Progesterone Enhances the Sensitivity of Ovarian Cancer Cells to Poly (ADP-Ribose) Polymerase (PARP) Inhibitors by Suggesting a Role for Transcription-Replication Conflict-Related Pathways: An In Vitro Study
Eri Suizu, Takahiro Koyanagi, Yasushi Saga, Yoshifumi Takahashi, Kohei Tamura, Akiyo Taneichi, Yuji Takei, Hiroaki Mizukami, Hiroyuki Fujiwara

TL;DR
This study shows that progesterone makes ovarian cancer cells more sensitive to PARP inhibitors by affecting pathways related to transcription-replication conflicts.
Contribution
The study reveals a novel mechanism by which progesterone enhances PARP inhibitor efficacy through non-genomic actions and TRC-related pathways.
Findings
Progesterone significantly increased the sensitivity of ovarian cancer cells to PARP inhibitors.
Progesterone reduced the expression of TRC-protective factors like PARP1/2/3 and TOPO-I.
The effect of progesterone was reversed by DRB, suggesting a TRC-related mechanism.
Abstract
Objective: Ovarian cancer is often diagnosed at an advanced stage with peritoneal dissemination and ascites. Despite initial chemosensitivity, most patients eventually relapse. Poly (ADP-ribose) polymerase (PARP) inhibitors have become important maintenance therapies, particularly for tumors with homologous recombination deficiencies. Transcription-replication conflicts (TRCs) are increasingly recognized as a key mechanism related to PARP inhibitor-induced cytotoxicity. Progesterone exerts rapid non-genomic effects via membrane progesterone receptors (mPRs), suppresses topoisomerase I (TOPO-I), and enhances irinotecan cytotoxicity in ovarian cancer cells. We hypothesized that combining progesterone with PARP inhibitors could enhance antitumor effects by modulating TRC-protective pathways. Methods: The BRCA1/2 wild-type ovarian cancer cell line SHIN-3 (PR-negative and mPR-positive),…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
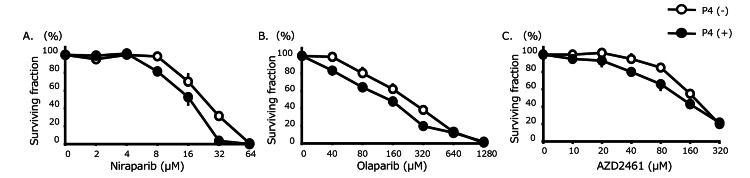 Figure 1
Figure 1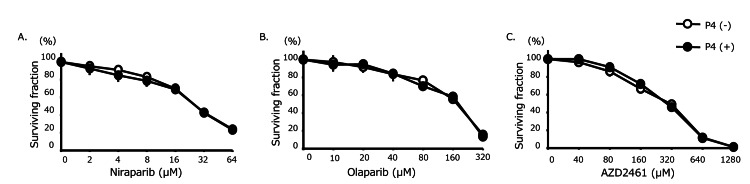 Figure 2
Figure 2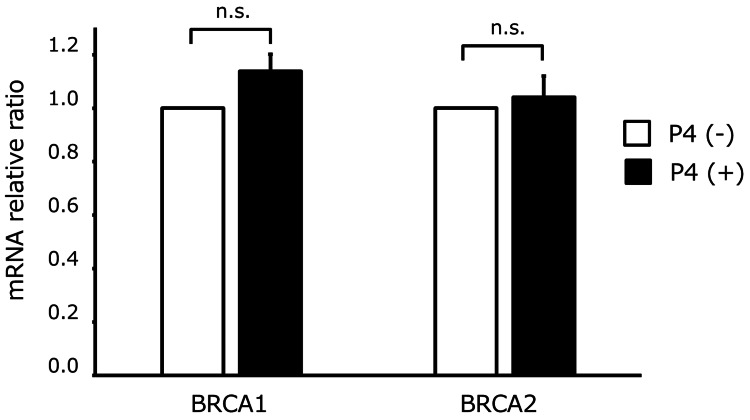 Figure 3
Figure 3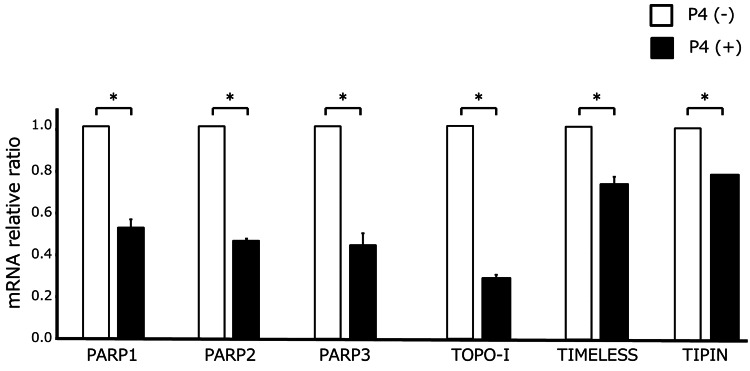 Figure 4
Figure 4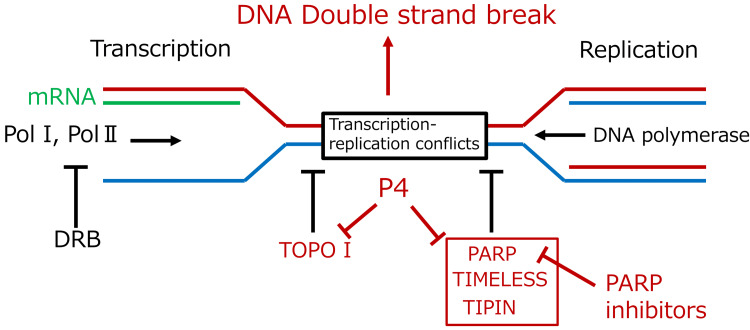 Figure 5
