Development and characterization of polymorphic microsatellite markers for Macrotermes carbonarius (Blattodea: Termitidae)
Li Lim, Kok Yean Von, Abdul Hafiz Ab Majid

TL;DR
This study develops and validates microsatellite markers for the termite Macrotermes carbonarius to better understand its genetic structure and population dynamics.
Contribution
The study introduces new polymorphic microsatellite markers specifically developed for Macrotermes carbonarius using next-generation sequencing.
Findings
Seven highly polymorphic microsatellite loci were validated with high heterozygosity and polymorphic information content.
Population genetic analyses revealed significant genetic differentiation and limited gene flow among colonies.
The markers are suitable for studying genetic diversity and colony structure in M. carbonarius.
Abstract
Macrotermes carbonarius is a fungus-growing termite widely distributed in Southeast Asia and is ecologically important in both natural and urban environments. In Malaysia, this species may act as a secondary pests following the suppression of primary termite infestations, making an understanding of colony structure and spatial connectivity particularly relevant. Microsatellite markers are widely used for such population-level studies; however, the performance and informativeness of loci developed for related taxa remain uncertain for M. carbonarius. In this study, genome-derived microsatellite markers were developed using Illumina-based next-generation sequencing. Seven loci showing strong amplification and high polymorphism were validated across 60 individuals from 6 locations in Penang, Malaysia. The number of alleles per locus ranged from 9 to 13, with expected heterozygosity (HE)…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
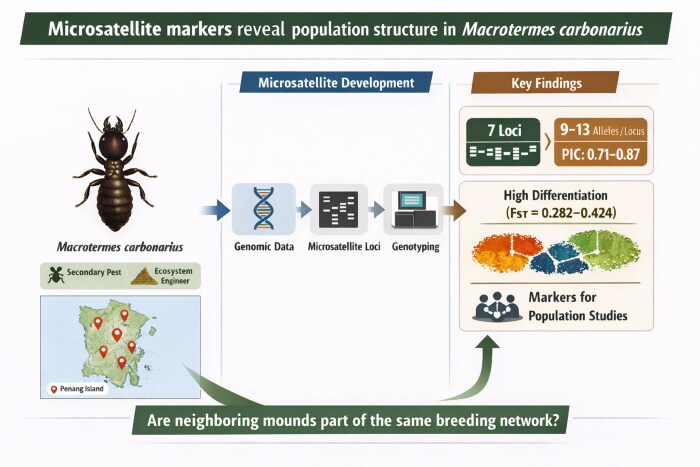 Figure 1
Figure 1| Location | Latitude | Longitude |
|---|---|---|
|
| 5°21′27″N | 100°18′13″E |
|
| 5°25′55″N | 100°17′51″E |
|
| 5°23′1″N | 100°18′39″E |
|
| 5°23′16″N | 100°16′49″E |
|
| 5°19′24″N | 100°16′7″E |
|
| 5°20′47″N | 100°16′15″E |
| Locus | Repeat motif | Primer sequences |
| Expected size (bp) | Observed allele range (bp) |
|
|---|---|---|---|---|---|---|
|
| (AG)37 |
F: GTCAACGAATCATGAGGCAG R: TGAGGGATGATGTGGTAGCAT | 56 | 131 | 110 to 139 | 12 |
|
| (AAT)8 |
F: TCACGATGTCTGTCCTCGAC R: TTTACGGACCGTGTATTCTACC | 55 | 131 | 131 to 139 | 11 |
|
| (ATC)8 |
F: CACACCGTATTCTAATGACAGA1AA R: CAACAGCCATGAATACGGAA | 53 | 109 | 109 to 129 | 11 |
|
| (AAAG)8 |
F: AAGTAGGCAGAAATGCGTCA R: ATTTGCAGGTTTATTGGGCA | 54 | 124 | 122 to 130 | 3 |
|
| (AATT)22 |
F: CCTGGCTTTGGAATAACGAA R: GGTTAGTCACTCACTCACTCACTCAT | 55 | 111 | 111 to 124 | 11 |
|
| (AAGG)16 |
F: CAGGCTCGGTAGTTCGTGAG R: TCAGTCGGCTGACTTCCTTT | 58 | 107 | 105 to 117 | 8 |
|
| (ATTC)13 |
F: AACCGATTTACATTGCCTGAA R: GGACCTGTCGTCAGACAGAA | 53 | 132 | 119 to 132 | 12 |
|
| (CCAAG)18 |
F: TCCGTTAGAATTTGGCTTGG R: GTGCTGTGGCAGTTTGATGT | 55 | 140 | 132 to 140 | 9 |
|
| (AAATT)10 |
F: TCTGTATGGATTGCATGGGA R: TCATGAAAGCACCTGTGTCAA | 55 | 117 | 117 to 142 | 13 |
| Locus | Micro-checker | Cervus | |||||
|---|---|---|---|---|---|---|---|
| Stuttering | Allele dropout | Present of null alleles | NA | HE | HO | PIC | |
|
| No | No | No | 12 | 0.888 | 0 | 0.869 |
|
| No | No | No | 11 | 0.857 | 0 | 0.833 |
|
| No | No | No | 11 | 0.858 | 0 | 0.834 |
|
| No | No | No | 11 | 0.846 | 0 | 0.821 |
|
| No | No | No | 12 | 0.872 | 0 | 0.850 |
