, blaCTX-M1, blaCTX-M9) among 56 ESBL-producing Escherichia coli (Mendonça et al., 2007) and Klebsiella pneumoniae (Badr et al., 2022) in relation to antibiotic resistance profiles. Hierarchical clustering separated the isolates into six major clusters (I: VI), reflecting distinct host-associated resistance and gene distribution patterns.
Overall, the majority of isolates harbored at least one ESBL gene, with blaTEM (51.8%) being the most frequently detected, followed by blaCTX-M1 (32.1%), whereas blaSHV (26.8%) is less common and there is no blaCTX-M9 detected. A considerable proportion of isolates carried multiple ESBL genes simultaneously.
Phenotypically, the majority of isolates were resistant to third-generation cephalosporins, with the highest resistance against CTX (89.3%) and CAZ (69.6%). Very high resistance rates were also observed for AM (87.5%), TE (71.4%), and CT (80.3). Resistance to CIP and C was moderate (both 57.1%) followed by AMC (53.5%), SXT (46.4%), and ATM (42.8%). In contrast, carbapenem resistance (meropenem) was the lowest (26.8%), indicating that carbapenems remain the most effective antibiotics against these isolates.
Hierarchical clustering divided the 56 ESBL-producing E. coli and K. pneumoniae isolates into six distinct clusters, with clear sub-structuring by host origin, gene carriage, and antibiotic resistance profiles. Cluster I, dominated by dairy cow isolates (n = 5) along with two environmental isolates, showed the lowest resistance levels and primarily carried blaTEM. Cluster II consisted mainly of environmental isolates (n = 4) and mixed-source isolates (n = 7; including three from dairy cows, and two each from human and poultry). This cluster displayed heterogeneous but generally moderate resistance phenotypes, with the majority harboring blaTEM, while only three strains carried blaSHV. Cluster III, associated with human (n = 3) and environmental (n = 2) isolates, exhibited the highest levels of multidrug resistance and frequent co-carriage of blaCTX-M1. Cluster IV comprised mainly poultry (n = 5) and human (n = 4) isolates, with additional contributions from dairy cows (n = 2) and the environment (n = 2). This cluster showed high resistance profiles and was predominantly associated with blaCTX. Cluster V, dominated by human isolates (n = 11) along with two poultry and one environmental isolate, demonstrated high resistance levels, with most carrying blaSHV and only three harboring blaTEM. Finally, Cluster VI, composed mainly of dairy cow isolates (n = 3) and one isolate each from human, poultry, and environment, displayed generally moderate resistance profiles, with all members carrying blaTEM. These findings underscore the host- and environment-specific dynamics of ESBL gene dissemination, with clinical isolates serving as primary reservoirs of multidrug resistance, while animal and environmental isolates contribute to resistance gene circulation and spillover.
Correlation analysis of ESBL genes and antibiotic resistance profiles
3.6
Correlation analysis between resistance genes (blaTEM, blaSHV, blaCTX-M1) and antibiotic susceptibility profiles revealed several statistically significant associations (Figure 6 and Supplementary Table S8).
Correlation analysis of ESBL genes and antibiotic resistance profiles among 56 ESBL-producing Escherichia coli and Klebsiella pneumoniae isolates. Positive and negative correlations are denoted by the red and blue color boxes, respectively. The correlation coefficient’s (r) numerical value is equivalent to the color’s intensity. Boxes with non-significant correlations were left empty, and correlations were calculated at p < 0.05.
A strong negative correlation was observed between blaTEM and blaSHV (r = −0.546, p = 1.34 × 10^−5^), indicating that the presence of blaTEM is inversely associated with blaSHV carriage. Similarly, blaTEM showed moderate negative correlations with blaCTX-M1 (r = −0.331, p = 0.013), CAZ (r = −0.310, p = 0.020), and CRO (r = −0.313, p = 0.019). In contrast, blaSHV demonstrated positive correlations with several β-lactams, including CRO (r = 0.490, p = 1.26 × 10^−4^), CAZ (r = 0.305, p = 0.022), and CEF (r = 0.359, p = 0.0066). Notably, blaCTX-M1 was positively correlated with SXT (r = 0.516, p = 4.7 × 10^−5^), suggesting a potential co-selection mechanism.