Figure 5| Gene | Accession number | Primer sequences (5'-3') |
| BRCA1 (F) | CTGAAGACTGCTCAGGGCTATC | |
| BRCA1 (R) | AGGGTAGCTGTTAGAAGGCTGG | |
| BRCA2 (F) | GGCTTCAAAAAGCACTCCAGATG | |
| BRCA2 (R) | GGATTCTGTATCTCTTGACGTTCC | |
| PARP1 (F) | CCAAGCCAGTTCAGGACCTCAT | |
| PARP1 (R) | GGATCTGCCTTTTGCTCAGCTTC | |
| PARP2 (F) | GGTGGCTTGTTCAGGCAATCTC | |
| PARP2 (R) | GGTGGCATAGTCCATCTGTAGC | |
| PARP3 (F) | TCTCTGAGCAGGAGAAGACGGT | |
| PARP3 (R) | TGTGGTTGCTGCCAGTCTGTTC | |
| TOPOI (F) | GAACAAGCAGCCCGAGGATGAT | |
| TOPO I (R) | TGCTGTAGCGTGATGGAGGCAT | |
| TIMELESS (F) | AAGTGGTCCAGGTGTCGGAGAA | |
| TIMELESS (R) | GTGGGCACTATTCTGCTGGTAG | |
| TIPIN (F) | CCAGAGAGACAAGATGGTGAAGG | |
| TIPIN (R) | CTCTGAGCATCCAGCTTGGGTA | |
| GAPDH (F) | ACCACAGTCCATGCCATCAC | |
| GAPDH (R) | CATCACGCCACAGTTTCCCG |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPARP inhibition in cancer therapy · Ovarian cancer diagnosis and treatment · Estrogen and related hormone effects
Introduction
Ovarian cancer ranks sixth among the causes of cancer-related mortality in women in the United States, with approximately 21,000 new cases and 13,000 deaths reported annually [1]. Because early-stage ovarian cancer usually presents without symptoms, over half of patients are diagnosed at an advanced stage with peritoneal dissemination and ascites. The prognosis of ovarian cancer is highly stage-dependent, with five-year survival rates of approximately 70-91% in localized or regional diseases, but only approximately 31% in cases with distant metastasis [2]. The standard treatment for advanced ovarian cancer includes cytoreductive surgery, followed by platinum-based combination chemotherapy. Although ovarian cancer is initially highly sensitive to chemotherapy, many patients achieve remission with multimodal therapy. Approximately 80% of patients achieve complete remission after primary treatment. However, more than half of these cases eventually experience relapse. Recurrent disease is often associated with reduced chemosensitivity, ultimately leading to death from disease progression [3,4]. These findings highlight the need for effective maintenance therapies.