|
| No | No | No | 9 | 0.751 | 0 | 0.707 |
|
| No | No | No | 13 | 0.882 | 0 | 0.863 |
| Locus |
|
|
|
|---|---|---|---|
|
| 1 | 0.282 | 1 |
|
| 1 | 0.3 | 1 |
|
| 1 | 0.361 | 1 |
|
| 1 | 0.291 | 1 |
|
| 1 | 0.424 | 1 |
|
| 1 | 0.383 | 1 |
|
| 1 | 0.31 | 1 |
|
| 1 | 0.336 | 1 |
- —Industry Research
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect and Arachnid Ecology and Behavior · Animal Behavior and Reproduction · Plant and animal studies
Introduction
Termites play essential roles in ecosystem functioning, particularly in tropical and subtropical regions, where they contribute significantly to the decomposition of organic matter and soil nutrient cycling (Jouquet et al. 2011). Among them, Macrotermes carbonarius, a fungus-growing termite species found across Southeast Asia, is notable for its large colony sizes and its ecological importance in forested and urban landscapes (Eggleton and Tayasu 2001, Yamada et al. 2005).
In Malaysia, M. carbonarius could be a secondary pest following Coptotermes infestations (Lee et al. 2007). When primary colonies are suppressed, M. carbonarius could occupy the same environment, prolonging damage and complicating control efforts as this species responds poorly to standard baiting programs and even fills the stations with soil rather than feed (Iqbal et al. 2017, Lee and Lee 2025). Because of this, understanding colony origin, spatial spread, and genetic relatedness between nearby mounds is key for answering two practical questions: where do replacement colonies originate, and are neighboring mounds part of the same breeding network? Such information is particularly important for eusocial termites, where colonies are typically composed of closely related individuals and dispersal of reproductives is limited. Addressing such questions require molecular markers capable of resolving fine-scale genetic structure. Markers with insufficient polymorphism may fail to distinguish between nearby but independent colonies, reducing their usefulness for both ecological and applied studies.
Microsatellite markers, also known as simple sequence repeats, are highly polymorphic DNA regions composed of short tandemly repeated nucleotide motifs. These markers are abundant, codominant, and widely distributed across eukaryotic genomes, making them powerful tools for studying genetic diversity, population structure, gene flow, and evolutionary relationships (Bruford and Wayne 1993, Buschiazzo and Gemmell 2006, Bhargava and Fuentes 2010). Although microsatellite loci have been developed for several termite species, including some within Macrotermitinae, their informativeness, amplification consistency, and allele diversity vary among species. Previous studies have demonstrated that cross-species amplification may occur in some cases; however, transferred loci often display limited allele richness or variable performance, and their suitability for resolving colony-level structure in M. carbonarius has not been evaluated (Singham et al. 2012, Husseneder et al. 2013).
The advent of next-generation sequencing (NGS) has revolutionized marker development by allowing rapid identification of microsatellite loci at the whole-genome scale (Lim et al. 2021, Tsai et al. 2024, Tamaoki et al. 2025). Given the need for markers with high PIC and reproducible amplification, a genome-derived approach was adopted to develop microsatellite loci directly from M. carbonarius.