Among antibiotics, the strongest associations were observed between cephalosporins themselves. For example, CAZ was strongly correlated with CRO (r = 0.593, p = 1.45 × 10^−6^), CTX (r = 0.504, p = 7.36 × 10^−5^), and CEF (r = 0.658, p = 3.58 × 10^−8^). Likewise, AM resistance was very strongly associated with CAZ (r = 0.740, p = 7.11 × 10^−11^). Interestingly, resistance of quinolones (CIP) also showed significant positive correlations with multiple β-lactams (e.g., CAZ: r = 0.480, p = 1.80 × 10^−4^; CEF: r = 0.385, p = 0.0034), highlighting possible cross-resistance or co-resistance phenomena.
Discussion
4
The dissemination of E. coli and K. pneumoniae across the animal–human–environment interface represents a major One Health challenge, particularly given their ability to acquire and disseminate resistance traits such as ESBLs. Understanding the baseline prevalence of these organisms and the seasonal factors influencing their distribution is essential for assessing the risk of antimicrobial-resistance spread. The present study recovered in total 385 E. coli and 68 K. pneumoniae isolates from poultry, dairy cows, farm environment, farm workers, and hospitalized patients to assess seasonal impact on prevalence of ESBL-traits.
First, the prevalence analysis of isolates showed clear seasonal differences in dairy cattle with higher rates detected in fall compared to winter. This may be attributed to changes in temperature, humidity, and farm management practices that favor bacterial survival and transmission (Balta et al., 2024). Similarly, seasonal fluctuation was observed in farm environment, with higher recovery rates in summer and winter, reflecting the influence of environmental stressors and water contamination on bacterial persistence. In contrast, farm workers consistently carried a high prevalence of E. coli across seasons, indicating persistent occupational exposure and highlighting the role of humans as reservoirs and potential disseminators within the farm setting. This is greatly aligned with a study in Scotland, which reported that E. coli shedding in cattle is seasonally associated with warmer months and reflecting the seasonality of human infections (Ogden et al., 2004).
K. pneumoniae prevalence followed a distinct pattern, showing generally lower rates across all samples from different host groups but with significant variation in environmental samples and among dairy cows. This highlights the combined influence of both season and source on its occurrence. These findings are consistent with a review study from Germany, which emphasized the environmental health impact of K. pneumoniae, noting its significant resistance to disinfectants and frequent detection in farm-related environmental matrices such as water and sewage (Wareth and Neubauer, 2021). It has been also recently demonstrated that food chain-animals serve as a major reservoir for E. coli, while K. pneumoniae is less prevalent but more commonly found in environmental samples (Tsitsos et al., 2025).
Interestingly, hospitalized patients did not exhibit clear seasonal trends. This is likely reflecting their continuous exposure to hospital-associated risk factors such as close contact with other carriers rather than external environmental conditions. Similarly, poultry isolates remained relatively stable across seasons which may be driven by the hygiene and biosafety measures and antibiotics use.
Our findings in general revealed source-specific and seasonal differences in bacterial distributions, suggesting that both environmental and management factors contribute to shaping the prevalence of E. coli and K. pneumoniae organisms. This provides a baseline for subsequent ESBL phenotype and genotype characterization.
Phenotypic and genotypic ESBL analysis revealed notable seasonal and host-associated variations in the prevalence of ESBL-producing E. coli and K. pneumoniae. In poultry, ESBL- E. coli remained relatively stable between summer and fall but slightly increased in winter, whereas K. pneumoniae showed a marked seasonal fluctuation, being absent in fall but reaching its highest prevalence in winter. In dairy cows, ESBL-producing E. coli were observed in summer and fall but disappeared in winter, suggesting a possible link to management practices or antimicrobial use during warmer months.