To address this issue, molecular targeted agents, such as bevacizumab and poly (ADP-ribose) polymerase (PARP) inhibitors, have been widely used as maintenance therapies [5]. Large-scale clinical trials have demonstrated that PARP inhibitors, such as olaparib and niraparib, significantly prolong progression-free survival when used as maintenance therapy in patients with advanced ovarian cancer [6,7]. Their therapeutic benefits are particularly evident in patients with BRCA mutations or homologous recombination deficiencies (HRD), although broader patient populations also derive benefit. PARP inhibitors were initially thought to exert cytotoxic effects through PARP trapping in DNA, which blocks replication fork progression and leads to DNA double-strand breaks [8]. There have been no reports to date that show a relationship between progesterone and these mechanisms. More recently, transcription-replication conflicts (TRCs) have been reported as another key mechanism underlying their efficacy [9]. PARP-1 is a key factor in preventing TRCs, and its inhibition increases TRC formation, ultimately resulting in DNA double-strand breaks. Other factors involved in TRC prevention include topoisomerase I (TOPO-I), TIMELESS, and TIMELESS-interacting protein (TIPIN) [10].
Progesterone is a steroid hormone mainly produced by the corpus luteum and binds to intracellular progesterone receptors to form a regulatory complex that interacts with nuclear DNA promoter regions to modulate gene expression [11]. Clinically, it is used to treat hormone-sensitive cancers, such as endometrial and breast cancers [12,13], but the detailed mechanism of tumor suppression remains unclear.
Recently, membrane progesterone receptors (mPRs) have been identified on cell membranes [14]. Unlike nuclear receptors, which act through slower transcription-dependent mechanisms, mPRs mediate the rapid, nongenomic effects of progesterone via membrane-associated pathways [15]. We previously reported that progesterone induces rapid cell death in ovarian cancer cells through non-genomic mechanisms within 30 min at 10-400 μM [16]. More recently, we showed that progesterone suppresses TOPO-I expression and markedly enhances the cytotoxicity of irinotecan, a TOPO-I-targeting chemotherapeutic agent [17].
In this in vitro study, our primary objective was to determine whether progesterone alters the sensitivity of ovarian cancer cells to PARP inhibitors, as assessed by changes in IC₅₀ values. We further hypothesized that progesterone increases sensitivity to PARP inhibitors through TRC-related pathways and explored this indirectly using gene expression analysis and transcriptional inhibition with DRB. Secondary objectives included evaluating changes in the expression of PARP1/2/3, TOPO-I, TIMELESS, and TIPIN, and examining the effect of transcriptional inhibition on progesterone-mediated sensitization. These experiments were performed exclusively in vitro using a single BRCA-wild-type, PR-negative, and mPR-positive ovarian cancer cell line (SHIN-3) and therefore represent an exploratory, hypothesis-generating investigation.