The objective of this study is to develop and characterize microsatellite markers for M. carbonarius using Illumina-based NGS. Following marker identification, a subset of polymorphic loci was validated across individuals collected from different colonies within a local area. The markers developed will serve as valuable tools for future ecological, evolutionary, and population-level studies of M. carbonarius.
Materials and Methods
Sample Collection and DNA Extraction
Macrotermes carbonarius specimens were collected in January and August 2025. Table 1 presents the 6 sampling locations of M. carbonarius within Penang, Malaysia. All sites were vegetation-rich areas such as parks and residential gardens. The coordinates (latitude and longitude) are listed to indicate the precise collection points, which range from Georgetown in the north to Bayan Lepas in the south. These spatially distinct sites provided representative samples for assessing genetic variation among local colonies.
One individual (soldier termite) was randomly selected for whole-genome sequencing. Genomic DNA was extracted from the head of the insect using the HiYield Plus Genomic DNA Mini Kit (Blood/Tissue/Cultured Cells) (Real Biotech Corp., Taipei, Taiwan), following the manufacturer’s protocol. A total of 50 µl of DNA was obtained and quantified using a NanoDrop 2000c spectrophotometer (Thermo Fisher Scientific, Massachusetts, United States). Illumina paired-end (PE) libraries with an average insert size of 150 bp were prepared according to standard protocols (Illumina, San Diego, California, United States) and sequenced on the HiSeq platform. In addition, 60 individuals (10 from each sampling site) were selected for DNA extraction to evaluate the polymorphism of the designed microsatellite markers.
Microsatellite Marker Design
Microsatellite markers were identified from the draft genome sequencing dataset of M. carbonarius using OSTRFPD v0.01 (Mathema et al. 2019), with a minimum threshold of eight repeats for motifs ranging from mono- to hexanucleotides. Primer pairs flanking the identified microsatellite regions were designed using Primer3 (Untergasser et al. 2012) under default parameters. The primers were set to a length of 17 to 26 bp, with a melting temperature between 58 °C and 63 °C and a GC content of 20% to 80%. Additional criteria included an expected amplicon size of 100 to 150 bp and the absence of predicted secondary structures.
Microsatellite Marker Validation
Twenty microsatellite markers were selected from the designed set (Supplementary Table S1, available in the Figshare repository with DOI: 10.6084/m9.figshare.29654594) for polymorphism analysis based on the number of repeat units and the type of repeat motif. These markers were used to amplify DNA from 60 individual M. carbonarius specimens. PCR amplification was carried out using a Thermal Cycler (TaKaRa, Japan). Each 25 µl PCR reaction contained 12.5 µl of Master Mix with green buffer (NX, Nuclix Biosolution, Malaysia), 0.5 µl of 5 mM forward and reverse primers, 5 µl of DNA template, and sterile cold distilled water to make up the final volume. PCR conditions included an initial denaturation step, followed by 40 cycles of denaturation at 94 °C for 30 s, annealing at locus-specific temperatures (listed in Table 2) for 30 s, and extension at 75 °C for 45 s, with a final extension step at 75 °C for 5 min.
Fragment analysis was performed using the Fragment Analyzer Automated CE System (Agilent Technologies, California), and allele scoring was carried out using ProSize 3.0 software (Agilent Technologies, California). Of the 20 markers tested, 7 were selected for further validation based on motif type and amplification quality. These included 1 dinucleotide repeat (MCDi3), 2 trinucleotide repeats (MCTri1 and MCTri2), 2 tetranucleotide repeats (MCTetra4 and MCTetra8), and 2 pentanucleotide repeats (MCPenta3 and MCPenta5).
Micro-Checker v2.2.0.3 (Van Oosterhout et al. 2004) was used to analyze the fragment analysis results, specifically examining the observed and expected null allele frequencies to detect errors such as amplification failure, stuttering, or large allele dropout during PCR. Allele frequency analysis, including observed and expected heterozygosity, the number of alleles, and the PIC, was conducted using Cervus v3.0.7 (Kalinowski et al. 2007). Population differentiation (FST) was estimated following Weir and Cockerham (1984) using GENEPOP on the web (Rousset 2008).
Results and Discussion
Microsatellites were screened from 15.3 million raw sequence reads. The identified microsatellites were distributed as follows: 8.72% mononucleotide, 42.45% dinucleotide, 35.41% trinucleotide, 12.29% tetranucleotide, 1.11% pentanucleotide, and 0.029% hexanucleotide repeats. From these, 82,243 primer pairs flanking the screened microsatellites were designed, as detailed in Supplementary Table S1.