Environmental isolates showed seasonal fluctuating in ESBL detection with K. pneumoniae more prevalence in winter. Contaminated environments are rich with commensals such as E. coli and other Enterobacteriaceae, which often carry plasmids, integrons, and transposons that facilitate resistance genes mobilization. Seasonal changes in temperature and humidity may act as stressors that support bacterial persistence and promote genetic exchange. Such ecological pressures, combined with excessive input of antibiotic residues from farming practices, likely create hotspots that facilitate the transfer of ESBL genes among bacterial populations (Larsson and Flach, 2022). This was highlighted in number of studies demonstrated that farm animals and their surrounding environments served as reservoirs to ESBL-producers (Kamaruzzaman et al., 2020) and (Badr et al., 2022). Similarly, environmental sources have been identified as significant contributors to the dissemination of ESBLs in sub-Saharan Africa, especially among E. coli isolates (Olaitan et al., 2025). Comparable observations have been reported in recent studies from Brazil (Requena-Castro et al., 2024) and New Zealand (Gray et al., 2025), where surface waters were considered major hotspots for ESBL-producing bacteria.
In farm workers, ESBL- producing E. coli prevalence peaked in summer, followed by a slight decrease in fall and complete absence in winter. This seasonal pattern along with the observed increase in the farm environment, suggests a potential risk of zoonotic transmission during warmer months through direct contact, contaminated surfaces, airborne routes, vectors such as flies, and environmental sources like slurry and dairy farm effluents (Dahms et al., 2015). Notably, hospitalized patients exhibited the highest prevalence in fall and summer compared to winter, highlighting the potential for seasonal amplification of resistant strains in clinical settings. Comparably, seasonal and high incidence of ESBL- E. coli and Klebsiella species among hospitalized patients in German was reported in warmer months (Kaier et al., 2010). Another study also highlighted the seasonal variations of ESBL-carriage among the general Dutch population, which were associated with the warmer months (Wielders et al., 2020).
These results corroborate the influence of seasonal and ecological factors on the dissemination of ESBL genes, with E. coli predominating in fall across most sources, while K. pneumoniae was more frequently associated with winter, particularly in poultry and environmental samples. This is consistent with the observations of Pak and King (2022), who demonstrated that low temperatures and wind contribute to increased antibiotic resistance and the environmental spread of resistant microbes. A German study also identified environmental and anthropogenic factors particularly seasonal variations, extreme weather events, and cattle density as significant determinants influencing the occurrence of ESBL/AmpC-producing E. coli in wildlife (Günther et al., 2022). The complete absence of ESBL-producing K. pneumoniae in fall among the tested hosts may be attributed to seasonal climatic conditions. In particular, local temperature can strongly influence the persistence of ESBL-carrying plasmids in the environment and affect how efficiently these plasmids are transferred between bacteria. Plasmid stability varies not only between bacterial species but also between strains of the same species, and temperature can influence this stability. For example, Yang et al. reported that some plasmids (such as blaKPC-IncF and blaNDM-IncX3) were most stable at 25–30 °C and became less stable at 37 °C (Yang et al., 2024). Other experimental work in E. coli has shown that plasmid loss is lower at 20 °C than at 37 °C (Wein et al., 2019). In addition, some conditions may limit the spread of ESBL plasmids. Haweky et al. surveyed ESBL strain and plasmid transmission over one year in a hospital setting and found that ESBL plasmid transmission events were generally rare (Hawkey et al., 2022).