Materials and methods
Cell line and culture
The human ovarian serous adenocarcinoma cell line SHIN-3 [18], which harbors wild-type BRCA1/2 [19], was obtained from the manufacturer. In our previous study, these cells were confirmed to lack progesterone receptor (PR) expression, but to express various mPRs [16]. Cells were cultured in Dulbecco’s Modified Eagle Medium/F12 (DMEM/F12; Thermo Fisher Scientific, Inc., Waltham, MA) supplemented with 10% heat-inactivated fetal bovine serum (FBS; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and 1% penicillin/streptomycin (Thermo Fisher Scientific, Inc.) at 37 °C under 5% CO₂. The number of cells at the time of receipt was unknown; however, all experiments were performed within 20 passages from the original stock. Cells were confirmed to be mycoplasma-negative at the time of receipt. To minimize variability in drug response, the same lot of FBS was used throughout the study. Cells were routinely passaged at approximately 80% confluency.
Colorimetric assay
Tumor cells (1 × 10³ cells/well) were seeded into a 96-well plate and treated with progesterone (P4; FUJIFILM Wako Pure Chemical Co., Osaka, Japan) at 0 or 100 μM [16], with or without 50 μM of 5,6-dichloro-1-β-D-ribofuranosylbenzimidazole (DRB), an RNA polymerase II inhibitor [20], for 30 min. These drugs were prepared in DMSO, with the final concentration of DMSO not exceeding 0.5%. Following P4 and/or DRB treatment, the medium was removed and replaced to prevent direct interference between P4 and the subsequent PARP inhibitors. Tumor cells were then exposed to niraparib (Selleck Biotech, Tokyo, Japan) at concentrations of 2-64 μM for 72 hours, or to olaparib (Selleck Biotech) at concentrations of 40-1,280 μM for 72 hours, or to AZD2461, a PARP1/2/3 inhibitor (Selleck Biotech), at concentrations of 10-320 μM for 72 hours. PARP inhibitors were continuously present for 72 hours without medium exchange during this period. Stock solutions of P4 were prepared in DMSO at 70 mM and stored at 4 °C protected from light. Stock solutions of niraparib and olaparib were prepared in DMSO at 50 mM, and AZD2461 at 10 mM, and stored at 4 °C protected from light. The viable cell count, measured by a colorimetric assay using the Premix WST-1 Cell Proliferation Assay System (Takara Bio Inc., Tokyo, Japan), is presented as a percentage relative to the control untreated with niraparib, olaparib, or AZD2461. WST-1 reagent was incubated for 24 hours before measurement, and absorbance was read at 450 nm using a microplate reader. Blank correction was performed. Each experiment was performed in triplicate and independently repeated at least three times. IC₅₀ values were calculated as the mean of these independent dose-response experiments. No specific measures were taken to control for edge effects; however, no apparent edge-related bias was observed in preliminary experiments.
RT-qPCR
Tumor cells (5 × 10⁵ cells/well) seeded into a six-well plate were exposed to progesterone at a concentration of 400 μM for 30 minutes. These conditions were the same as in previous experiments [17]. Cellular mRNA was extracted using a RNeasy Mini Kit (Qiagen, Valencia, CA) according to the manufacturer’s instructions. RT-qPCR was performed using a Thermal Cycler Dice Real-Time System II (Takara Bio, Inc.) with the One Step TB GreenR PrimeScript™ PLUS RT-PCR Kit (Takara Bio, Inc.), following the manufacturer’s instructions. The PCR was carried out using 40 cycles of heating at 95 °C for 15 seconds, 58 °C for 15 seconds, and 72 °C for 20 seconds. Melt-curve analysis confirmed single amplicons. mRNA levels were determined relative to the fluorescence signal of glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The primer sequences are listed in Table 1. Each assay was performed with three technical replicates, and experiments were independently repeated at least three times. GAPDH Ct values did not differ significantly between control and progesterone-treated cells, confirming its stability under the experimental conditions.