From the large set of designed primers, 20 primer pairs were selected for experimental screening. Most of these primers successfully genotyped 60 M. carbonarius individuals from 6 populations and produced varying numbers of alleles. Primers yielding 3 or more alleles are presented in Table 2.
Then, 7 markers that showed higher numbers of alleles were selected for further validation. These markers demonstrated high genetic diversity, producing 9 to 13 alleles per locus among samples from 6 locations (Table 3). All loci showed clear and consistent amplification without evidence of stuttering, allele dropout, or null alleles, as confirmed by Micro-Checker. The PIC values for all loci ranged from 0.707 to 0.869, indicating that all 7 loci are highly informative (Botstein et al. 1980). These results demonstrate that the newly developed markers are robust and suitable for population genetic studies of this species.
Although the observed heterozygosity (HO) values across loci were low, the expected heterozygosity (HE) values were relatively high, ranging from 0.751 to 0.888. The discrepancy between HO and HE may be attributed to the reproductive biology of termites, which are known for their eusocial structure and high levels of relatedness within colonies. Colonies typically originate from a single reproductive pair and may later include neotenic reproductives derived from the same lineage. Such reproductive systems lead to inbreeding, reduced heterozygosity, and deviations from the Hardy-Weinberg equilibrium (Shellman-Reeve 1997, Vargo 2003).
Population differentiation, as indicated by F-statistics (Table 4), revealed varying levels of genetic structuring among colonies. FST values ranged from 0.282 (MCDi3) to 0.424 (MCTetra8), with an average of 0.336, suggesting high differentiation among populations. The highest genetic differentiation observed in MCTetra8 indicates substantial divergence between colonies, likely due to limited dispersal of reproductive alates and the spatial isolation of colonies. Since M. carbonarius reproductives have short flight ranges and rely on suitable environmental conditions for successful colony establishment, restricted dispersal could promote local genetic differentiation (Thorne et al. 1999).
The consistently high inbreeding coefficient (FIS = 1) across all loci further supports the presence of inbreeding within colonies. This pattern aligns with the termite’s hierarchical breeding system, in which individuals within a colony are typically descendants of a single reproductive pair or related neotenics. The resulting genetic homogeneity and reduced heterozygosity are characteristic of termite populations with restricted outcrossing opportunities (Calleri et al. 2006, Bankhead‐Dronnet et al. 2015).
Supplementary Material
ieag008_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bankhead‐Dronnet S , Perdereau E, Kutnik M, et al. 2015. Spatial structuring of the population genetics of a European subterranean termite species. Ecol. Evol. 5:3090–3102. 10.1002/ece 3.156626357538 PMC 4559052 · doi ↗ · pubmed ↗
- 2Bhargava A , Fuentes FF. 2010. Mutational dynamics of microsatellites. Mol. Biotechnol. 44:250–266.20012711 10.1007/s 12033-009-9230-4 · doi ↗ · pubmed ↗
- 3Botstein D , White RL, Skolnick M, et al. 1980. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 32:314–331.6247908 PMC 1686077 · pubmed ↗
- 4Bruford MW , Wayne RK. 1993. Microsatellites and their application to population genetic studies. Curr. Opin. Genet. Dev. 3:939–943. 10.1016/0959-437X(93)90017-J 8118220 · doi ↗ · pubmed ↗
- 5Buschiazzo E , Gemmell NJ. 2006. The rise, fall and renaissance of microsatellites in eukaryotic genomes. Bioessays. 28:1040–1050. 10.1002/bies.2047016998838 · doi ↗ · pubmed ↗
- 6Calleri DV , Mc Grail Reid E, Rosengaus RB, et al. 2006. Inbreeding and disease resistance in a social insect: effects of heterozygosity on immunocompetence in the termite Zootermopsis angusticollis. Proc. Biol. Sci. 273:2633–2640. 10.1098/rspb.2006.362217002949 PMC 1635464 · doi ↗ · pubmed ↗
- 7Eggleton P , Tayasu I. 2001. Feeding groups, lifetypes and the global ecology of termites. Ecol. Res 16:941–960.
- 8Iqbal N , Wijedasa LS, Evans TA. 2017. Bait station preferences in two Macrotermes species. J. Pest Sci. 90:217–225.