The distribution of ESBL genes among E. coli and K. pneumoniae isolates demonstrated clear seasonal variation by host and environments in current one health setting, although some resistance traits remained stable. In summer, E. coli isolates from poultry, dairy, and the environment were dominated by blaTEM (100, 75, and 50%, respectively), a pattern that persisted into the fall where poultry and dairy isolates remained universally positive (100%). Contrarily, human E. coli showed no blaTEM in summer but displayed substantial carriage in fall (42.85%), together with blaSHV as the most frequent gene (57.1%). Environmental E. coli also shifted between seasons, harboring blaCTX-M1 predominantly in fall after being associated with both blaTEM (50%) and blaCTX-M group 1 (50%) in summer. Winter presented a distinct profile, with blaTEM disappearing from E. coli across all sources but emerging in K. pneumoniae from poultry (50%) and environment (100%). At the same time, E. coli in winter became enriched by blaSHV (poultry 100%, humans 100%, environment 66.6%) while blaCTX-M group 1 was confined to environmental E. coli (33.3%) and poultry K. pneumoniae (50%). Comparably, a study conducted in El-Sharkia Governorate between December 2019 and April 2021 demonstrated that blaTEM and blaSHV were the most frequently detected ESBL genes in poultry (Salem et al., 2023). The seasonal occurrence of blaSHV, particularly its predominance in humans and its fluctuating detection in poultry and environmental isolates during winter and fall, mirrors its previously reported dominance across multiple reservoirs including humans, livestock, and poultry in northern Egypt (Nossair et al., 2022). In the contrary, blaCTX-M group 9 was consistently absent across all three seasons, suggesting limited introduction or persistence of this lineage in the studied ecosystem.
These patterns indicate that blaTEM is a stable trait in poultry E. coli, while both blaSHV and blaCTX-M group 1 fluctuate seasonally across hosts and niches, possibly reflecting differences in antimicrobial usage, environmental pressures, and bacterial adaptability. Likewise, a study on Dutch poultry farm environments showed that the vast majority of E. coli isolates harbored blaCTX-M-1, blaSHV, and blaTEM (Blaak et al., 2015) Additionally, a meta-analysis from sub-Saharan Africa, which included subgroup analysis by sample source, identified animals as the predominant reservoir of ESBL-producing E. coli, with blaCTX-M reported as the most prevalent ESBL gene (Olaitan et al., 2025).
Hierarchical analysis of ESBL-gene distribution and antibiotic-resistance profiles showed clustering of isolates belonging to different hosts. The results showed that blaTEM tends to be inversely associated with other ESBL genes (blaSHV, blaCTX-M1) and certain cephalosporins, whereas blaSHV and blaCTX-M1 are significantly linked with cephalosporins and other non-β-lactam antibiotics. The clustering of correlations among cephalosporins indicates that resistance to these agents may be strongly co-selected, possibly reflecting the underlying genetic determinants of ESBL production.
The association of ESBL genes with resistance to antibiotic classes not inhibited by ESBL genes has previously been demonstrated in several studies (Badr et al., 2022; Nossair et al., 2022; Yousfi et al., 2016). Our data reported a strong correlation between blaCTX-M1and SXT resistance which suggests cross resistance and co-location of multiple resistance traits on mobile genetic elements. The sul1 gene, which confers resistance to sulfonamides, is commonly carried within class 1 integrons, genetic elements known for capturing and expressing multiple resistance gene cassettes (Jiang et al., 2019). Detection of mobile genetic elements such as integrons in ESBL- isolates from feces of farm and domestic animals has been linked to phenomenon of extensive resistance to non- cephalosporin antibiotics (Ejaz et al., 2021). Several studies have shown that class 1 integrons containing sul1 can occur on the same mobile genetic elements such as ISCR1-associated transposons that also harbor blaCTX-M genes (Mendonça et al., 2007; Shahid et al., 2012). This close physical linkage means that the resistance determinants can move together on a single plasmid or integron. As a result, the observed co-resistance pattern likely reflects a shared genetic background that enables both traits to be maintained and transmitted simultaneously.
This finding and previous studies identified ESBL-producers as a One Health threat both as pathogen and disseminators to multi-resistance traits (Tello et al., 2022). ESBL-producing isolates showed multidrug-resistant patterns, including resistance to colistin, which is a last-resort antibiotic. Several studies have also reported a link between ESBL production and colistin resistance in E. coli and K. pneumoniae(Lay et al., 2021; Hide et al., 2024). For example, Trongjit et al. (2022) found that ESBL-producing bacteria from pigs could transfer both bla genes and the colistin-resistance gene mcr-3 to Salmonella. This suggests that some strains may carry both resistance traits together (Trongjit et al., 2022).