Statistical analysis
Statistical analyses were performed using EZR software (Saitama Medical Center, Jichi Medical University, Saitama, Japan). Since all results were two-group comparisons, two-tailed Student's t-tests were used. p-values < 0.01 were considered statistically significant. All experiments were performed using independent biological replicates. Prior to t-test application, equality of variances was confirmed using an F-test. Because all analyses involved two-group comparisons, no correction for multiple testing was applied.
Results
Chemosensitivity
To determine whether progesterone enhances the sensitivity of ovarian cancer cells to PARP inhibitors, we examined the effects of progesterone on SHIN-3 cells treated with different PARP inhibitors. As shown in Figure 1, the IC₅₀ for niraparib in SHIN-3 cells with progesterone was 16.8 ± 2.4 μM, which was significantly lower than that of SHIN-3 cells without progesterone (24.9 ± 1.7 μM) (p = 0.0080). Similarly, the IC₅₀ for olaparib in SHIN-3 cells with progesterone was 151 ± 9 μM, significantly lower than that of SHIN-3 cells without progesterone (237 ± 12 μM) (p = 0.0006). Moreover, the IC₅₀ for AZD2461 in SHIN-3 cells with progesterone was 152 ± 12 μM, again significantly lower than that of SHIN-3 cells without progesterone (195 ± 14 μM) (p = 0.0024).
Sensitivity to PARP inhibitorsThe IC50 for each PARP inhibitor in SHIN‐3 was as follows. Niraparib: With progesterone, 16.8 ± 2.4 μM versus without progesterone, 24.9 ± 1.7 μM (1.5‐fold higher sensitivity, p = 0.0080). Olaparib: With progesterone, 151 ± 9 μM versus without progesterone, 237 ± 12 μM (1.6‐fold higher sensitivity, p = 0.00006). AZD2461 (PARP1, 2, 3 inhibitors): With progesterone, 152 ± 12 μM versus without progesterone, 195 ± 14 μM (1.3‐fold higher sensitivity, p = 0.0024). Data are shown as means and SD (n = 3).P4, progesteron; PARP, poly (ADP-ribose) polymerase
To assess whether these effects depended on transcriptional activity, SHIN-3 cells were treated with DRB, a transcriptional inhibitor. As shown in Figure 2, no significant difference was observed in the IC₅₀ values for niraparib, olaparib, and AZD2461 in SHIN-3 cells with or without progesterone (27.7 ± 2.7 μM for niraparib with progesterone vs. 27.7 ± 0.5 μM without progesterone; 251 ± 19.3 μM for olaparib with progesterone vs. 255 ± 7.0 μM without progesterone; and 182 ± 9.0 μM for AZD2461 with progesterone vs. 189 ± 12.3 μM without progesterone).
Sensitivity to PARP inhibitors when combined with DRBThe IC50 for each PARP inhibitor in SHIN‐3 when combined with DRB were as follows. Niraparib: With progesterone, 27.7 ± 2.7 μM versus without progesterone, 27.7 ± 0.5 μM (not significant). Olaparib: With progesterone, 251 ± 19.3 μM versus without progesterone, 255 ± 7.0 μM (not significant). AZD2461 (PARP1, 2, 3 inhibitors): With progesterone, 182 ± 9.0 μM versus without progesterone, 189 ± 12.3 μM (not significant). Data are shown as means and SD (n = 3).P4, progesterone; DRB, 5, 6-Dichloro-1-β-D-ribofuranosyl-1H-benzimidazole; n.s., not significant; PARP, poly (ADP-ribose) polymerase
Gene expression
To explore the mechanisms underlying the enhanced sensitivity to PARP inhibitors, we investigated the effect of progesterone on gene expression in SHIN-3 cells. As shown in Figure 3, progesterone did not affect the expression of BRCA1 or BRCA2. However, as shown in Figure 4, progesterone reduced the expression of PARP1/2/3 (p = 0.0001), TOPO-I (p = 0.0001), TIMELESS (p = 0.0018), and TIPIN (p = 0.0001) in SHIN-3 cells. These are all mRNA results; protein validations are pending.