A key limitation of this study is the lack of seasonal antimicrobial use data, which may have influenced the observed variations in ESBL prevalence. Additionally, not all known ESBL genes were tested, so the presence of other resistance determinants cannot be excluded. In a future study, whole genome sequencing of the isolates will be conducted to provide deeper molecular insights and clarify the roles of gene mobilization, plasmids, and integrons.
Conclusion
5
This study investigated seasonal influence on the prevalence and dissemination of ESBL-producing E. coli and K. pneumoniae within a One Health context. The results revealed a notable seasonal pattern associated with both animal and human hosts, as well as with specific bacterial species, while also highlighting the farm environment as an important reservoir for ESBL producers. Furthermore, a correlation was observed between harboring ESBL genes and resistance to non-β-lactam antibiotics, rather than to β-lactams themselves. These findings highlight the significance of ESBL-producing bacteria as both pathogens and carriers, driving bacterial evolution while promoting cross-resistance and gene mobilization within shared ecological niches. This underscores the need for seasonally tailored antimicrobial interventions and integrated One Health surveillance, including enhanced hygiene measures during high-risk seasons and closer monitoring of plasmid-carrying strains to limit the spread of ESBL genes.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Badr H. Reda R. M. Hagag N. M. Kamel E. Elnomrosy S. M. Mansour A. I. . (2022). Multidrug-resistant and genetic characterization of extended-spectrum beta-lactamase-producing E. coli recovered from chickens and humans in Egypt. Animals 12:346. doi: 10.3390/ani 12030346, 35158668 PMC 8833359 · doi ↗ · pubmed ↗
- 2Balta I. Lemon J. Murnane C. Pet I. Vintila T. Mc Cleery D. . (2024). The one health aspect of climate events with impact on foodborne pathogens transmission. One Health 19:100926. doi: 10.1016/j.onehlt.2024.100926, 39559751 PMC 11570983 · doi ↗ · pubmed ↗
- 3Bauer A. W. Kirby W. M. Sherris J. C. Turck M. (1966). Antibiotic susceptibility testing by a standardized single disk method. Am J Clin Pathol.; 45:493–6., 5325707 · pubmed ↗
- 4Blaak H. van Hoek A. H. Hamidjaja R. A. van der Plaats R. Q. Kerkhof-de Heer L. de Roda Husman A. M. . (2015). Distribution, numbers, and diversity of ESBL-producing E. coli in the poultry farm environment. P Lo S One 10:e 0135402. doi: 10.1371/journal.pone.0135402, 26270644 PMC 4536194 · doi ↗ · pubmed ↗
- 5Bubpamala J. Khuntayaporn P. Thirapanmethee K. Montakantikul P. Santanirand P. Chomnawang M. T. (2018). Phenotypic and genotypic characterizations of extended-spectrum beta-lactamase-producing Escherichia coli in Thailand. Infect. Drug Resist. 11, 2151–2157. doi: 10.2147/IDR.S 174506, 30464558 PMC 6223337 · doi ↗ · pubmed ↗
- 6Dahms C. Hübner N.-O. Kossow A. Mellmann A. Dittmann K. Kramer A. (2015). Occurrence of ESBL-producing Escherichia coli in livestock and farm workers in Mecklenburg-Western Pomerania, Germany. P Lo S One 10:e 0143326. doi: 10.1371/journal.pone.0143326, 26606146 PMC 4659621 · doi ↗ · pubmed ↗
- 7Dierikx C. M. van der Goot J. A. Smith H. E. Kant A. Mevius D. J. (2013). Presence of ESBL/Amp C-producing Escherichia coli in the broiler production pyramid: a descriptive study. P Lo S One 8:e 79005. doi: 10.1371/journal.pone.0079005, 24244401 PMC 3820706 · doi ↗ · pubmed ↗
- 8Ejaz H. Younas S. Abosalif K. O. Junaid K. Alzahrani B. Alsrhani A. . (2021). Molecular analysis of Bla SHV, bla TEM, and Bla CTX-M in extended-spectrum β-lactamase producing Enterobacteriaceae recovered from fecal specimens of animals. P Lo S One 16:e 0245126. doi: 10.1371/journal.pone.0245126, 33412564 PMC 7790543 · doi ↗ · pubmed ↗