BRCA1 and BRCA2 gene expression after exposure to progesteroneThe exposure to progesterone did not affect BRCA1 and BRCA2 gene expression in SHIN-3 cell line (not significant). Data are shown as means and SD (n = 3). P4, progesterone; n.s., not significant
*Gene expression of TRC-protective factors after exposure to progesteroneProgesterone significantly reduced the expression of PARP1/2/3, TOPO-I, TIPIN, and TIMELESS in SHIN-3 cells. Data are shown as means and SD (n = 3).TRC, transcription-replication conflict; P4, progesterone; PARP, poly (ADP-ribose) polymerase; TOPO-I, topoisomerase I; TIPIN, TIMELESS-interacting protein. p < 0.01.
Discussion
This study investigated the combined effects of progesterone and various PARP inhibitors using the ovarian cancer cell line SHIN-3 to develop a novel therapeutic strategy. To clarify the mechanism by which progesterone enhances PARP inhibitor sensitivity, we evaluated BRCA1/2 expression, key genes strongly associated with HRD. Furthermore, to elucidate the mechanism of the combined effect, we investigated the relationship between TRC-protective factors and the enhancement of PARP inhibitor sensitivity by progesterone.
Progesterone was found to enhance the sensitivity of ovarian cancer cells to PARP inhibitors. In contrast, when combined with the TRC inhibitor DRB, an RNA polymerase II inhibitor, progesterone had no effect on PARP inhibitor sensitivity. Examination of BRCA1/2 gene expression, which strongly influences PARP inhibitor sensitivity, revealed that progesterone did not affect its expression. In contrast, progesterone significantly reduced the expression of PARP, TOPO-I, TIPIN, and TIMELESS, all of which prevented TRCs. These results suggest progesterone may promote TRC-related vulnerability to PARP inhibition (Figure 5); however, direct TRC measurements were not performed, and alternative mechanisms (e.g., effects on DNA repair gene expression or replication dynamics) cannot be excluded.
Schematic summary of the resultsProgesterone suppresses the expression of TRC-protective factors (PARP, TOPO-I, TIMELESS, TIPIN), inducing TRCs and subsequent DNA double-strand breaks. Combining progesterone and PARP inhibitors could show a synergistic antitumoral effect on PR-negative and mPR-positive ovarian cancer.TRC, transcription-replication conflict; P4, progesterone; PARP, poly (ADP-ribose) polymerase; TOPO-I, topoisomerase I; TIPIN, TIMELESS-interacting protein; Pol I, RNA polymerase I; Pol Ⅱ, RNA polymerase Ⅱ; DRB; 5, 6-dichloro-1-β-D-ribofuranosyl-1H-benzimidazole
We have previously shown that progesterone triggers rapid cell death in ovarian cancer cells in vitro [15]. Because all of the ovarian cancer cells were PR-negative and mPR-positive, and cell death occurred within 30 min, the effect was concluded to be likely mediated by non-genomic actions through mPRs, rather than nuclear PR-dependent genomic actions. Because the ovarian cancer cell line used in the present study, SHIN-3, is also PR-negative and mPR-positive, the observed effect of progesterone on enhancing PARP inhibitor sensitivity is thought to be mediated through non-genomic actions.
Sensitivity to PARP inhibitors is largely influenced by BRCA1 and BRCA2, which are responsible for repairing DNA double-stranded breaks. The ovarian cancer cell line SHIN-3 used in this study possessed wild-type BRCA1 and BRCA2 [19]. Examination of BRCA1/2 expression after treatment with progesterone revealed that progesterone did not affect its expression. Next, we focused on TRCs, a recently proposed novel mechanism of action of PARP inhibitors. Specifically, PARP inhibitors block the function of TRC-protective factors, such as PARP, TIMELESS, and TIPIN, thereby promoting TRCs and inducing DNA double-stranded breaks. TOPO-I has also been identified as a protective factor [21]. Analysis of the expression of these TRC-protective factors following progesterone treatment demonstrated that progesterone decreased their expression levels. Furthermore, when DRB, an RNA polymerase II inhibitor that suppresses transcription and thereby protects against TRCs, was used, progesterone did not enhance the sensitivity to PARP inhibitors.
The clinical application of PARP inhibitors in ovarian cancer treatment has begun mainly in the setting of maintenance therapy following conventional cytotoxic chemotherapy [7,22,23]. Because the prognosis for recurrent ovarian cancer is poor [24], augmentation of sensitivity to PARP inhibitors might contribute to an improved prognosis. Another challenge is the economic burden and specific side effects, particularly when treatment duration increases. Progesterone is inexpensive and clinically used in other contexts [25], making it an attractive candidate for repurposing, but pharmacokinetic, dosing, and safety studies in cancer settings are required. However, progesterone is rapidly metabolized in vivo by multiple hydroxylases produced in the liver [26]. Further studies are required to determine the appropriate dosage and administration method for progesterone.
This study has several limitations. All experiments were carried out using a single ovarian cancer cell line; therefore, findings obtained from one model may not be fully generalizable to the broad heterogeneity of ovarian cancer, and additional cell lines will be needed to confirm the reproducibility of these results. In addition, the study was conducted exclusively through in vitro assays, which preclude direct evaluation of biological complexity in living organisms. Furthermore, although several genes were identified as downregulated following progesterone treatment, protein-level validation for these changes was not performed. TRCs were not directly assessed because no dedicated assays were performed and their involvement was inferred only indirectly from downstream phenotypes. Finally, the involvement of mPR was not examined through perturbation experiments, leaving its precise contribution unresolved. Further studies will be required to clarify the underlying mechanisms and to determine the extent to which these findings may be relevant to future clinical applications.
Conclusions
Progesterone suppresses the expression of TRC-protective factors, including PARP, TIPIN, TIMELESS, and TOPO-I, through non-genomic actions mediated by mPRs. This suppression promotes TRCs and enhances the sensitivity of ovarian cancer cells to PARP inhibitors.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cancer statistics, 2025 CA Cancer J Clin Siegel RL Kratzer TB Giaquinto AN Sung H Jemal A 10457520253981767910.3322/caac.21871 PMC 11745215 · doi ↗ · pubmed ↗
- 2Cancer stat facts: ovarian cancer 1 2026 2025 https://seer.cancer.gov/statfacts/html/ovary.html
- 3Cancer of the ovary N Engl J Med Cannistra SA 2519252935120041559095410.1056/NEJ Mra 041842 · doi ↗ · pubmed ↗
- 4Recent advances in systemic treatments for ovarian cancer Cancer Imaging Banerjee S Gore M 3053091220122302288710.1102/1470-7330.2012.9002 PMC 3460563 · doi ↗ · pubmed ↗
- 5Safety of bevacizumab and olaparib as frontline maintenance therapy in advanced ovarian cancer: expert review for clinical practice Front Oncol Romero I Guerra E Madariaga A Manso L 13043031320243834812210.3389/fonc.2023.1304303 PMC 10859514 · doi ↗ · pubmed ↗
- 6Maintenance olaparib in patients with newly diagnosed advanced ovarian cancer N Engl J Med Moore K Colombo N Scambia G 2495250537920183034588410.1056/NEJ Moa 1810858 · doi ↗ · pubmed ↗
- 7Niraparib in patients with newly diagnosed advanced ovarian cancer N Engl J Med González-Martín A Pothuri B Vergote I 2391240238120193156279910.1056/NEJ Moa 1910962 · doi ↗ · pubmed ↗
- 8The underlying mechanism for the PARP and BRCA synthetic lethality: clearing up the misunderstandings Mol Oncol Helleday T 387393520112182147510.1016/j.molonc.2011.07.001PMC 5528309 · doi ↗ · pubmed ↗